We are arranging a 2-day conference together with the Swedish Society for Developmental Biology (SWEDBO), Finnish Society for Developmental Biologists, and Danish & Norwegian Developmental Biologists the 3rd Nordic Meeting on Development, Stem Cells and Regeneration in Copenhagen in October 5-7th, 2022
The line up of invited speakers is outstanding and brings together experts in developmental and stem cell biology and regeneration! Attending the meeting is a great opportunity to meet with developmental biologists, from the Nordic countries and beyond. All speakers will be there ‘in person’ allowing for lots of networking.
There are many opportunities for short talks, poster prizes, and student and postdoc activities are planned in addition. The registration is now open and it includes membership in one of the Nordic Developmental Biology associations! Follow the link to register: https://nordicdevelopmentalbiology.com
Dr. Denise Allen and Dr. Tomasz Nowakowski at the University of California, San Francisco recently published an article in Science where they revealed a dual origin for astrocytes in the human cortex. Using a combination of fate mapping and single cell analysis, they revealed that the two stem cell niches in the developing cortex give rise to spatially, morphologically, and molecularly distinct populations of astrocytes. The Node asked them to give us a behind the scenes look at how the story came together:
How did you get started on this project?
TJN: For a very long time, we have been interested in the question of why the brains of primates and humans are so much larger and complex than the brains of mice, which we study frequently in the laboratory. Differences in brain size can be found very early on during development, and therefore it was plausible to hypothesize that differences in the way radial glia, which act as neural stem cells, develop could contribute to these differences. In our prior work, we found that animals with large brains, such as primates or humans, contain a greater diversity of radial glia subtypes compared to mice. In particular, we found that based on gene expression profiles, radial glia could be divided into truncated radial glia and outer radial glia, which are located in two anatomically distinct niches of the developing cortex.
DA: During my undergraduate neuroscience classes I was always struck by the deep knowledge we have about the development of neurons in the cerebral cortex, but astrocytes and other glia seemed to be so often overlooked. During my rotation in the Nowakowski lab, I became fascinated with Tom’s preliminary data that suggested distinct subtypes of radial glia could give rise to distinct astrocyte populations. I was really excited by the fact that large brain mammals, including primates and humans, seem to have a different repertoire of radial glia compared to rodents, as well as much more complex astrocytes. So the possibility to study the unique features of human development with a focus on astrocytes was a dream come true.
What was already known about the developmental trajectories of radial glia in the developing brain prior to your work?
A lot of work has been done probing the differentiation of outer radial glia (also known as basal radial glia). Numerous papers have shown that they give rise to neurons, oligodendrocytes and supposedly the majority of astrocytes, but the role of truncated radial glia has not been studied in great detail. Previous studies have suggested that because few mitotic cells can be found in the ventricular zone stem cell niche during midgestation in primates and humans, that the truncated radial glia that reside in this zone are unlikely to serve as a major source of new cells. We decided to challenge this assumption by labeling progenitors in the ventricular zone and determining the fates of the resulting cells. To our surprise, we found that neurons, oligodendrocytes, and astrocytes continue to be produced by ventricular zone progenitors.
Can you summarize your findings? What was the key experiment?
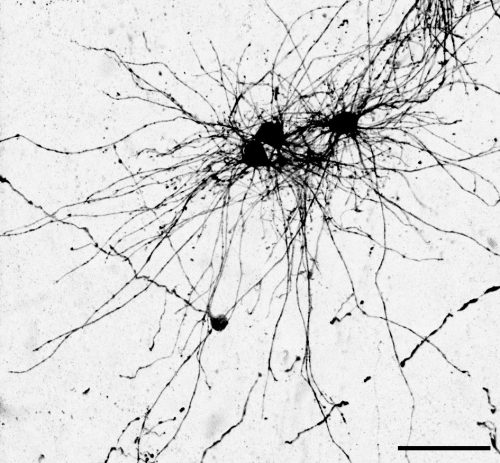
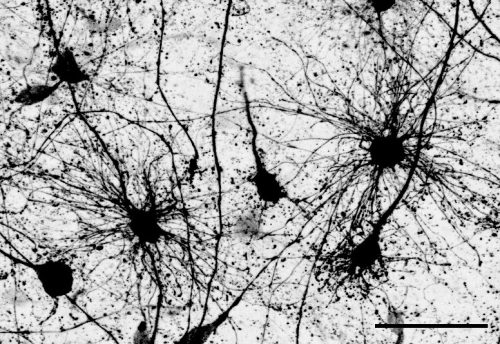
The key experiment involved labeling progenitor cells that occupy anatomically distinct niches that truncated and outer radial glia cells, and tracing the fates of cells that they produce. We found that while both populations broadly produce similar cell types (neurons, astrocytes and oligodendrocytes), they produce very distinct subtypes of astrocytes. In a series of very striking results, we found that truncated radial glia give rise to astrocytes that migrate to the cortical plate, while outer radial glia give rise to astrocytes that do not migrate, and instead differentiate locally in the outer subventricular zone.
