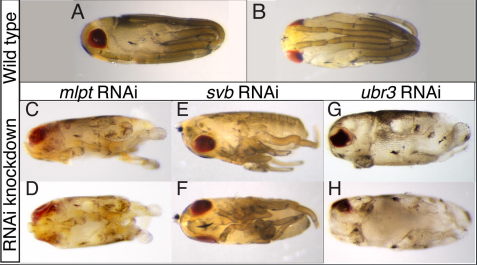
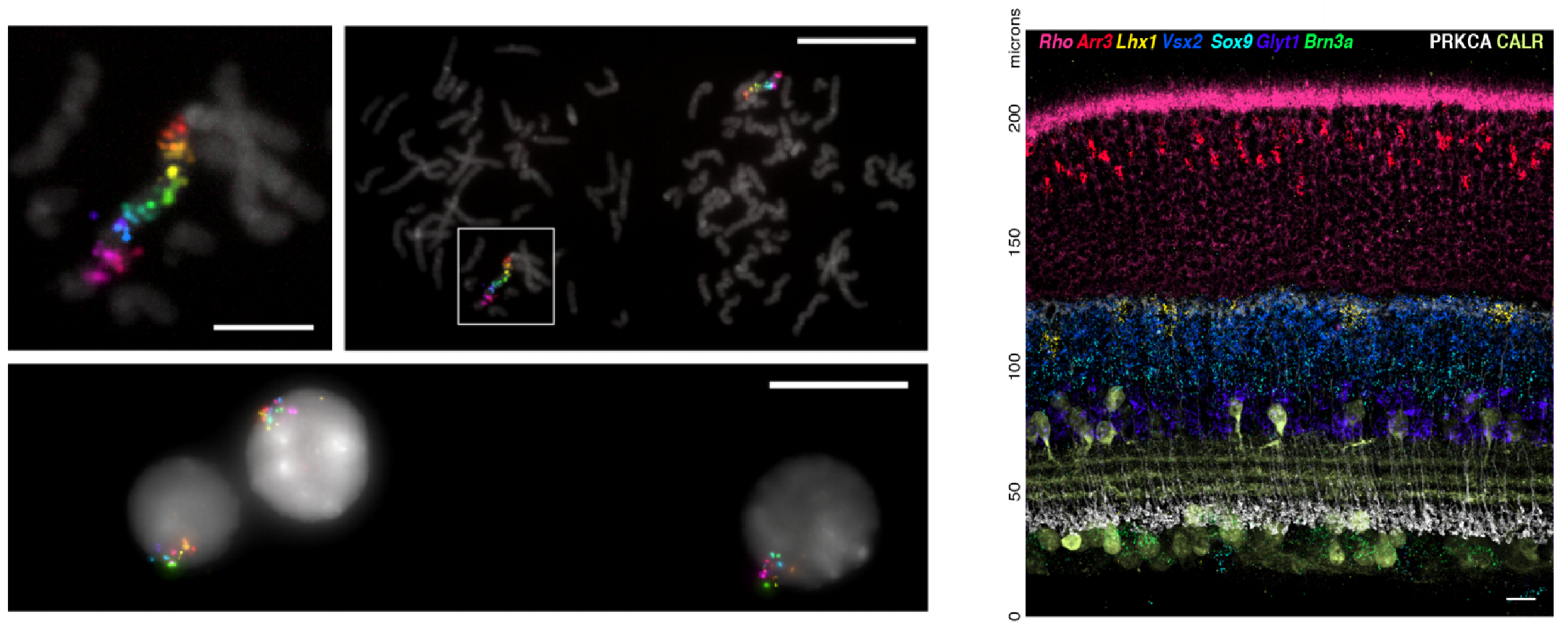
Four Year (1+3) PhD Programme in Stem Cell Biology & Medicine
Posted by stemcellsjobs, on 16 October 2018
Closing Date: 15 March 2021
Department/Location: Wellcome – MRC Cambridge Stem Cell Institute
Closing date: 4 January 2019
Interviews will be held: 30 January – 1 February 2019
Studentships starting October 2019
https://www.stemcells.cam.ac.uk/join-us/study
Stem Cell Biology
Stem cells are defined by the dual capacity to self-renew and to differentiate. These properties sustain homeostatic cell turnover in adult tissues and enable repair and regeneration throughout the lifetime of the organism. In contrast, pluripotent stem cells are generated in the laboratory from early embryos or by molecular reprogramming. They have the capacity to make any somatic cell type, including tissue stem cells.
