Two Postdoctoral Fellow positions are available in Gabsang Lee laboratory at Johns Hopkins University. These positions are NOT for stem cell expert, rather for who has OUTSTANDING expertise on either biochemistry, skeletal muscle biology, optogenetics OR single-cell studies. Previous experience on human pluripotent stem cells is not necessarily required, and we value more on the ‘non-stem cell’ experience. Research project will be developed based on the applicants’ talent and interest, in a conjunction with our research topics.
The Lee lab has been establishing novel methodologies to specify human induced pluripotent cells (hiPSCs), and to direct convert somatic cells, into multiple lineages, including induced neural crest (Kim et al., Cell Stem Cell, 2014), peripheral neurons (Oh et al., in revision), Schwann cells (Mukherjee-Clavin et al., in revision) and skeletal muscle cells (Choi et al., in revision) using multiple genetic reporter systems. We continue to study human developmental and degenerative disorders (pain disorders, demyelination disorders, and muscular dystrophies) to unravel the underlying cellular/molecular mechanism toward realistic therapeutic approaches.
Areas of research topics of these positions are below.
– Neural cell fate determinations
– Pain disorders
– Demyelination disorders
– Insulin resistance
– Muscular dystrophies
– Satellite cell biology
Applicants can send (by Feb 10th, 2016) a CV and the names of three references to the address listed below:
Melina Schuh received her diploma degree in biochemistry from the University of Bayreuth, Germany, where she completed her Diploma thesis with Stefan Heidmann and Christian Lehner. She went on to do her PhD with Jan Ellenberg at the European Molecular Biology Laboratory in Heidelberg, Germany. In 2009, after a bridging postdoc with Jan, Melina started her own group at the MRC Laboratory of Molecular Biology in Cambridge, UK. Since January 2016, she is a Director at the Max Planck Institute for Biophysical Chemistry in Göttingen, Germany, and will establish a new department focussing on meiosis. She is an EMBO Young Investigator and a recipient of the 2014 Lister Institute Research Prize, the 2014 Biochemical Society Early Career Award and the 2015 John Kendrew Young Scientist Award. Her lab is studying meiosis in mammalian oocytes, including human oocytes.
What motivated you to become a scientist?
To some degree I became a scientist by chance. I had many different interests in school, and I was struggling to decide whether I wanted to become a journalist, an architect or a scientist. My friends and family encouraged me to study biochemistry because they thought that it suited me most. But I don’t think I knew at that stage what it would be like to work as a scientist. Studying biochemistry was a very good choice, though, and I never had any regrets. I loved the way science was taught at university.
In your PhD you developed methods for studying meiosis in mouse oocytes. What problems did you have to overcome, how did you devise your system?
There were very few high-resolution studies of mouse oocytes when I started my PhD in Jan Ellenberg’s lab at EMBL in Heidelberg. Jan’s lab had worked with starfish oocytes but had no experience with mammalian oocytes. I wanted to develop a system that would allow us to image meiosis in oocytes at high resolution by using a confocal microscope. This was not trivial because oocytes normally develop inside the body and they are very sensitive cells. I had to find out how to culture the oocytes outside the body on a confocal microscope, how much light they could tolerate, and how to label and follow structures – like the chromosomes – over the entire course of meiosis, which takes more than 12 h. Much of it was learning by doing, but I also got very valuable advice from various people around me.
You continue to work on meiosis in mouse oocytes. What are the particular questions that your group is currently trying to answer?
Our main aim is to understand how defects at the interface between chromosomes and the cytoskeleton lead to aneuploid eggs and pregnancy loss in mammals. We have also recently started to study meiosis and chromosome segregation directly in live human oocytes, which has not been possible before. This is a new direction of research in my lab that I am very excited about, as it should allow us to investigate why human eggs are so likely to be aneuploid.
What parts of the cytoskeleton are you specifically looking at?
We have evidence that, in human oocytes, the way that the spindle assembles is different from how it assembles in mitotic cells. Spindle assembly takes more than half a day in human oocytes. In mitosis, it takes just 30 minutes. So the question is: why are human oocytes so slow in assembling a spindle? Oocytes from many species assemble the spindle without centrosomes. Instead, they use acentriolar microtubule organising centres (aMTOCs), which functionally replace the centrosomes. We could not find these aMTOCs in human oocytes. We discovered that human oocytes assemble the spindle by a mechanism that is chromosome-dependent and mediated by the small GTPase Ran. The spindles need to be extensively reorganized while the chromosomes become aligned in the spindle centre. This could be one of the reasons why spindle assembly takes so long. These are, at least to our knowledge, the first studies of chromosome segregation in live human oocytes.
