As scientists, we are all are works in progress and continually developing in our own ways. Each of us brings unique strengths and skills along with challenges, and these can change during the course of our careers. Still, there are some common themes to what makes for an effective scientist, and these can provide useful benchmarks for evaluating our progress and planning the path ahead. This article on helpimascientist.com defines several stages of scientific development, with the intention to promote honest self reflection, open conversations, and aid in helping to set goals.
Here we showcase work from Craig Zuckerman, a digital fine artist whose work draws on scientific imagery to create immersive visual environments. With a background in medical illustration and animation, he now works at the intersection of science and fine art, using digital tools to explore form, light, and colour in ways that invite reflection and quiet attention.
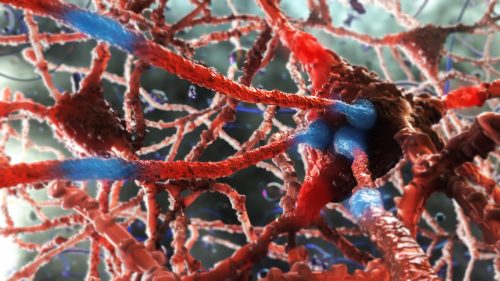
Dynamic surface: lipid membrane surface proteins
Can you describe your artistic practice and how science informs it?
My work transforms cellular and subcellular structures into generative frameworks, moving beyond scientific description into poetic abstraction. Microscopy becomes a language for exploring form, colour, and spatial complexity. Each piece creates a tension between what is biologically recognisable and what is purely atmospheric.
I use scientific structures as scaffolding and use them for creating cinematic environments that resist literal interpretation. My digital work becomes a place to sculpt form, colour, and space with precision, constructing environments that feel immersive and contemplative. Microscopic systems are expanded into vast, navigable landscapes. This shift in scale invites viewers to inhabit the unseen, reframing the body’s interior as a place of wonder, serenity, and emotional resonance.
Luminous currents: daughter cell budding off of host cell
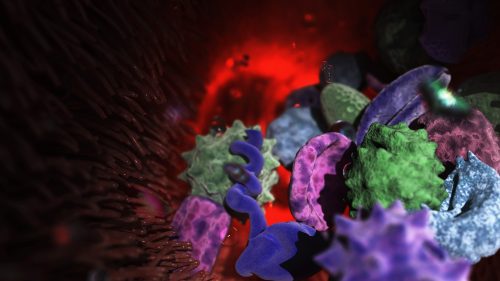
Structures in transition: organic cell demise
How do light, colour, and materiality function in your work?
Light functions as a structural force, creating depth and atmosphere. Colour becomes a psychological and emotional driver, guiding the viewer’s experience and transforming biological forms into meditative spaces. These environments invite contemplative gazing. They create a sense of inwardness, mirroring the quiet intelligence of living systems and offering viewers a space for reflection, grounding, and calm.
My limited edition prints on aluminium and plexiglass emphasise physicality, durability, and concreteness. Editioning becomes a conceptual gesture, establishing boundaries around reproducibility and reinforcing the singularity of each work.
Synaptic pulse: neurotransmitters traveling along neuronal axons
Where does your inspiration come from, and how has it informed your artistic practice?
I am a digital fine artist whose work is inspired by science, with a background in medical illustration and animation. My practice marks a deliberate shift from applied science visualisation to autonomous fine art, using cellular structures as generative frameworks rather than clinical subjects.
Now working exclusively in the digital discipline, I construct immersive, cinematic environments in various 3D software that occupy the intersection of abstraction and representation. Colour, light, and composition function as primary structural elements, transforming microscopic systems into expansive spatial experiences. I now use science as a starting point to create biolandscapes.
I have always been influenced by prominent illustrators, from the golden age of illustration through the 1980s, as well as artists from the Renaissance, landscape artists, and sculptors. I continue to create more work in this space, constantly challenging myself with respect to technique, colour, composition, and scientific knowledge.
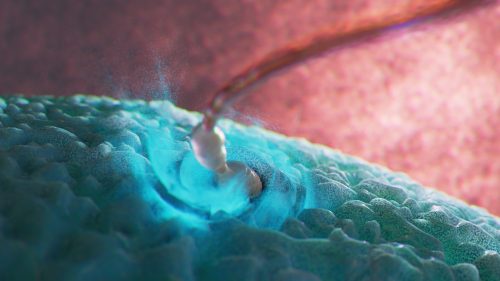
Conception: sperm cell penetrating egg cell
What advice would you give to others interested in your SciArt approach and where can they find more of your work?
To anyone who has interest in pursuing this approach, it is most important to grow as a visual artist — i.e. use of colour, composition, lighting, drawing and painting skills, or in the software of your choice.
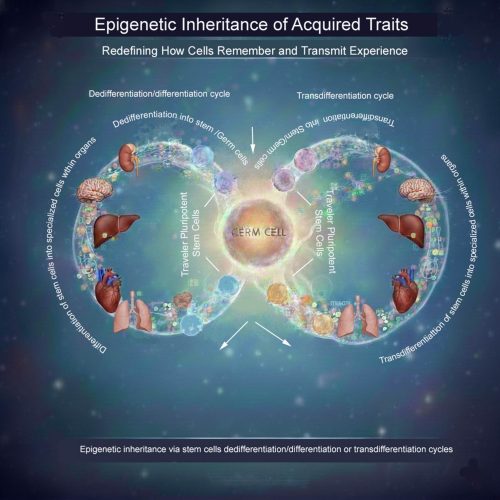
Do cells carry memories of the whole body into the next generation?
For more than two millennia, biologists and philosophers have debated whether traits acquired during life can be inherited. From Hippocrates and Aristotle to Lamarck and Darwin, this idea repeatedly surfaced but was ultimately set aside due to the absence of a convincing biological mechanism.
