And the winner of the latest roundof images from the Woods Hole embryology course is… the skate embryo!
Here are the full results:
– SEM of butterfly scale: 93 votes
– Drosophila embryos: 37 votes
– Skate embryo: 241 votes
– Swallowtail wing: 79 votes
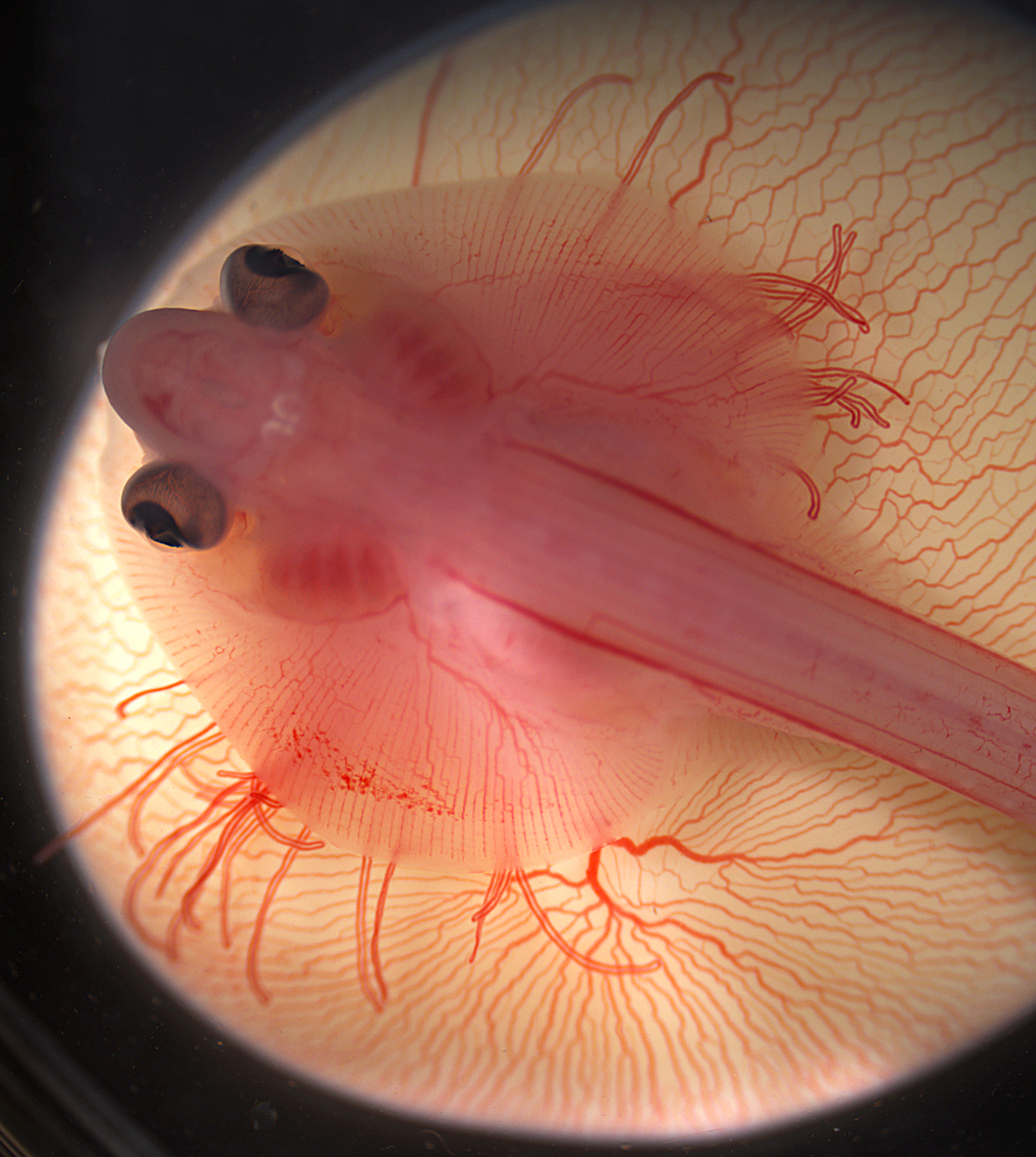
Many congratulations to Mary Colasanto (University of Utah, USA) and Emily Mis (Yale University, USA) who took this image at the 2013 course. It shows a living embryo of the little skate (Leucoraja erinacea), sitting atop its yolk at approximately ten weeks of development. Imaged on a Zeiss Discovery.V20. This image will feature in the cover of a coming issue of Development.
The other great images in this round were taken by Lara Linden (butterfly scale), Wiebke Wessels (Drosophila embryos), and Emily Mis and Misty Riddle (swallowtail wing).
Our jobs page was again very busy this month, so make sure to check it out! Here are some of the other highlights:
Research:
– Manuel wrote about his Development paper, where he examines how the eyes of the cavefish P. andruzii develop, and what that tells us about the evolution of eye loss in different cavefish species.
The Node is almost 5 years old, and it is now time to revise its design and functionalities. To help us with this, during the last month we asked you to fill in a survey and give us your feedback. In total 169 people completed the survey, and many of you left comments and suggestions. Thank you all for taking the time to help us make the Node better!
Many congratulations as well to Paolo Panza (PhD student at the Max Planck Institute for Developmental Biology in Tübingen, Germany), who is the lucky winner of our prize draw! Your bag of goodies from the Node and Development is on its way!
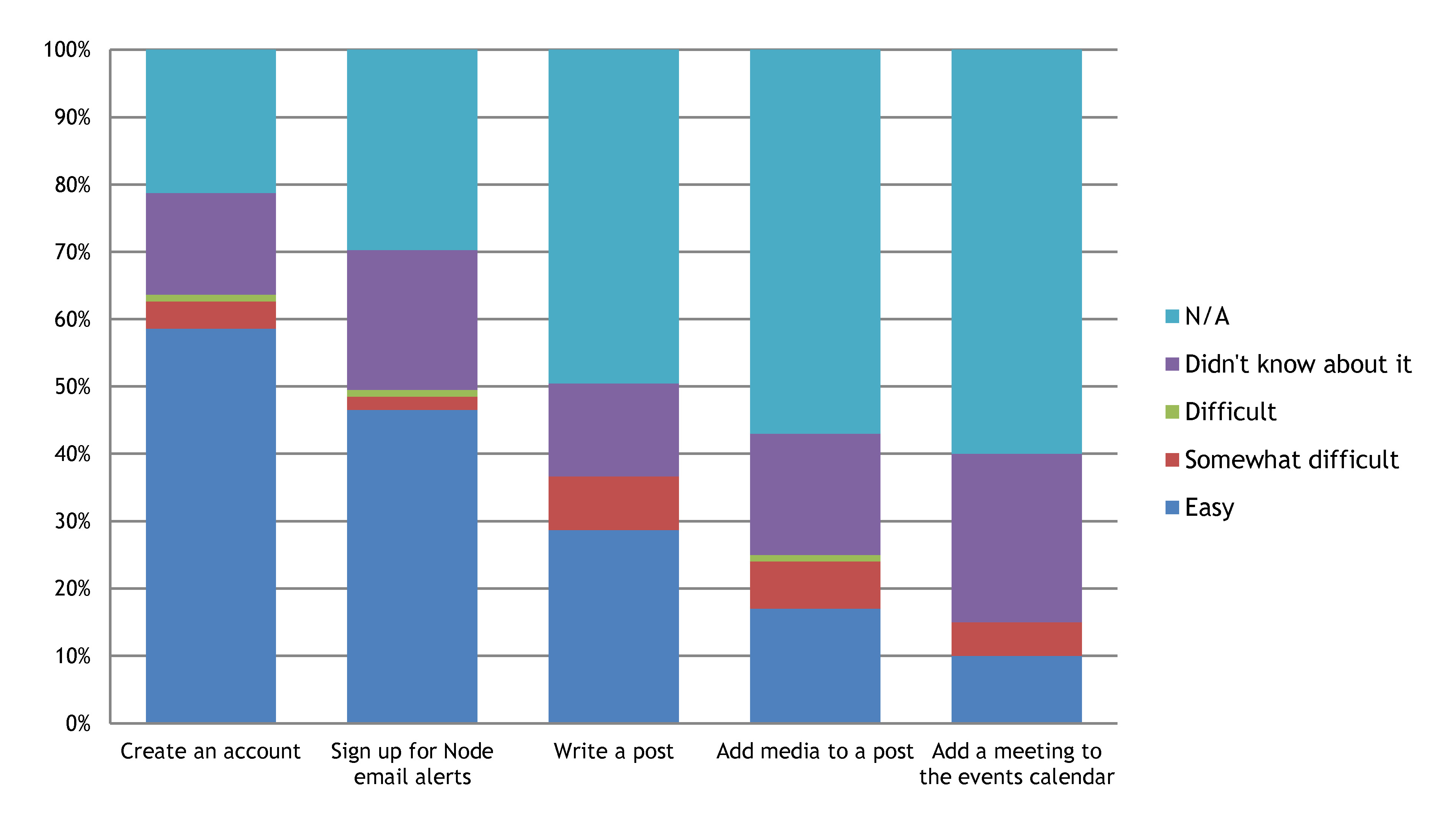
So what were the main conclusions of the survey?