Stem cell biology aims to identify and characterise which cells are true stem cells, and to elucidate the physiological, cellular and molecular mechanisms that govern self-renewal, fate specification and differentiation. This research should provide new foundations for biomedical discovery, biotechnological and biopharmaceutical exploitation, and clinical applications in regenerative medicine.
Cambridge Stem Cell Community
The University of Cambridge is exceptional in the depth and diversity of its research in Stem Cell Biology, and has a dynamic and interactive research community that is ranked amongst the foremost in the world. By bringing together members of both the Schools of Biology and Medicine, this four year PhD programme will enable you to take advantage of the strength and breadth of stem cell research available in Cambridge. Choose from over 50 participating host laboratories using a range of experimental approaches and organisms.
Programme Outline
During the first year students will:
- Perform laboratory rotations in three different participating groups working on both basic and translational stem cell biology.
- Study fundamental aspects of Stem Cell Biology through a series of teaching modules led by leaders in the field.
- Learn a variety of techniques, such as advanced imaging, flow cytometry, and management of complex data sets.
Students are expected to choose a laboratory for their thesis research by June 2020, and will then write a research proposal to be assessed for the MRes Degree in Stem Cell Biology. This assessment will also be used to determine whether students continue on to a 3-year PhD.
Physical Biology of Stem Cells
Incorporated into the ‘Stem Cell Biology’ Programme, opportunities are available specifically for candidates with a Physical, Computational or Mathematical Sciences background, wanting to apply their training to aspects of Stem Cell Biology *.
Great inroads have been made towards understanding how stem cells generate tissue and sustain cell turnover, most of which have been made by studying the biochemistry of stem cells. Less is known of their function across scales – from molecules to tissue – or interaction with their physical environment. We aim to identify the importance of physical, chemical, mathematical and engineering considerations in stem cell functionality. This could include mathematical modelling, engineering controlled environments to control stem cell function, single molecule approaches to study molecular interactions, systems biology, or investigating stem cell’s response to forces in its environment.
Eligibility
We welcome applications from those who hold (or expect to receive) a relevant first degree at the highest level (minimum of a UK II.i Honours Degree or equivalent). You must have a passion for scientific research.
Stem Cell Biology and Medicine Programme (funding by Wellcome)
We welcome applications from EU and non-EU candidates. The Wellcome Trust provide full funding at the ‘Home/EU’ rate. Funding does not include overseas fees, so non-EU applicants will need to find alternative funding sources to cover these.
‘Physical Biology of Stem Cells’ Programme (funding by the MRC)
We welcome applications from UK/EU candidates, with a Physical Sciences, Mathematical or Computational Sciences background. *The Medical Research Council provide full funding for UK applicants only. Applicants from EU countries other than the UK, are generally eligible for a fees-only award. Please check your eligibility status at https://www.mrc.ac.uk/skills-careers/studentships/studentship-guidance/student-eligibility-requirements/ before applying.
Application Process
Visit https://www.stemcells.cam.ac.uk/join-us/study for full details, including how to apply.
Enquiries are welcome via email to sci-phd@stemcells.cam.ac.uk.
