This is part of the ‘Lab meeting’ series featuring developmental and stem cell biology labs around the world.
Where is the lab?
Andrea: You can find the Ditadi lab at Ospedale San Raffaele, as part of the San Raffaele Telethon Institute for Gene Therapy, in the north-east corner of Milan, Italy. Milan is a great spot for both science and life, with a myriad of places to visit, plenty of things to do and a rich community of great labs to collaborate with.
Andrea: We want to understand how human blood cells form. For this, we use human pluripotent stem cells as a model, integrating developmental, cell and molecular biology, as well as a bit of immunology. We study human developmental biology in a dish: we study early mesoderm patterning and follow the process all the way to mature blood cells, including hematopoietic stem cells, trying to work out which signals guide each step. We are developmental biologists working at an institute that focuses on genetic diseases and their therapy, so we also use the cells we generate to look at diseases from a developmental perspective. At the same time, we explore how to engineer and arm these cells in unique ways so they can be used in clinical settings in the future.
Lab roll call
Let’s start in order of length of service in the lab.
We have Lauren Randolph, post-doctoral fellow, who is studying how hemogenic cells give rise to blood.
Claudia Castiglioni, PhD student, who aims to identify the earliest commitment to blood cell fate.
Riccardo Piussi, former Master’s student now PhD student-wannabe (and hopefully soon-to-be), working to decipher the regulation of self-renewal in emerging HSCs.
Deborah Donzel, a postdoctoral fellow, and Nikita Pinto, another former Master’s student now turned research assistant, are partners in modeling a ribosomopathy that affects red blood cells only postnatally to decipher proteostasis regulation across different stages of hematopoietic development.
Elena Morganti, a postdoctoral fellow, and Bianca Nesti, a Master’s student, who teamed up to model a pediatric autoimmune disease as a way to understand the role of embryonic lymphocytes in health and disease.
Alessandra Guerreschi, a Master’s student who recently joined our lab and is gearing up to investigate the multiple roles of Notch signaling in hematopoietic development.
Favourite technique, and why?
Andrea: It is not exactly a technique, but my favourite moment in the lab is simply watching cells under the microscope. We do not do much imaging; most of our days are spent in the hood doing cell culture. Even now, when I am sadly not doing many experiments any longer, I still have this habit that I actually stole from my postdoc advisor. When I need a break from the desk and the administrative tasks, I go to the lab for what I call a bit of “cell therapy”. I grab a few plates and spend some time simply looking at cells under the microscope. I love it. Observing cells in cultures is very informative, cells talk to us all the time.
If I need to choose a proper technique, I would choose flow cytometry. We use it a lot. It may not be as high-throughput as some newer methods, but it gives us robust full gene expression data at the single-cell level, and we can learn a lot from what comes out of the cytometer.
Andrea, apart from your own research, what are you most excited about in developmental and stem cell biology?
Andrea: Recently, I have been following the evo-devo field with a lot of interest. I find it fascinating to think about how cells and tissues evolved, and for a lab like ours that tries to recreate how blood cells are formed in vitro, understanding how they appeared in the first place feels very relevant.
Another field that I find extremely exciting is synthetic biology. I am fascinated by how we can now “prod” cells and systems and modify their responses. I remain a developmental biologist at heart, but the environment where we work has opened my eyes to how we can push the boundaries of therapeutic innovation. Alongside the clinical application of stem cells, synthetic biology is transforming the way we think about medicine and how we might design future therapies.
Andrea, how do you approach managing your group and all the different tasks required in your job?
Andrea: I am not sure I can say I am set into one approach, at least yet. I think it is always evolving, as the people in the lab, as well as the lab itself, need change over time. In general, I try to spend time getting to know the people in my group, recognizing the strengths and weaknesses, and trying to exploit the former while helping them work on the latter. I often think of the group as an orchestra or a music band. First, I need to hear the sound of each instrument, help them get tuned and then my job is to compose some music that fits them. Let’s say that some composition takes more time than others. But in the end, the goal is to nurture the love and passion for the true privilege of doing research for everyone.
As for managing the different tasks, I often wish I had more hours in the day; that would be a great superpower. So, I try to clear out the things I do not enjoy, the administrative duties and emails, as quickly as possible. This gives me protected time for what I love: reading, thinking and spending time with the team in the lab. I am not sure I always find enough time for that, but I try very hard.
What is the best thing about where you work?
AD: Without a doubt, being surrounded by young and bright people. It is energizing and another privilege of this job.
CC: The thing I value most about being at SR-Tiget is the stimulating environment, where science truly comes alive. Ideas are shared freely, we have the resources to bring them to life, and we constantly get to learn from seminars by scientists from around the world.
NP: The best thing about working at SR-Tiget in Milan is the combination of different scientific topics and a truly collaborative environment, where you can walk into a lab or an office to ask for help and know that someone will genuinely take the time to help you solve a problem.
RP: What I like most about where I work is the general drive of the institute to do high-level science and to set ambitious goals. In the lab, I really appreciate the way we reason scientifically and the fact that we constantly challenge our ideas by asking questions every day.
LNR: The best thing about where I work is the science and the people. I really enjoy the project that I am working on and find it both challenging and engaging. I am also really lucky to work with incredibly collaborative and supportive colleagues who really treat the lab as a family. It makes it a joy to spend time with them, both in and out of work, and to do and talk science together.
DD: The best things about where I work are the research topic and the people I work with. My enthusiasm for the project keeps me focused and driven, even during challenging periods. I’m also fortunate to work with colleagues who are open to sharing ideas and knowledge, which creates a collaborative environment that helps us move forward together.
BN: What I appreciate most about working at SR-Tiget is the highly stimulating scientific environment, both at the institute level and within my own laboratory. The presence of diverse expertise, frequent seminars and strong resources fosters a continuous exchange of ideas and supports high-quality research.