1- Most of you first found out about the Node via Development or by word of mouth
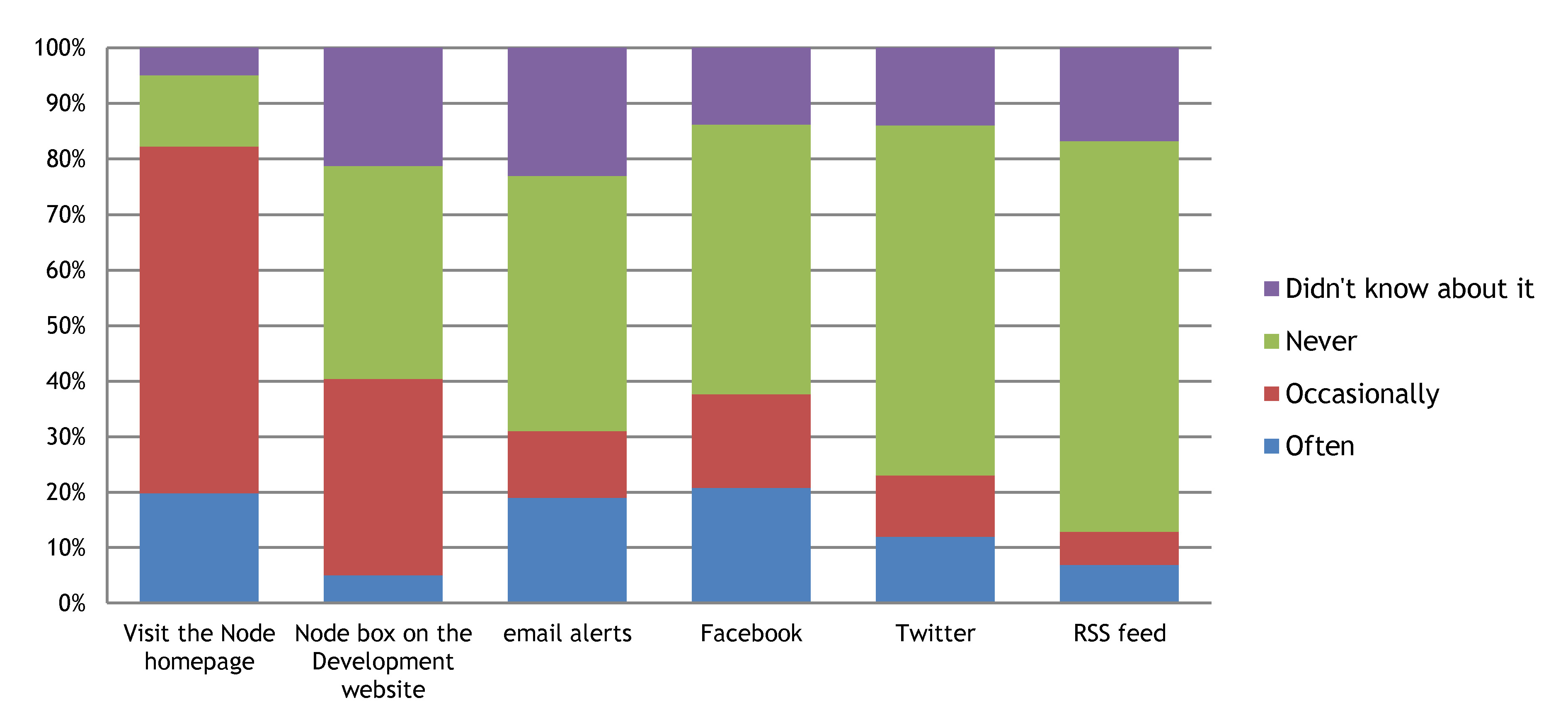
2- Most of you keep up with new Node content by visiting the Node homepage
The Node box on the Development website and facebook are also popular. Interestingly, very few of you thought the Node needed to be present in additional social media sites (only 2 people suggested LinkedIn, and there was only 1 mention of Google+).
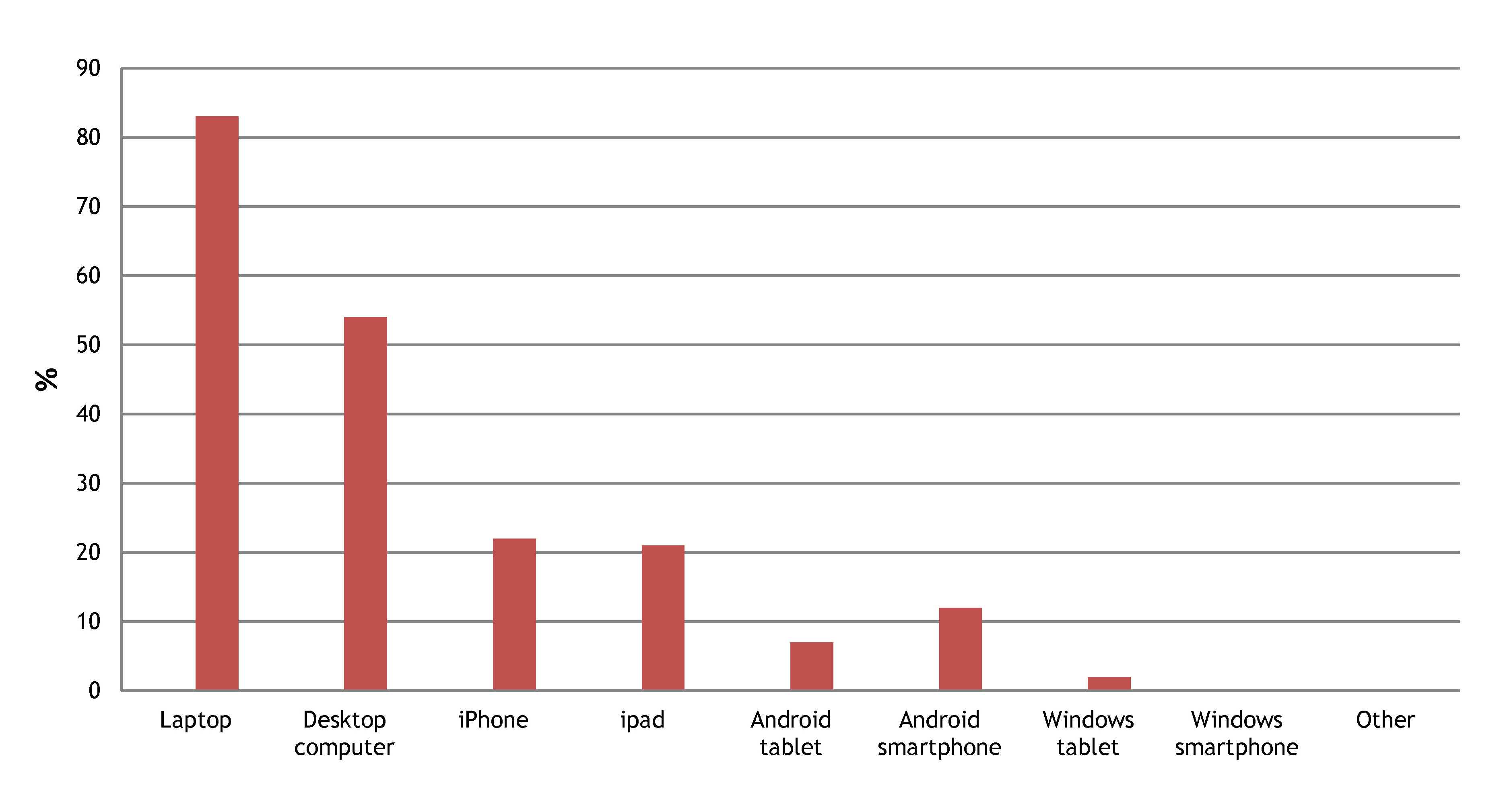
3- Laptops and desktop computers are the devices most often used to access the Node
This question was important so that we know how much effort to invest on the mobile versions of the site. However, if you access the Node mostly via your mobile or tablet please rest assured that we will continue to have a mobile version of the Node!