We often speak about the diversity of neurons, but classical studies have also shown that astrocytes can be remarkably diverse, even in early development. I wanted to further explore the diversity of the astrocyte subtypes we identified, but it is challenging to connect these classical descriptions of astrocyte subtypes to modern-day descriptions such as those derived from single cell sequencing. To solve this problem, I took advantage of a method called Patch-seq which gave us the ability to aspirate the contents of a cell that was previously defined based on its morphology and position, and then performing sequencing of that cellular contents to determine a molecular identity. This analysis was key for bridging our cellular definitions based on morphology and developmental cell lineage, and linking them to molecular markers. This allowed me to bring the story in full circle.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
The very first experiment I performed that involved labeling these two different stem cell niches resulted in a distribution of cell types that could not have been more different. Many comparisons in developmental biology rest upon small differences between conditions. To see such a stark difference in the distribution of cells–especially of glia–was an exciting moment that defined the course of the project very early on in my PhD.
Another surprising finding was when we started closely comparing classical drawings by Ramon y Cajal and Retzius and to images of our astrocyte subtypes. Remarkably, we found that our “dense bulbous” astrocytes were clearly depicted in those early records, but these cells have rarely been mentioned in modern literature. This realization gave us a lot of confidence that the cells we were observing were a real phenomenon and not an experimental artifact. These cells had just been lying in wait, waiting for someone to put the spotlight on them.
And what about the flipside? Any frustrations or despair?
The Universe really conspired against us when we were trying to finish experiments for the revision of the paper. I set up the last three revision experiments in late December, when we suddenly found out that it was time to move our lab to a new building at UCSF. We came up with an elaborate system to keep the cultures going while we moved and they seemed to have survived, until someone suddenly noticed that the incubator had failed and the alarm hadn’t gone off. What followed was two months of issue after issue trying to repeat these last two experiments, but we finally got there in the end!
Where will this story take the lab?
This work has inspired several new projects in the lab. We are excited to examine if similar findings can be replicated in other models of brain development such as cerebral organoids, what these unique subpopulations look like in the adult brain, and what role they might play in disease states. I’m also hoping this work will also attract more trainees interested in glial development to the lab!
What is next for you/the lab after this paper? Let us know if you are continuing this research, or starting/looking for a new position.
Denise has graduated and is currently interviewing for computational biologist roles in biotech. She is looking forward to delving into the “big data” side of biology, and working towards making a significant impact on patients’ lives.
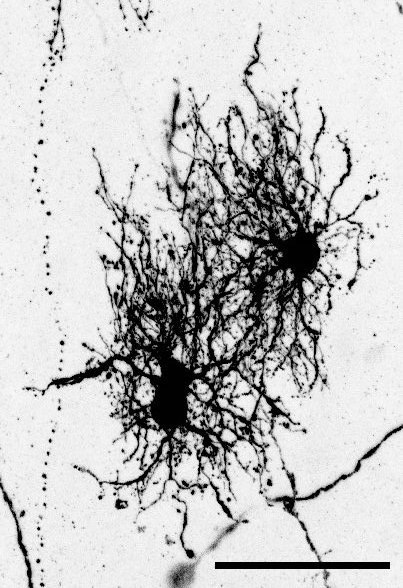
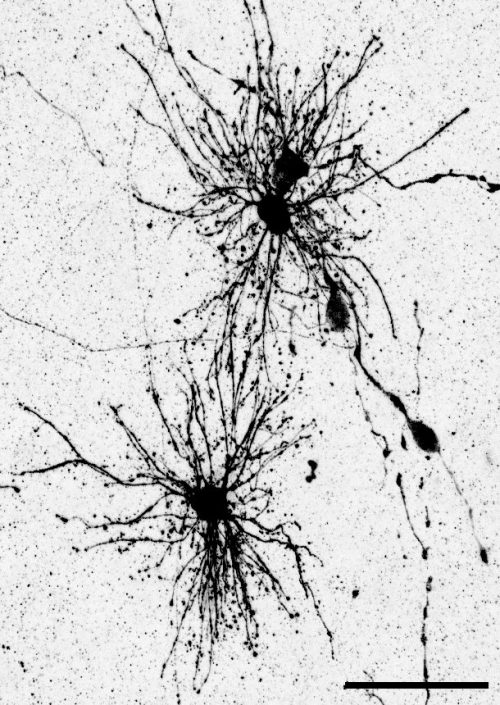
“Dense bulbous” astrocytes derived from the VZ“Dense bulbous” astrocytes derived from the VZ“Dense bulbous” astrocytes derived from the VZ “Dense smooth” astrocytes derived from the OSVZ“Dense smooth” astrocytes derived from the OSVZ (1 votes) Loading...
Development’s biennial meeting, From Stem Cells to Human Development, will be taking place in mid-September (11-14) at beautiful Wotton House in Surrey, UK. After a very successful virtual meeting in 2020, we’re excited to be meeting in person again, but we wanted to explore ways of making part of the meeting accessible to the broader community. We’re therefore delighted to announce that we’ll be livestreaming one session of this meeting, and the recording is available below.