EM: What I like most about where I work is the young and supportive environment. I feel that people around me are genuine and open-minded, and this makes my days very pleasant and enjoyable.
AG: Even though I haven’t been here long, I’ve really noticed how welcoming and supportive everyone is. It makes it easy to ask questions, learn quickly and feel like part of the team right away.
What’s there to do outside of the lab?
AD: Despite being in love with my job and not feeling the need to escape, life is too short, and I have so many interests – books, music, sport, hiking, biking, food, friends, etc. – so I try to do a bit of it all. To be coordinated with the family, in particular, two kids who keep me happy and busy.
CC: Outside of the lab, I really enjoy canoeing on the Navigli, the famous canals in Milan. Being on the water allows me to slow down and take a break from the busy pace of the lab. I love the feeling of paddling along the canals, enjoying the surroundings and reconnecting with the city.
NP: Having grown up in Milan, I sincerely love this city and everything it offers. Outside of the lab, I like different things, from baking and crocheting to spending time with family and friends while enjoying the city’s cultural life, like its aperitivo culture and different neighborhoods. Recently, I also joined the Red Cross as a volunteer, where I am involved in social inclusion activities with homeless people, as well as assistance roles during public events. These experiences help me stay grounded, connected to the community and maintain balance alongside research.
RP: This job takes a lot of time and energy, but outside of the lab I really enjoy spending time with my family and friends. I also love fishing. I enjoy it for its unpredictability and complexity; it requires analyzing many variables and accepting failure without expectations. Every small decision can make a difference, and while nothing is guaranteed, everything is possible, like in science.
LNR: Outside of the lab, I enjoy traveling, reading, and all things food-related. In Milan, I particularly enjoy access to the ballet, opera, and theater.
DD: Outside of the lab, I really enjoy going for walks—especially in parks or outside the city, where I can reconnect with nature. Living in Milan, I also like going to the theater and meeting friends for an aperitivo.
BN: Outside the lab, I enjoy spending time reading, as it offers a break from continuous scientific reflection while still keeping my mind engaged in a pleasant way. I also like to take advantage of the many cultural and recreational initiatives that Milan has to offer, often in the company of my friends.
EM: I usually try to spend time in nature and clean air when I am not in the lab. Milano is really close to beautiful mountains and lake,s and those are my favorite spots for the weekend. I also enjoy food, art and history.
AG: In my free time, I enjoy reading and spending time in the mountains outside of Milan, whether it’s hiking, skiing or horseback riding. Skiing, in particular, is a great way to unwind on the weekends and enjoy the outdoors. Being able to combine outdoor activities with some quiet time to read makes my free time really enjoyable.
Biology probes form and function of Life. Form is easy to grasp: cells under a microscope, subcellular structures in electron micrographs, or organisms on camera readily present their shapes.
Figure 1: Examples of microscopy images that allow understanding of Life: Fruit fly ovaries development and the changing RNA localizations during the process. Jambor, 2015
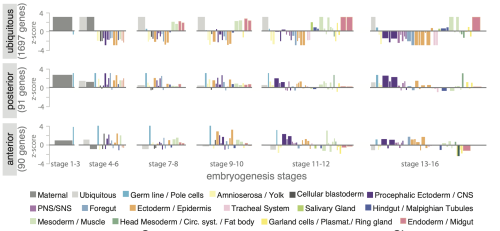
Function is different: it emerges from molecular compositions, interactions, and temporal changes. Such data is not directly visible – we use statistics to make sense of it. But summaries and p-values alone rarely reveal how complex biological systems are organized, the variability in the samples and the resulting uncertainty in the data, or unexpected relationships and pattersn. As datasets grow larger and more complex, these insights only become accessible when data are visualized.
Figure 2: Statistical chart comparing RNA localizations across development Jambor et al.
Despite being used widely, data visualization is still treated as a final step in research, a way to communicate results once the real analysis is finished. In reality, visualization plays a much earlier and more fundamental role. Visuals expose batch effects, hidden subpopulations, nonlinear behaviors, and experimental artifacts that often remain invisible to summary statistics alone. These insights directly shape which data can be trusted, which controls are needed, and which experiments should come next.
Figure 3: Examples of plots used during the research process to visualize technical and sample variability. Figure 3: Examples of plots used during the research process to visualize technical and sample variability.
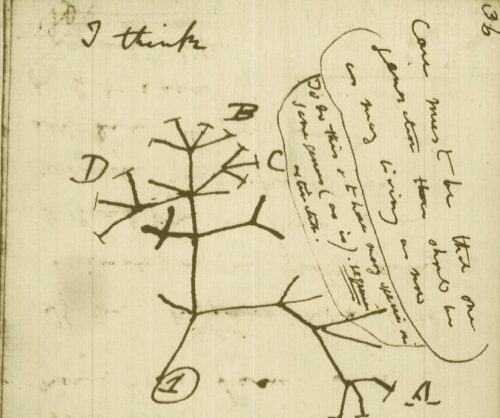
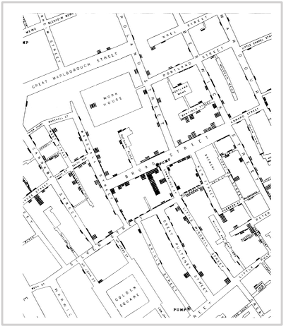
While the urgency to visualize data feels modern, the principle itself is not new. Seeing has always been central to biological understanding. Darwin’s and Linnaeus’s classification of species relied on careful visual comparison. In the nineteenth century, Florence Nightingale pioneered statistical charts to reform healthcare, while John Snow’s maps of cholera outbreaks transformed how disease transmission was understood. In the twentieth century, Michaelis and Menten introduced the kinetic plot as a standardized visual language for enzyme activity, and more recently, interactive genome browsers have made entire genomes navigable at nucleotide resolution.