Have we been looking at an incomplete system when studying meiosis in mouse oocytes?
Mouse oocytes still resemble human oocytes in many aspects. We will need to study human oocytes in much more detail to evaluate how closely they resemble mouse oocytes.
How did your collaborations influence your research? Do you have any advice on collaborating?
It is helpful to collaborate with scientists that you enjoy interacting with and where the communication is working well. Which is crucial to a successful collaboration.
Everyone makes mistakes. How do you deal with them in the lab?
It is unavoidable that you make mistakes at some point in your career. And you should, of course, try to learn from your mistakes. But it is equally important that you don’t get hung up on these mistakes. You need to accept that you cannot be perfect. Try to always remind yourself of all the things that are working, the things you are doing well and to simply enjoy the science.
What advice on how to establish a successful academic career would you give?
I would recommend that you try to do something new and exciting, even if it is difficult. Be creative, ambitious and fearless in your approaches. Do not think that something cannot be done just because nobody has done it yet. It is good if you try to develop something new, because this helps you to stand out from the crowd, become known for what you are doing, advance your career and get tenure.
What do you think about the feasibility of being both a good parent and a good scientist?
I think this is a challenge for many parents. Pregnancy, maternity or paternity leave, the many sleepless nights that come with a baby, and the time involved in looking after children will all have an impact on your productivity. There is not much that can be done about this. It does help to have excellent day care, ideally subsidised by institutes. It is also important that line managers and institutions in general are understanding that there will be a transient period of time when progress is going to be a bit slower. Having this support will help you to stay focussed on your work and to get through this busy time with confidence.
You left your home country, Germany, to start your own research group here in Cambridge. Do you think of returning to Germany one day?
I have been offered a great position at the Max Planck Institute for Biophysical Chemistry in Göttingen. I very much enjoy working at the LMB, but my husband is still working in Germany while I am in the UK with two kids. Moving back to Germany will allow us to live together as a family while doing research at a fantastic institute – it couldn’t be better.
This obituary was written by Ross Cagan andEyal Gottlieb, and first appeared in Disease Models & Mechanisms.
With the untimely death of Marcos Vidal, we have lost a good friend and a creative, brilliant colleague who made important contributions to the field of cancer biology through fruit fly research. Marcos began his research into Drosophila at Ross Cagan’s laboratory in 2003 first at Washington University in St. Louis and later at Mount Sinai Hospital in New York. In 2009 Marcos was appointed as Research Group Leader at the Beatson Institute for Cancer Research in Glasgow.
I (Ross) had just finished giving a talk in the Renal Division at Washington University School of Medicine when a young scientist approached me. He was a biochemist but liked the idea of using fruit flies to study diseases. Marcos Vidal was an impressive young man and I was excited to have him join the laboratory. A few days later a woman approached with the same request. Julia Cordero assured me that her relationship with Marcos would not be a problem, that they were used to working together and that they never fought. I sat them on opposite sides of the laboratory.
Marcos and Julia were a brilliant young couple who instantly made the laboratory smart and fun. Marcos took up cancer research, the second person to ever work on this disease in the laboratory. He loved everything about conducting science and he was ambitious. He worked crazy hours. With a laconic Argentinian accent, he kept up a running science conversation that lasted until now. Julia worked on separate topics and they lived balanced lives. Marcos loved board sports from snowboarding to kite- and wind-surfing and was superb at them. Marcos and Julia made friends easily. The Facebook postings and the emails we’ve received and the conversations we’ve had over the past few days are a reminder just how much they were loved. We all rooted for their success because they were really good people.