Recent advances in epigenetics have reopened this question.
In a recent paper, DOI: 10.1016/j.cdev.2024.203928 I propose a conceptual framework for how environmentally induced epigenetic information might be transmitted from somatic tissues to germ cells—not solely through diffusible molecules, but through cellular movement combined with fate plasticity.
A traveler stem cell hypothesis
The central idea is that certain pluripotent or highly plastic adult stem cells—potentially including germline-associated stem cells—may act as epigenetic travelers. These cells could circulate through the body, enter developmentally active or regenerating tissues, and undergo cycles of differentiation or transdifferentiation in response to local cues. During these transitions, they would acquire tissue-specific epigenetic modifications.
Importantly, these cells would not remain terminally committed. Through dedifferentiation or further transdifferentiation, they could revert to an uncommitted state while retaining accumulated epigenetic information. During gametogenesis, such cells might be recruited back to the gonads, where they ultimately contribute to germ cells—carrying with them epigenetic memory collected across multiple somatic environments.
What the image illustrates
The accompanying schematic visualizes this concept: pluripotent “traveler” stem cells move between tissues, repeatedly cycling through differentiation, dedifferentiation, and transdifferentiation. Over time, they integrate epigenetic inputs from diverse organs before re-entering the germline, offering a potential cellular route for soma-to-germline information transfer.
Existing biological foundations
Crucially, elements of this process are not purely hypothetical. Across many multicellular organisms—including plants, invertebrates, and vertebrates—intergenerational and transgenerational epigenetic inheritance has already been experimentally demonstrated. Epigenetic information can persist through extensive developmental reprogramming events and across multiple generations, indicating that biological systems possess robust mechanisms for preserving epigenetic memory.
Moreover, it has been shown that germ cells or germline-associated stem cells are not irreversibly restricted to reproductive fate. Under specific developmental or experimental conditions, germ cells have been observed to generate diverse somatic cell types. Conversely, somatic or pluripotent stem cells can be induced to acquire germ cell identity and contribute to functional gametes. These bidirectional fate transitions challenge a strict interpretation of the soma–germline barrier and establish that germline and somatic identities are more plastic than traditionally assumed.
Together, these observations provide a biological foundation for considering mobile, fate-plastic cells as integrators and carriers of epigenetic information across tissues.
Why this matters
This framework does not contradict existing models of epigenetic inheritance involving small RNAs or other molecular mediators. Instead, it complements them by addressing a key unresolved problem: how complex, tissue-specific epigenetic states accumulated across an organism’s lifetime might be integrated and transmitted coherently to the next generation.
If experimentally validated, this idea could have implications for developmental biology, evolution, aging, regenerative medicine, and disease inheritance.
A question for the community
If highly plastic stem cells can act as mobile carriers of epigenetic memory, how might we experimentally trace their movements, fate transitions, and epigenetic histories across tissues and generations?
I would welcome thoughts on experimental strategies—or alternative interpretations—that could test or challenge this hypothesis.
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Recognizing dUTPase as a mitotic factor essential for early embryonic development Nikolett Nagy, Otília Tóth, Eszter Oláh, László Henn, Gergely Attila Rácz, Edit Szabó, György Várady, Fanni Beatrix Vigh, Zita Réka Golács, Martin Urbán, Tímea Pintér, Orsolya Ivett Hoffmann, László Hiripi, Hilde Loge Nilsen, Angéla Békési, Miklós Erdélyi, Elen Gócza, Gergely Róna, Judit Tóth, Beáta G. Vértessy
Single-cell spatially resolved transcriptomic characterization of the developing mouse cochlea Philippe Jean, Sabrina Mechaussier, Amrit Singh-Estivalet, Céline Trébeau, Aurore Gaudin, Laura Barrio Cano, Andrea Lelli, Fabienne Wong Jun Tai, Sébastien Megharba, Sandrine Schmutz, Sarra Loulizi, Sophie Novault, David Hardy, Carolina Moraes-Cabe, Milena Hasan, Christine Petit, Raphael Etournay, Nicolas Michalski
Scalable high-fidelity human vascularized cortical assembloids recapitulate neurovascular co-development and cell specialization Shubhang Bhalla, Belda Gulsuyu, Damian Sanchez, Jayden M. Ross, Santhosh Arul, Adnan Gopinadhan, Muhammet Öztürk, Tanzila Mukhtar, Jonathan J. Augustin, Jerry C. Wang, Joseph Kim, Chang N. Kim, Sena Oten, Yohei Rosen, John M. Bernabei, Vijay Letchuman, Shantel Weinsheimer, Helen Kim, Elizabeth E. Crouch, Edward F. Chang, David Haussler, Mircea Teodorescu, Arnold R. Kriegstein, Tomasz J. Nowakowski, Ethan A. Winkler
Novel repressors of cambium activity in Arabidopsis Xing Wang, Jingyi Han, Emma K. Turley, Riikka Mäkilä, Anne-Maarit Bågman, Julia M. Kraus, Qing He, Hanan Alhowty, Joanna Edwards, Yuqi Li, Raluca Blasciuc, Wiktoria Fatz, Wenbin Wei, Miguel de Lucas, Siobhán M. Brady, Shixue Zheng, Chunli Chen, Ari Pekka Mäh-önen, J. Peter Etchells
In vitro sexual dimorphism establishment in schistosomes Remi Pichon, Magda E Lotkowska, Jude L. D. Bulathsinghalage, Madeleine McMath, Mary Evans, Benjamin J. Hulme, Kirsty Ambridge, Geetha Sankaranarayanan, Simon Kershenbaum, Sarah D. Davey, Josephine E. Forde-Thomas, Karl F. Hoffmann, Matthew Berriman, Gabriel Rinaldi
Leveraging a hybrid cross-disciplinary training model to accelerate global bioinformatics capacity Taras K. Oleksyk, Daryna Yakymenko, Sylwia Bożek, Viorel Munteanu, Wojciech Pilch, Zoia Comarova, Victor Gordeev, Grigore Boldirev, Dumitru Ciorbă, Viorel Bostan, Christopher E. Mason, Alexander G. Lucaci, Nadiia Kasianchuk, Daria Nishchenko, Victoria Popic, Andrei Lobiuc, Mihai Covasa, Martin Hölzer, Joanna Polanska, Alex Zelikovsky, Vasili Braga, Mihai Dimian, Paweł Łabaj, Serghei Mangul
Cloud-Connected Pluripotent Stem Cell Platform Enhances Scientific Identity in Underrepresented Students Samira Vera-Choqqueccota, Drew Ehrlich, Vladimir Luna-Gomez, Sebastian Hernandez, Jesus Gonzalez-Ferrer, Hunter E. Schweiger, Kateryna Voitiuk, Yohei Rosen, Kivilcim Doganyigit, Isabel Cline, Rebecca Ward, Erika Yeh, Karen H. Miga, Barbara Des Rochers, Sri Kurniawan, David Haussler, Kristian López Vargas, Mircea Teodorescu, Mohammed A. Mostajo-Radji
OurJanuary webinar featured two early-career researchers studying development, evolution and the environment. Here, we share the talks from Chee Kiang (Ethan) Ewe (Tel Aviv University) and Max Farnworth (University of Bristol).