Darwin’s phylogenetic treeSnow’s Cholera mapightingale’s chart invented to document health care reformsMarey’s animation of human locomotion
Today, data visualization is however still poorly formalized in the life sciences. It lacks dedicated training programs, shared standards, and institutional recognition. This gap matters, as data visualization leads to hypotheses generation, insightful data presentation, and builds trust in the results.
Just as early scientists needed training in scientific drawing to accurately document what they observed, today’s researchers must learn to engineer and interpret data visualizations with comparable rigor. In an era where biology increasingly unfolds in data rather than images alone, learning how to see again has never been more important.
PS – I wrote this looking for discussions on this topic – feel free to reach out helena.jambor – at – fhgr.ch
Reflections from a Workshop by Nivedita Mukherjee1 and Mateus de Oliveira Lisboa2
1National Centre for Biological Sciences, Tata Institute of Fundamental Research, 560065, India (NCBS-TIFR)
2Core for Cell Technology, School of Medicine and Life Sciences, Pontifícia Universidade Católica do Paraná (PUCPR)
Last November, a unique gathering took place at Fanhams Hall in Hertfordshire, UK, for The Company of Biologists’ workshop titled “Decoding Whole Genome Doubling: Insights from Evolution, Development, and Disease.” Organised by Renata Basto and Zuzana Storchova, the workshop brought together a highly interdisciplinary cohort of scientists studying whole-genome doubling (WGD) across organisms, from flies and frogs to plants and humans.
As two of ten early-career researchers selected to participate, we had the rare opportunity to engage in close discussions with leaders in the field, present our own work, and explore the diverse biological contexts in which the entire chromosomal complement of a cell is doubled, changing its ploidy – the number of copies of the genome. Across three intense days, we uncovered how such changes can drive cellular adaptations, precipitate disease, and drive evolutionary innovation.
A Biological Balancing Act
Ploidy variation is ubiquitous in biology. Even within a single organism, cells often carry different numbers of genome copies depending on the tissue type and developmental stage. For example, while humans are diploid as a species, polyploid cells are routine in organs such as the liver, heart and pancreas. These can arise from either programmed or accidental WGD, resulting from the skipping of one or more cell-cycle steps or through cell fusion. Once polyploidy arises, the cell is thrust into a new regime, one that demands extensive adaptation if it is to continue functioning or proliferating without compromising genome integrity.
Initial responses to genome doubling can be defensive. DNA damage and stress-response pathways are activated, and cell death is a common outcome unless these surveillance mechanisms are suppressed. Survival alone, however, is not enough. To continue to divide successfully, polyploid cells must carefully segregate their excess chromosomes. This often involves re-establishing functional mitotic spindles despite the presence of supernumerary centrosomes. Several talks highlighted molecular strategies that allow polyploid cells to survive and divide by rewiring apoptotic signalling and stabilising centrosome numbers.
Bigger Cells, Bigger Questions
Polyploid cells aren’t just more complex; they’re physically larger. This increase in volume comes with consequences: intracellular transport must traverse greater distances, metaphase spindles must span wider plates, and increased metabolic demands must be sustained. Several speakers described how these physicochemical constraints can be systematically probed using in silico models alongside in vitro perturbations.
Beyond these geometric and energetic challenges, it is tempting to assume that increased DNA content translates to increased gene expression. However, multiple presentations demonstrated that gene expression scales unevenly and nonlinearly with ploidy, with pronounced disparities between transcript abundance and protein yield. In some contexts, specific genes are selectively up- or down-regulated, rewiring the cell’s regulatory network in unexpected ways. This decoupling of genome content from functional output overturns the view of polyploid cells as merely “amplified” versions of their diploid counterparts.
Genomic Instability and Interventions
For a cell, doubling its genome is not always a deliberate developmental strategy or a trivial bookkeeping error; it is often a high-risk gamble. Cardiomyocytes, for instance, become increasingly polyploid with age or heart disease, accompanied by a substantial accumulation of mutations. Whether polyploidy in such settings is adaptive, maladaptive, or merely tolerated remains an open question—one that echoes broader uncertainties about where normal physiology ends, and pathology begins.
Polyploidy-related genomic instability is also characteristic of cancer, with genome-doubled tumours showing many more chromosomal aberrations than diploid tumours. While high levels of such aberrations can be lethal, genome doubling also casts a paradoxical safety net by creating “genomic backups” that restore essential functions. At its extremes, WGD-induced genomic instability can be spectacular. One striking example is chromothripsis, where mis-segregated chromosomes trapped within micronuclei shatter and reassemble in chaotic ways, creating the genomic equivalent of a misassembled jigsaw puzzle.
A silver lining is that, while such drastic mutational events can accelerate cancer cell evolution, their dependence on these aberrant states may also be their undoing. Because WGD imposes unique cellular stresses, polyploid cancer cells acquire distinct vulnerabilities that may be therapeutically exploitable. The workshop highlighted several efforts to leverage these weaknesses, offering hope for selective treatments that preferentially target cancer cells while sparing normal tissue.
Innovation Across Evolutionary Time
At the macro scale, genome doubling acts as a powerful driver of evolutionary change. Ancient WGDs have profoundly shaped the genomes of present-day microorganisms, plants, and vertebrates, with recent events producing drought-resistant plants, pest-resistant crops, and amphibians adapted to arid environments. Polyploid lineages often adapt rapidly, restructuring physiological and metabolic pathways, and frequently adopting self-pollination or asexual reproductive strategies to overcome early barriers to evolutionary establishment.
Across the tree of life, WGD is frequently followed by episodes of rapid diversification, though these bursts typically occur only after a delay. This “radiation lag” is thought to reflect the time required for re-diploidisation, during which redundant genomic content is reduced, and ohnologues or duplicated genes diverge in function. As gene dosage is re-balanced and new regulatory networks emerge, WGD becomes a springboard for long-term evolutionary innovation.
