Thanks to everyone who has contributed to the Node in 2021. We hope you have enjoyed interacting with the site over the past year. As a reminder, the Node is a community site so anyone with an interest in developmental biology or stem cells can, once registered, post on the website. This could be a research story, a commentary, an interview or a job or event listing. If you have any suggestions for new features or would like advice from the Node team you can contact us at thenode@biologists.com. But before we look ahead to 2022, we focus on 2021 in numbers:
Regeneration is the mechanism by which a species can restore damaged or missing cells, tissues, organs, or body parts. Different living organisms have vastly differing regenerative capabilities. Humans, while capable of regenerating some organs following damage or disease, most notably the liver, are very limited in their capacity. Other organisms, however, have remarkable regenerative capacities that facilitate re-growth of entire limbs, parts of nervous systems, or even whole-body regeneration like planarians.
Despite the relative prevalence of regeneration and the potential contributions of its research to modern medicine, the field is widely understudied, especially from a comparative evolutionary perspective (Lai and Aboobaker 2018 Dev Bio).
This summer, the BDSB Summer Studentship gave me the opportunity to undertake an exciting research project, further studying the regenerative abilities of the European Garden Spider, Araneus diadematus, within the McGregor Laboratory at Oxford Brookes, under the supervision of Research Fellow and spider expert, Anna Schönauer.
Fritz Vollrath’s work (Vollrath, 1990) describes the remarkable ability of juvenile A. diadematus (Figure 1, A) to quickly regenerate functional legs from the coxa-trochanter joint following autotomy (Figure 1, B). This functionality is crucial to their survival, as the spiders rely on the limbs’ sensory-motor abilities for weaving their geometrically complex webs, which are intricately designed to facilitate capture of prey (Reed et al 1965).
Regeneration is understood to be an ancestral trait in arachnid species (Goss, 1992) and although some lineages have lost this ability (Vollrath, 1990), the evolutionary relationships giving rise to these differences remain unclear. Research into the development of closely related spider species, with and without regenerative capabilities, offers potential insight into the changes that have led to the loss of this trait. Further, as outlined in Karl Ernst von Baer’s Laws of Embryology (Wanninger, 2015), studying the early development of a species offers essential contributions to the uncovering of evolutionary patterns and relationships of characteristics.
Figure 1. Fourth Instar Female Araneus diadematus (A) Overview of specimen with regenerated, left first walking leg (L1). (B) The coxa-trochanter joint.
The aim of my project was to improve understanding of the regeneration walking legs in the Araneus diadematus. Initially my Objectives comprised:
Generating a time series to document the species’ post-embryonic development
Comparing the regeneration of the first walking legs between different instars
Analysing the leg transcriptome of an A. diadematus leg, to compare with a species unable to regenerate lost limbs – Parasteatoda tepidarioum.
Due to impediments imposed by Covid19, in addition to further challenges thrown up by the unpredictable nature of scientific research, the outcomes and overall procedure of my project ultimately deviated significantly from the parameters of my original aims. Covid regulations limited my lab access early enough in the year, and as a result, I was unable to accurately document the beginning of the time series for the species, obstructing my first aim.
However, acquisition of A. diadematus embryos, contributed an additional branch to the project and supplemented my first objective. Embryos were frozen at different times in development, prior to being peeled, DAPI stained, and microscopically imaged (Figure 2). This contributed to the creation of a partial embryonic time-series, in place of the initially intended post-embryonic time-series.
The examination and comparison of the imaged A. diadematus embryos, with pre-existing images of the P. tepidarioum (Mittman and Wolff 2012) also presents the opportunity to identify significant disparities within the development of the two species, with the potential to propose relevant evolutionary relationships of regeneration, signposted through embryonic development.
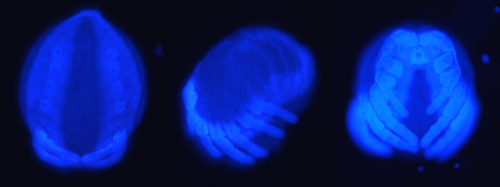
Figure 2: The Prosomal, Lateral and Frontal View (left to right) of a DAPI Stained Araneus diadematus embryo
The findings from the second objective, analyzing and comparing the regeneration and emergent legs (Figure 3) of individuals within the third and fourth instar, prompted a secondary investigation, examining the effects and implications of the leg regeneration of individuals within the fifth instar. Throughout these analyses, I thoroughly enjoyed the weekly imaging and recording of my experimental subjects’ regenerative development, but nothing quite compared to the excitement of discovering the emergence of a new regenerated leg on my visits to the lab’s spider room first thing in the morning.
Figure 3: The Regenerated Left, First Walking Leg of a Third Instar Female, 21 Days (and 1 molt) After Leg Loss
The final objective (obtaining, analyzing, and comparing the A. diadematus leg transcriptome) is still ongoing and I am learning a lot about the patience required for bioinformatics!
My experiences in the lab have emphasized the fluid and unpredictable nature of scientific study, reinforcing the importance of patience, open mindedness, and flexibility. The Gurdon Summer Studentship has taught me so much that I would not otherwise have had access to in the ordinary course of my undergraduate program. I’ve been involved in lab meetings and journal clubs, worked alongside an amazing group of PhD students, and gained experience using cutting-edge equipment and techniques. I’ve developed lab skills, learned specialized spider husbandry techniques, and grown as a scientist, thanks to the lab team I have been privileged to be a small part of.
I did not underestimate the incredible opportunity being afforded to me and was excited before the project started, but I could not have imagined just how much I would enjoy the process. The unpredictable nature of the field of developmental biology has made for an exciting summer and I owe an enormous debt of gratitude to the BDSB. Further I am beyond grateful to Alistair McGregor and to Anna Schönauer for this incredible opportunity and for their support, encouragement, and training. Their passion for their subject is infectious and would inspire anyone to study further in the field of developmental biology!
References
Goss, R. J. (1992) “The Evolution of Regeneration: Adaptive or Inherent?,” Journal of theoretical biology, 159(2), pp. 241–60.
Lai, A. G. and Aboobaker, A. A. (2018) “Evoregen in Animals: Time to Uncover Deep Conservation or Convergence of Adult Stem Cell Evolution and Regenerative Processes,” Developmental Biology, 433(2), pp. 118–131. doi: 10.1016/j.ydbio.2017.10.010.
