In Development this week (Vol. 141, Issue 18)
Posted by Seema Grewal, on 2 September 2014
Here are the highlights from the current issue of Development:
Mcc: a new player in gastrulation
 The mutated in molorectal cancer (Mcc) gene has been described as a tumour suppressor, and has been shown to interact with β-catenin and thus limit Wnt signalling. However, various data also indicate a potential role in regulating the cytoskeleton. Ray Dunn and colleagues set out to investigate this further in zebrafish and Xenopus (p. 3505). In both species, they find that morpholino-induced mcc knockdown leads to phenotypes typical of defects in convergence and extension during gastrulation. Importantly, these phenotypes can be fully rescued by co-injection withMcc RNA. In zebrafish, the defects in cellular behaviour are very similar to those seen upon disruption of non-canonical Wnt pathway. Through epistasis and biochemical analysis, the authors provide evidence that Mcc is likely involved in transmitting the Wnt signal from Vangl2 to the downstream effectors RhoA and JNK. Although the detailed molecular mechanism remains unclear, these data identify an important role for Mcc as a component of the non-canonical Wnt pathway that coordinates anamniote gastrulation.
The mutated in molorectal cancer (Mcc) gene has been described as a tumour suppressor, and has been shown to interact with β-catenin and thus limit Wnt signalling. However, various data also indicate a potential role in regulating the cytoskeleton. Ray Dunn and colleagues set out to investigate this further in zebrafish and Xenopus (p. 3505). In both species, they find that morpholino-induced mcc knockdown leads to phenotypes typical of defects in convergence and extension during gastrulation. Importantly, these phenotypes can be fully rescued by co-injection withMcc RNA. In zebrafish, the defects in cellular behaviour are very similar to those seen upon disruption of non-canonical Wnt pathway. Through epistasis and biochemical analysis, the authors provide evidence that Mcc is likely involved in transmitting the Wnt signal from Vangl2 to the downstream effectors RhoA and JNK. Although the detailed molecular mechanism remains unclear, these data identify an important role for Mcc as a component of the non-canonical Wnt pathway that coordinates anamniote gastrulation.Chromatin dynamics in sperm
During spermatogenesis, the chromatin undergoes significant changes in architecture, with histones being largely replaced by protamines, inducing genome-wide condensation. Two papers provide insights into the regulators and mechanisms of this histone-to-protamine transition and its importance for male fertility.
 On p. 3495, Leonard Guarente and colleagues analyse the consequences of SirT1 deletion in pre-meiotic spermatids. SirT1 is an NAD+-dependent deacetylase, the whole body deletion of which has been shown to impair male and female fertility via a systematic effect on reproductive hormone levels. However, whether it is also required in the germ cells themselves is not known. In this article, the authors show that SirT1 depletion specifically in the testis leads to a cell-autonomous defect in sperm maturation and male fertility. They find that histone hyperacetylation – one of the first steps in the chromatin changes of the histone-to-protamine transition – is significantly impaired, with consequent defects in the recruitment of downstream proteins required for histone removal and protamine deposition. Moreover, their data suggest that loss of SirT1 may accelerate reproductive ageing. Although the molecular mechanisms by which SirT1 regulates histone acetylation remain unclear, these data uncover an important role for this protein in the male germline.
On p. 3495, Leonard Guarente and colleagues analyse the consequences of SirT1 deletion in pre-meiotic spermatids. SirT1 is an NAD+-dependent deacetylase, the whole body deletion of which has been shown to impair male and female fertility via a systematic effect on reproductive hormone levels. However, whether it is also required in the germ cells themselves is not known. In this article, the authors show that SirT1 depletion specifically in the testis leads to a cell-autonomous defect in sperm maturation and male fertility. They find that histone hyperacetylation – one of the first steps in the chromatin changes of the histone-to-protamine transition – is significantly impaired, with consequent defects in the recruitment of downstream proteins required for histone removal and protamine deposition. Moreover, their data suggest that loss of SirT1 may accelerate reproductive ageing. Although the molecular mechanisms by which SirT1 regulates histone acetylation remain unclear, these data uncover an important role for this protein in the male germline. Meanwhile, Paul Knoepfler and co-workers investigate the role of the histone H3 variant H3.3 in regulating spermatogenesis (p. 3483). H3.3 is generally associated with active transcription. Here, the authors generate a knockout of one of the two genes encoding H3.3, H3f3b, which leads to a strong reduction in H3.3 levels in the male germline. The mutant mice display a severe defect in sperm morphology and production, and hence in male fertility. Analysis of the chromatin state of H3f3b knockout germ cells reveals an increase in the levels of the repressive mark H3K9me3, and a decrease in H3K4 methylation, a mark of active chromatin. Importantly, the authors uncover defects in the histone-to-protamine transition, with both protamine levels and incorporation being reduced. These data provide the first insights into the role of H3.3 in mammalian spermatogenesis, and indicate an important role for this histone variant in regulating the striking changes in chromatin structure that accompany sperm formation.
Meanwhile, Paul Knoepfler and co-workers investigate the role of the histone H3 variant H3.3 in regulating spermatogenesis (p. 3483). H3.3 is generally associated with active transcription. Here, the authors generate a knockout of one of the two genes encoding H3.3, H3f3b, which leads to a strong reduction in H3.3 levels in the male germline. The mutant mice display a severe defect in sperm morphology and production, and hence in male fertility. Analysis of the chromatin state of H3f3b knockout germ cells reveals an increase in the levels of the repressive mark H3K9me3, and a decrease in H3K4 methylation, a mark of active chromatin. Importantly, the authors uncover defects in the histone-to-protamine transition, with both protamine levels and incorporation being reduced. These data provide the first insights into the role of H3.3 in mammalian spermatogenesis, and indicate an important role for this histone variant in regulating the striking changes in chromatin structure that accompany sperm formation.