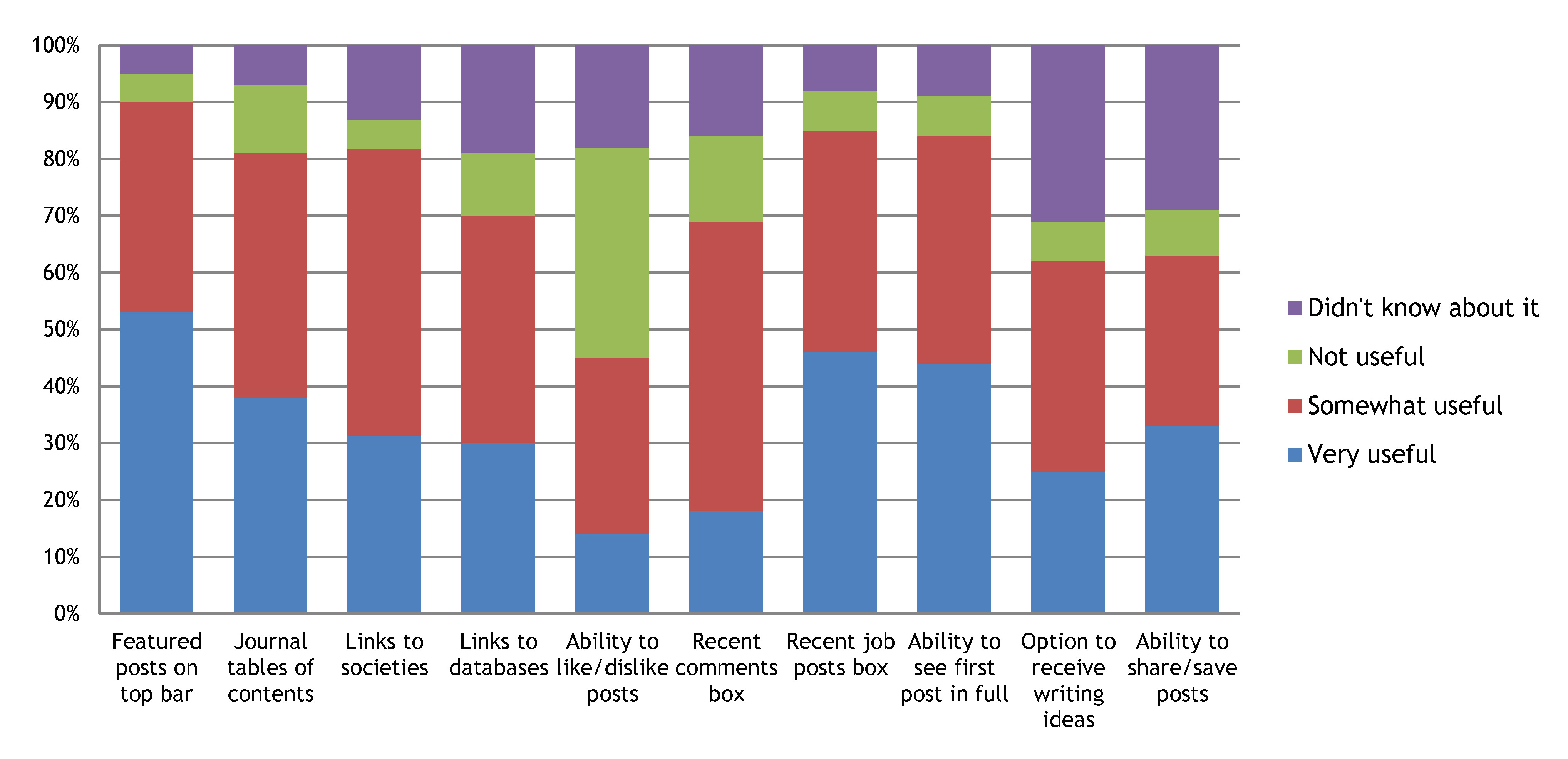
4- Most of the features that we listed are considered useful
The most popular feature listed were the highlighted posts on the top bar. Several of you also used this opportunity to point out that many functions on the site are not working properly, such as the like/dislike buttons, or the search box. We have made a record of all the problems you spotted and we are already in the process of fixing some of them!