If you are in the pancreas field, you may be either part of the endocrine or the exocrine band. Now, this may not be like the Sharks and the Jets in West Side Story, but you better know your position. Whether this separation reflects the actual spatial segregation of both compartments and their different embryonic development is an idea perhaps worth exploring. In any case, our question was linked precisely to that spatial segregation: why do both compartments develop in different regions of the organ?
First of all, we know that what lies around the pancreatic epithelium (what we call the microenvironment) is crucial for its development. Since the 60s1, great works have progressively characterised the microenvironment with greater and greater detail, from early elegant experiments using explants, to more elaborate mouse genetics studies where specific cellular components and signalling pathways were perturbed2,3. The single-cell revolution brought a new twist: the degree of cellular heterogeneity populating the microenvironment, especially mesenchymal cells, was much higher than anticipated. The question then was: how is this heterogeneity spatially distributed?
Mapping the pancreas and deciphering maps.
Spatial transcriptomics (ST) appeared to us the best way to answer the question, but at the time we started the project, sequencing-based approaches did not provide the resolution needed to map a small, branched organ like the embryonic pancreas. On the other hand, image-based approaches only allowed for mapping the expression of a handful of markers. Thanks to the early discussions Francesca Spagnoli (PI of the lab) had with Cartana, the biotech at Karolinska Institute, which developed the In Situ Sequencing (ISS) technology and was later acquired by 10x Genomics, we were able to pioneer this approach. In parallel, access to the first single-cell RNASeq datasets of the murine embryonic pancreas -from our lab and others in the field4– enabled us to identify the most informative set of marker genes and design robust panels for the ISS experiments. Running the ISS technology on pancreas was not immediately immediately straightforward; it required considerable effort and a series of optimization experiments carried out by me and another postdoc in the lab., Jean Francois Darrigrand. Finally, by profiling the spatial distribution of sets of markers, we were able to create a cartography of the mouse embryonic pancreas (Fig. 1).
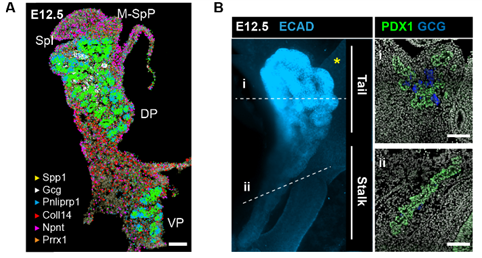
Fig. 1 ISS image of selected marker genes in E17.5 pancreas. Close-ups of selected probe genes and their spatial distribution in the tissue are shown in (i) and (ii) dashed boxes.
But a map is only an instrument, and the information obtained from it will largely depend on how you read it. When analysing a geographical map, your answers may vary depending on the level of aggregation: you can look at it from the country perspective, zoom in and separate by region or zoom in even more and analyse every city and small town independently. Similarly, when observing an organ, one can use different magnification lenses. First, the pancreas originates from two groups of progenitor cells growing independently (dorsal and ventral pancreatic buds), until they fuse around E14.5 in the mouse embryo. As shown in the 3D images below, generated by a PhD student in the lab, Anna Salowka, the architecture of each bud is not homogeneous along its axes. At the organ level, we discovered that the mesenchyme surrounding the ventral and dorsal pancreas is distinct (Fig. 2A). Furthermore, along the dorsal pancreas -from the duodenum to the region next to the spleen- specific mesenchyme subsets are selectively enriched (Fig. 2B).
Fig. 2 (A) Representative ISS image showing selected genes in dorsal pancreas (DP) and ventral pancreas (VP) at E12.5. Scale bar, 100 μm. (B) Representative three-dimensional (3D) rendering of light-sheet fluorescent microscopy image (left) and confocal microscopy images (right) of E12.5 pancreas stained with indicated antibodies. Right: Confocal IF images show transverse cryosections of DP at tail (i) and stalk (ii) levels. Hoechst was used as nuclear counterstain. Scale bars, 100 μm. Asterisk indicates approximate position of the spleen.