Mittmann, B. and Wolff, C. (2012) “Embryonic Development and Staging of the Cobweb Spider Parasteatoda Tepidariorum C. L. Koch, 1841 (syn.: Achaearanea Tepidariorum; Araneomorphae; Theridiidae),” Development genes and evolution, 222(4), pp. 189–216. doi: 10.1007/s00427-012-0401-0.
Reed, C. F. Witt, P. N. and Jones, R. L. (1965) “The Measuring Function of the First Legs of Araneus Diadematus Cl,” Behaviour, 25(1-2), pp. 98–119.
Vollrath, F. (1990) “Leg regeneration in web spiders and its implications for orb weaver phylogeny”. Basel (Suiza): Zoologisches Institute.
Wanninger, A. (2015) “Evolutionary developmental biology of invertebrates”. Wien: Springer (Online access: Springer (t). doi: 10.1007/978-3-7091-1868-9.
In our final SciArt profile of 2021 we meet Jessica Richardson, a final year PhD student in Kate Poole’s group at the University of New South Wales, Sydney. In addition to using her artistic skills in her lab work, Jessica works as a freelance illustrator and writer.You can find Jessica on Twitter and LinkedIn.
Where are you originally from and what do you work on now?
I grew up and still live in Sydney, Australia, where I completed my Bachelor’s degree in Neuroscience and Physiology. I am currently in the final year of my PhD in Physiology and Pharmacology at UNSW Sydney working in Associate Professor Kate Poole’s lab. For my PhD project, I am investigating how cells can sense mechanical forces using specialised proteins known as mechanically activated ion channels.
Outside of my research, I am a freelance illustrator and writer and am currently working on various projects, including contributing writing and drawings for high school science textbooks, as well as creating promotional material for commercial organisations in the STEMM space.
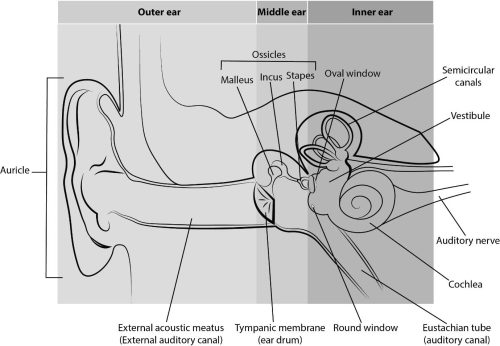
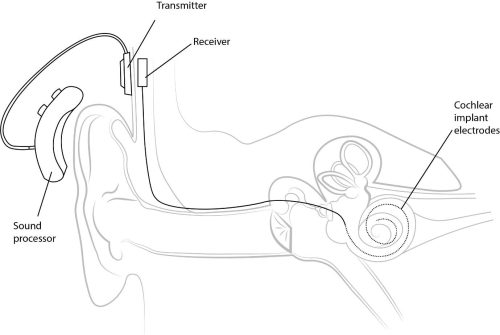
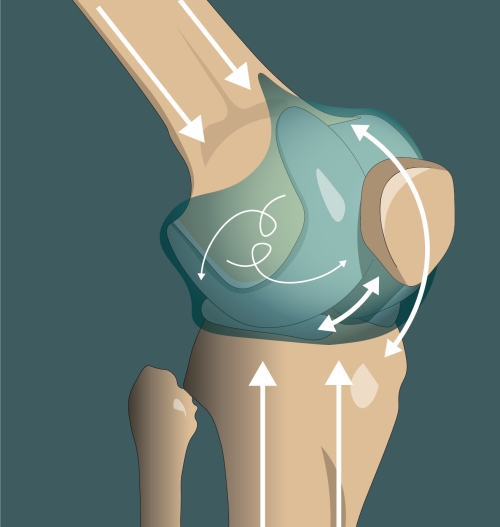
Left: Ear Anatomy – Anatomy of the human ear Middle: Cochlear Implant – Basic diagram of the components of a cochlear implant Right: Knee Joint – Illustration of the forces acting on cartilage within the knee joint, including compressive forces from weight bearing, shear forces of articulating surfaces gliding over one another, and movements of the synovial fluid within the joint capsule.
Were you always going to be a scientist?
I have always had a passion for science since childhood and was the annoying child that constantly asked ‘why?’ and ‘how does that work?’, so you could say so. However, my journey towards becoming a scientist hasn’t been a linear path! I’ve always struggled to choose between pursuing a career in STEMM or pursuing one in the creative industries. For a long time during high school, I really wanted to be creative writer and/or a comic artist. At some point, my love for science won out and I decided to pursue a degree in neuroscience. Neuroscience had always fascinated me as a subject, especially regarding how we sense the world around us and how that differs so greatly from person to person.
But that wasn’t the end of the story. Early during my degree, my university made a double-degree available: a Bachelor of Advanced Science alongside a Bachelor of Fine Arts. At around the same time, I also was introduced to the possibility of a career in science illustration which would combine two of my passions, so I immediately transferred into this degree.
Unfortunately, the Art and Design school was on a different campus from the main campus and trying to run across town between my lab tutorials and my art classes with my huge art book (and supplies) and safety glasses still on was too much. I made the sad decision to transfer to science only, knowing that while I could always find the time to make art on my own while completing my science degree, the reverse would not be true.
Ion Channels at the Cell-Substrate Interface – Illustration of the interface between a cell and its surrounding microenvironment including proteins which may be important for cellular sensing of mechanical forces. This diagram features the cytoskeleton inside the cell (bottom right), proteins such as integrins and mechanically activated ion channels within the cell membrane, and the extracellular matrix outside of the cell (top left).
And what about art – have you always enjoyed it?
I have always enjoyed making art since I was a very young child. I was always doodling in class in school and always had to have a sketch book on hand. Throughout primary school and high school, I experimented a lot with different mediums, and to this day, while I’ve found my preferred mediums, I still like to dabble occasionally in different techniques.
T-cell – Illustration of a T-cell (or T-lymphocyte)
What or who are your most important artistic influences?
Generally speaking, I have always been influenced by genres that use monochromatic black and white colours and heavy shadowing. Think the stark look of noir films and black and white comics, as well as high contrast black and white photography. Looking back to my childhood dreams of being a comic book artist, I was definitely inspired by comic book artists like Frank Miller and James O’Barr, and still am, in recent years, by newer comic artists like Nicola Scott and Fiona Staples. I definitely incorporate these influences into my science illustration work and figures, and often try to create a cartoon-like feel. However, especially for illustrating cellular environments at a molecular level, I am heavily inspired by David Goodsell, who creates absolutely stunning works of art to visualise such complex concepts about the microscopic world.