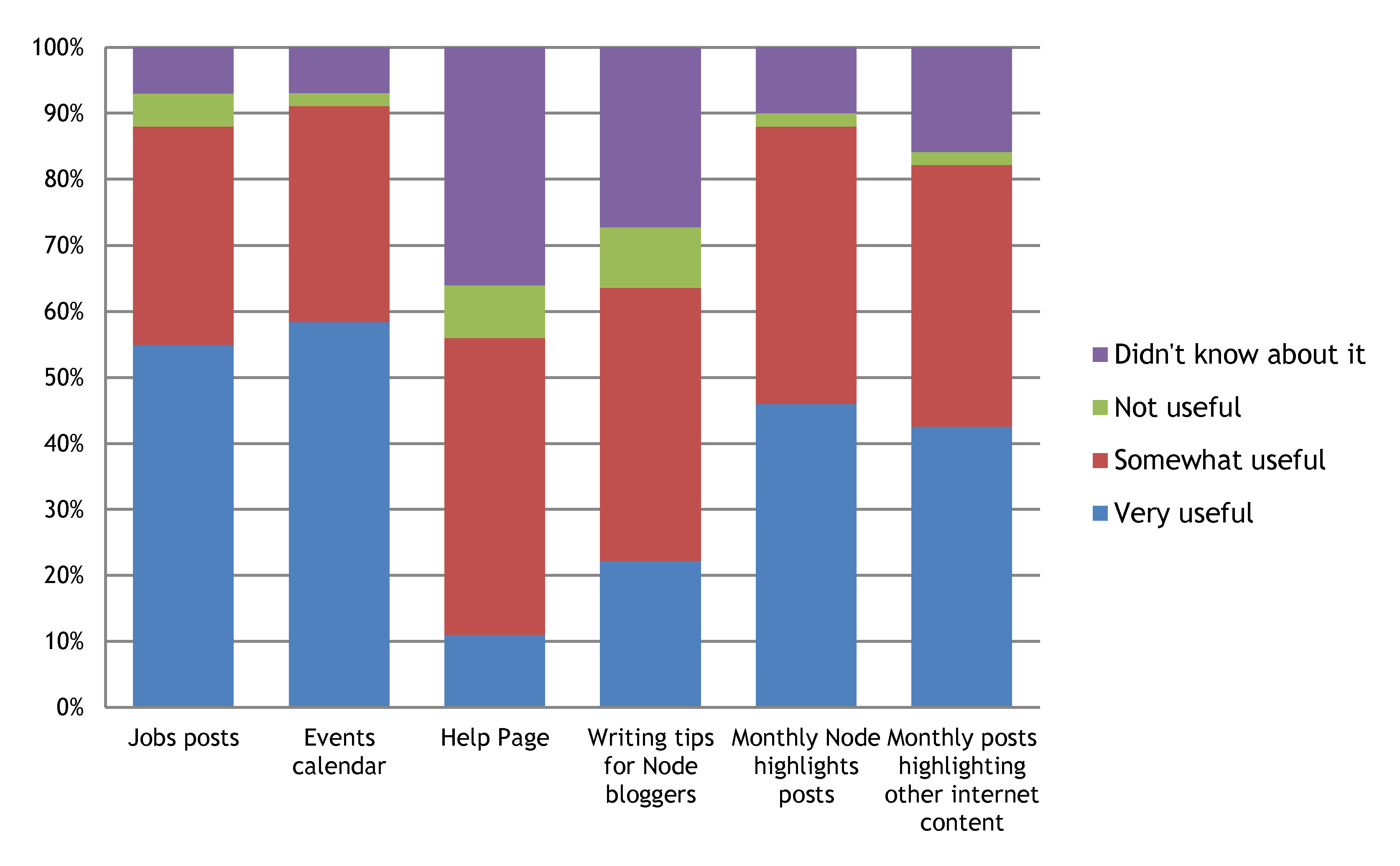
5- Most of the content that we listed is considered useful, particularly the job posts and events calendar
Many of you didn’t know about the help page and the writing tips for Node bloggers though.
6- Most of the actions listed are easy to perform
But many of the people who completed the survey haven’t tried them yet.
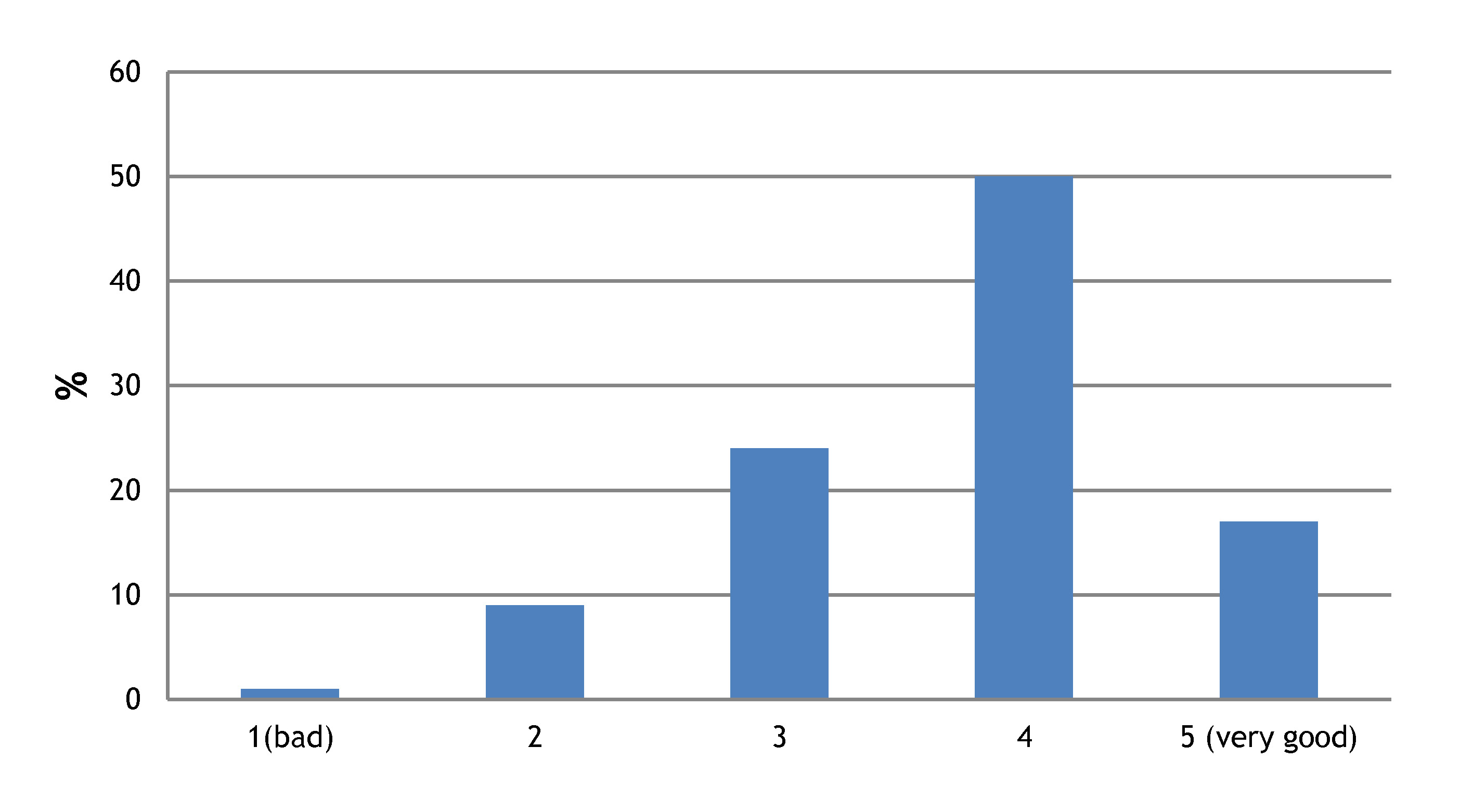
7 – The majority of you quite like the visual design of the Node
However, many people pointed out that the current look is cluttered and old fashioned. So although you don’t want the look of the Node to change completely, a sleeker, more modern look would be welcome!
8- Most of you find the Node to be both valuable and enjoyable
Several of you left very positive comments, encouraging us to keep going and to continue providing a service to the community. Thank you for your encouragement!