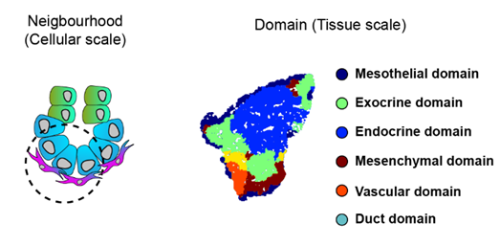
To increase the resolution of our analysis to meso- and micro- scales (Fig. 3), Gabriel Herrera (at the time rotation student in the lab) brough into the project his bioinformatic skills to implement pipelines to analyse the spatial data. What we found is that the tissue is organised in concentrical niches enriched in mesothelial, mesenchymal, exocrine or endocrine cells. When comparing exocrine and endocrine niches, we found that proliferative mesenchyme was preferentially located around acinar cells, whereas another subset, which we termed Mesenchyme (M)-II, was enriched in the endocrine niche.
Fig. 3 Schematics of the spatial analysis frameworks: At cellular scale (left), spatial neighborhoods encompassing the 10 closest cells around each cell were used to calculate cluster pair neighborhood enrichment; at tissue scale (right), tissue areas with similar local cell type composition were clustered to identify tissue domains.
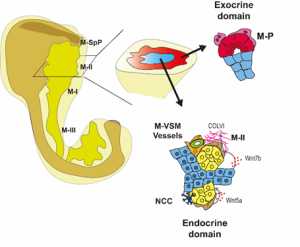
We then focused on the latter association and identified putative Ligand:Receptor interactions between M-II and endocrine cells (Fig. 4). In particular, Wnt5a and Collagen VI molecules caught our attention because of their potential role in creating a niche favourable for endocrine and, specifically, beta-cell differentiation. Consistently, functional experiments using mouse pancreatic explants demonstrated that blocking Wnt5a signaling hampered endocrinogenesis by perturbing the JNK pathway. On the other hand, explants treated with Collagen VI showed a higher number of endocrine cells. By examining human foetal pancreatic tissue, Georgina Goss, a postdoc in the lab, showed that Collagen VI is also enriched around human endocrine cells. Finally, I went on embedding human iPSC-derived endocrine cells in hydrogels containing different ECM mixes, and discovered that Collagen VI, in a conserved fashion, increased the number of beta-cells in the cultures.
To complete our study, we decided to have a glimpse of the adult pancreas. What we found is that different mesenchyme subsets are enriched inside and around islets of Langerhans, ducts and acini. A long-standing question in the field is to track the origin of the adult pancreatic mesenchyme. Our dataset enabled us to fill this gap. Using in silico analysis, we identified fate trajectories connecting the embryonic and adult mesenchyme. Our results suggested that the Spleno-Pancreatic mesenchyme could be one of the origins of the adult mesenchyme which we confirmed using in vivo lineage tracing.
Fig 4: Spatial organization of the pancreatic mesenchyme during embryonic development
What’s next?
Several questions remain open, and several arose during the project. If the pancreatic tissue is carefully distributed, how is that architecture shaped? What signals link epithelial compartments to the formation of their surrounding microenvironment? Our results also raise questions regarding the function of the different levels of organisation: Why does pancreas development need gradients of signalling along the proximodistal axis? It would be interesting to test whether the disruption of that axis causes defects in the separation of the pancreas and surrounding organs. Further research is also needed to understand the function of the secretion of specific ECM components, such as Collagen VI, around exocrine and endocrine cells. In the case of Collagen VI, it would be interesting to investigate how it affects tissue stiffness, as it has been shown that control of the mechanotransducer YAP is crucial for endocrinogenesis. Finally, the spatial organization of the microenvironment during human embryonic development needs further characterization, but using similar approaches we are now beginning to understand it, so if you want to know a little bit more about it, check out the new preprint from the lab5.
Access the article: Torres-Cano, A., Darrigrand, J. F., Herrera-Oropeza, G., Goss, G., Willnow, D., Salowka, A., Ma, S., Chitnis, D., Rouault, M., Vigilante, A., & Spagnoli, F. M. (2025). Spatially organized cellular communities shape functional tissue architecture in the pancreas. Sci Adv, 11(46), eadx5791. https://doi.org/10.1126/sciadv.adx5791
References
1. Golosow, N. & Grobstein, C. Epitheliomesenchymal interaction in pancreatic morphogenesis. Developmental Biology4, doi:10.1016/0012-1606(62)90042-8 (1962/04/01).
2. L, L. et al. Pancreatic mesenchyme regulates epithelial organogenesis throughout development – PubMed. PLoS biology9, doi:10.1371/journal.pbio.1001143 (2011 Sep).
3. C, C. et al. A Specialized Niche in the Pancreatic Microenvironment Promotes Endocrine Differentiation – PubMed. Developmental cell55, doi:10.1016/j.devcel.2020.08.003 (10/26/2020).
4. Byrnes, L. E. et al. Lineage dynamics of murine pancreatic development at single-cell resolution. Nature Communications 2018 9:19, doi:10.1038/s41467-018-06176-3 (2018-09-25).
5. Goss, G. et al. Mesodermal-niche interactions direct specification and differentiation of pancreatic islet cells in human multilineage organoids. bioRxiv, 2025.2012.2013.694117, doi:10.64898/2025.12.13.694117 (2025).
This is part of the ‘Lab meeting’ series featuring developmental and stem cell biology labs around the world.
Where is the lab?
Andrea: You can find the Ditadi lab at Ospedale San Raffaele, as part of the San Raffaele Telethon Institute for Gene Therapy, in the north-east corner of Milan, Italy. Milan is a great spot for both science and life, with a myriad of places to visit, plenty of things to do and a rich community of great labs to collaborate with.