Marcos pushed the fly as a discovery platform further than almost anybody else. He was enthusiastic to exploit the power of drug screening. He had a feel for the organism and, perhaps most importantly, he had a feel for what constituted an important biological question. In 2005 he validated Vandetanib as a candidate therapeutic for medullary thyroid carcinoma. The drug was approved in 2011 for clinical use. He explored the role of Src in oncogenesis, providing evidence and mechanisms that position metastasis as an immediate early event, promoted at least in part by local cell interactions within the epithelium. Later on, as an independent Group Leader in Glasgow, Marcos focused on the cancer promoting role of the tumor microenvironment and the immune system. Using the fruit fly as a cancer model, Marcos demonstrated that the genetic composition of a tumor is a key determinant as to whether Tumor Necrosis Factor (TNF) acts as a tumor suppressing or tumor promoting factor. He further characterized the crosstalk between epithelial tumors and the innate immune response, demonstrating that the TNF-Toll non cell-autonomous signaling cascade dictates tumor cell fate. Marcos used the fruit fly cancer model creatively, and he did it with a style unmistakably his own.
As a couple, Marcos and Julia were a true science power: young and smart and handsome. In Glasgow, Marcos started out as a Junior Group Leader at the Beatson Institute, becoming my (Eyal) valued neighbor and peer. Julia helped him set up his laboratory, and later started her own. They lived the lives of young professionals, with an international twist. They worried about keeping two leadership research positions, worried about their two young children and whether Argentinians could adapt to the Scottish weather. But they soon became central to the Beatson Institute, got involved with the local community and established successful professional and personal lives in Scotland. Beyond his work in the lab, Marcos dedicated his energy and free time to his family. He spent long, happy hours passing his boarding and drawing skills to his son Lautaro, and riding his bike with his daughter Mara.
Marcos fell ill 18 months ago and those extremely difficult times were a testimony to the central role his family and friends played in his life. Marcos worked as hard as he could to make a recovery. He received tremendous support from his colleagues and friends and, most of all, from Julia. Despite struggling with his illness, Marcos sailed through his Tenure Review with flying colors and in July 2015 he became a Senior Research Group Leader at the Beatson Institute and a Professor at the University of Glasgow.
Marcos Vidal died on January 2, 2016. Julia, Lautaro and Mara lost a warm-hearted, dedicated and loving husband and father; the science community has lost a brilliant young scientist and a good friend.
Recommended by Jane Langdale (University of Oxford)
If you read about plant development in textbooks you will be told that the shoot meristem, which gives rise to all the stems, leaves and flowers, is formed of distinct cell layers. In dicotyledonous flowering plants there are three layers: the outermost layer (L1) will give rise to the epidermis; the L2 layer will produce most of the subepidermal tissues of leaves and flowers; and the innermost L3 layer forms the stem and vasculature tissues. That these layers exist and that they make different contributions to the shoot does not seem so surprising now, but there was a time when even the number of the then called ‘germ’ layers was a topic of debate. There had been much interest in meristems since the mid 19th century, and scientists like Hanstein had already suggested that meristems might have 3 germ layers in some plants. There was the need to carefully determine whether that was the case in all higher plants.
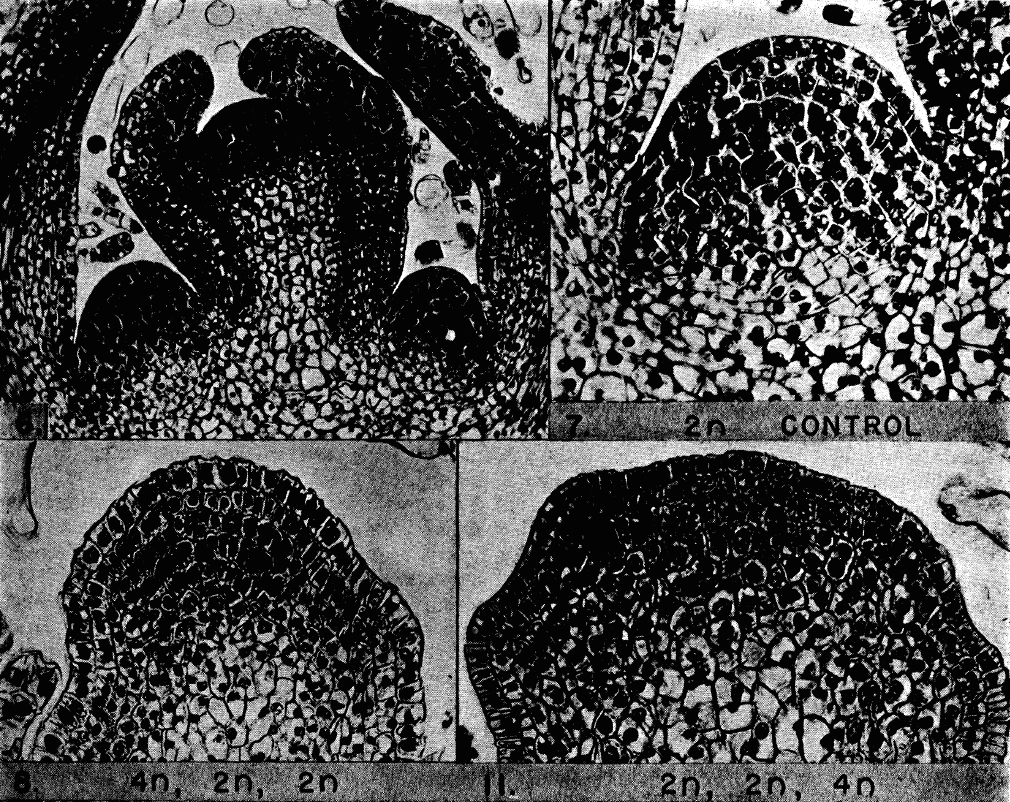