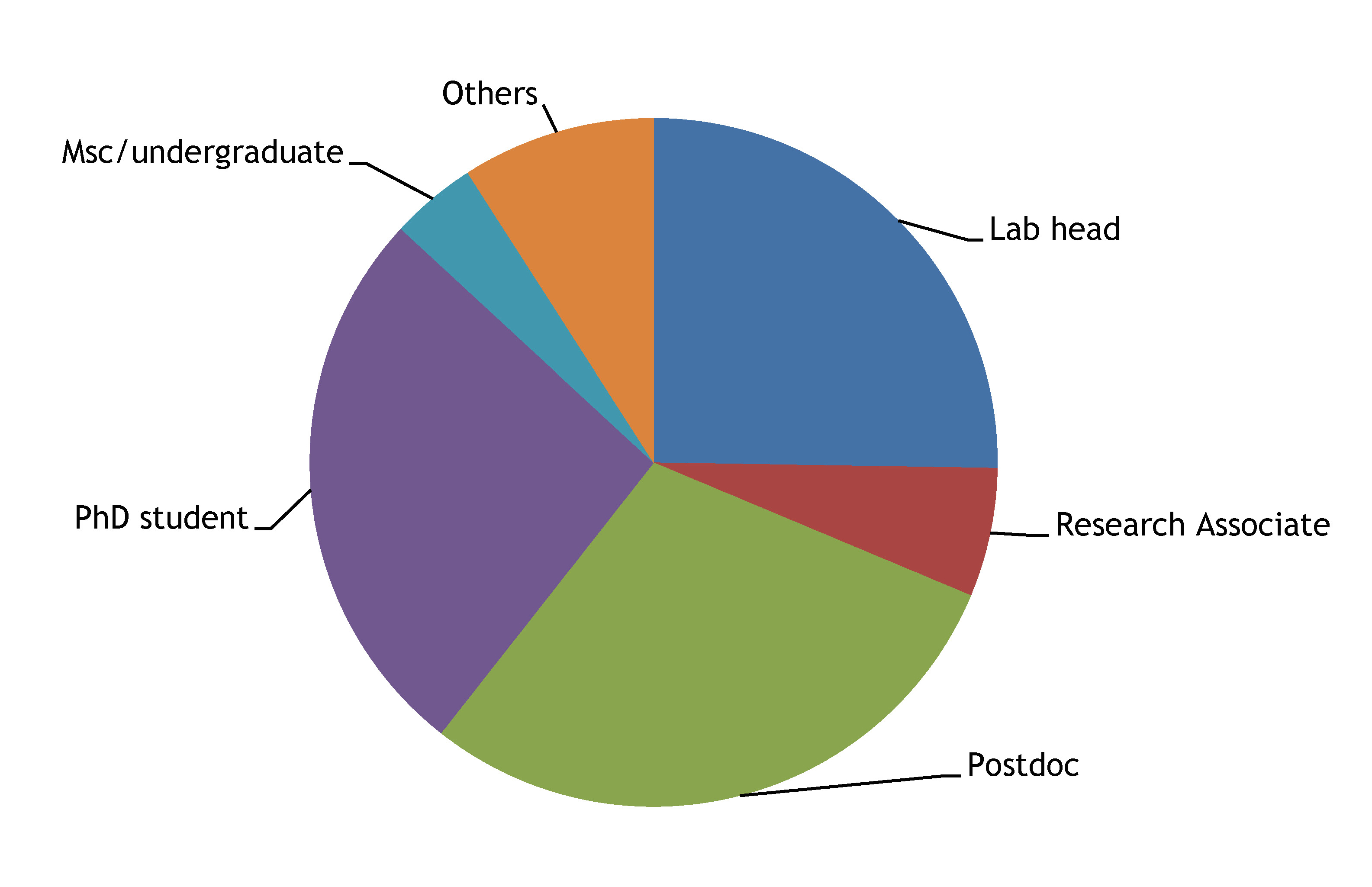
9- The Node is read by an equal proportion of PhD students, postdocs and group leaders (at least among survey responders).
Other suggestions:
Many of you used the comment boxes to leave more specific feedback and make suggestions. Some of the most common ones were:
– Have more discussion posts that generate debate
– We should find a better way to organise the information of the Node, such as creating collections or categories for different model organisms or fields
– There were also specific suggestions for new content, such as protocols and technical tips, funding, historical anecdotes on major discoveries in the field, short biographies of eminent biologists, career advice, and even a Node podcast!
We will consider all your suggestions, big or small, and they are invaluable in helping us make the Node a better community site. Many of these changes can’t only come from us though. As you know, anyone in the community can write for the Node. You only need to create an account and you can post without needing to ask us for permission. So you can help us improve the content of the Node by contributing! Write about what you find interesting, start a discussion or leave a comment. The Node is here for you and together we can make it better and more relevant!
Finally, please keep giving us your feedback. You can get in touch us via this contact form and we are always keen to hear your thoughts and suggestions!
Understanding fundamental roles of the cytoskeleton downstream of Wnt/PCP signalling in neurodegeneration
A fully funded studentship open to home/EU students is available. We are seeking a highly motivated and collaborative individual to join the dual PhD studentship of the Universities of Liverpool (UK) and Szeged (Hungary). The successful candidate will work under the supervision of Dr Natalia Sanchez-Soriano (Institute of Translational Medicine, University of Liverpool) and Dr József Mihály (Biological Research Centre, Hungarian Academy of Sciences). You will work the first 2 years of your project at the University of Liverpool and the following 1.5 years at the Biological Research Centre, Szeged in Hungary. Candidates should have a master’s degree or equivalent.
You will investigate the links between the Wnt/PCP pathway, cytoskeleton remodelling and neurodegenerative diseases such as Alzheimer’s disease, using the model organism Drosophila. You will gain experience in Drosophila neurobiology, molecular/cell biology and imaging. See more details about our work. Note that there are two further positions available for a postdoctoral researcher and technician. Details can be found here.
Candidates should email Natalia Sanchez- Soriano (Natalia1[at]liverpool.ac.uk) or Dr József Mihály (mihaly.jozsef[at]brc.mta.hu) with an expression of interest, a CV and suggestions for 2 referees.
A new position is now available in the National Xenopus Resource (NXR) at the Marine Biological Laboratory (MBL). This is an NXR Manager position that involves daily operational and scientific oversight of the MBL’s P40 NIH funded National Xenopus Resource.
The NXR Manager will supervise a staff of 3-5 research assistants and technicians, coordinate custom transgenic and mutant orders and purchases, help organize workshops, and troubleshoot the frog housing systems. In addition, the individual will work on scientific projects that serve to improve the NXR, including genome editing, sperm cryopreservation and health screening.
The NXR Manager will work with researchers from the Xenopus community who visit the NXR for short term research projects through our research hotel service or as MBL Research Awardees.
The successful candidate must have a PhD in Biological Sciences and is essential that they have previous training and/or experience with the Xenopus model. Individual must be highly motivated, independent, enthusiastic and able to work well with others. Several years postdoctoral experience is highly beneficial.
The Node is off to its first conference of the year- the Annual Drosophila Research Conference, i.e. the fly meeting! We will be tweeting from Chicago using the hashtag #DROS2015, and you can also expect a meeting report here on the Node.
If you are attending the meeting, why not say hello to our community manager Cat? She will be around during the meeting and she is always keen to chat with Node readers. Feel free to drop her an email via here if you would like to meet her!
Hello! I am Annie, a research assistant in the group of Molecular and Developmental Complexity, under the supervision of Dr. Luis Alfredo Cruz-Ramírez. Our group is part of the National Laboratory of Genomics for Biodiversity (LANGEBIO) at Irapuato, Mexico. We are a relatively new group as we only started a year ago. Our research focuses on the molecular networks underlying cell reprogramming events and their influence in regeneration and developmental complexity. We are very interested in deciphering the first signals that trigger cell reprogramming for blastema formation during Ambystoma limb regeneration.
The Axolotl is a salamander species belonging to the family Ambystomatidae and part of the Ambystoma genus. This genus includes 32 species in North America, distributed from southeastern Alaska to the southern boundary of Mexico. Half of the global diversity of the genus Ambystoma (16 species) is endemic to Mexico. The 16 species are distributed along the so-called Transmexican Volcanic Belt (TVB). This region is the most populated in Mexico, and most of the endemic species of Ambystoma found there are threatened by changes to their habitat.
Wild type and albino mutant Axolotls (Ambystoma mexicanum)
The Axolotl in mexican culture
Within the Ambystoma genus Ambystoma mexicanum, the Axolotl, is the one with the greatest importance in Aztec culture. According to Aztec mythology the Axolotl is one of the representations of the god Xolotl, the monstrous twin brother of Quetzalcoatl. The word Axolotl comes from the Nahuatl word for water (Atl) and for monster (Xolotl). The Axolotl is therefore an aquatic monster, the aquatic representation of one of the most important gods in Aztec culture.