My absolute biggest artistic influence and inspiration, however, has to be my oldest sister, Sara, who first got me interested in drawing and gave me my very first lessons on proportions, perspective, shading, and drawing human faces. Looking through old sketchbooks from when I was a child, there are plenty of beautiful drawings by her and many more poor emulations from me. I really have her to thank for my artistic endeavours.
Bubbles – Illustration of my niece blowing bubbles
How do you make your art?
My favourite things to draw with are ink and fine-tipped pens. I like to use stippling and heavy shadows and contrast in my artworks and using a lot of black ink really gets this job done well! When making my more formal science illustrations and figures, however I like to use a graphics tablet and Adobe Illustrator and Photoshop.
Left: Resonance – Drawing of the cardiovascular system, inspired by magnetic resonance imaging and other medical imaging techniques that allow us to see into the living human body. Right: Blue Ringed Octopus – Illustration of a blue ringed octopus
Does your art influence your science at all, or are they separate worlds?
I didn’t realise how much my art influenced my science until I began to present my scientific work. Trying to explain my results and ideas, and more importantly trying to communicate them quickly during my presentations and in publications made me focus on making figures and diagrams that could easily convey complex information. While it is a cliché that ‘a picture is worth a thousand words’, I strongly believe this to be true, and that really helps when you have a set word limit!
While it is a cliché that ‘a picture is worth a thousand words’, I strongly believe this to be true, and that really helps when you have a set word limit!
I think that the mental and physical act of trying to represent complicated ideas visually can teach you a lot about what is important in your own work and hence where to go next in your experiments. For instance, even having to ask simple questions like “what can I draw in a high level of detail? What aspects can I only draw very vaguely?” can give insight into where there are holes in your own knowledge, or even what might be unknown in the field and waiting to be discovered.
Freeze – Illustration of a cryo-electron microscopy sample investigating bacterial chaperonin proteins.
What are you thinking of working on next?
I am currently finishing the final stages of my PhD and am really excited to see what’s next for me in research. I hope to continue making illustrations to communicate my own research (and others!). In the future, I’d really love to start a science web comic as well as design online visual educational tools for learning STEMM subjects. I have long-term ambitions about creating or joining a consultancy for science communication, helping other scientists to create figures and drawings which are not only useful and accurate, but visually appealing too.
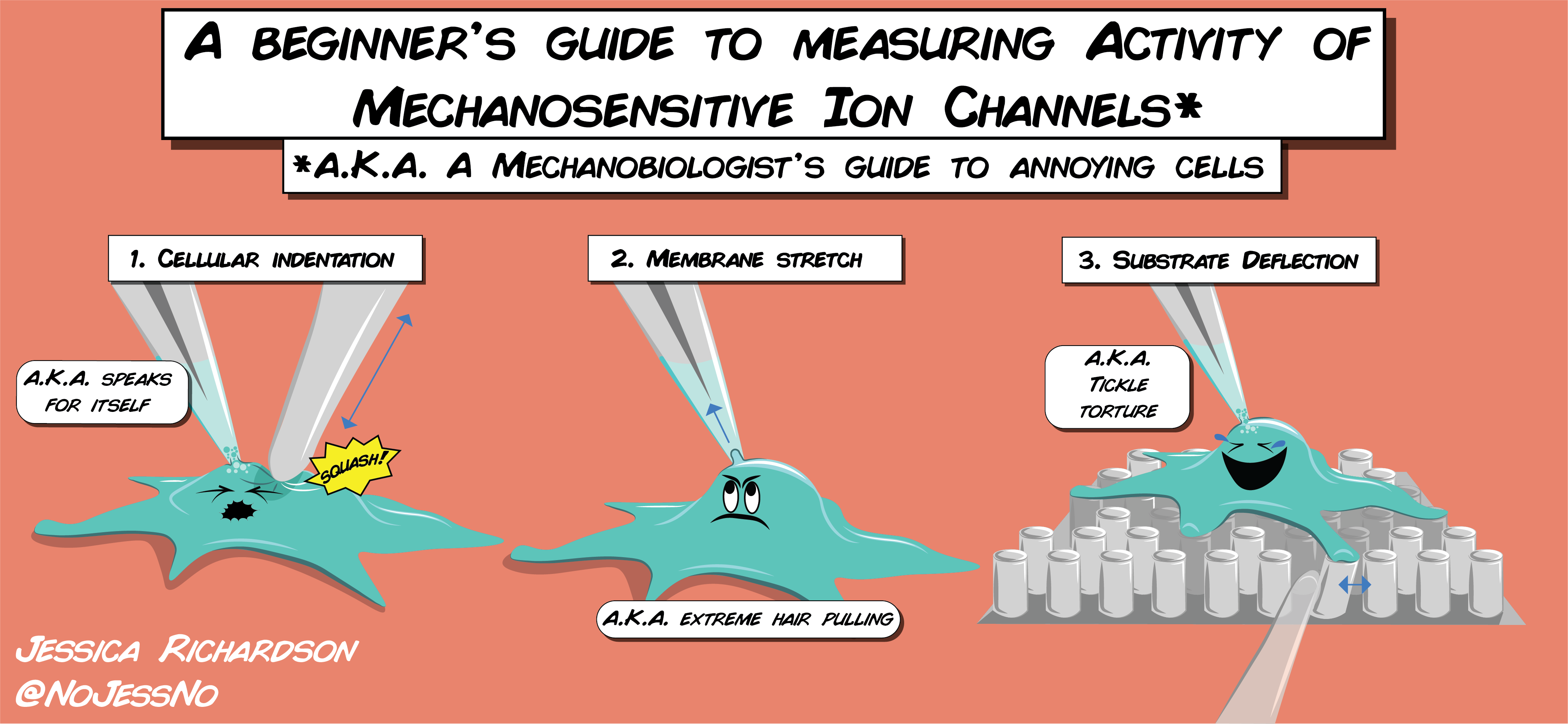
A Beginner’s Guide – A beginner’s guide to measuring mechanically activated ion channel activity in cells. Many mechanical stimuli can be applied experimentally to cells including: 1) cellular indentation, where the cell is physically compressed using a blunt glass probe 2) high speed pressure clamp which applies membrane stretch by applying pressure changes to the cell membrane via a micropipette 3) substrate deflections, where cells can be cultured onto a bed of elastic cylindrical pili which can be individually deflected to apply a mechanical stimulus at the base of cells All of these techniques can be combined with single-cell patch clamp electrophysiology to measure electrical activity in cells.Lab Waste – Poster illustration for a lab campaign to raise awareness about the environmental impact of life science research and areas of focus to decrease our footprint.