Such innovations may confer resilience during periods of ecological upheaval. Compelling evidence for this comes from the observation that WGD events in the evolutionary history of species are disproportionately clustered around major extinction events, such as the Cretaceous–Palaeogene boundary. Interestingly, environmental stressors, such as extreme heat or cold, can induce the formation of unreduced gametes, providing a direct link between ecological pressures and the origin of polyploid species.
A Shared Language for Polyploidy
Amid the breadth of scientific discussions at the workshop, one unifying theme stood out: WGD is neither an anomaly nor a biological accident. From plants surviving climate shifts to tumours evading physiological checks, genome doubling repeatedly emerges as a powerful strategy in life’s toolkit. Yet it remains a double-edged sword, capable of driving adaptation or unleashing instability.
This raises big questions. In healthy tissues, polyploidy seems to balance on a knife-edge between careful regulation and stochasticity. Understanding this balance could reveal why certain cells become polyploid, how tissues keep their abundance in check, and what stops them from turning cancerous. In cancer cells, it’s still unclear if WGD is a driving force or a downstream manifestation of genomic instability. Moreover, scientists are still figuring out how evolutionary pressures shape newly formed and established polyploid lineages, and how the opportunities for adaptation and diversification play out across different tissues and species.
What the field now needs is a shared language across disciplines. Polyploidy in microbes, plants, animals, cancer, and development has too often been studied in silos. By integrating these perspectives, we may finally decipher how a single genomic event reverberates from the scale of individual cells all the way to the evolution of multicellular species.
This workshop marked a step in that direction, and for us, a turning point in our understanding of what it means to live with, adapt to, and evolve through an extra genome.
Mateus de Oliveira Lisboa is a PhD student at the Core for Cell Technology, PUCPR (Brazil), studying how whole-genome doubling shapes cell fate, evolution, and disease. With a background in chromosome biology, stem cells, and the cancer microenvironment, he integrates molecular biology and bioinformatics to explore the causes and consequences of large-scale genomic alterations, always through an evolutionary lens. Outside the lab, he explores mountains, photography, astronomy, and birds.
Nivedita Mukherjee is a PhD student at the National Centre for Biological Sciences (NCBS–TIFR), Bengaluru, India, where she studies cancer evolution through the computational analysis of large-scale genomics datasets. Her research integrates statistical genomics and evolutionary theory to examine how whole-genome doubling reshapes selective pressures in cancer. Beyond research, Nivedita writes popular science articles and enjoys singing, travelling, photography, and reading.
[Editorial from Development by James Briscoe, Swathi Arur, Anna Bigas, Dominique Bergmann, Benoit G. Bruneau, Cassandra G. Extavour, Paul François, Anna-Katerina Hadjantonakis, Haruhiko Koseki, Thomas Lecuit, Matthias Lutolf, Irene Miguel-Aliaga, Samantha A. Morris, Kenneth D. Poss, Elizabeth J. Robertson, Peter Rugg-Gunn, Debra L. Silver, James M. A. Turner, James M. Wells, Steve Wilson.]
Every researcher knows the anticipation and trepidation that come with submitting a paper to a journal. Years of effort have been distilled into a few thousand words and a handful of figures containing the metaphorical (and often literal) sweat from long hours and hard toil in the lab. What will the reviewers say? How will the editor deal with it? At Development, we understand the anxiety and the investment that goes with a paper submission. Our mission is to provide the kind of expert, constructive review that not only evaluates your work but helps it achieve its full potential for lasting scientific impact. But we know that the comments provided by reviewers don’t always live up to this expectation. We’ve heard the concerns. Some of these frustrations reflect deeper, systemic issues across scientific publishing: the feeling that revision requests can expand beyond what is feasible, that editorial decisions are not always transparent, and that standards can seem uneven across subfields such as developmental and stem cell biology. These are not challenges unique to Development, but we acknowledge them and continue to refine our editorial practices to address them wherever possible.
We’ve been told that Development is ‘too hard to publish in’, that reviews are unnecessarily harsh, that revisions are excessive and time-consuming. These criticisms matter to us. Even though sometimes it might be more perception than reality, we won’t pretend that there isn’t some truth in these criticisms. As active research scientists ourselves, we Editors face similar frustrations with our own papers. We want to be transparent about Development’s reviewing process and explain how we endeavour to get the balance right so that it serves both authors and the scientific community.