The Axolotl is endemic to Xochimilco, a town that has existed since pre-Hispanic times. This town was declared a site of Cultural Heritage for Humanity by UNESCO due to the series of water canals that house the few remaining Axolotls, since the main lakes and lagoons that made up most of the Valley of Mexico have already disappeared to make way to today’s Mexico City. Due to these drastic changes to its habitat since 1975 that the Axolotl is included in the Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), to regulate its international trade. In Mexico, it was placed under protection by the programmer of Norma Official Mexicana (NOM- 059-ECOL-1994) and has been under special protection since 2001, although this category changed to endangered in 2010. Today, the Axolotl is one of the 51 species of the Priority Conservation Program Priority Species (PROCER). The International Union for Conservation of Nature (IUCN) changed the status of the species from vulnerable to critically endangered in 2006, indicating that this species is under a high risk of extinction.
Ambystoma species as a model for regeneration studies
Salamanders of the genus Ambystoma are the vertebrates with the greatest capacity for cellular and tissue regeneration. The most studied model species is the Axolotl. The great capacity for regeneration and tissue repair displayed by the axolotl has surprised researchers for a long time. The axolotl is therefore the most studied model species in this genus and it has been a study model for over 100 years. Moreover, it is one of the salamanders most easily bred in captivity and for which the most complete and comprehensive genetic, genomic, and transgenic tools have been developed. Previous studies have reported the ability of Axolotls to regenerate retina, tail, legs, spinal cord and some tissues of the heart, after injury or amputation. This makes the Axolotl an important experimental model in the field of regenerative medicine and developmental biology.
Ambystoma species endemic to Mexico
In addition to its regenerative capacity the Axolotl is of scientific interest because it retains the gross morphology of larvae as a reproductive adult, a phenomenon known as neoteny or paedomorphosis. In addition to the Axolotl, three more species (A. andersoni, A. taylori, A.dumerilii) are obligated neotenic, while some others are facultative neotenic like A. velasci. All these species are endemic to Mexico. The 16 endemic species are A. mexicanum, A. taylori, A. andersoni, A. dumerilii (obligated neotenic), A. velasci, A. silvense, A. rosaceum, A. ordinarium, A. lermaensis, A. altamirani, A. amblycephalum (facultative neotenic), A. flavipiperatum, A. rivulare, A. bombypellum, A. leorae and A. granulosum (metamorphic), all of them are endangered at some level.
Ambystoma velasci, different phases of metamorphosis
A typical day in our lab
We normally arrive in the lab at 9 a.m. and go directly to our animal facility, to check that everything is well with the Ambystomas. We do physicochemical tests periodically (measurements of temperature, pH, nitrite, ammonia), and fully wash the dirty tanks while in other tanks only a partial replacement of dirty water for clean and fresh water is made. Water quality is essential to the welfare of Ambystomas; they need temperature of 15-18 °C, pH 6.5-8, nitrite levels no higher than 1.6 mg/L, and ammonia and chlorine concentrations of 0 mg/L. We feed them once a day, or twice when they are juvenile, with earthworms and pellets for carnivorous turtles (see video 1- feeding). These animals are carnivores, and therefore must be well fed as otherwise they may start biting and eating the limbs or gills of their neighbors. Cannibalism is something that occurs more frequently in young animals because they need more food. They are very charismatic animals and have a characteristic big smiling mouth. Ambystomas are very quiet and rarely aggressive except when hungry. When you enter the animal facility and they detect you they swim to the surface of the water indicating they are already hungry, and we feed them well to avoid any nutritional stress, illness or cannibalism.
Video 1. Feeding an Axolotl
In our animal facility we have five of the 16 Mexican species: A. mexicanum, A. dumerilii, A. velasci, A. rivulare and A. granulosum. All of them are endangered species but three of them are in critical condition (A. mexicamun, A. dumerilii and A. granulosum).
A. mexicanum
A. dumerilii
A. velasci A. rivulare A. granulosum
We perform different types of studies with these animals. We are interested in analyzing the molecular circuits that work during regeneration (see video 2), at different time-points. We can amputate a limb and examine the expression pattern of a given transcript or protein by qRT-PCR, in situ hybridization or immunolocalization. We are also interested in the functional characterization of genes and pathways involved in metamorphosis. Since we are not a conservation laboratory, we are less involved in this area, however we are helping in the preservation of these animals by collaborating in genetic and molecular analyses with a conservation group at the Metropolitan Autonomous University campus in Xochimilco. This group has been working in ex situ and in situ conservation of the Axolotl for the last 15 years by designing and applying diverse strategies to preserve them. In our lab we are willing to generate global transcript profiles by RNA-seq for some of the mexican Ambystoma species, using individuals collected from the wild, this is quite important since most genetic and genomic studies have been done so far using animals that have been many years in captivity in labs, since they were collected from Xochimilco (in the case of the Axolotl) almost certainlythese collections have developed a great level of endogamy since, which generates drastic changes in the genetic pool.
In summary, in a normal day, besides keeping the animals and facilities highly controlled and in good shape, we collect samples or dissect tissues from the animals and go back to the bench to perform any of the several experimental strategies previously described. The animals are rarely sacrificed and dissections are made after anesthesia.
Video 2. Axolotl limb regeneration
Perspectives
As I mentioned before, it has not been long since we started working with Ambystomas and my experience with breeding has been a bit complicated. We have tried an artificial method using human chorionic gonadotropin hormone (HCG) in order to know the exact time of fertilization, and we got very good response in the female since we were able to recover a good number of ovules by simple massage. Unfortunately, when we tried the same procedure to extract sperm from the male, to avoid animal sacrifice and pull the vas deferens, we were not successful. We are currently, trying natural breeding and we will test if we are able to standardize the collection of zygotes in the first division-stage or preferably in the one cell-stage (to try transgenesis). Currently, all the animals we work with are obtained from a laboratory dedicated to ex situ reproduction and conservation of Axolotls in conditions as close as possible to the wild.
In the medium and long term, we want to study as many Ambystoma species as possible, to determine the molecular circuits that control developmental aspects ruled by cell reprogramming such as regeneration and metamorphosis. To approach functional genetics, one of our major goals is to establish transgenesis successfully, in order to thoroughly study those genes that are part of the circuits that define regeneration and metamorphosis developmental processes. We are also concerned about how endangered several of these species are. To help with this, we are trying to collect all the Ambystoma species endemic to Mexico in collaboration with experts in Axolotl conservation. We aim to generate a living genetic reservoir and design strategies for ex situ reproduction and re-introduction to the wild, being very careful with the genetic pool, avoiding endogamy and securing wild type-like variability.
“There was a time when I thought a great deal about the axolotls. I went to see them in the aquarium at the Jardin des Plantes and stayed for hours watching them, observing their immobility, their faint movements. Now I am an axolotl.”
From Julio Cortázar’s short story “Axolotl”
Acknowledgements
We thank Tania Quintana Gómez, Gerardo Alejo Jacuinde and Hugo Varela Rodríguez for Video and photo making. We also acknowledge the support of Dr. Luis Herrera-Estrella, Dr. Luis Brieba de Castro and Dr. Mario Arteaga Vázquez and from CIBAC-UAM Xochimilco Biólogo Arturo Vergara and Prof. Fernando Arana.
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
A Postdoctoral Scientist position is available in the Horb Laboratory of the Eugene Bell Center for Regenerative Biology & Tissue Engineering at the Marine Biological Laboratory (http://www.mbl.edu/horb/). The position will focus on the creation of Xenopus models of human disease through the generation of mutants using CRISPR/Cas and TALENs and to work on developing and establishing genome editing in Xenopus.
The postdoc will be focused on creating specific mutants, mainly in X. tropicalis but also in X. laevis, for the Xenopus research community in the US and abroad. In addition, the postdoc will coordinate the efforts of a team of 3-4 personnel to generate 100-200 mutants over 5 years for the research community. The individual will be required to interact with Xenopus researchers on a regular basis in designing and creating these mutants. The individual will also work on knock-in strategies in Xenopus using CRISPR/Cas. The Horb lab is scientifically integrated with the National Xenopus Resource (NXR) at the MBL and the postdoc will be expected to interact with NXR staff on a regular basis.