 (No Ratings Yet)
(No Ratings Yet)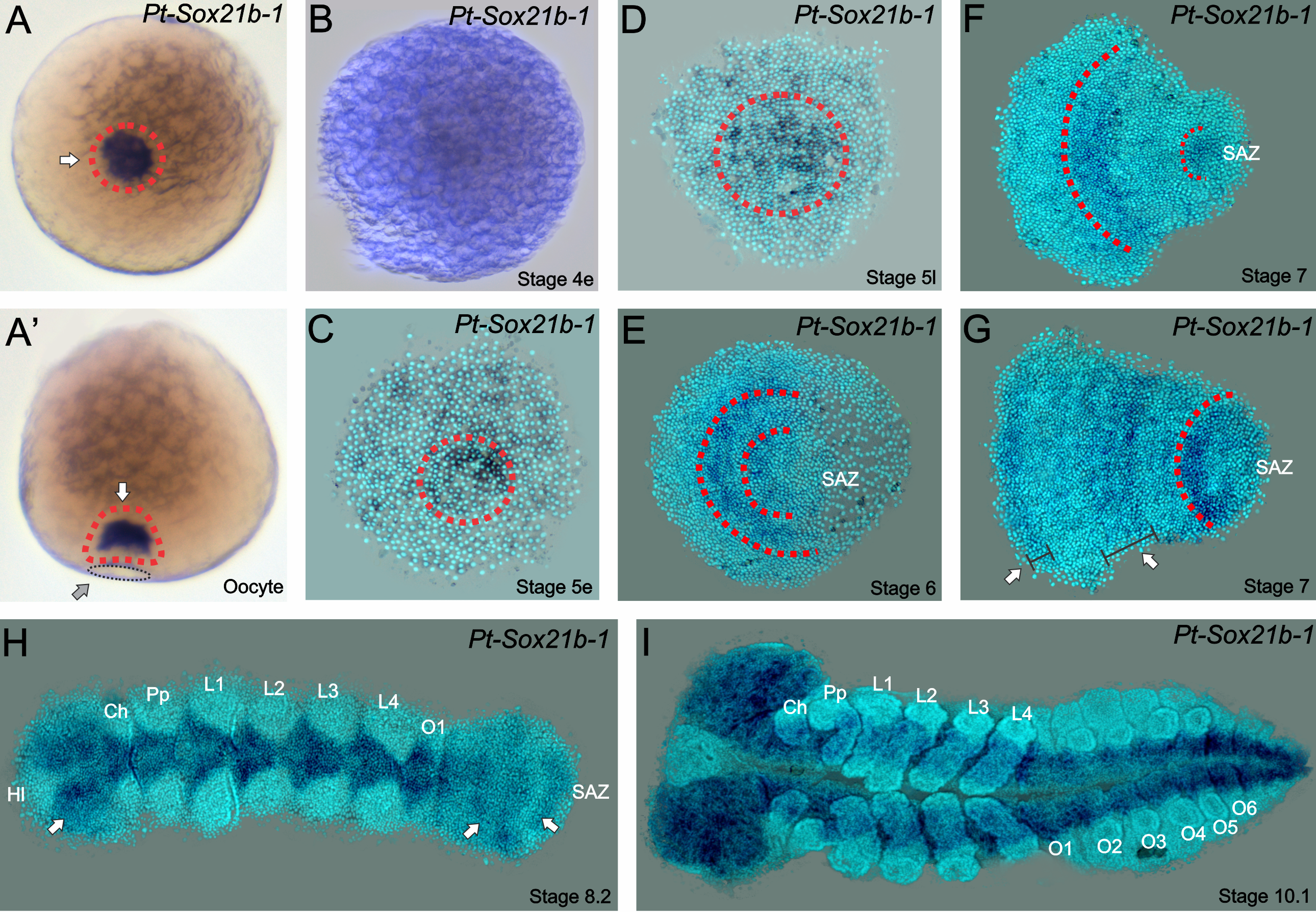
 (3 votes)
(3 votes)