First, the numbers behind the perception. The bottom line is that, for the past 10 years, 35-45% of papers submitted to Development ultimately get published. Let us break this down. Roughly 65% of the manuscripts submitted to Development are sent out for peer review. We only editorially reject papers when the topic of the article is beyond our scope and expertise, or it is clear to us that the study would not be supported by our peer reviewers. A rapid rejection allows authors to quickly redirect their manuscript to more appropriate venues and, where relevant, we facilitate direct transfers to our sister journal, Biology Open. Of the papers we send for peer review, well over 50% receive positive reviews from reviewers and we ask the authors to revise and resubmit these…
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Developmental regulation of intestinal best4+ cells Abhinav Sur, Ella X. Segal, Michael P. Nunneley, Jason W. Sinclair, Morgan Kathleen Prochaska, Louis E. Dye, Yalan Wu, Liezhen Fu, Yun-Bo Shi, James Iben, Benjamin Feldman, Jeffrey A. Farrell
Cytoplasmatic polyadenylation of mRNA by TENT5A is critical for enamel mineralization Goretti Aranaz-Novaliches, Olga Gewartowska, Frantisek Spoutil, Seweryn Mroczek, Pavel Talacko, Karel Harant, Ana-Matilde Augusto-Vale, Irena Krejzova, Carlos Eduardo Madureira Trufen, Pawel Krawczyk, Ales Benda, Vendula Novosadová, Radislav Sedlacek, Andrzej Dziembowski, Jan Prochazka
Functional architecture of cardiac TF regulatory landscapes in control of mammalian heart development Virginia Roland, Johannes Tüchler, Andrea Esposito, Mattia Conte, Matteo Zoia, Ekapaksi Wisnumurti, Virginie Tissières, Julie Gamart, Raquel Rouco Garcia, Ines J. Marques, Akshay Akshay, Vincent Rapp, Brandon J. Mannion, Jennifer A. Akiyama, Prateek Arora, Harry Walker, Ali Hashemi Gheinani, Beth A. Firulli, Gretel Nusspaumer, Anthony B. Firulli, Guillaume Andrey, Axel Visel, Nadia Mercader, Javier Lopez-Rios, Mario Nicodemi, Iros Barozzi, Marco Osterwalder
A Single Cell Atlas of the Newt Iris During Lens Regeneration Olivia M. Williams, Kelsey E. Ahearn, Joseph L. Sevigny, Nicole Farber, Disha Hegde, Kenneth J. Lampel, Jenna Loporcaro, Leo Napoleon, Jacob Nipoti, Timothy Ralich, Brooklyn Wallace, W. Kelley Thomas, Konstantinos Sousounis
The germline-restricted chromosome orchestrates germ cell development in passerine birds Niki Vontzou, Yifan Pei, Israel Campo-Bes, Wolfgang Forstmeier, Moritz Hertel, Manuel Irimia, Bart Kempenaers, Sylvia Kuhn, Katrin Martin, Jakob C. Mueller, Kim Teltscher, Annelie Mollbrink, Xesús Abalo, Matthew T. Biegler, Simone Immler, Francisco J. Ruiz-Ruano, Alexander Suh
Development of the Early Childhood Duodenum across Ancestry, Geography and Environment Joshua de Sousa Casal, Krishnan Raghunathan, Chelsea Asare, Abigail Plone, Nazanin Moradinasab, Junaid Iqbal, Lianna F. Wood, Elsy M. Ngwa, Xia Chen, S. Fisher Rhoads, Clara Baek, Dur-e Shahwar, Neha S. Dhaliwal, Madison Wong, Max Garrity-Janger, Lily P. Gillette, Stephanie Regis, Fatima Zulqarnain, Asra Usmani, Jason D. Boisvert, Casey R. Johnson, Jackson Larlee, Michael D. Anderson, Daniel Zeve, Elisa Saint-Denis, Thomas G. Wichman, Jeffrey La, Ashish Jain, Liang Sun, Lauren Scudari, Natalie N. Bhesania, Zehra Jamil, Michelle Galeas-Pena, Adam R. Greene, Aneeta Hotwani, Fedaa Najdawi, Shyam S. Raghavan, Donald E. Brown, Christopher A. Moskaluk, Heather H. Burris, Piotr Sliz, Phyllis R. Bishop, Scott B. Snapper, Kamran Sadiq, Sarah C. Glover, Muhammad Imran Nisar, Sana Syed, Jocelyn A. Silvester, Jose Ordovas-Montanes, Jay R. Thiagarajah
The 2026 Loke CTR annual meeting theme, “Epigenetics of embryogenesis and placentation” brings together leading clinical and basic scientists to explore topics including the interplay of environment, genetics and epigenetics in DOHaD, mechanisms of cell fate and lineage development, genomic imprinting, X chromosome inactivation, and the role of repetitive elements in trophoblast and early development.
Where: Old Divinity School, St John’s College, All Saints Passage, Cambridge CB2 1TP
Pedro Martinez. Departament de Genètica, Microbiologia i Estadística, Universitat de Barcelona
Gene Regulatory Networks: An Introduction and Their Historical and Conceptual Context.
The concept of a gene regulatory network has, by now, a long history. It was essentially developed in the papers that Britten and Davidson published in 1969 and later. In these works, they proposed the idea that genes—particularly transcription factors—constitute the underlying mechanisms that causally explain development and evolution. This paradigm shifted earlier views of developmental processes as “linear” epistatic relationships, which had incorporated concepts such as “master regulatory genes” positioned at the top of epistatic hierarchies. Although ideas like pleiotropy had long been recognized, implying that linear sequences of gene action were unrealistic representations of how genes build structures, the notion that development could still be depicted as a set of linear processes in which transcription factors act on one another remained attractive and was, in some sense, reinforced by classical developmental genetics.
The advent of genomic technologies and the ability to study interactions between numerous transcription factors and cis-regulatory sequences enabled a more complex view of development. In this framework, many factors act through mutual interactions, in hierarchical architectures, driving processes forward in a largely directional manner. Gene regulatory networks (GRNs, from now on) were instrumental in giving mechanistic form to older concepts such as developmental trajectories in Waddingtonian landscapes, where developmental decisions were visualized as the movement of a system downstream along a valley shaped by both external physical constraints and internal constraints built into the architecture of the network. Moreover, GRNs offered hints about what “emergent properties” may depend on—phenomena in which novel, nonlinear properties arise at different biological scales, even though the proposed underlying explanations were never entirely clear.
However, as with many concepts introduced in science, the use of the term gene regulatory network has gradually lost precision. In a substantial fraction of articles, it is employed in a very loose way. This trend has run parallel to the pressure to use “fashionable” terminology in papers, grant proposals, reports, and other scientific outputs—a problem that is not foreign to many disciplines.
Gene Regulatory Networks: Clarifying What Counts—and What Does Not
A Gene Regulatory Network is a graphical representation of the physical interactions between transcription factors (TFs) and their target cis-regulatory regions that drive a specific developmental process. The representation aims at completeness: it should indicate the full set of transcription factors involved, as well as all the cis-regulatory elements to which they bind (often located in other transcription factors). Needless to say, TFs also regulate other classes of genes, including those coding for signalling molecules and structural proteins. These gene products are, of course, essential for establishing intercellular communication and for generating the specific phenotypes of cells and tissues in which they are expressed. However, TF networks are the most relevant component because they constitute the driving engine of the process. TFs are the true, physical effectors that modulate new gene activities and are ultimately responsible for pushing development forward.