Root nodule fate map revealed
 While most plants derive nitrogen (N) from the soil, some plants, such as legumes, can create their own supply by forming specialized root nodules that house N2-fixing bacteria. Nodule tissue ultimately derives from root cells that are reprogrammed to a nodule cell fate during nodule initiation, but exactly which root cells can contribute to the developing nodule primordial has not been fully established. Now, on p. 3517, Ton Bisseling and colleagues unveil a detailed fate map of the origin of different cell types within the Medicago truncatula root nodule. By combining detailed and careful microscopy with promoter-reporter expression analyses, the authors analyse the fate of each root cell layer during nodule initiation. They show that the root inner cortex, endodermis and pericycle divide and transdifferentiate into about 16 cell layers that are located in the basal part of the nodule, whereas the middle cortex reprograms into the nodule meristem. The authors then use these data to re-evaluate previously published root nodule mutants, providing important contextual information for key developmental events during root nodule formation.
While most plants derive nitrogen (N) from the soil, some plants, such as legumes, can create their own supply by forming specialized root nodules that house N2-fixing bacteria. Nodule tissue ultimately derives from root cells that are reprogrammed to a nodule cell fate during nodule initiation, but exactly which root cells can contribute to the developing nodule primordial has not been fully established. Now, on p. 3517, Ton Bisseling and colleagues unveil a detailed fate map of the origin of different cell types within the Medicago truncatula root nodule. By combining detailed and careful microscopy with promoter-reporter expression analyses, the authors analyse the fate of each root cell layer during nodule initiation. They show that the root inner cortex, endodermis and pericycle divide and transdifferentiate into about 16 cell layers that are located in the basal part of the nodule, whereas the middle cortex reprograms into the nodule meristem. The authors then use these data to re-evaluate previously published root nodule mutants, providing important contextual information for key developmental events during root nodule formation.Out of the niche: exploring unknown pathways
 In May 2014, approximately 200 stem cell scientists from all over world gathered near Copenhagen in Denmark to participate in ‘The Stem Cell Niche’, part of the Copenhagen Bioscience Conferences series. The meeting covered an array of different stem cell systems from pluripotent stem cells and germ cells to adult stem cells of the lung, liver, muscle, bone and many more. In addition to the stem cell niche, the meeting focused on a number of cutting edge topics such as cell fate transitions and lineage reprogramming, as well as stem cells in ageing and disease, including cancer. Here, Kateri Moore and Giulio Cossu describe the exciting work that was presented and some of the themes that emerged from this excellent meeting. See the Meeting Review on p. 3441
In May 2014, approximately 200 stem cell scientists from all over world gathered near Copenhagen in Denmark to participate in ‘The Stem Cell Niche’, part of the Copenhagen Bioscience Conferences series. The meeting covered an array of different stem cell systems from pluripotent stem cells and germ cells to adult stem cells of the lung, liver, muscle, bone and many more. In addition to the stem cell niche, the meeting focused on a number of cutting edge topics such as cell fate transitions and lineage reprogramming, as well as stem cells in ageing and disease, including cancer. Here, Kateri Moore and Giulio Cossu describe the exciting work that was presented and some of the themes that emerged from this excellent meeting. See the Meeting Review on p. 3441Roles for Hedgehog signaling in adult organ homeostasis and repair
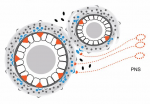 The hedgehog (HH) pathway is well known for its mitogenic and morphogenic functions during development, and HH signaling continues in discrete populations of cells within many adult mammalian tissues. Here, Ralitsa Petrova and Alex Joyner review recently identified functions of HH in modulating the behavior of tissue-specific adult stem and progenitor cells during homeostasis, regeneration and disease. See the Review on p. 3445
The hedgehog (HH) pathway is well known for its mitogenic and morphogenic functions during development, and HH signaling continues in discrete populations of cells within many adult mammalian tissues. Here, Ralitsa Petrova and Alex Joyner review recently identified functions of HH in modulating the behavior of tissue-specific adult stem and progenitor cells during homeostasis, regeneration and disease. See the Review on p. 3445
piRNAs: from biogenesis to function
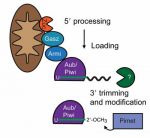 Since their discovery less than a decade ago, Piwi-interacting RNAs (piRNAs) have been shown to repress transposable elements in the germline and, hence, have been at the forefront of research aimed at understanding the mechanisms that maintain germline integrity. More recently, roles for piRNAs in gene regulation have emerged. In their Review, Eva-Maria Weick and Eric Miska highlight recent advances made in understanding piRNA function, highlighting the divergent nature of piRNA biogenesis in different organisms, and discussing the mechanisms of piRNA action during transcriptional regulation and in transgenerational epigenetic inheritance. See the Review on p. 3458
Since their discovery less than a decade ago, Piwi-interacting RNAs (piRNAs) have been shown to repress transposable elements in the germline and, hence, have been at the forefront of research aimed at understanding the mechanisms that maintain germline integrity. More recently, roles for piRNAs in gene regulation have emerged. In their Review, Eva-Maria Weick and Eric Miska highlight recent advances made in understanding piRNA function, highlighting the divergent nature of piRNA biogenesis in different organisms, and discussing the mechanisms of piRNA action during transcriptional regulation and in transgenerational epigenetic inheritance. See the Review on p. 3458
 (No Ratings Yet)
(No Ratings Yet)

 (1 votes)
(1 votes)
![admin-ajax[1]](https://thenode.biologists.com/wp-content/uploads/2014/08/admin-ajax1.jpg)











 (8 votes)
(8 votes)