Thanks to Jessica and all the other SciArtists we have featured so far.We’re looking for new people to feature in this series – whatever kind of art you do, from sculpture to embroidery to music to drawing, if you want to share it with the community just email thenode@biologists.com (nominations are also welcome!)
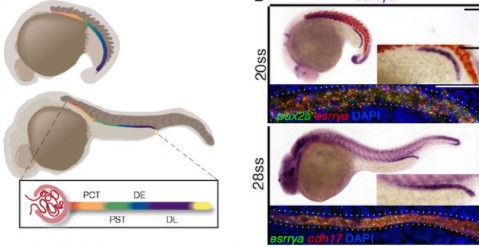
On Wednesday 8 December, Development hosted three talks to celebrate the 25th anniversary of our zebrafish special issue.
Below you’ll find each of the talks, plus reflections on the impact of the special issue, 25 years on from its publication, from our guest chair Alex Schier.
Alex Schier (Harvard University and Biozentrum, University of Basel) Reflections on the 25th anniversary of the ‘zebrafish issue’
Caren Norden (Instituto Gulbenkian Ciência) ‘Lamination in the vertebrate retina: An interplay of diverse modes of cell migration’
You can read the preprint of the research presented by Caren here.
Frieda Leesch (Research Institute of Molecular Pathology) ‘A molecular network of conserved factors keeps ribosomes dormant in the egg’
The research presented by Frieda is available as a preprint on biorxiv
Brad Cairns (Huntsman Cancer Institute) ‘Maternally-inherited anti-sense piRNAs antagonize transposon expression in zebrafish and medaka embryos’
You can read the preprint of the research presented by Brad here.
Doing great science depends on teamwork, whether this is within the lab or in collaboration with other labs. However, sometimes the resources that support our work can be overlooked. In our new series, ‘Featured Resource’, we aim to shine a light on these unsung heroes of the science world. For our third article, Yvonne Bradford and Leyla Ruzicka tell us about the work of ZFIN. Read on to find out a little about the history of ZFIN, the people running the site and what resources are available.
When was ZFIN established and how has it evolved?
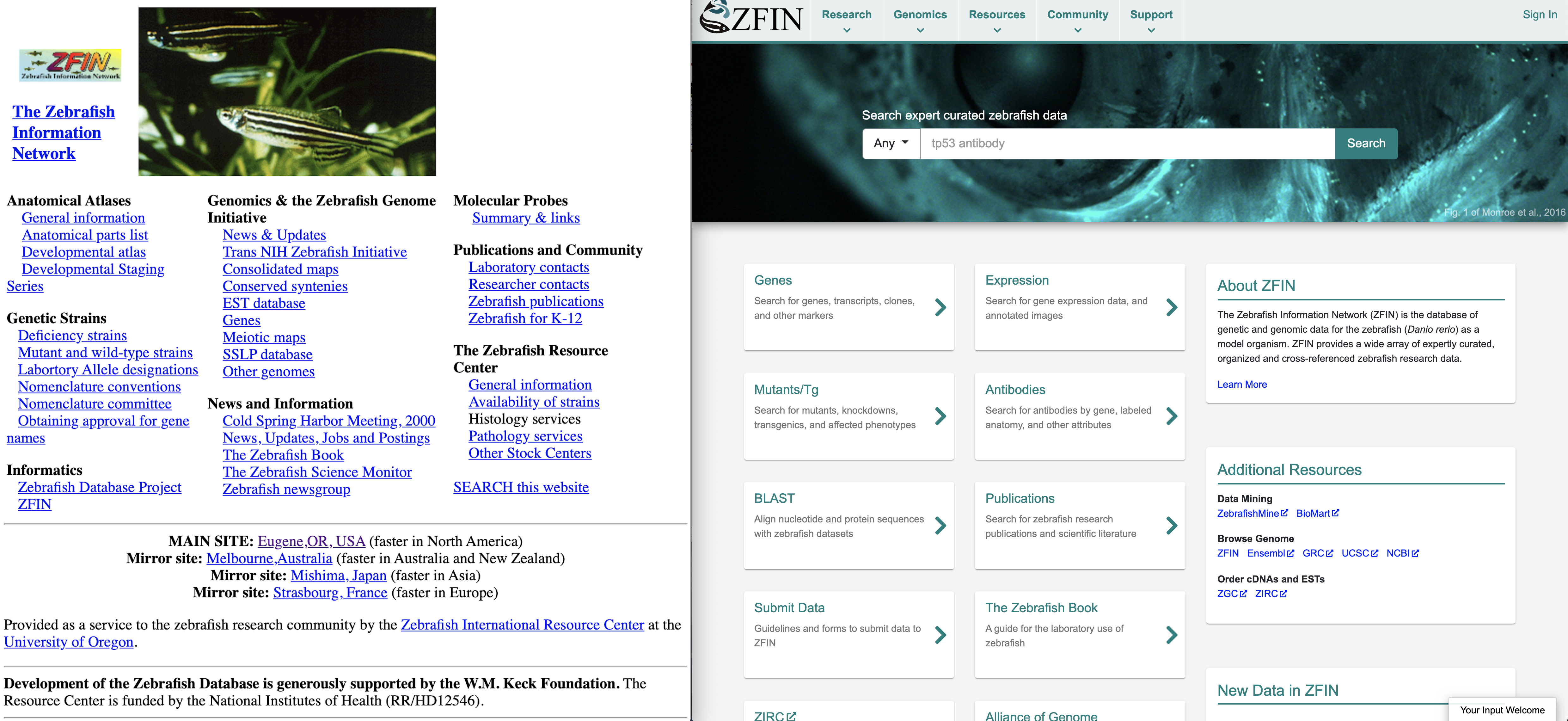
The Zebrafish Information Network (ZFIN) was established by Monte Westerfield in 1994 following the first international Zebrafish research meeting at Cold Spring Harbor. In its infancy, ZFIN played a large role as a community contact resource for zebrafish researchers and students. By 1997 ZFIN curators were establishing nomenclature guidelines for genes and mutant lines, as well as curating mutant line information from scientific publications1. Leap to 2003 and ZFIN had established cross links and integration of ZFIN data with other genomic resources including GenBank, Vega, UniProt and Ensembl, and curators were annotating additional data types that included gene expression, orthology, and Gene Ontology data2. Over the next several decades, ZFIN curators continued to expand the data supported at ZFIN to include complex genotypes, transgenic lines, phenotypes, morpholinos, CRISPRs, TALENs, antibodies, and transcripts34567. Over the past 27 years, ZFIN has grown into a mature knowledgebase for zebrafish researchers providing information about genes and alleles, gene expression and phenotype data, human disease associations and models, Gene Ontology data for gene function, orthology, reagent information and protocol wikis, BLAST and Genome Viewers, as well as deep integration with genetic and genomic data service providers. Throughout ZFIN’s history, serving the research community has been central to the development of the resource. ZFIN has a core aim of providing accurate, accessible and interoperable data to researchers, students, bioinformaticians, and other online databases to facilitate the use of Danio rerio as a model organism to understand developmental biology, genetics, genomics, phenomics, and ultimately human biology.