In the late 1930s, Amos Avery and his colleagues were on the case. But how do you investigate cell lineages when you are 60 years too early to use ‘basic’ labelling techniques like GFP-tagged proteins? What was needed was a way to genetically modify some of the cells of the plant so that you could identify their descendants and hence follow their fate, but it didn’t necessarily have to be a precise change. Realising this, Avery and colleagues took a blunt approach. They soaked seeds of Datura plants in colchicine to induce chromosome duplications. As colchicine inhibits spindle formation, chromosome duplication occurs without cell division, generating cells of different ploidy within a single seed – a chimera. It is important to note that the shoot meristem is fully formed in the embryo and hence these treated seeds contained shoot meristems in which some cells were polyploid and others were not.
Reproduced with permission of the American Journal of Botany
The seeds were grown in fields over the summers of 1938 and 1939, and the plants were regularly pruned to encourage the formation and outgrowth of lateral shoot meristems (which were ultimately derived from subsets of cells in the primary embryonic shoot meristem). Such an experiment is of course a game of chance, and as a result Avery and his colleagues obtained a huge range of chimera types, which they then used to study different aspects of plant development. In this particular paper, they examined the shoot meristem. Using cell size as a broad readout of ploidy, they examined how the cells of the shoot meristem were affected by the procedure. They noticed that you could find plants in which all the cells of the first layer had a different ploidy to the rest of the meristem. Sometimes they found plants in which the change in ploidy happened only in the cells of the second layer, and sometimes only in the cells of the third layer. And very occasionally you could find a plant where all three layers had different ploidy, e.g. the first layer was 2n, the second layer was 4n and the third layer 8n. Since ‘the response of any one layer to colchicine is independent of the response of the other two’, the authors concluded that the shoot meristem of Datura must have three independent germ layers. They also observed another phenomenon that is now textbook fact, namely that the cells in the 1st and 2nd layer (L1 and L2) mainly undergo anticlinal cell divisions (where the division plane is perpendicular to the cell layer) while the cells of the third layer can undergo divisions in any orientation.
This paper essentially reported the results of a single experiment, and did not go on to give further details on the subsequent development of the germ layers and on which organs originated from which layer. This is perhaps something we would expect in a modern paper, but science was different in those days so the authors saved those experiments for a subsequent paper. However, one should not underestimate the importance of this work. As Jane Langdale (who recommended this paper) put it, this was the ‘first demonstration that shoot apical meristems of flowering plants operate in the context of functionally distinct layers. A concept that is now so embedded that students rarely wonder how it was first shown.’
Further thoughts from the field
This study is a prime example of an early clonal analysis in plants. It provided direct information about the cell division patterns in shoot apical meristems and laid the basis for the tracing of cell lineages and the origin of tissues. Most importantly, it established the existence of three germ layers in plants.
Kay Schneitz, TU Munich (Germany)
Though molecular genetics has shown the importance of communication among layers in shoot meristem organization, we are far away from understanding its complexity and how this communication orchestrates layer-specific gene expression and the identities of individual cell layers. In the data rich science that we practice today, this paper is a must read for every developmental biologist, as it conveys the power of a single and carefully thought about experiment.
Venugopala Gonehal, UC Riverside (USA)
The American Journal of Botany has kindly provided free access to this paper until the end of June 2016!
—————————————–
by Cat Vicente
This post is part of a series on forgotten classics of developmental biology. You can read the introduction to the series here and read other posts in this series here.