Andrea: We want to understand how human blood cells form. For this, we use human pluripotent stem cells as a model, integrating developmental, cell and molecular biology, as well as a bit of immunology. We study human developmental biology in a dish: we study early mesoderm patterning and follow the process all the way to mature blood cells, including hematopoietic stem cells, trying to work out which signals guide each step. We are developmental biologists working at an institute that focuses on genetic diseases and their therapy, so we also use the cells we generate to look at diseases from a developmental perspective. At the same time, we explore how to engineer and arm these cells in unique ways so they can be used in clinical settings in the future.
Lab roll call
Let’s start in order of length of service in the lab.
We have Lauren Randolph, post-doctoral fellow, who is studying how hemogenic cells give rise to blood.
Claudia Castiglioni, PhD student, who aims to identify the earliest commitment to blood cell fate.
Riccardo Piussi, former Master’s student now PhD student-wannabe (and hopefully soon-to-be), working to decipher the regulation of self-renewal in emerging HSCs.
Deborah Donzel, a postdoctoral fellow, and Nikita Pinto, another former Master’s student now turned research assistant, are partners in modeling a ribosomopathy that affects red blood cells only postnatally to decipher proteostasis regulation across different stages of hematopoietic development.
Elena Morganti, a postdoctoral fellow, and Bianca Nesti, a Master’s student, who teamed up to model a pediatric autoimmune disease as a way to understand the role of embryonic lymphocytes in health and disease.
Alessandra Guerreschi, a Master’s student who recently joined our lab and is gearing up to investigate the multiple roles of Notch signaling in hematopoietic development.
Favourite technique, and why?
Andrea: It is not exactly a technique, but my favourite moment in the lab is simply watching cells under the microscope. We do not do much imaging; most of our days are spent in the hood doing cell culture. Even now, when I am sadly not doing many experiments any longer, I still have this habit that I actually stole from my postdoc advisor. When I need a break from the desk and the administrative tasks, I go to the lab for what I call a bit of “cell therapy”. I grab a few plates and spend some time simply looking at cells under the microscope. I love it. Observing cells in cultures is very informative, cells talk to us all the time.
If I need to choose a proper technique, I would choose flow cytometry. We use it a lot. It may not be as high-throughput as some newer methods, but it gives us robust full gene expression data at the single-cell level, and we can learn a lot from what comes out of the cytometer.
Andrea, apart from your own research, what are you most excited about in developmental and stem cell biology?
Andrea: Recently, I have been following the evo-devo field with a lot of interest. I find it fascinating to think about how cells and tissues evolved, and for a lab like ours that tries to recreate how blood cells are formed in vitro, understanding how they appeared in the first place feels very relevant.
Another field that I find extremely exciting is synthetic biology. I am fascinated by how we can now “prod” cells and systems and modify their responses. I remain a developmental biologist at heart, but the environment where we work has opened my eyes to how we can push the boundaries of therapeutic innovation. Alongside the clinical application of stem cells, synthetic biology is transforming the way we think about medicine and how we might design future therapies.
Andrea, how do you approach managing your group and all the different tasks required in your job?
Andrea: I am not sure I can say I am set into one approach, at least yet. I think it is always evolving, as the people in the lab, as well as the lab itself, need change over time. In general, I try to spend time getting to know the people in my group, recognizing the strengths and weaknesses, and trying to exploit the former while helping them work on the latter. I often think of the group as an orchestra or a music band. First, I need to hear the sound of each instrument, help them get tuned and then my job is to compose some music that fits them. Let’s say that some composition takes more time than others. But in the end, the goal is to nurture the love and passion for the true privilege of doing research for everyone.
As for managing the different tasks, I often wish I had more hours in the day; that would be a great superpower. So, I try to clear out the things I do not enjoy, the administrative duties and emails, as quickly as possible. This gives me protected time for what I love: reading, thinking and spending time with the team in the lab. I am not sure I always find enough time for that, but I try very hard.
What is the best thing about where you work?
AD: Without a doubt, being surrounded by young and bright people. It is energizing and another privilege of this job.
CC: The thing I value most about being at SR-Tiget is the stimulating environment, where science truly comes alive. Ideas are shared freely, we have the resources to bring them to life, and we constantly get to learn from seminars by scientists from around the world.
NP: The best thing about working at SR-Tiget in Milan is the combination of different scientific topics and a truly collaborative environment, where you can walk into a lab or an office to ask for help and know that someone will genuinely take the time to help you solve a problem.
RP: What I like most about where I work is the general drive of the institute to do high-level science and to set ambitious goals. In the lab, I really appreciate the way we reason scientifically and the fact that we constantly challenge our ideas by asking questions every day.
LNR: The best thing about where I work is the science and the people. I really enjoy the project that I am working on and find it both challenging and engaging. I am also really lucky to work with incredibly collaborative and supportive colleagues who really treat the lab as a family. It makes it a joy to spend time with them, both in and out of work, and to do and talk science together.
DD: The best things about where I work are the research topic and the people I work with. My enthusiasm for the project keeps me focused and driven, even during challenging periods. I’m also fortunate to work with colleagues who are open to sharing ideas and knowledge, which creates a collaborative environment that helps us move forward together.
BN: What I appreciate most about working at SR-Tiget is the highly stimulating scientific environment, both at the institute level and within my own laboratory. The presence of diverse expertise, frequent seminars and strong resources fosters a continuous exchange of ideas and supports high-quality research.
EM: What I like most about where I work is the young and supportive environment. I feel that people around me are genuine and open-minded, and this makes my days very pleasant and enjoyable.
AG: Even though I haven’t been here long, I’ve really noticed how welcoming and supportive everyone is. It makes it easy to ask questions, learn quickly and feel like part of the team right away.
What’s there to do outside of the lab?
AD: Despite being in love with my job and not feeling the need to escape, life is too short, and I have so many interests – books, music, sport, hiking, biking, food, friends, etc. – so I try to do a bit of it all. To be coordinated with the family, in particular, two kids who keep me happy and busy.