ZFIN has grown and changed over the years. Left panel is a screenshot of the ZFIN home page circa 2000. Right panel is the current ZFIN home page.
Who are the people behind the resource?
ZFIN is located at the University of Oregon in Eugene, Oregon. The ZFIN team consists of scientific curators who hold a PhD or MSc and have expertise in genetics, developmental, cellular, molecular, and evolutionary biology, as well as software developers skilled in Java web architecture, PostgreSQL database development, and many modern coding languages to implement user interfaces. Although the following list represents ZFIN staff as of 2021, many prior staff members significantly contributed to the success of ZFIN over the years.
Principal Investigator – Monte Westerfield
Project Manager – Yvonne Bradford
Scientific Curation Team:
David Fashena
Ken Frazer
Doug Howe
Holly Paddock
Sridhar Ramachandran
Leyla Ruzicka
Amy Singer
Wei-Chia Tseng
Ceri Van Slyke
Software Development Team:
Ryan Martin
Christian Pich
Ryan Taylor
Literature Acquisition Assistant – Kaia Fullmer
Where does funding come from?
Initially, ZFIN was funded by the W.M Keck Foundation and the National Science Foundation. In 1998, Dr. Westerfield was awarded NIH funding for the Zebrafish International Resource Center (ZIRC) that included database support for ZFIN. Since 2002, ZFIN has been independently funded by the National Human Genome Research Institute of the National Institutes of Health.
What is available for researchers?
ZFIN hosts a diverse collection of data, as well as genomic and community resources. ZFIN provides expertly curated data on zebrafish, including information on genes, alleles, transgenic lines, human disease models, gene expression, mutant phenotypes, gene function, orthology, publications, researchers, labs, and reagents. ZFIN has data-specific web pages for genes, alleles, fish, antibodies, constructs, morpholinos, CRISPRs, TALENs, clones, publications, and ontology terms. Researchers can search for data using the single box search on the home page (https://zfin.org/) with results returned in the single box search interface, which allows for easy refinement of results using filters. ZFIN data are also available for download (https://zfin.org/downloads). ZFIN is the nomenclature authority for zebrafish genes, alleles, and transgenic lines. ZFIN provides nomenclature support to the community through the ZFIN Nomenclature Coordinator who works with the Zebrafish Nomenclature Committee, which is composed of active zebrafish researchers, to approve names and symbols for gene loci and transgenic lines. Researchers can contact the ZFIN Nomenclature Coordinator at nomenclature@zfin.org.
ZFIN provides access to genomic resources through the Genomics menu on the Home Page. Researchers can access BLAST resources and genome browsers at ZFIN, NCBI, UCSC, and Ensembl. In addition, the ZFIN gene page has genome browser images and links to sequence analysis tools in the sequences and transcripts sections.
Researchers can access community resources from the Resources and Community menus at the top of the ZFIN Home Page. The Resources menu provides links to The Zebrafish Book, Anatomy Atlases, Zebrafish Resource Centers, and community wikis. ZFIN hosts the zebrafish research community wiki that has sections for protocols and antibodies. The protocols section provides an area where zebrafish researchers can share experimental protocols and tips covering a range of topics from general zebrafish care to molecular and behavioural methods, and the antibodies section focuses on antibody usage notes and protocols. The Community menu offers researchers access to zebrafish News, Job postings, and Meeting announcements, as well as links to search for researchers, labs, and companies.
How can the community contribute to ZFIN?
In addition to annotating zebrafish research publications, ZFIN accepts direct submissions from users. From the Support menu at the top of the home page, under ‘Using ZFIN’, there is a link to submit data to ZFIN. The link leads to a page that has links to the various ways users can submit data directly. To help researchers submit data, a guide that outlines the minimal set of information for data that are submitted for inclusion in ZFIN is available in the Methods in Cell Biology chapter: A Scientist’s guide for submitting data to ZFIN8.
How can the community help ZFIN?
The research community can help ZFIN in many ways:
Respond to ZFIN surveys. ZFIN sends out user surveys to registered users yearly. The surveys determine which features will be developed at ZFIN
Keep your person/lab/company pages in ZFIN up to date
Help facilitate the curation of your publications:
Contact ZFIN about gene and allele/transgenic line nomenclature before publication
Make sure data in papers are unambiguous – use identifiers, accession numbers, and approved names whenever possible for genes, alleles, and reagents.
Respond to requests for information by ZFIN curators on details in papers including genes, alleles, transgenes, sequence information, and reagents.
What are the most used features on ZFIN?