You just started your research career, or maybe you just moved fields. The first thing on your to-do list is to catch up with the literature. What has been the latest progress? What are the open questions in the field? Ideally, you would go all the way back to the beginnings of the field and acquire a broad perspective of how research has changed and how the earlier works have impacted on current research. But time is short. The literature is just too vast, and the bench is calling, urging you to get started on your experiments. So you read a couple of recent comprehensive reviews, a bunch of papers from the last few years and a handful of the most significant earlier studies and move on to start your research project.
There is nothing wrong with this approach- there really is a vast number of papers out there, and time is limited. Unfortunately, this means that many important papers are overlooked. Certain ‘facts’ are taken for granted, and how they were obtained is not questioned. That chance encounter with an old result that could change the course of your research never takes place.
Aware of these issues, we have decided to launch a new series of posts called ‘Forgotten Classics of Developmental Biology’. We asked prominent researchers from all areas of developmental biology to suggest their favourite hidden gems and to explain why we should all revisit them. In this series we will provide you with a summary of each paper, alongside comments from the person who recommended it as well as from other researchers in the field. For those papers still not freely available online, we also aim to ask the publishers if they will provide free access to each paper for a limited period of time after the post is published. We hope this will give everyone a chance to benefit from re-reading these forgotten classics!
The list of papers that we plan to feature in this series is still growing, so if there is a ‘Forgotten Classic’ that you think we should cover, please get in touch. We would love to hear your suggestions.
The signalling systems that conduct the orchestra of embryonic development are fantastically complex and dynamic. We owe much of our knowledge of in vivo signalling dynamics to advances in microscopy and protein tagging with fluorescent reporters that have allowed visualization of signalling proteins. Looking forward, however, it is clear that simply analyzing the localization patterns of proteins is only the first step to fully understand signalling processes. Of course, a protein’s presence at a particular time or place in the embryo does not tell us if it is actively signalling or lying dormant until its activity is required. Therefore, generic methods to distinguish actively signalling proteins from passive proteins will be tremendously useful advances for researchers interested in developmental processes.
A few years ago, Darren Gilmour at the European Molecular Biology Laboratory (EMBL) in Heidelberg, Germany, realized that since many signalling events lead to the degradation, stabilization, or relocalization of participating proteins, methods for detecting changes in protein turnover could provide powerful proxies for in vivo signalling activity. Gilmour and his graduate student Erika Donà therefore took advantage of a dual-color genetically encoded fluorescent reporter called a “tandem timer”, newly developed by Michael Knop in the next-door lab at the EMBL. Tandem timers report on the age of protein populations through the relative fluorescence of adjacent slow- and fast-maturing fluorophores (Khmelinskii et al. Nat. Biotechnol 2012). In collaboration with Knop and his postdoc Anton Khmelinskii, Gilmour and Donà tagged the chemokine receptor Cxcr4b with a tandem timer with a view to observing Cxcl12a chemokine signalling activity across a migrating tissue, the zebrafish posterior lateral line primordium. They wagered that this strategy was likely to inform on chemokine signalling activity since Cxcr4b receptors undergo rapid internalization and degradation upon ligand-mediated activation. Higher Cxcl12a levels would increase Cxcr4b internalization rate and this should be reflected by a decrease in the receptor’s population age (increased protein turnover). Gilmour and Donà also understood the potential long-term implications of their attempt; a successful proof-of-principle study might lead to the adoption of the tandem timer as a generic tool for the detection of signalling activities.
At the time I was a postdoctoral researcher in Wolfgang Huber’s group at EMBL, and having collaborated with Knop and Khmelinskii on the original tandem timer paper, I was happy to get involved in this first project using tandem timers in embryos. Using image processing and analysis tools from R and Bioconductor software, I measured tandem timer signalling readouts from 3D images of migrating tissues, and used a mathematical model of timer behaviour to interpret the results. The model describes the maturation kinetics of the two fluorophores on the timer, as well as production and degradation rates of the tagged protein. What surprised and delighted me throughout my collaboration with Gilmour and Donà, was the extent to which quantitative analysis and modelling was able to feed back on subsequent experimental design, and positively influence later results. Ultimately our tandem timer observations, along with other data, allowed us to demonstrate that a self-generated chemokine gradient guides the migration of the zebrafish posterior lateral line primordium (Donà et al. Nature 2013).