Session details (all times GMT+1): 16:00 Sarah Teichmann (Wellcome Sanger Institute, UK): Human development: one cell at a time 16:30 Sergiu Pasca (Stanford University, USA): From stem cells to assembloids: constructing and deconstructing human nervous system development and disease 17:00 Panel discussion: Technical, ethical and legal challenges of studying early human development Chair: Patrick Tam (University of Sydney, Australia) Panellists: Amander Clark (University of California Los Angeles), Robin Lovell-Badge (The Francis Crick Institute, UK), Sergiu Pasca, Sarah Teichmann, Magdalena Zernicka-Goetz (University of Cambridge, UK and CalTech, USA) 18:00 Close
The method was straightforward: take a bunch of writers, novelists, a playwright, science communicators and scientists from all over the world, from fields as diverse as astrophysics and climate science to materials engineering, neurobiology, and evolution, confine them to a 16th century manor, and get them talking. The result: The Company of Biologists’ whacky and wonderful Creative Science Writing Workshop.
Wiston House, South Downs National Park: the experiment site.
Good scientific writing has the power to help communities make more informed decisions, and a creative route could make it more accessible and exciting for all. But here, without the certainties of a regular science meeting where everyone shares a common interest in a method, organism or question, and data and information that forms the focus of most scientific discussions, what would happen was anyone’s guess.
Any disquiet quickly evaporated in the opening session where everyone introduced themselves through stories, childhood memories, and mementos: a bottled book, a handmade urn, the steady pulse of a ticking metronome, the clinking beads of a treasured necklace, a traditional Indian kolam drawn in flour before our eyes. It was immediately clear that despite our diverse backgrounds and experiences, everyone shared a passion for storytelling and science in all its forms. The Workshop began with a bang!
The programme included a variety of topics and activities. We read S.J. Gould, Primo Levi and the brilliant Karen Joy Fowler, looking for the ingredients of great writing and pondering the authors’ process and intent. We critiqued the work of other delegates, providing structured feedback – a rather nerve-racking exercise when one considers the differences between academic and creative writing. In conventional science writing, the authors present new facts and relationships that help us better understand our world, without ever revealing themselves. In creative writing the opposite holds true; every piece exposes the writer, their style, their quirks of character, beliefs, and passions. This Workshop managed the juxtaposition remarkably well.
There were also sessions on structuring writing, getting published, and the role of agents. Most importantly, there was time to write, review, and revise. Inside oak-panelled rooms, within the verdant grounds, and in the sunny conservatory, relationships grew, word counts climbed, and inspiration abounded.
Over a period of three and a half days, the Workshop exposed delegates to the world of writing and publishing. Aspiring writers learned from one another as well as from the established writers – who were incredible mentors. It was a journey that traversed science and writing, lab and the field, life and the page, lyrical prose and cold hard facts. In doing so, a new community was formed.
Applications will remain open until the position is filled.
The Gambetta lab is recruiting a Postdoctoral researcher to study how genome architecture impacts gene regulation in development.
Project
The advertised project addresses the fundamental question of how regulatory elements are guided to their target genes. You will study a paradigm we uncovered in which genes are controlled by regulatory elements located at flexible and surprising large genomic distances. This model is powerful and unique.
You will use genetics (Drosophila genome-engineering, genetic screens, comparative evolution), genomics (transcriptomics, chromatin accessibility, chromosome conformation capture, genome-wide screens, single-cell genomics), imaging (fixed and live), and biochemistry (proteomics).
This work is expected to continue to reveal new evolutionary perspectives into the relevance of 3D genome folding for correctly wiring genes to their regulatory elements. For more information on our research check our lab website: http://www.gambettalab.org
Job information
Expected start date in position : as soon as possible or to be agreed
Contract length : 1 year, renewable 2 x 2 years, maximum 5 years
Your responsibilities
You will use multidisciplinary approaches such as genomics, genetics, imaging, and/or biochemistry in the fruit fly Drosophila melanogaster. You will present your results during seminars with gene regulation research labs in Lausanne. You will collaborate with other labs in Lausanne and abroad. Full funding for the position is available, but application to fellowships is also expected.
Your qualifications
You are a dynamic and rigorous scientist with a PhD degree in Biology or a related discipline. You have experience in genomics (next-generation sequencing library preparation), molecular biology, genetics, biochemistry or imaging. You are a critical thinker, a team player eager to participate in scientific discussions but able to work independently. You have a strong interest in developing your skills in multidisciplinary experimental strategies to understand basic mechanisms in gene regulation.
Your benefits
The Gambetta lab is hosted at the Center for Integrative Genomics (CIG) at the University of Lausanne (UNIL), a vibrant, well-funded institute with a focus on functional genomics and equipped with modern core facilities (see www.unil.ch/cig).
It is embedded in the broader Lausanne research environment that includes two universities (UNIL, EPFL), the Swiss Institute of Bioinformatics, Ludwig Center for Cancer Research, university hospital, and a cluster of biotech companies flourishing in the larger lake Geneva area.