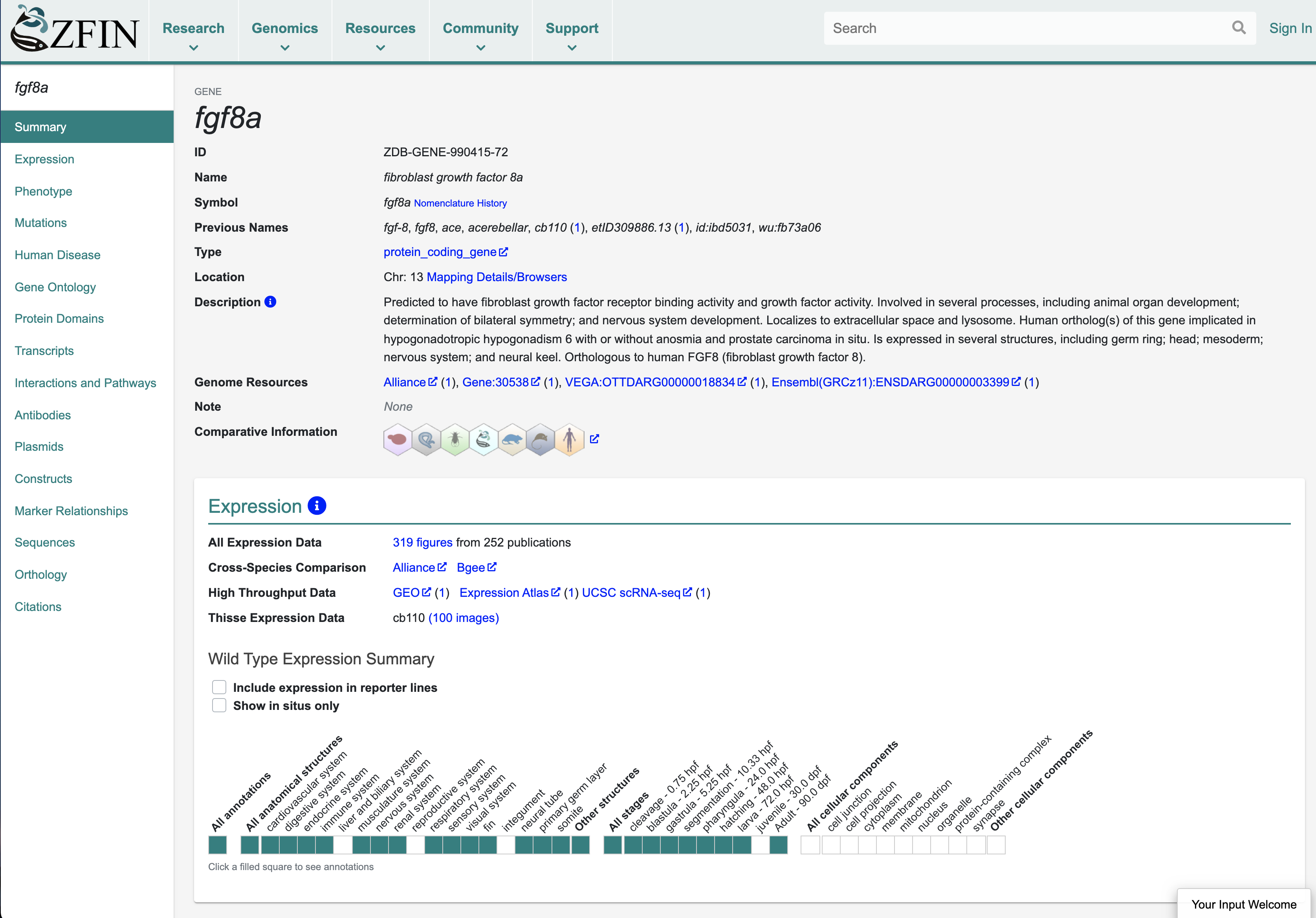
Gene pages are the most highly accessed pages at ZFIN. The gene page integrates all data pertinent to a gene and is a hub of information for researchers. Other ZFIN resources that are highly accessed include gene expression and phenotype data, publications and associated images, and search interfaces. ZFIN person and lab pages are also very popular, as is The Zebrafish Book9, the classic handbook for zebrafish research.
The gene page serves as a hub of information and integrates all data for a gene. Here is a screenshot of a portion of the fgf8a gene page.
Any new developments that you can recommend, which researchers might be less aware of?
ZFIN is a founding member of the Alliance of Genome Resources (The Alliance), a consortium of six Model Organism databases (Mouse Genome Database, Rat Genome Database, Saccharomyces Genome Database, WormBase, FlyBase, and ZFIN) and the Gene Ontology database. The Alliance aims to develop and maintain sustainable genome information resources that facilitate the use of model organisms in understanding the genetic and genomic basis of human biology and disease10. Follow the “Alliance” links from ZFIN gene pages to explore comparative data on Orthology, Human Disease, Gene Ontology, and Gene Expression, plus details about Variants and Phenotypes. The Alliance also provides download files and APIs for programmatic access to model organism data.
References
1. Westerfield M, Doerry E, Kirkpatrick A, Driever W, Douglas S. An on-line database for zebrafish development and genetics research. Seminars in cell & developmental biology. 1997;8(5):477–88. doi:10.1006/scdb.1997.0173
2. Sprague J, Clements D, Conlin T, Edwards P, Frazer K, Schaper K, Segerdell E, Song P, Sprunger B, Westerfield M. The Zebrafish Information Network (ZFIN): the zebrafish model organism database. Nucleic acids research. 2003;31(1):241–243. doi:10.1093/NAR/GKG027
3. Sprague J, Bayraktaroglu L, Bradford Y, Conlin T, Dunn N, Fashena D, Frazer K, Haendel M, Howe DGDG, Knight J, et al. The Zebrafish Information Network: the zebrafish model organism database provides expanded support for genotypes and phenotypes. Nucleic acids research. 2008;36(Database issue):D768-72. doi:10.1093/nar/gkm956
4. Bradford YM, Conlin T, Dunn NA, Fashena D, Frazer K, Howe DG, Knight J, Mani P, Martin R, Moxon SAT, et al. {ZFIN:} enhancements and updates to the zebrafish model organism database. Nucleic Acids Res. 2011;39(Database-Issue):822–829. doi:10.1093/nar/gkq1077
5. Howe DG, Bradford YM, Conlin T, Eagle AE, Fashena D, Frazer K, Knight J, Mani P, Martin R, Moxon SAT, et al. ZFIN, the Zebrafish Model Organism Database: increased support for mutants and transgenics. Nucleic Acids Research . 2013;41(Database issue):D854-60. doi:10.1093/nar/gks938
6. Ruzicka L, Bradford YM, Frazer K, Howe DG, Paddock H, Ramachandran S, Singer A, Toro S, Van Slyke CE, Eagle AE, et al. ZFIN, The zebrafish model organism database: Updates and new directions. Genesis. 2015;53(8). doi:10.1002/dvg.22868
7. Howe DG, Ramachandran S, Bradford YM, Fashena D, Toro S, Eagle A, Frazer K, Kalita P, Mani P, Martin R, et al. The zebrafish information network: Major gene page and home page updates. Nucleic Acids Research. 2021;49(D1):D1058–D1064. doi:10.1093/nar/gkaa1010
8. Howe DGG, Bradford YMM, Eagle A, Fashena D, Frazer K, Kalita P, Mani P, Martin R, Moxon STT, Paddock H, et al. A scientist’s guide for submitting data to ZFIN. Methods in Cell Biology. 2016;135:451–481. doi:10.1016/bs.mcb.2016.04.010
9. Westerfield M. The zebrafish book: a guide for the laboratory use of zebrafish (Danio rerio). 4th ed. Eugene, OR: University of Oregon Press; 2000.