While all of the above may seem almost self-evident, what is striking in many papers is that the links between TFs and their regulatory binding sites are often hardly demonstrated. There is no such thing as a network of factors if the interactions are not experimentally established; otherwise, the implied causality collapses. Once again, we end up with mere correlations between TF activities, without any explanation of how the interactions drive specific parts of the developmental process. A network cannot be based on the assumption that correlations between expression patterns—whether in natural conditions or after gene perturbations—and the presence of putative open chromatin sites are sufficient, even if all these data appear together in correlated datasets (e.g., ATAC-seq profiles or transcriptomic datasets). TF binding relationships must be demonstrated. Moreover, the binding events must not only occur but also be functionally significant. For instance, binding sites may be occupied without being functional.
In this context, only those representations whose nodes and edges have been experimentally verified can truly be called networks. Moreover, we should be cautious about calling a small set of genes a network. There is an obvious impossibility in representing a bona fide network with a very limited number of components. A good network should aspire to completeness, and this requires a substantial number of experimentally demonstrated interconnections.
Alternatively, what we often have is merely a collection of correlated data, which may generate preliminary hypotheses. These hypotheses must then be rigorously tested if they are to become genuine GRNs with any degree of predictive power.
It is the completeness of a GRN that provides a genuinely mechanistic understanding of a developmental process—not merely the description of the actors involved, even when some interactions can be demonstrated. By definition, causal explanatory frameworks (Johansson et al., 2024) should always include:
Linking antecedent causes to subsequent effects;
Describing the pathways from cause to effect (not just establishing a correlation);
A clear temporal sequence, in which the cause must precede the effect; and
An explanation of the “why” of a process, offering reasons for the observed events or states.
In this context, only GRNs reveal the causal and mechanistic relationships needed for a veritable (and verifiable) understanding of development. This does not mean that GRNs are the only explanatory tool—physical forces and complex cell behaviors also account for complementary aspects of developmental processes. However, these should be integrated into a holistic framework in which the core explanatory level is provided by the nuclear events that contribute to build a GRN.
REFERENCES
Britten RJ, Davidson EH. Gene regulation for higher cells: a theory. Science. 1969 Jul 25;165(3891):349-57. doi: 10.1126/science.165.3891.349.
Johansson, LG. et al. (2024). Causal Explanations. In: A Primer to Causal Reasoning About a Complex World. Springer Briefs in Philosophy. Springer, Cham. https://doi.org/10.1007/978-3-031-59135-8_8
Peter, I. S., & Davidson, E. H. (2015). Genomic control process : development and evolution / Isabelle S. Peter, Eric H. Davidson. (1st ed.). Elsevier. (for a general review)
1Department of Microbiology and Immunology, University of Rochester Medical Center, Rochester, NY 14642, USA
2Department of Environmental Medicine, University of Rochester Medical Center, Rochester, NY 14626, USA
Introduction
Plastic particles and fibers shed from plastic debris, termed microplastics (MP), have become ubiquitous environmental pollutants found everywhere globally throughout marine and freshwater ecosystems (reviewed in [1]). Improper disposal, accidental loss, and fragmentation of plastic materials have led to an increase in MPs, which range in size from as large as 5 mm to as small as 1 mm. These MPs which can also be airborne, pollute environments as varied as urban landscapes, remote terrestrial regions, aquatic ecosystems. One of the highest waterborne MP concentrations reported in the USA is where Rochester’s Genesee River meets Lake Ontario [2]. In the air, soil and water, these MPs are consumed by a wide variety of organisms from invertebrates including mollusks and crustaceans to vertebrates such as fish, amphibians and ultimately humans. In humans, MPs accumulate in breast milk as well as various organs and tissues including the brain , liver, and placenta [3, 4]. Recently, MPs present in human tissues have increased from 2016 to 2024, especially in the brain [5]. While there is increasing evidence suggesting that MPs pose serious threats to aquatic ecology and human health, many aspects of their potential biological activity remain unclear. Notably, little is known about the potential lasting impacts of exposure to MPs during early development on immunity.
The study of biological effects of MPs has revealed multiple challenges including the wide diversity of plastic types that may or may not induce similar effects and the need for reliable biological models. While it is estimated that there are over 5000 different types of plastics composed of different polymers (e.g., nylon, polyethylene terephthalate, etc.) and chemical additives (e.g., flame retardants, plasticizers, etc.), the large majority of biological studies of MPs have used manufactured sterile polyethylene or polystyrene spherical beads of uniformized size. Moreover, under the actions of UV, temperature and pH in the environment, plastics fragment into a myriad of sizes and shapes, while their physical properties (e.g., porosity, hydrophobicity) are further altered during this process of aging. Finally, plastic debris sinking in water is associated with the formation of biofilms composed of diverse microbial communities, which may include pathogens [6].
To help fill these current gaps in knowledge surrounding MPs, we leveraged the amphibian Xenopus laevis as a robust comparative model. The X. laevis Research Resource for Immunology at the University is specialized in the development and use of Xenopus for immunological research. Fully aquatic tadpoles are ideal experimental organisms for addressing the acute and persistent biological impacts resulting from exposure to MPs, because their post-embryonic development, including the immune system differentiation is external and not protected by the maternal environment, which increases their sensitivity to perturbations by water pollutants. Furthermore, the development and physiology, as well as the immune system of Xenopus, are remarkably similar to that of humans which has led to fundamental discoveries about pathophysiology, development, and medical immunology [7-9].