CC: Outside of the lab, I really enjoy canoeing on the Navigli, the famous canals in Milan. Being on the water allows me to slow down and take a break from the busy pace of the lab. I love the feeling of paddling along the canals, enjoying the surroundings and reconnecting with the city.
NP: Having grown up in Milan, I sincerely love this city and everything it offers. Outside of the lab, I like different things, from baking and crocheting to spending time with family and friends while enjoying the city’s cultural life, like its aperitivo culture and different neighborhoods. Recently, I also joined the Red Cross as a volunteer, where I am involved in social inclusion activities with homeless people, as well as assistance roles during public events. These experiences help me stay grounded, connected to the community and maintain balance alongside research.
RP: This job takes a lot of time and energy, but outside of the lab I really enjoy spending time with my family and friends. I also love fishing. I enjoy it for its unpredictability and complexity; it requires analyzing many variables and accepting failure without expectations. Every small decision can make a difference, and while nothing is guaranteed, everything is possible, like in science.
LNR: Outside of the lab, I enjoy traveling, reading, and all things food-related. In Milan, I particularly enjoy access to the ballet, opera, and theater.
DD: Outside of the lab, I really enjoy going for walks—especially in parks or outside the city, where I can reconnect with nature. Living in Milan, I also like going to the theater and meeting friends for an aperitivo.
BN: Outside the lab, I enjoy spending time reading, as it offers a break from continuous scientific reflection while still keeping my mind engaged in a pleasant way. I also like to take advantage of the many cultural and recreational initiatives that Milan has to offer, often in the company of my friends.
EM: I usually try to spend time in nature and clean air when I am not in the lab. Milano is really close to beautiful mountains and lake,s and those are my favorite spots for the weekend. I also enjoy food, art and history.
AG: In my free time, I enjoy reading and spending time in the mountains outside of Milan, whether it’s hiking, skiing or horseback riding. Skiing, in particular, is a great way to unwind on the weekends and enjoy the outdoors. Being able to combine outdoor activities with some quiet time to read makes my free time really enjoyable.
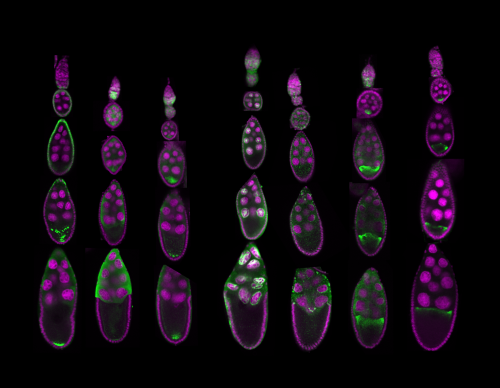
Biology probes form and function of Life. Form is easy to grasp: cells under a microscope, subcellular structures in electron micrographs, or organisms on camera readily present their shapes.
Figure 1: Examples of microscopy images that allow understanding of Life: Fruit fly ovaries development and the changing RNA localizations during the process. Jambor, 2015
Function is different: it emerges from molecular compositions, interactions, and temporal changes. Such data is not directly visible – we use statistics to make sense of it. But summaries and p-values alone rarely reveal how complex biological systems are organized, the variability in the samples and the resulting uncertainty in the data, or unexpected relationships and pattersn. As datasets grow larger and more complex, these insights only become accessible when data are visualized.
Figure 2: Statistical chart comparing RNA localizations across development Jambor et al.
Despite being used widely, data visualization is still treated as a final step in research, a way to communicate results once the real analysis is finished. In reality, visualization plays a much earlier and more fundamental role. Visuals expose batch effects, hidden subpopulations, nonlinear behaviors, and experimental artifacts that often remain invisible to summary statistics alone. These insights directly shape which data can be trusted, which controls are needed, and which experiments should come next.
Figure 3: Examples of plots used during the research process to visualize technical and sample variability. Figure 3: Examples of plots used during the research process to visualize technical and sample variability.
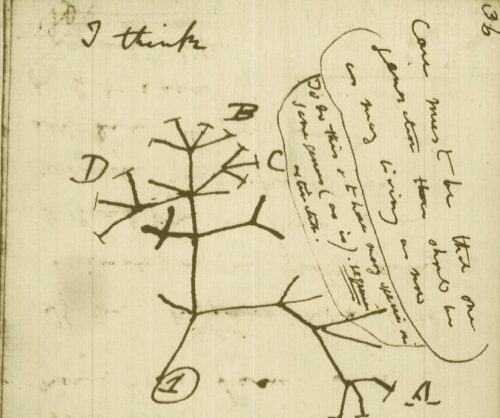
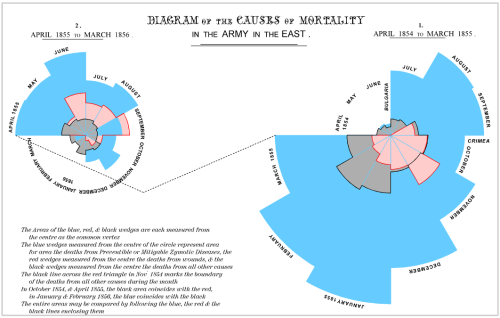
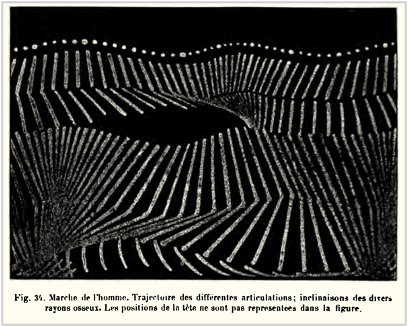
While the urgency to visualize data feels modern, the principle itself is not new. Seeing has always been central to biological understanding. Darwin’s and Linnaeus’s classification of species relied on careful visual comparison. In the nineteenth century, Florence Nightingale pioneered statistical charts to reform healthcare, while John Snow’s maps of cholera outbreaks transformed how disease transmission was understood. In the twentieth century, Michaelis and Menten introduced the kinetic plot as a standardized visual language for enzyme activity, and more recently, interactive genome browsers have made entire genomes navigable at nucleotide resolution.