In addition, the postdoc is expected to work with scientists who visit the NXR throughout the year, since many of them come here to do genome editing. The NXR supports the community by providing transgenic and mutant frogs as well as hosting workshops, providing custom transgenic and mutant services, and hosting scientists through their research hotel service (http://www.mbl.edu/xenopus/).
A postdoctoral position is available immediately to study the transcriptional regulation of retinal progenitor cell proliferation and its interface with cellular bioenergetics. Our lab utilizes a multi-disciplinary approach that combines traditional mouse developmental genetics and molecular biology with live, dynamic tissue imaging and high throughput genomics. The ideal candidate for this position will be within a year of completion of her/his PhD and have expertise in mouse genetics, cell biology, protein and mRNA analysis, recombinant DNA technologies and bioinformatics. Expertise in retinal development, ChIP-seq, 2D-gel Mass Spec. and the characterization of metabolic phenotypes is also highly desirable.
We are a young and vibrant research group located at Baylor College of Medicine within the Texas Medical Center in Houston. Being in the largest medical center in the World has the advantage of fostering a very collaborative research environment and we firmly believe that merging scientific expertise and interests ultimately drive innovation. Therefore, highly creative, independent, but also interactive applicants are particularly encouraged to apply.
To apply please submit your CV, a brief description of your research interests and career goals and the name and contact information of 3 references to Dr. Ross Poché, Department of Molecular Physiology and Biophysics, Baylor College of Medicine. Email: poche@bcm.edu
Here are the highlights from the current issue of Development:
pancRNAs in early development
Promoter-associated noncoding RNAs (pancRNAs) are a class of long noncoding RNAs, transcribed from bidirectional promoters and thought to be involved in promoting expression of the gene transcribed from the opposite strand. Takuya Imamura and colleagues (p.910) now set out to investigate the prevalence and potential functions of such pancRNAs during early mouse development. Focussing on the 2-cell stage, when the major wave of zygotic gene activation (ZGA) occurs, they use directional RNA-seq technology to identify several hundred pancRNAs upregulated in concert with their cognate coding RNA. To assess the potential functional relevance of this co-regulation, the authors analyse three candidates, including Il17d/pancIl17d. In each case, siRNA-mediated knockdown of the pancRNA impairs expression of the mRNA and also prevents the normal DNA demethylation associated with gene upregulation at the ZGA. They further find that depletion of pancIl17d leads to defects in blastocyst survival and in embryonic stem cell colony formation – phenotypes that can be rescued by the provision of recombinant IL17D. Although the mechanisms by which pancRNAs act remain poorly understood, these data provide evidence for an important physiological role for pancRNAs in promoting expression of their partner mRNAs during early development.
Uncovering neurogenic potential of striatal astrocytes
Adult neurogenesis in mammals is generally confined to specific regions where astrocytic cells produce particular neuronal types throughout life. Outside these areas, the capacity for neurogenesis is limited. However, on p.840, Paolo Peretto, Federico Luzzati and co-workers provide evidence that striatal astrocytes can be activated to a neurogenic program in an adult mouse model of Huntington’s disease. Following treatment with the toxin quinolinic acid to induce a lesion, the authors observe the appearance of proliferating progenitors and neuroblasts. Fate-mapping experiments identify local striatal astrocytes as the source of this neurogenic program. Importantly, neurogenesis is only observed upon lesion, suggesting that these astrocytes are normally quiescent but possess latent neurogenic potential upon damage. Further analysis is required to understand the programs regulating this neurogenesis, to determine the final fate of the newly born neuroblasts and to assess whether a similar phenomenon might exist in humans. However, these data open the possibility of harnessing the neurogenic potential of striatal astrocytes for therapeutic purposes.
Timing degradation to tune development
A key issue in developmental biology is how particular processes are coordinated in time: for example, what determines when development of a particular organ starts and how quickly it proceeds? Jennifer Nemhauser and colleagues explore this problem in Arabidopsis, using auxin-mediated regulation of lateral root development as a model (p. 905). Specifically, they investigate the consequence of manipulating the degradation rate of the auxin-responsive transcriptional co-repressor IAA14, the mutation of which is known to affect lateral root formation. Using a synthetic biology approach, the authors generate several versions of IAA14 that show varying degradation kinetics upon auxin stimulation, and then generate transgenic plants expressing each version from the wild-type promoter. They find that both the density and timing of lateral root emergence inversely correlates with IAA14 stability: more stable variants show fewer lateral roots and these roots take longer to initiate. IAA14 is part of a large family of Aux/IAA repressors and these data suggest that regulating the degradation rate of these proteins could act as a tunable timer for developmental progression in plants.
Getting to the heart of heart cell identity
Cardiac progenitor cells differentiate into multiple cell types that make up the functional heart: cardiomyocytes (CMs), smooth muscle cells (SMCs), endothelial cells (EDCs) and fibroblasts. These lineage decisions can be modelled by differentiation of embryonic stem cells (ESCs), but it is not fully clear how closely these in vitrosystems reflect in vivo developmental progression, or how much variability there is within the progenitor population – either in culture or in the embryo. On p.846, Sean Wu and co-workers use single-cell quantitative PCR and lineage-tracing assays on embryonic and adult mouse cardiac cells, as well as mouse ESCs differentiated down the cardiac lineage, to define a gene expression signature for each of the various cell types. Amongst the wealth of data generated, a number of key findings emerge. Firstly, the authors find that ESC-derived CMs closely resemble embryonic and neonatal endogenous CMs, but adult CMs diverge. Secondly, embryonic and ESC-derived cardiac progenitors show different potential: both generate CMs, but embryonic cells can differentiate to EDCs while ESC-derived progenitors produce SMCs. These data demonstrate the power of the single-cell profiling approach and provide valuable insights into lineage choices during cardiac development.
Asymmetric division and fate in the retina
In many systems, neural progenitor cells divide asymmetrically to generate a self-renewing progenitor and a committed neuron. How is this fate segregation controlled, and what defines the balance of proliferation and differentiation? Using live imaging in the developing zebrafish retina, Lucia Poggi and colleagues (p. 832) address these questions, focussing on the role of Anillin – a protein involved in cytokinesis – in the cell divisions that generate retinal ganglion cells (RGCs). By following individual divisions, the authors find that Anillin is itself asymmetrically inherited between daughter cells, and directs the asymmetric inheritance of actin and the polarity protein Par3. Cells with reduced Anillin levels tend to divide symmetrically, generating two RGCs rather than a progenitor and a RGC. Globally, this results in a retina with more RGCs. The authors further show that anillin expression is itself regulated by the RGC fate determinant Ath5, suggesting that there may be feedback loops involving Ath5 and Anillin that control the balance of proliferation and differentiation. How Anillin acts to regulate asymmetric division and fate choice remains unclear, but this technically challenging study demonstrates the importance of this protein in the control of neurogenesis in the retina.
PLUS…
Neural development and regeneration: it’s all in your spinal cord
The latest advances in the field of spinal cord development and regeneration were discussed at a recent EMBO workshop entitled ‘Spinal cord development and regeneration’, which was held in Sitges, Spain, in October, 2014. Here, Catherina Becker and Ruth Diez del Corral provide a review of the workshop. See the Meeting Review on p. 811
A pathway to bone: signaling molecules and transcription factors involved in chondrocyte development and maturation
Decades of work have identified the signaling pathways that regulate the differentiation of chondrocytes during bone formation, from their initial induction from mesenchymal progenitor cells to their terminal maturation into hypertrophic chondrocytes. Here, Elena Kozhemyakina, Andrew Lassar and Elazar Zelzer review how multiple signaling molecules, mechanical signals and morphological cell features are integrated to activate a set of key transcription factors that determine and regulate the genetic program that induces chondrogenesis and chondrocyte differentiation. See the Review on p. 817
Beautiful imaging of zebrafish development
This issue’s featured movie shows the development of a zebrafish, from early embryo to larval stage, imaged using a combination of optical tomography and SPIM. Read the Techniques and Resources article by Huisken and colleagues here.