The Gambetta lab tightly networks with other gene regulation research laboratories at UNIL and EPFL, and collaborates with the on-site Bioinformatics Competence Center. There are regular possibilities to present and participate in local or international conferences and workshops. Hard and soft skill, and career development courses are offered on campus.
We offer a nice working place in a multicultural, diversified and dynamic academic environment.
Your application
Please email lab head Prof. Maria Cristina Gambetta (mariacristina.gambetta@unil.ch). Provide your CV, a brief description of your research experience, and why you think your research interests complement ours.
Posting my lab report from last summer again since I can’t log into my last account. Update: I’ll be starting an Mres looking at the mutual suppression of the cardiac versus skeletal muscle programme!
Biomedical research tries to understand among other things, how during development, gene expression determines cell fates. One of the aspects that we look at is how cells are recruited to the heart and how to apply this knowledge to cardiac therapy. However, cell fate decisions in the head mesoderm, the tissue responsible for delivering the heart, craniofacial muscle, parts of the skull and vasculature, are poorly understood. Last summer, I had the opportunity to work with Dr. Susanne Dietrich, who studies the formation of muscles in early development, including, but not only, genes responsible for committing cells into a mesodermal fate. I was part of ongoing research addressing this question, at her lab in Portsmouth. The Dietrich lab has shown that initially, the entire head mesoderm has cardiac competence. However, at early neurula stages of development, the cardiac inducer Bmp2 fails to induce the cardiac programme and instead, it induces Msc, a craniofacial precursor marker. It is not yet clear how this switch in developmental competence is achieved. We hypothesize that Wnt may be responsible for the switch, inducing an early expression of craniofacial precursor markers and downregulating cardiac markers. To test this idea, I grafted heparin-coated acrylic beads soaked in recombinant Wnt3a, the Wnt inhibitor Sfrp2 and Bmp2 or bovine serum albumin as control, into HH7/8 embryos. Embryos were cultured for 6 hours, the time sufficient for Bmp to induce Msc. I then used In situ hybridization to analyse the expression of Msc, the cardiac marker Nkx2.5 and the Wnt responsive genes Pax3 and Axin2. We found that Wnt did not upregulate and Sfrp2 did not downregulate Pax3 and Axin2 (data not shown), probably because it takes more than 6 hours to change the expression of these genes. However, Wnt3a did downregulate Nkx2.5 as expected (Fig.1). Nonetheless, Wnt did not upregulate Msc (Fig.1), suggesting that the concentration used and the 6-hour time period might not have been enough for Wnt to participate in the activation of Msc. This was against our hypothesis, and we wondered why that might be. So, we decided to test the effect of Wnt on the paraxial head mesoderm marker Cyp26C1, an inhibitor of retinoic acid signalling. We found that Cyp26C1 was suppressed (Fig.1). Thus, Wnt may in fact suppress paraxial head mesoderm features.
I am now faced with new questions: What really is the role of Wnt? Does it suppress heart and paraxial mesodermal features because it posteriorizes the tissue? To answer this question, I will have to analyse if Wnt causes an ectopic expression of posterior information markers (e.g: Raldh2 and Hoxb1). And our original question is not answered: what facilitated the switch from cardiac to skeletal muscle competence? I am intrigued by these results, and I do want to find out the right mechanism that causes this switch. I will be working on this project throughout the next academic year, looking at different embryonic stages, different concentrations of Wnt, different Wnt inhibitors (e.g.: Dkk) and possibly, longer culture periods. I am hoping to find results that tell us if Wnt signalling is or not responsible. If Wnt is not involved, what else could it be? Many other signalling cascades converge on the head mesoderm, and they could be tested using similar approaches. Alternatively, I could use small molecule inhibitors of signalling cascades on embryos cultured as Cornish pasties. We also have to consider that the epigenetic landscape might change over time, and cardiac genes might be put out of use. This would require a different approach, chromatin immunoprecipitation. I would love to learn about chromatin immunoprecipitation and work on this approach during my master’s or PhD. I want to continue working on these questions with Dr. Susanne throughout the next years of my academic life. I hope that with my 3rd year module “genes and development” I will gain more insight into new experimental methods used in developmental biology and maybe use them as an approach in my project. I would love to work with different model organisms and upgrade my knowledge with new techniques that may facilitate the research. Working with Dr. Susanne and her team made me grow as a scientist. I remember the week before starting on my project I was so nervous I even had nightmares about it. But the people in the lab were very kind and helpful and they made me feel at home. During the summer, I was faced with some of the ups and downs of science. In situs that did not work, embryos that were accidentally lost, beads not sticking, and all of that (Particularly the last one), allowed me to develop my problem-solving skills and patience (especially while grafting beads). Being part of a research group made me realize that I do not see myself doing anything else. I love planning my experiments and I love the practical part. I am also very interested in presenting and explaining my results to other people. I look forward to continue working on developmental biology throughout my studies. I plan on continue my education with Dr. Susanne, working on finding the mechanism behind this cardiac to skeletal muscle switch, and other projects.