10. Bult CJ, Blake JA, Calvi BR, Cherry JM, DiFrancesco V, Fullem R, Howe KL, Kaufman T, Mungall C, Perrimon N, et al. The alliance of genome resources: Building a modern data ecosystem for model organism databases. Genetics. 2019;213(4):1189–1196. doi:10.1534/genetics.119.302523
A comprehensive series of temporal transcription factors in the fly visual system Nikolaos Konstantinides, Anthony M. Rossi, Aristides Escobar, Liébaut Dudragne, Yen-Chung Chen, Thinh Tran, Azalia Martinez Jaimes, Mehmet Neset Özel, Félix Simon, Zhiping Shao, Nadejda M. Tsankova, John F. Fullard, Uwe Walldorf, Panos Roussos, Claude Desplan
Competition for endothelial cell polarity drives vascular morphogenesis Pedro Barbacena, Maria Dominguez-Cejudo, Catarina G. Fonseca, Manuel Gómez-González, Laura M. Faure, Georgia Zarkada, Andreia A. Pena, Anna Pezzarossa, Daniela Ramalho, Ylenia Giarratano, Marie Ouarné, David Barata, Isabela Fortunato, Lenka H. Misiková, Ian Mauldin, Yulia Carvalho, Xavier Trepat, Pere Roca-Cusachs, Anne Eichmann, Miguel O. Bernabeu, Cláudio A. Franco
Pan-primate DNA methylation clocks Steve Horvath, Amin Haghani, Joseph A. Zoller, Ake T. Lu, Jason Ernst, Matteo Pellegrini, Anna J. Jasinska, Julie A. Mattison, Adam B. Salmon, Ken Raj, Markus Horvath, Kimberly C. Paul, Beate R. Ritz, Todd R. Robeck, Maria Spriggs, Erin E. Ehmke, Susan Jenkins, Cun Li, Peter W. Nathanielsz
Transgenerational impact of aberrant inflammation in rat pregnancy Takafumi Ushida, Tiziana Cotechini, Nicole Protopappas, Aline Atallah, Charlotte Collyer, Shannyn K. Macdonald-Goodfellow, M. Yat Tse, Louise M. Winn, Stephen C. Pang, Michael A. Adams, Maha Othman, Tomomi Kotani, Hiroaki Kajiyama, Charles H. Graham
Transcription factor induction of vascular blood stem cell niches in vivo Elliott J. Hagedorn, Julie R. Perlin, Rebecca J. Freeman, Samuel J. Wattrus, Tianxiao Han, Clara Mao, Ji Wook Kim, Inés Fernández-Maestre, Madeleine L. Daily, Christopher D’Amato, Michael J. Fairchild, Raquel Riquelme, Brian Li, Dana A.V.E. Ragoonanan, Khaliun Enkhbayar, Emily L. Henault, Helen G. Wang, Shelby E. Redfield, Samantha H. Collins, Asher Lichtig, Song Yang, Yi Zhou, Balvir Kunar, Jesus Maria Gomez-Salinero, Thanh T. Dinh, Junliang Pan, Karoline Holler, Henry A. Feldman, Eugene C. Butcher, Alexander van Oudenaarden, Shahin Rafii, J. Philipp Junker, Leonard I. Zon
Circadian regulation of lung repair and regeneration Amruta Naik, Kaitlyn Forrest, Yasmine Issah, Utham Valekunja, Akhilesh B Reddy, Elizabeth Hennessy, Thomas S. Brooks, Apoorva Babu, Mike Morley, Gregory R. Grant, Garret A. FitzGerald, Amita Sehgal, G. Scott Worthen, David B. Frank, Edward E Morrisey, Shaon Sengupta
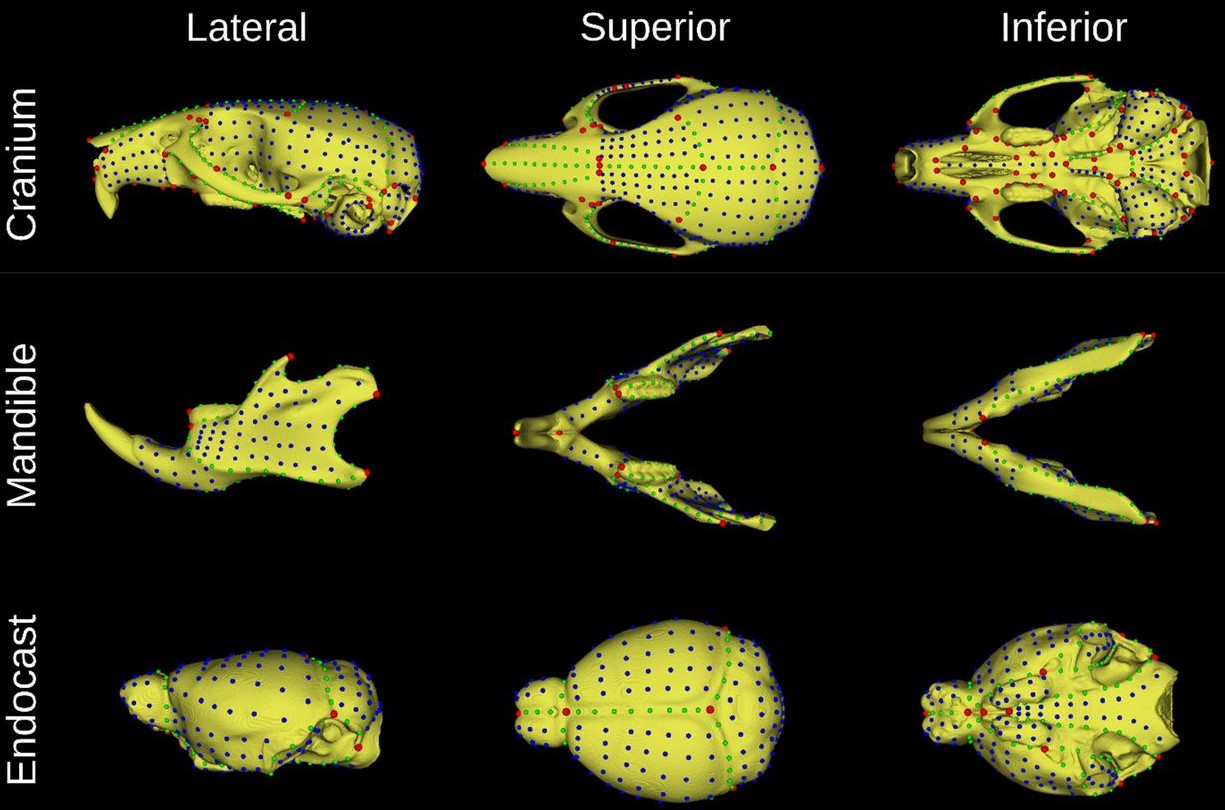
Mouse adult reference skull atlas from Devine, et al.