Darwin’s phylogenetic treeSnow’s Cholera mapightingale’s chart invented to document health care reformsMarey’s animation of human locomotion
Today, data visualization is however still poorly formalized in the life sciences. It lacks dedicated training programs, shared standards, and institutional recognition. This gap matters, as data visualization leads to hypotheses generation, insightful data presentation, and builds trust in the results.
Just as early scientists needed training in scientific drawing to accurately document what they observed, today’s researchers must learn to engineer and interpret data visualizations with comparable rigor. In an era where biology increasingly unfolds in data rather than images alone, learning how to see again has never been more important.
PS – I wrote this looking for discussions on this topic – feel free to reach out helena.jambor – at – fhgr.ch
Reflections from a Workshop by Nivedita Mukherjee1 and Mateus de Oliveira Lisboa2
1National Centre for Biological Sciences, Tata Institute of Fundamental Research, 560065, India (NCBS-TIFR)
2Core for Cell Technology, School of Medicine and Life Sciences, Pontifícia Universidade Católica do Paraná (PUCPR)
Last November, a unique gathering took place at Fanhams Hall in Hertfordshire, UK, for The Company of Biologists’ workshop titled “Decoding Whole Genome Doubling: Insights from Evolution, Development, and Disease.” Organised by Renata Basto and Zuzana Storchova, the workshop brought together a highly interdisciplinary cohort of scientists studying whole-genome doubling (WGD) across organisms, from flies and frogs to plants and humans.
As two of ten early-career researchers selected to participate, we had the rare opportunity to engage in close discussions with leaders in the field, present our own work, and explore the diverse biological contexts in which the entire chromosomal complement of a cell is doubled, changing its ploidy – the number of copies of the genome. Across three intense days, we uncovered how such changes can drive cellular adaptations, precipitate disease, and drive evolutionary innovation.
A Biological Balancing Act
Ploidy variation is ubiquitous in biology. Even within a single organism, cells often carry different numbers of genome copies depending on the tissue type and developmental stage. For example, while humans are diploid as a species, polyploid cells are routine in organs such as the liver, heart and pancreas. These can arise from either programmed or accidental WGD, resulting from the skipping of one or more cell-cycle steps or through cell fusion. Once polyploidy arises, the cell is thrust into a new regime, one that demands extensive adaptation if it is to continue functioning or proliferating without compromising genome integrity.
Initial responses to genome doubling can be defensive. DNA damage and stress-response pathways are activated, and cell death is a common outcome unless these surveillance mechanisms are suppressed. Survival alone, however, is not enough. To continue to divide successfully, polyploid cells must carefully segregate their excess chromosomes. This often involves re-establishing functional mitotic spindles despite the presence of supernumerary centrosomes. Several talks highlighted molecular strategies that allow polyploid cells to survive and divide by rewiring apoptotic signalling and stabilising centrosome numbers.
Bigger Cells, Bigger Questions
Polyploid cells aren’t just more complex; they’re physically larger. This increase in volume comes with consequences: intracellular transport must traverse greater distances, metaphase spindles must span wider plates, and increased metabolic demands must be sustained. Several speakers described how these physicochemical constraints can be systematically probed using in silico models alongside in vitro perturbations.
Beyond these geometric and energetic challenges, it is tempting to assume that increased DNA content translates to increased gene expression. However, multiple presentations demonstrated that gene expression scales unevenly and nonlinearly with ploidy, with pronounced disparities between transcript abundance and protein yield. In some contexts, specific genes are selectively up- or down-regulated, rewiring the cell’s regulatory network in unexpected ways. This decoupling of genome content from functional output overturns the view of polyploid cells as merely “amplified” versions of their diploid counterparts.
Genomic Instability and Interventions
For a cell, doubling its genome is not always a deliberate developmental strategy or a trivial bookkeeping error; it is often a high-risk gamble. Cardiomyocytes, for instance, become increasingly polyploid with age or heart disease, accompanied by a substantial accumulation of mutations. Whether polyploidy in such settings is adaptive, maladaptive, or merely tolerated remains an open question—one that echoes broader uncertainties about where normal physiology ends, and pathology begins.
Polyploidy-related genomic instability is also characteristic of cancer, with genome-doubled tumours showing many more chromosomal aberrations than diploid tumours. While high levels of such aberrations can be lethal, genome doubling also casts a paradoxical safety net by creating “genomic backups” that restore essential functions. At its extremes, WGD-induced genomic instability can be spectacular. One striking example is chromothripsis, where mis-segregated chromosomes trapped within micronuclei shatter and reassemble in chaotic ways, creating the genomic equivalent of a misassembled jigsaw puzzle.
A silver lining is that, while such drastic mutational events can accelerate cancer cell evolution, their dependence on these aberrant states may also be their undoing. Because WGD imposes unique cellular stresses, polyploid cancer cells acquire distinct vulnerabilities that may be therapeutically exploitable. The workshop highlighted several efforts to leverage these weaknesses, offering hope for selective treatments that preferentially target cancer cells while sparing normal tissue.