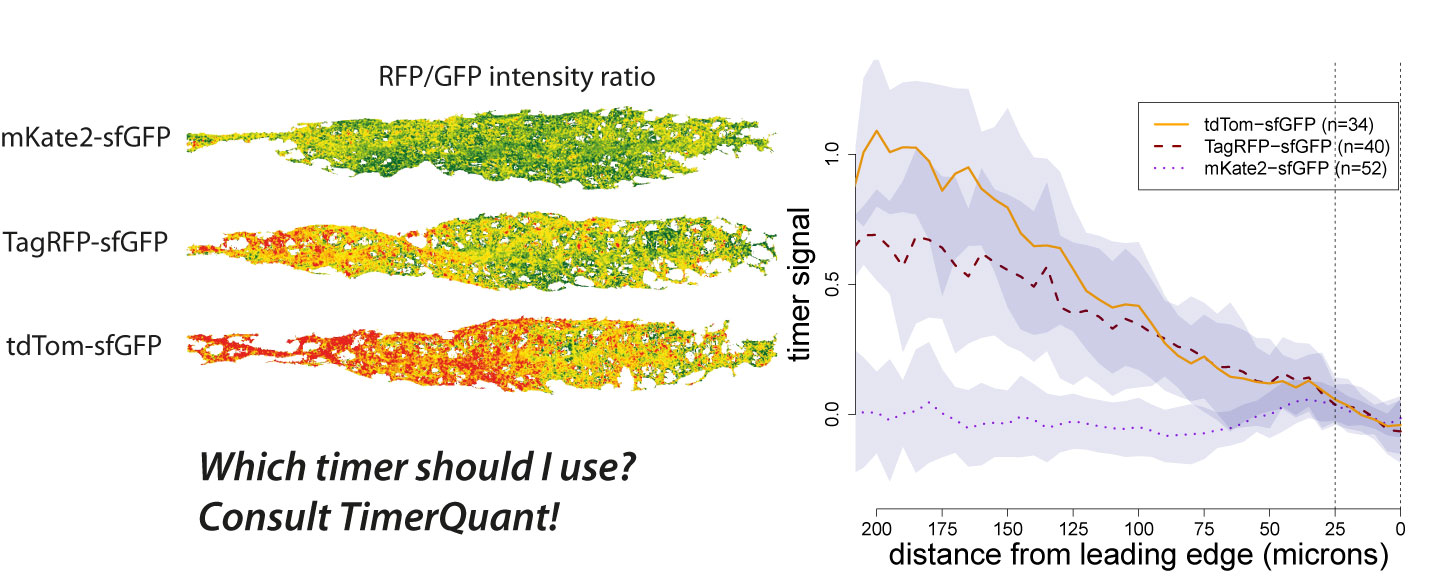
Tandem timers can be constructed with virtually any combination of spectrally separated fluorescent proteins with different maturation rates. Therefore, the first question an experimentalist typically asks when using tandem timers is: which fluorophores should I choose? We have been contacted by many groups asking exactly this question. As our ability to design timers and interpret timer data has benefited greatly from the use of modelling, we decided to write a paper outlining how our modelling tools could be used by other researchers interested in tandem timer research. To make these tools easily accessible we developed an interactive web application, TimerQuant, and made all our code available through the open-source software platform Bioconductor. For our new paper, which is now published in the journal Development (Barry et al. Development 2016), we systematically investigated the effect of relevant experimental parameters on timer signal, a measure of how good a particular timer is at detecting differences between two protein half-lives. To validate the predictions of our model, we reinvestigated the Cxcr4b signalling gradient identified in our original study using three tandem timers that had the same fast-maturing fluorophore (sfGFP), but different slow-maturing fluorophores (mKate2, mCherry, TagRFP). We found that timer signal decreased as the maturation times of the slow- and fast-maturing fluorphores became more similar. Readouts became noisier as protein abundance decreased. Timer signal increased with the maturation time of the slow-maturing flurophore, albeit at the expense of noisier readouts. While these conclusions might already be expected from a more qualitative reasoning, the quantitative model also led to some unexpected and intriguing findings. For example, Förster resonance energy transfer (FRET) between the fast- and slow-maturing fluorophores actually increased timer signal, a prediction that was borne out in our experiments, and which may be an important consideration for future timer designs.