“Purple male and white female, or white male and purple female: whichever way he did the cross, he got the same results. And Mendel realised that was telling him something really, really important. It takes two.”
Professor Alison Woollard
In the latest episode of the Genetics Unzipped podcast, we celebrate the 200th birthday of Gregor Mendel and learn about the latest research into the genetics of human social traits that would have blown his mind. Professor Alison Woollard from the University of Oxford shares some of the less well-known stories about the forefather of genetics, Professor Greg Radick from the University of Leeds ponders what would have happened if Darwin had read Mendel’s papers, and Dr Rosa Cheesman from the University of Oslo explains how genetics can help us understand complex human traits and behaviours such as education.
Reproduction is the process whereby organisms pass on their genetic information to the next generation. Reproductive strategies are highly varied and success is measured not only by the number of offspring produced but also the reproductive success of the next generation. Mammals (and some cartilaginous fish and reptiles) have evolved a strategy whereby fertilised eggs are retained inside the female, the embryo receives nourishment from the mother via a placenta and the offspring a born alive. Further maternal resources in the form of milk are exclusively provided by the mother alongside high quality maternal care. This intimate and bidirectional relationship between the mammalian mother and her offspring increases the chances of offspring surviving to adulthood to reproduce but places a substantial burden on the mother. Direct nurturing by the mother also creates an excellent opportunity for exploitation both by the offspring she carries and cares for, and by the male parent. While these adaptations have led to the global success of mammals, they are also linked to highly common pregnancy complications which impact the health of the mother, her offspring and, in some cases, her offspring’s offspring. This meeting will cover establishment of the mammalian germline, embryonic and placental development and common pregnancy complications finishing with a lively debate on the evolution of pregnancy led by invited speakers.
There will also be a Genetics Society Medal talk by Professor Robin Lovell-Badge (CBE, FRS FMedSci).
Spatial mapping of embryoid body models to gastrulating marmoset embryos from Castillo-Venzor, et al.
Origin and segregation of the human germline Aracely Castillo-Venzor, Christopher A. Penfold, Michael D. Morgan, Walfred W. C. Tang, Toshihiro Kobayashi, Frederick C. K. Wong, Sophie Bergmann, Erin Slatery, Thorsten E. Boroviak, John C. Marioni, M. Azim Surani
Exocyst Inactivation in Urothelial Cells Disrupts Autophagy and Activates non-canonical NF-κB Michael A. Ortega, Ross K. Villiger, Malia Harrison-Chau, Suzanna Lieu, Kadee-Kalia Tamashiro, Amanda J. Lee, Brent A. Fujimoto, Geetika Y. Patwardhan, Joshua Kepler, Ben FogelgrenMichael A. Ortega, Ross K. Villiger, Malia Harrison-Chau, Suzanna Lieu, Kadee-Kalia Tamashiro, Amanda J. Lee, Brent A. Fujimoto, Geetika Y. Patwardhan, Joshua Kepler, Ben Fogelgren
CITED2 Is A Conserved Regulator Of Deep Hemochorial Placentation Marija Kuna, Pramod Dhakal, Khursheed Iqbal, Esteban M. Dominguez, Lindsey N. Kent, Masanaga Muto, Ayelen Moreno-Irusta, Keisuke Kozai, Kaela M. Varberg, Hiroaki Okae, Takahiro Arima, Henry M. Sucov, Michael J. Soares
A molecular mechanism for membrane chaperoning by a late embryogenesis abundant protein Xiao-Han Li, Conny W.H. Yu, Natalia Gomez-Navarro, Viktoriya Stancheva, Hongni Zhu, Cristina Guibao, Andal Murthy, Boer Xie, Michael Wozny, Benjamin Leslie, Marcin Kaminski, Ketan Malhotra, Christopher M. Johnson, Martin Blackledge, Balaji Santhanam, Douglas R. Green, Junmin Peng, Wei Liu, Jinqing Huang, Elizabeth A. Miller, Stefan M.V. Freund, M. Madan Babu
Histone demethylome map reveals combinatorial gene regulatory functions in embryonic stem cells Yogesh Kumar, Pratibha Tripathi, Pushkar Dakle, Majid Mehravar, Varun K. Pandey, Michael J. Bullen, Zhongming Zhang, Dhaval Hathiwala, Marc Kerenyi, Andrew Woo, Alireza Ghamari, Alan B. Cantor, Lee H. Wong, Jonghwan Kim, Kimberly Glass, Guo-Cheng Yuan, Luca Pinello, Stuart H. Orkin, Partha Pratim Das