Innovation Across Evolutionary Time
At the macro scale, genome doubling acts as a powerful driver of evolutionary change. Ancient WGDs have profoundly shaped the genomes of present-day microorganisms, plants, and vertebrates, with recent events producing drought-resistant plants, pest-resistant crops, and amphibians adapted to arid environments. Polyploid lineages often adapt rapidly, restructuring physiological and metabolic pathways, and frequently adopting self-pollination or asexual reproductive strategies to overcome early barriers to evolutionary establishment.
Across the tree of life, WGD is frequently followed by episodes of rapid diversification, though these bursts typically occur only after a delay. This “radiation lag” is thought to reflect the time required for re-diploidisation, during which redundant genomic content is reduced, and ohnologues or duplicated genes diverge in function. As gene dosage is re-balanced and new regulatory networks emerge, WGD becomes a springboard for long-term evolutionary innovation.
Such innovations may confer resilience during periods of ecological upheaval. Compelling evidence for this comes from the observation that WGD events in the evolutionary history of species are disproportionately clustered around major extinction events, such as the Cretaceous–Palaeogene boundary. Interestingly, environmental stressors, such as extreme heat or cold, can induce the formation of unreduced gametes, providing a direct link between ecological pressures and the origin of polyploid species.
A Shared Language for Polyploidy
Amid the breadth of scientific discussions at the workshop, one unifying theme stood out: WGD is neither an anomaly nor a biological accident. From plants surviving climate shifts to tumours evading physiological checks, genome doubling repeatedly emerges as a powerful strategy in life’s toolkit. Yet it remains a double-edged sword, capable of driving adaptation or unleashing instability.
This raises big questions. In healthy tissues, polyploidy seems to balance on a knife-edge between careful regulation and stochasticity. Understanding this balance could reveal why certain cells become polyploid, how tissues keep their abundance in check, and what stops them from turning cancerous. In cancer cells, it’s still unclear if WGD is a driving force or a downstream manifestation of genomic instability. Moreover, scientists are still figuring out how evolutionary pressures shape newly formed and established polyploid lineages, and how the opportunities for adaptation and diversification play out across different tissues and species.
What the field now needs is a shared language across disciplines. Polyploidy in microbes, plants, animals, cancer, and development has too often been studied in silos. By integrating these perspectives, we may finally decipher how a single genomic event reverberates from the scale of individual cells all the way to the evolution of multicellular species.
This workshop marked a step in that direction, and for us, a turning point in our understanding of what it means to live with, adapt to, and evolve through an extra genome.
Mateus de Oliveira Lisboa is a PhD student at the Core for Cell Technology, PUCPR (Brazil), studying how whole-genome doubling shapes cell fate, evolution, and disease. With a background in chromosome biology, stem cells, and the cancer microenvironment, he integrates molecular biology and bioinformatics to explore the causes and consequences of large-scale genomic alterations, always through an evolutionary lens. Outside the lab, he explores mountains, photography, astronomy, and birds.
Nivedita Mukherjee is a PhD student at the National Centre for Biological Sciences (NCBS–TIFR), Bengaluru, India, where she studies cancer evolution through the computational analysis of large-scale genomics datasets. Her research integrates statistical genomics and evolutionary theory to examine how whole-genome doubling reshapes selective pressures in cancer. Beyond research, Nivedita writes popular science articles and enjoys singing, travelling, photography, and reading.
[Editorial from Development by James Briscoe, Swathi Arur, Anna Bigas, Dominique Bergmann, Benoit G. Bruneau, Cassandra G. Extavour, Paul François, Anna-Katerina Hadjantonakis, Haruhiko Koseki, Thomas Lecuit, Matthias Lutolf, Irene Miguel-Aliaga, Samantha A. Morris, Kenneth D. Poss, Elizabeth J. Robertson, Peter Rugg-Gunn, Debra L. Silver, James M. A. Turner, James M. Wells, Steve Wilson.]
Every researcher knows the anticipation and trepidation that come with submitting a paper to a journal. Years of effort have been distilled into a few thousand words and a handful of figures containing the metaphorical (and often literal) sweat from long hours and hard toil in the lab. What will the reviewers say? How will the editor deal with it? At Development, we understand the anxiety and the investment that goes with a paper submission. Our mission is to provide the kind of expert, constructive review that not only evaluates your work but helps it achieve its full potential for lasting scientific impact. But we know that the comments provided by reviewers don’t always live up to this expectation. We’ve heard the concerns. Some of these frustrations reflect deeper, systemic issues across scientific publishing: the feeling that revision requests can expand beyond what is feasible, that editorial decisions are not always transparent, and that standards can seem uneven across subfields such as developmental and stem cell biology. These are not challenges unique to Development, but we acknowledge them and continue to refine our editorial practices to address them wherever possible.
We’ve been told that Development is ‘too hard to publish in’, that reviews are unnecessarily harsh, that revisions are excessive and time-consuming. These criticisms matter to us. Even though sometimes it might be more perception than reality, we won’t pretend that there isn’t some truth in these criticisms. As active research scientists ourselves, we Editors face similar frustrations with our own papers. We want to be transparent about Development’s reviewing process and explain how we endeavour to get the balance right so that it serves both authors and the scientific community.
First, the numbers behind the perception. The bottom line is that, for the past 10 years, 35-45% of papers submitted to Development ultimately get published. Let us break this down. Roughly 65% of the manuscripts submitted to Development are sent out for peer review. We only editorially reject papers when the topic of the article is beyond our scope and expertise, or it is clear to us that the study would not be supported by our peer reviewers. A rapid rejection allows authors to quickly redirect their manuscript to more appropriate venues and, where relevant, we facilitate direct transfers to our sister journal, Biology Open. Of the papers we send for peer review, well over 50% receive positive reviews from reviewers and we ask the authors to revise and resubmit these…
 (4 votes)
(4 votes) Loading...
Loading...

 (No Ratings Yet)
(No Ratings Yet)