In the initial model protein production (expression) and degradation rates were kept constant. Given that in developmental contexts these are likely to change over time, we decided to explore the effect of dynamics on timer ratio profiles. We modelled constant increases and decreases of protein expression and degradation rates as well as sudden bursts of expression and degradation, and looked at model solutions over time. The simulation results showed timer ratio profiles over time producing clear, characteristic responses to expression and degradation dynamics. We were surprised by the extent to which expression and degradation responses were distinguishable from one another. These additions to the model further help in the interpretation of timer ratio results in non-steady state conditions, which is often the situation when studying dynamic developmental processes. Moreover, as degradation rate could be experimentally fixed, for example by using non-degradable (or slowly degradable) versions of tandem timers, these model predictions open up the exciting possibility of using tandem timers to observe gene expression dynamics with time-lapse microscopy.
Here on the Node you can sign up for several mailing lists, but since they were implemented at different stages each sign up page was in a different place. To makes things simpler and easier, we have now created a single page where you can sign up to our three mailing lists:
Node Weekly emails– a list of all new posts published on the Node in the last week.
Occasional writing ideas– for those who want to post on the Node but need some inspiration.
Occasional news from The Company of Biologists– updates from our sister journals, workshops and grants.
If you haven’t sign up to these lists yet, or only signed up to some, you can do it now in our new Sign up page. We also added a new Sign up box on the right hand side column of the Node, so that this new page is easy to find. New users will also be given the option to sign up to these email lists upon registration.
You will also notice that that our daily update emails have now become weekly. In our recent survey several of you commented that our update emails were sent too frequently, so we hope that a weekly email will work better. Our first weekly email will arrive to your inbox tomorrow, also featuring a new look. As always, we welcome your feedback on these changes. Please feel free to leave a comment here or drop us an email!
TheBSDB Newletter 2015 is out! As you might have noticed, there was no BSDB summer newsletter this year. The reason for this is that the BSDB committee has taken the sensible decision to reduce them to one edition a year. Newsletters clearly do no longer play the same role as they used to. In times of the internet, they are no longer needed to update members on new developments of society matters or in the area of Developmental Biology. This is now done far better through the BSDB website and through The Node (also see page 9 of the Newletter). However, we feel that providing an overview of the year still is a service we owe to BSDB members, and which might provide an informative document also for future generations.
This year, the BSDB has implemented the new Cheryll Tickle Prize and we proudly announce Abigail Tucker as the well-deserved inaugural Awardee of 2016 (page 14). This prize is for outstanding women in their mid-career and our chair, Ottoline Leyser, explains the underlying rationale in her welcome note (page 2). As usual, this issue contains the reports by our secretary Kim Dale (page 3), meetings officer Joshua Brickman (page 4), treasurer Chris Thompson (page 7), communications officer (page 9) and, for the first time, a joined note by our graduate student rep Alexandra Ashcroft and postdoc rep Michelle Ware who explain to us their plans to improve services for young BSDB members (page 10). We congratulate all BSDB awardees of 2015 (page 6), introduce our new committee members (page 11) and present Ana Ribeiro’s report of the Autumn Meeting 2015 (page 5). Claudio Stern makes an important case for promoting developmental biologists for prizes and politically important memberships (to which the BSDB committee is responding already; page 16), and we explain our recent decision to become a full member of the Royal Society of Biology (page 23). Finally, we present four reports of the 2015 Gurdon/The Company of Biologists Summer Studentship awardees which clearly demonstrate the success of this scheme (page 17).
I hope you enjoy this issue and, in the name of the BSDB committee, would like to wish you a happy and successful new year 2016.
Andreas Prokop (Communications officer)
P.S. If you are interested in BSDB newletters of the last 10 years, they are archived on our website.
A running joke amongst avian developmental biologists is that the chicken (Gallus gallus) is the tastiest of the model organisms. A typical response from some of my mouse, frog or fish friends, would be that that is where the advantages end – the lack of the ability to do genetics in birds present limitations in the types of molecular analyses that could be done. Despite being so tasty, the chick is not the most cost effective lab animal to keep: It is big, and requires a large amount of space for housing, and it has a slow generation time of almost a year. These points make the development of genetic techniques in chick prohibitive to most labs. About 5 years ago, we decided to see what could be done with the zebra finch. Zebra finches have been widely used in studying behaviour and neural circuitry. They have the advantage of being small and have a generation time of about 3-4 months, the same as that for mouse. With support from the RIKEN CDB Director’s fund (Masatoshi Takeichi at that time) and from the Animal Facility (particularly Shinichi Aizawa, its then head), establishing our finch colony in the RIKEN CDB was straightforward. We bought the founders from pet shops in the Kansai region of Japan and housed them in our newly appointed finch room at the CDB. At this point we realised that many of the published protocols for finch husbandry were not optimal for egg production and so we decided to invest time optimizing these protocols.