Symbiosis-driven development in an early branching metazoan Aki H. Ohdera, Justin Darymple, Viridiana Avila-Magaña, Victoria Sharp, Kelly Watson, Mark McCauley, Bailey Steinworth, Erika M. Diaz-Almeyda, Sheila A. Kitchen, Angela Z. Poole, Anthony Bellantuono, Sajeet Haridas, Igor V. Grigoriev, Lea Goentoro, Elizabeth Vallen, David M. Baker, Todd C. LaJeunesse, Sandra Loesgen, Mark Q. Martindale, Matthew DeGennaro, William K. Fitt, Mónica Medina
Multi-chamber cardioids unravel human heart development and cardiac defects Clara Schmidt, Alison Deyett, Tobias Ilmer, Aranxa Torres Caballero, Simon Haendeler, Lokesh Pimpale, Michael A. Netzer, Lavinia Ceci Ginistrelli, Martina Cirigliano, Estela Juncosa Mancheno, Daniel Reumann, Katherina Tavernini, Steffen Hering, Pablo Hofbauer, Sasha Mendjan
Odd skipped-related 1 controls the pro-regenerative response of Fibro-Adipogenic Progenitors Georgios Kotsaris, Taimoor H. Qazi, Christian H. Bucher, Sophie Pöhle-Kronawitter, Vladimir Ugorets, William Jarassier, Stefan Börno, Bernd Timmermann, Claudia Giesecke-Thiel, Pedro Vallecillo-García, Aris N. Economides, Fabien Le Grand, Petra Knaus, Sven Geissler, Sigmar Stricker
Emergent dynamics of adult stem cell lineages from single nucleus and single cell RNA-Seq of Drosophila testes Amelie A. Raz, Gabriela S. Vida, Sarah R. Stern, Sharvani Mahadevaraju, Jaclyn M. Fingerhut, Jennifer M. Viveiros, Soumitra Pal, Jasmine R. Grey, Mara R. Grace, Cameron W. Berry, Hongjie Li, Jasper Janssens, Wouter Saelens, Zhantao Shao, Chun Hun, Yukiko M. Yamashita, Teresa M. Przytycka, Brian Oliver, Julie A. Brill, Henry M. Krause, Erika L. Matunis, Helen White-Cooper, Stephen DiNardo, Margaret T. Fuller
Accurate simultaneous sequencing of genetic and epigenetic bases in DNA Jens Füllgrabe, Walraj S Gosal, Páidí Creed, Sidong Liu, Casper K Lumby, David J Morley, Tobias W B Ost, Albert J Vilella, Shirong Yu, Helen Bignell, Philippa Burns, Tom Charlesworth, Beiyuan Fu, Howerd Fordham, Nick Harding, Olga Gandelman, Paula Golder, Christopher Hodson, Mengjie Li, Marjana Lila, Yang Liu, Joanne Mason, Jason Mellad, Jack Monahan, Oliver Nentwich, Alexandra Palmer, Michael Steward, Minna Taipale, Audrey Vandomme, Rita Santo San-Bento, Ankita Singhal, Julia Vivian, Natalia Wójtowicz, Nathan Williams, Nicolas J Walker, Nicola C H Wong, Gary Yalloway, Joanna D Holbrook, Shankar Balasubaramanian
Maximizing CRISPRi efficacy and accessibility with dual-sgRNA libraries and optimal effectors Joseph M. Replogle, Jessica L. Bonnar, Angela N. Pogson, Christina R. Liem, Nolan K. Maier, Yufang Ding, Baylee J. Russell, Xingren Wang, Kun Leng, Alina Guna, Thomas M. Norman, Ryan A. Pak, Daniel M. Ramos, Michael E. Ward, Luke A. Gilbert, Martin Kampmann, Jonathan S. Weissman, Marco Jost
Adam Shellard, a postdoc in Roberto Mayor’s lab, was the winner of the 2022BSCB Postdoctoral Research Medal. We caught up with Adam over Teams to find out more about his career path so far, his evolving research interests and the Cell Migration webinar series that he started during the pandemic.
Where are you originally from?
I grew up in London, before going to the University of Manchester for my undergraduate studies. As part of this course, I completed a year-long internship at Thomas Jefferson University in Philadelphia, USA in Renato Iozzo’s lab. I spent a lot of time doing western blots and qPCRs but it was a great experience, both in the lab and having the opportunity to travel.
Why did you choose Roberto Mayor’s lab for your PhD?
I was on the Wellcome Trust Stem Cell and Developmental Biology programme at UCL, which meant I spent my first year doing rotations in different labs. When I started on the programme, I was interested in everything and didn’t have a special interest in any particular topic. I tried to choose labs where I could learn different techniques. I went to a lab that did more biochemistry; I went to one that used electron microscopy; and I did mouse work and live imaging for the first time. The rotations were a great opportunity to try lots of different techniques and topics to discover what I was most interested in. In the end, a big reason for choosing Roberto’s lab was that it was a good environment, and I really liked the people there. The topic didn’t matter so much at that stage because I felt that I could become interested in anything! I liked the fact they had lots of microscopes, and a lot of cool projects were going on at that time.
Can you tell us about your PhD research?