 (1 votes)
(1 votes)

 (No Ratings Yet)
(No Ratings Yet)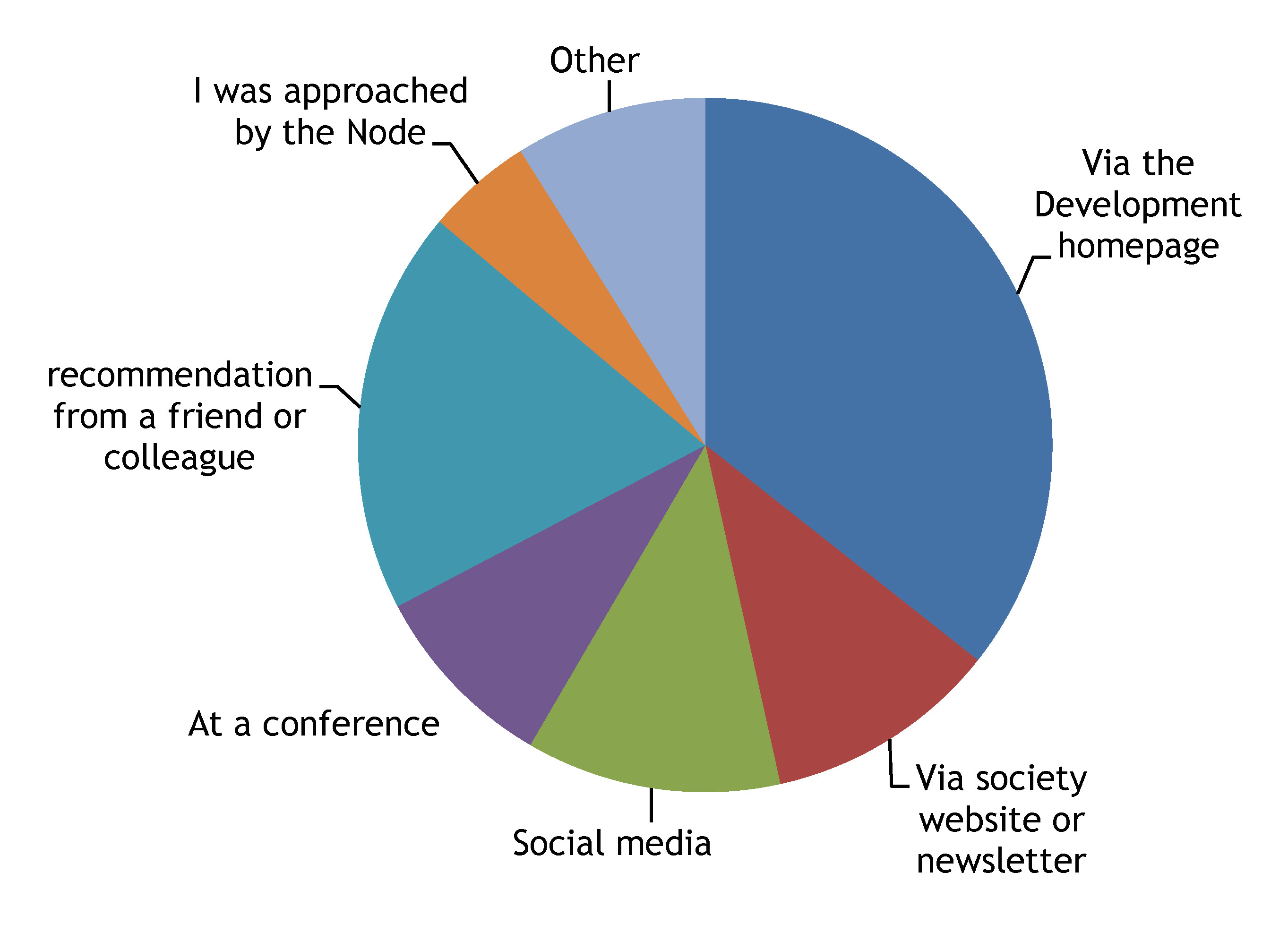









 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series  Promoter-associated noncoding RNAs (pancRNAs) are a class of long noncoding RNAs, transcribed from bidirectional promoters and thought to be involved in promoting expression of the gene transcribed from the opposite strand. Takuya Imamura and colleagues (p.
Promoter-associated noncoding RNAs (pancRNAs) are a class of long noncoding RNAs, transcribed from bidirectional promoters and thought to be involved in promoting expression of the gene transcribed from the opposite strand. Takuya Imamura and colleagues (p. Adult neurogenesis in mammals is generally confined to specific regions where astrocytic cells produce particular neuronal types throughout life. Outside these areas, the capacity for neurogenesis is limited. However, on p.
Adult neurogenesis in mammals is generally confined to specific regions where astrocytic cells produce particular neuronal types throughout life. Outside these areas, the capacity for neurogenesis is limited. However, on p. A key issue in developmental biology is how particular processes are coordinated in time: for example, what determines when development of a particular organ starts and how quickly it proceeds? Jennifer Nemhauser and colleagues explore this problem in Arabidopsis, using auxin-mediated regulation of lateral root development as a model (p.
A key issue in developmental biology is how particular processes are coordinated in time: for example, what determines when development of a particular organ starts and how quickly it proceeds? Jennifer Nemhauser and colleagues explore this problem in Arabidopsis, using auxin-mediated regulation of lateral root development as a model (p.  Cardiac progenitor cells differentiate into multiple cell types that make up the functional heart: cardiomyocytes (CMs), smooth muscle cells (SMCs), endothelial cells (EDCs) and fibroblasts. These lineage decisions can be modelled by differentiation of embryonic stem cells (ESCs), but it is not fully clear how closely these in vitrosystems reflect in vivo developmental progression, or how much variability there is within the progenitor population – either in culture or in the embryo. On p.
Cardiac progenitor cells differentiate into multiple cell types that make up the functional heart: cardiomyocytes (CMs), smooth muscle cells (SMCs), endothelial cells (EDCs) and fibroblasts. These lineage decisions can be modelled by differentiation of embryonic stem cells (ESCs), but it is not fully clear how closely these in vitrosystems reflect in vivo developmental progression, or how much variability there is within the progenitor population – either in culture or in the embryo. On p. In many systems, neural progenitor cells divide asymmetrically to generate a self-renewing progenitor and a committed neuron. How is this fate segregation controlled, and what defines the balance of proliferation and differentiation? Using live imaging in the developing zebrafish retina, Lucia Poggi and colleagues (p.
In many systems, neural progenitor cells divide asymmetrically to generate a self-renewing progenitor and a committed neuron. How is this fate segregation controlled, and what defines the balance of proliferation and differentiation? Using live imaging in the developing zebrafish retina, Lucia Poggi and colleagues (p.  The latest advances in the field of spinal cord development and regeneration were discussed at a recent EMBO workshop entitled ‘Spinal cord development and regeneration’, which was held in Sitges, Spain, in October, 2014. Here,
The latest advances in the field of spinal cord development and regeneration were discussed at a recent EMBO workshop entitled ‘Spinal cord development and regeneration’, which was held in Sitges, Spain, in October, 2014. Here,  Decades of work have identified the signaling pathways that regulate the differentiation of chondrocytes during bone formation, from their initial induction from mesenchymal progenitor cells to their terminal maturation into hypertrophic chondrocytes. Here,
Decades of work have identified the signaling pathways that regulate the differentiation of chondrocytes during bone formation, from their initial induction from mesenchymal progenitor cells to their terminal maturation into hypertrophic chondrocytes. Here,