Zebra finch pair
Siu-Shan Mak (Suzanne) the lead post-doc who really drove the study, and Anna Wrabel-Suzuki, the animal technician in charge of looking after the birds, spent the first year studying the behaviour of the birds, and checking the quality of the eggs, tweaking the diet, lighting, and cage conditions so that we could house them in the best conditions for optimal fertile egg production. It was during these checks of egg quality that Suzanne noticed that the finch embryo was at a younger stage than the chick at laying. A small note here; birds undergo some development in utero, before the egg is laid. Chicks and quails lay their eggs when the embryos are just about to gastrulate (Eyal-Giladi and Kochav 1976). We found that finches lay fairly early, with the embryo at a late blastula stage. We thought that this difference would allow us some additional insights into early avian development.
The zebra finch family
Around this time, the groups of Jennifer Nichols and Austin Smith published a series of papers describing the stage in mouse at which embryonic stem cells (ES cells) that showed naïve pluripotency could be derived (Boroviak et al. 2014). In this state, naïve ES cells can give rise to all cell types found in the embryo. It contrasts with primed pluripotency, where the cells (also called epiblast-derived stem cells or EpiSCs) have a more restricted potential (Nichols and Smith 2009). Another key difference of these cells is their sensitivity to the inhibition of an important signalling pathway intermediate MAP kinase: naïve ES cells were resistant to its inhibition whereas EpiSCs were not (Nichols and Smith 2012). The group showed that mice blastocysts at around E4 were able to generate naïve ES cells. At this stage the epiblast has been specified (as shown by molecular markers) but is yet to epithelialise. We found that the finch blastoderm at laying is similar: The epiblast, and indeed the hypoblast, are both specified but there is little morphological segregation, reminiscent of the pre-epithelialised epiblast. When we took cells from newly laid finch embryos, and cultured in the presence of a MAP kinase inhibitor, they retained the expression of pluripotent markers, and markers of naïve ES cells. Cells taken from later staged finches or from newly laid chicken embryos did not express pluripotent markers when MAP kinase was inhibited, behaving more like primed pluripotent cells. These results were presented in our recent paper in eLife (Mak et al. 2015).
We think that at the very least, the finch embryo could provide some comparative insights into the restriction of pluripotency in amniotes, and possibly would enable the generation of testable hypotheses when thinking about the evolution of pluripotency in mammals. Our hope that by combining new genome engineering tools with long-term finch ES cell cultivation and subsequent chimera generation, this work sets the stage for a tractable, genetic avian model system. These are some of the ideas we will be thinking about in our new labs in Kumamoto (Guojun) and in Bangalore (Raj).
 (No Ratings Yet)
(No Ratings Yet)

 (1 votes)
(1 votes) Marcos and Julia were a brilliant young couple who instantly made the laboratory smart and fun. Marcos took up cancer research, the second person to ever work on this disease in the laboratory. He loved everything about conducting science and he was ambitious. He worked crazy hours. With a laconic Argentinian accent, he kept up a running science conversation that lasted until now. Julia worked on separate topics and they lived balanced lives. Marcos loved board sports from snowboarding to kite- and wind-surfing and was superb at them. Marcos and Julia made friends easily. The Facebook postings and the emails we’ve received and the conversations we’ve had over the past few days are a reminder just how much they were loved. We all rooted for their success because they were really good people.
Marcos and Julia were a brilliant young couple who instantly made the laboratory smart and fun. Marcos took up cancer research, the second person to ever work on this disease in the laboratory. He loved everything about conducting science and he was ambitious. He worked crazy hours. With a laconic Argentinian accent, he kept up a running science conversation that lasted until now. Julia worked on separate topics and they lived balanced lives. Marcos loved board sports from snowboarding to kite- and wind-surfing and was superb at them. Marcos and Julia made friends easily. The Facebook postings and the emails we’ve received and the conversations we’ve had over the past few days are a reminder just how much they were loved. We all rooted for their success because they were really good people. (22 votes)
(22 votes)