Lake Ontario MicroPlastics Center
The research program using Xenopus is integrated into the Lake Ontario MicroPlastics Center (LOMP). This is a new collaborative initiative between the University of Rochester (UR) and the Rochester Institute of Technology (RIT). LOMP is a hub for research, translation, and community engagement, interested in investigating how different types of real-life plastics enter and move through the Great Lakes ecosystems and how MPs may affect human health under different environmental conditions. LOMP is one of six Centers for Oceans and Human Health jointly funded by the National Science Foundation and the National Institute of Environmental Health Sciences. As mentioned above, besides marine ecosystems, significant quantities of MPs have been detected in Upstate New York lakes, rivers, and the drinking water of cities such as Rochester, NY [10]. This has led to the establishment of this productive collaborative research program between UR and RIT. An important component of LOMP is its Materials and Metrology Core that develops standardized protocols and produces optimized materials for the different research teams. For example, the Core provides silicon nanomembranes used for water and air filtration to quantify environmental MPs; it also prepares by cryomilling lab-made MP stock solutions in defined size ranges to mimic real-life MPs (Fig. 1A-C).
Figure 1: Polyethylene terephthalate (PET) MPs. (A) Bright field image and (B) RFP image obtained with an epifluorescence microscope of real-life PET-MPs of variable shapes and sizes (1-20 μm) stained with fluorescent Nile red dye. (C) Photograph of silicone nanomembranes (SimPore, Inc.). (D) Bright field image, and (E) RFP image of PET-MPs isolated on a nanomembrane (2 μm × 50 μm microslits) from liver lysates of tadpoles exposed for 7 days.
Biodistribution and biological impact of MP exposure in water on Xenopus
As part of the LOMP program, the objectives of the Xenopus research project are to determine the biodistribution and biological impacts of MP water contaminants using a sensitive and reliable experimental platform in X. laevis. The overarching hypothesis is that the developmental exposure to MPs will induce composition and size dependent long-term perturbations of immune homeostasis, chronic inflammation, decreased resistance to microbial pathogens and poorer antimicrobial immunity.
Using X. laevis tadpoles, we are conducting a rigorous assessment of the biodistribution and biological effects of MP ingestion, and specifically their potential to affect immune homeostasis and weaken the immune response to viral pathogens. In contrast to studies often using unrealistically large amounts of spherical microspheres (greater than 1 g/L), we are using smaller amounts of MPs (from 25 to 0.1 mg/L) that are closer to what is found in the environment. As a point of comparison, it is estimated that mineral water bottle can contain up to 0.6 mg/L of MPs [11], while European drinking water has been reported to contain 4,889 MPs per liter, which would correspond to about 2.6 mg/L [12]. We are focusing on two types of environmentally relevant plastics: polyethylene terephthalate (PET), which is extensively used in the packaging industry, and is a significant contributor to environmental plastic pollution [13] but is under-investigated [14]; and nylon 66 (nylon), which despite being one of the most abundant MPs in the microenvironments and detected in high amount in human tissues [5], there is little research about its biological impact. In collaboration with the other LOMP research teams, we also plan to test MPs isolated from Lake Ontario.
We first defined in detail the biodistribution, accumulation, and persistence of PET-MPs cryomilled to different size ranges: 1–100 μm [15] and 1-20 μm. These MPs were fluorescently labelled with Nile red for their detection by fluorescence microscopy on whole mount tissues and isolated peritoneal macrophages. The biodistribution was also evaluated by enzymatic digestion and silicon nanomembrane filtration (Fig. 1D and E). Even at concentrations as low as 0.1 mg/L, there was a rapid intestinal transit of PET-MPs leading to their accumulation as early as 24 hrs. of exposure in tadpole intestines, liver, kidneys, brain, and peritoneal macrophages, where they persisted for over a week after the initial exposure. We were able to estimate that a 2–3-week-old tadpole weighing approximatively 300 grams could ingest up to 2 mg of PET-MPs during the 24-hr. exposure. When transferring exposed tadpoles into clean MP-free water, a total of 1.7 mg of the MPs were released within 7 days. Thus, we estimated that on average 0.3 grams of MPs were retained in tadpoles one week after exposure, which correspond to 0.5-1 mg of MPs per gram of tadpole tissue.
To determine whether exposure to PET-MP has any effect on tadpole immunity, we took advantage of the ranavirus FV3, a major amphibian pathogen, which is a large double-stranded DNA virus. We have extensively characterized the pathogenesis and immune response against FV3 in X. laevis (reviewed in [16]). Notably, exposure to PET-MPs at a concentration of 10 mg/L for 1 month significantly increased tadpoles susceptibility to viral infection and altered innate antiviral immunity without inducing overt inflammation [15]. Further analysis of gene expression by qPCR revealed an altered expression of several genes critical for macrophage function (e.g., IL-34, MHC-II), which suggest that exposure to PET-MPs induces some macrophage dysfunction. Regarding the effect on development, we also noted that 1 month of exposure to PET-MP significantly delayed metamorphosis completion.