When I started my PhD, I was supervised by Elena Scarpa, who is now a group leader in Cambridge. She had some preliminary data showing an actomyosin cable around the edge of a neural crest cell cluster when it was dissected out and imaged in vitro. My project was to look at the role of actin and myosin during neural crest cell migration as we didn’t know anything about it. This sounds a little crazy, because obviously actin and myosin play a role in migration, but we didn’t know much about how they were involved in the collective migration of the neural crest. So, it started from there. I tried lots of experiments and whilst they worked, there were a lot of negative results in the first three years. Then when I got to the final year, luckily, or serendipitously, a couple of techniques that I’d been trying to work out for a long time, started to work. I finally got laser ablations working on the microscope after searching for so long, so I could very specifically test actomyosin cable function. At the same time, Xavier Trepat’s lab had published some optogenetic constructs which controlled contractility, so I cloned those and used them as well. What we found was that the neural crest, as a cluster, had an actomyosin cable around its edge. And in the absence of any chemoattractant, the cable would contract around the edge, so it would look like the cluster was pulsing. But if you put on a chemoattractant like SDF1 and the cells move by chemotaxis, the SDF1 would inhibit contractility at the front of the cluster, whilst the contractility at the back continued. Using laser ablation and optogenetics, we found that the contractility specifically at the rear of the cluster was driving the directed migration of the neural crest. And we could do that in vitro and in vivo. Of course when you say it, it sounds really obvious because rear contractility contributes to the driving forces of migration in cells, we’ve known that for years. But the novelty was that we had seen the whole cluster was acting like a single cell, where many cells at the front had a protrusion, and many cells at the back had a contraction, which we described as a supracell. And so, the analogy of how a single-cell moves was essentially expanded up to the scale of a cluster. We had this idea for quite a while, but we never had the techniques to address it. We did initially use blebbistatin and attempted to use mosaics, but those methods were very crude, so it was difficult to get any specific conclusions.
Can you tell us about your decision to stay with Roberto for your postdoc and how your research focus evolve during this time?
When I was in my completing research status (CRS) year, which is supposed to be your writeup year, I was struggling to finish off the paper and at the same time I had a deadline to submit my thesis. I was trying to get both of those done. I managed to get the paper submitted and then in for the revisions. Then I had about three or four weeks to write my thesis; I just wrote non-stop for about a month and got the thesis submitted! Then I think I had a round of revisions to do for the paper, so I had to stay on for a little bit longer to do those. Then I had my viva and by that point, it was November or December of that year, and I was just exhausted. I had not planned or considered my future at all at that point. I know you’re supposed to be looking for positions at least six months in advance, you can’t just ring someone up. So, at that point it was Christmas and Roberto offered that I could stay. The idea initially was just to stay for a little bit so that I could continue working until I found a postdoc position. I started my postdoc with Roberto basically a month later. Then, of course, the good thing about staying in a lab is that you already know how to do everything, so you can be super productive. But I did want to push my skillset, because many of the ideas I had required new techniques. So I developed some new methods especially in the context of labelling tissues in vivo and measuring and manipulating mechanics in vivo, as I was keen to explore what I saw was an open question of how chemical and mechanical cues interact in vivo. Fortunately, the lab acquired a nanoindenter to do mechanical measurements at around the same time. The combination of new techniques to address what I thought was a big question, and some promising results, led me to stay for the project.
Can you summarise the main findings from your recent paper?
There are a few main findings, one of them is that we saw durotaxis in vivo. Durotaxis is moving along a stiffness gradient, typically from soft substrates to a stiff substrate, which has been known for 22 years, but there was scarce evidence in vivo. So, that was the first one; we found that the neural crest undergoes durotaxis in vivo as well as chemotaxis, which we previously knew. And then following on from that, we found that the stiffness gradient was being formed by the neural crest cells themselves. The neural crest mechanically modifies an adjacent tissue, the placodes, and in doing so they generate a gradient in their own substrate. That was a very cool and surprising finding. And then towards the end of the paper, we describe how the mechanical signals in durotaxis and chemical signals in chemotaxis interact, how there’s interplay between those two. So essentially, both of these guidance cues work on the same set of proteins, Rho, Rac and actomyosin, influencing contractility. They work together in a cooperative manner. I think that this is going to be a big question for many systems in the next decade or so: how do the chemical signals and the mechanical ones interact to control various biological processes?
DAPI staining of a cryosectioned Xenopus embryo pseudocoloured in green (neural crest) based on Twist in situ hybridisation, and purple-yellow (stiffness gradient) based on Sox2 position and nanoindentation stiffness measurements.
It’s interesting that the stiffness gradient moves with the cells.
Yes, so we had this result that there was a stiffness gradient. But at the time I was brand new to doing mechanical measurements, and as I was quickly learning, doing these measurements in embryos is really difficult. The embryos are super soft, which means that the cantilever you use also has to be really soft. All this means that even the tiniest thing can make a deflection and screw up your measurement; if it sticks, or if there’s a tiny piece of dust, anything like that. So, getting the data from the embryos was a really hard slog in the beginning. After we had observed the gradient, the obvious question is what happens at later time points when the cells move. We could have just seen that the gradient doesn’t move, that could totally make sense as well, the cells just move up the gradient. But, when we saw that the gradient moves, it was a very nice lab meeting slide!