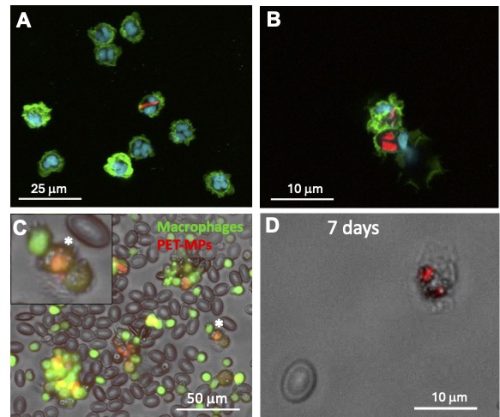
We are now focusing our investigation on the effects of MPs on macrophage function using both in vitro and in vivoapproaches. Interestingly, our preliminary results indicate that, compared to polystyrene or polyethylene manufactured spherical beads that are phagocytosed by a majority (>90%) of peritoneal macrophages from adult frogs at the same concentration, PET, and nylon 1-20 μm MPs fragments are internalized by only a minor fraction (~10%) of peritoneal macrophages. In addition, fewer PET and nylon MPs are internalized by individual peritoneal macrophage compared to manufactured spherical beads (Fig. 2A and B). We are in the process of developing a controlled aging process by UV treatment to determine if aged MPs might be phagocyted at a different rate than pristine MPs. To assess MP effect on macrophage function in vivo, we are taking advantage of the X. laevis transgenic line mpeg::GFP, where macrophages express the fluorescent GFP reporter. Intraperitoneal injection of a small amount of Nile red-stained PET-MPs in adult frogs allows the detection of GFP+ macrophages that have engulfed red fluorescent PET-MPs (Fig. 2C). Similarly, tadpole peritoneal macrophages can phagocytose red fluorescent PET-MPs following intraperitoneal injection. Moreover, we can detect macrophages with internalized PET-MPs up to 7 days after exposure (Fig. 2D), which suggests that, as also observed in vitro, MP engulfment does not induce marked cell death. This in vivo system will now allow us to follow the fate of macrophages with internalized MPs and determine whether their function in homeostasis, regeneration and immune response is altered.
Figure 2. In vitro and in vivo MP internalization assay. (A) Confocal microscopy images of adult X. laevis peritoneal macrophages (Mø) incubated with 10 μg/mL of Nile red fluorescently stained PET-MPs (1-20 μm) for 24 hours in vitro. (B) Similar peritoneal macrophages incubated with 10 μg/mL of Nile red fluorescently stained nylon-MPs (1-20 μm) for 24 hours in vitro. (C) Three days following the elicitation of Mø in the peritoneum, mpeg::gfp transgenic frogs with green fluorescent Mø were intraperitoneally injected with 10 μg of Nile red fluorescently stained PET-MPs. PLs were harvested 24 hrs. later by lavage and Mø with internalized MPs (orange) were visualized under a fluorescent microscope. (*) Magnification of a Mø with ingested PET-MPs. (D) Peritoneal macrophage from a tadpole (3 weeks of age) 7 days post-intraperitoneal injection with 10 μg Nile red fluorescently stained PET-MPs.
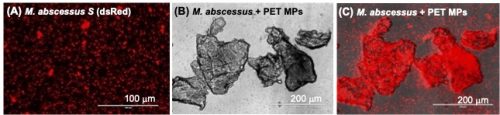
Regarding association of microorganisms with MPs, mycobacteria spp. were found in biofilms generated on plastic debris in a field mesocosm study in Lake Ontario and in the laboratory setting by our LOMP collaborators at RIT [6]. To further investigate the potential of MPs to interact with pathogens, we incubated PET-MPs with different mycobacteria species. M. marinum, a non-tuberculosis mycobacterium found in water around the world, can tightly bind to PET-MPs in vitro, as well as M. abscessus, which is a notable emerging human pathogen (Fig. 3). We plan to determine whether association of mycobacteria with MPs promotes their colonization in tadpoles and whether it affect infection and survival in macrophages.
Fig. 3: Tight association of non-tuberculous Mycobacterium abscessus with PET-MPs. M. abscessus expressing dsRED fluorescent reporter incubated overnight at 1×105 cfu in (A) 1 ml of amphibian PBS or (B, C) 10 µg of PET-MPs in 1 ml of amphibian PBS. The mixture was gently resuspended before examination under a fluorescent microscope.
In summary, our study using Xenopus carries substantial significance, raising developmental immunotoxicity (DIT) concerns not only for aquatic vertebrates but also for human health. The demonstrated impact on immune function underscores the broader ramifications of MP pollution, highlighting the need for comprehensive strategies to mitigate its pervasive DIT effects on both aquatic ecosystems and human populations.
Acknowledgements
The expert animal husbandry provided by Tina Martin is gratefully appreciated. We would like to thank Rachel F. Lombardo, Francisco De Jesus Andino and Hannah Turner for their significant experiment contribution as well as Drs. Lisa De Louise and Alison Elder for their critical review of this manuscript. The Lake Ontario MicroPlastics Center (LOMP) is jointly funded by NIEHS (P01 ES035526) and NSF (OCE-2418255). JR is also funded by NAID (R24-AI059830) and R.L. by the Toxicology Training Grant (T32-ES07026).
This video is the culmination of several years attempting to: (1) Figure out best practices for modeling protein-protein interactions; (2) Understand the outputs of programs like AlphaFold and adjacent software including quantitative metrics; and (3) Communicate my thoughts to unwitting victims through workshops. Hopefully, others like me (molecular biologists, geneticists, cell and developmental biologists) may find some value in the content.
My lab group is interested in understanding the genetic and developmental mechanisms behind the evolution of novel phenotypes (evo-devo). I am seeking a curious student interested in biomineralization, evolution, development, and/or molluscs to join my team. We are aiming to understand how the shell of molluscs has evolved into so many different forms by understanding the developmental mechanisms regulating biomineralization via the mantle.
We are a new lab helping to develop a slipper snail model (Crepidula atrasolea) to test fundamental evolution and development questions. As a member of our lab, you will have the freedom to tailor your project to your interests, ideally relating to the genetics, development, or role of the nervous system in shell formation in gastropods. Depending on your project and research goals, you will have the opportunity to learn microinjection, CRISPR, hybridization chain reaction in situ hybridization, immunohistochemistry, confocal microscopy, animal husbandry, electron microscopy, and more.
We are a welcoming group that celebrates diverse backgrounds and strives for an inclusive environment for biologists of any background, age, race/ethnicity, sexual orientation, gender, or dis/ability.




 (4 votes)
(4 votes)


 (No Ratings Yet)
(No Ratings Yet)