During lockdown, you set up the Cell Migration webinar series, was this something you already had in mind or was it prompted by the pandemic? Can you tell us about the series and why you think it has been so successful?
Yes, the initiation of the series was totally pandemic driven. I don’t think anyone had even thought about virtual seminars pre-pandemic. I initially thought of the idea maybe a week into lockdown, but I didn’t act on it. After about a month or two, I started seeing other seminars pop up and people discussing them on Twitter. It seemed that people were interested in attending, because my initial worry was about putting all the effort in, and then having no one show up! So, it was good to see that people were attending virtual meetings on other topics. And whilst the series was pandemic driven, I’m really happy that it’s still going on. It’s been two years now and it’s still regularly getting high attendance, which is great. I guess it’s popular because people are interested in seeing seminars on their research topic. The cell migration community is a lot more diverse and vibrant than I’d previously known, so it is still attracting a lot of interest! The success is also due to Becky Jones, Jen Mitchell and Ankita Jha, who has taken over my role in organising the webinars, because it is quite a lot of effort.
Do you have any plans for in-person meetings linked to the series, or will you stick with the current format?
I’m not sure, I know some attendees have suggested that maybe the webinars could be organised as a one-day meeting for early career researchers in migration, which I think Jen and Ankita might be considering. But with the return of in-person meetings in cell migration, like the Abercrombie meeting this year and the GRC next year, I’m not sure whether adding another meeting would be a bit overkill. So, I think the virtual meetings will be there for now.
What’s next for you, both short term and longer term?
Short term, I’m trying to finish off a project for which I’m developing a lot of new skills for! I’m hoping to submit the paper before the end of the year, that is my optimistic plan. And then next year, I will be moving on to ‘destination unknown’. I’m considering my options, perhaps a short postdoc in another lab, or maybe a fellowship where you do a few months in many different labs, just to learn some new skills and experience some different environments before applying for positions. That’s one option that I’m considering, but I’m not totally sure yet.
It sounds like you are a big fan of developing new tools and techniques, is that something you enjoy doing?
I enjoy it, but it’s incredibly frustrating. I do it because I have to, not because I want to! I’m kind of attracted to the high risk, high reward projects, the projects that have a lot of potential. But often those are the projects that would have been done if the tools already existed. For example, the project I’m working on now, I’m forced to make new tools. But every time I do this, I always remember how difficult it is and how many months you have to spend developing these tools just to do a single experiment. So yes, I do it because I’m forced to, not because I want to; I enjoy using other people’s tools more than I enjoy making my own!
So, is it more that the question comes first and then you have to find a way to answer it, even if that means tool development?
Yes, that’s absolutely it. For example, in my postdoc I was interested in looking at the neural crest in vivo, in Xenopus, which as anyone who works with Xenopus knows, doing in vivo imaging is really, really difficult. There weren’t even any good antibodies for the neural crest; in the past it had always been inferred by the fact that there’s a fibronectin ECM around it. I spent a few months just developing fluorescence in situ hybridization for the neural crest so I could co-label it with other markers. So yes, the question always comes first, and then whatever technique I need to use to address it as best I can, that comes second.
What do you think the big questions in developmental biology will be over the next ten years?
One of the things that I think will be important, as I mentioned before, is the integration of mechanical and chemical cues, or signals or factors, in trying to understand the cell behaviour in a holistic way. I think that comparatively, we know a lot about signalling pathways and step-by-step processes that are occurring in cells, and now, we’re even getting a decent amount of data about how mechanics affects those processes. But I think in terms of combining them we haven’t even scratched the surface of how these cues come together. And it’s not a trivial thing to do, because trying to do manipulations of those various things without having unwanted side effects is really, really challenging. I think that’s going to be one of the main questions for the next 10 years of developmental biology.
When you’re not in the lab, what do you do for fun?
I enjoy painting, especially with oil paints. I’m really liking ‘Duolingo’ at the minute because I’m awful at languages. I also enjoy travelling, which is a rarity, but I’m happy to accept invitations!

 (No Ratings Yet)
(No Ratings Yet)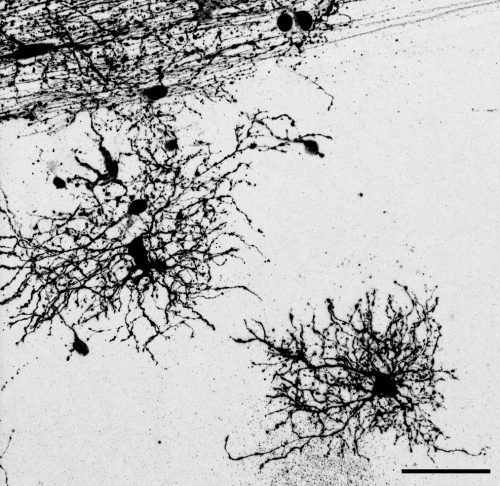
 (1 votes)
(1 votes)