MusMorph, a database of standardized mouse morphology data for morphometric meta-analyses Jay Devine, Marta Vidal-García, Wei Liu, Amanda Neves, Lucas D. Lo Vercio, Rebecca M. Green, Heather A. Richbourg, Marta Marchini, Colton M. Unger, Audrey C. Nickle, Bethany Radford, Nathan M. Young, Paula N. Gonzalez, Robert E. Schuler, Alejandro Bugacov, Campbell Rolian, Christopher J. Percival, Trevor Williams, Lee Niswander, Anne L. Calof, Arthur D. Lander, Axel Visel, Frank R. Jirik, James M. Cheverud, Ophir Klein, Ramon Y. Birnbaum, Amy E. Merrill, Rebecca R. Ackermann, Daniel Graf, Myriam Hemberger, Wendy Dean, Nils D. Forkert, Stephen A. Murray, Henrik Westerberg, Ralph S. Marcucio, Benedikt Hallgrímsson
Mark your diaries and get your applications in for the upcoming Workshops run by The Company of Biologists. Three Workshops that are of particular interest to developmental and stem cell biologists are currently accepting applications for funded places from early-career researchers:
Want to know more about the format of our Workshops, and how and why you should apply? Our Science Communications Officer, Laura Hankins explains more here.
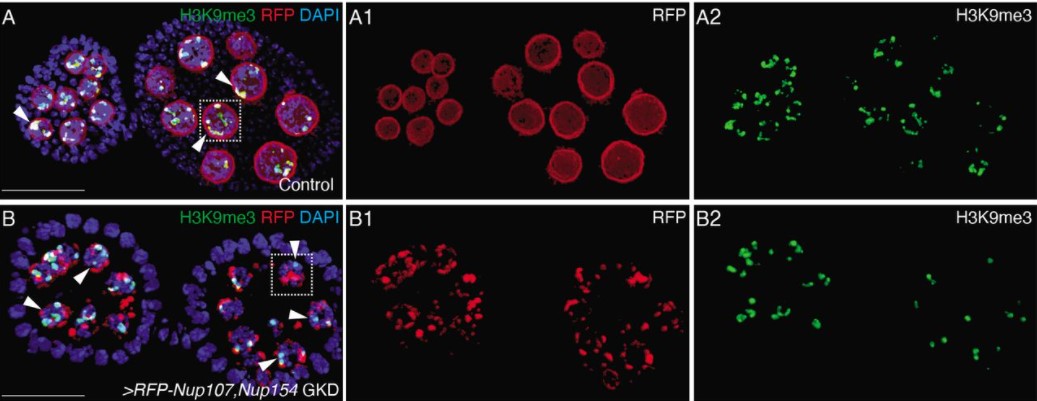
This project started in a somewhat roundabout way. Our lab (the Gross lab) has focused for many years on development of the retina, lens and optic cup, eye evolution, and more recently RPE regeneration. Several years ago, Taka Kuwajima, who is an expert in retinal ganglion cell (RGC) biology, and particularly axon regeneration, was hired into our Department. We share lab space and have a joint weekly meeting, and had wanted to work together on a project for some time but nothing quite got started. Then, in 2019, we had a fabulous MD/Ph.D. student, Si Chen, begin to work with us through a joint program between the University of Pittsburgh and Xiangya Hospital/Central South University in China. Si was interested in glaucoma, so we thought about what sort of RGC-focused project we could do together that would also finally get Taka and I collaborating.
RGCs in mice die rapidly after injury. The same is true in humans, and neither system can regenerate lost cells or axons if they are damaged. This is what leads to the loss of vision in glaucoma – once the RGC axons comprising the optic nerve are damaged, the cells begin to die and vision progressively deteriorates. Zebrafish have remarkable abilities to regenerate lost or damaged cells and tissues, and this is true for RGCs; when the axons of the optic nerve are damaged or even transected, zebrafish regenerate these projections and restore connections and function. Several labs are doing some really exciting work on axon regeneration including Lieve Moons, Ava Udvadia, and Dan Goldman. While the mechanism that facilitates RGC axon regeneration is a fascinating topic, what really interested us was a slightly different facet of zebrafish RGC biology coming from earlier studies where it was reported that ~75% of zebrafish RGCs were preserved after severe optic nerve injury, even to 7-weeks post-injury (Zou et al., 2013). Again, in mammals, the RGCs die rapidly after injury and to our knowledge, no one had followed up this observation that zebrafish RGCs stayed alive after completely severing their axons. We thought this would be a great project to bring our labs together and a great in vivo system to possibly discover something new about neuroprotection. Moreover, since there are no effective treatments to preserve RGCs in glaucoma, we thought this project might also have some exciting translational potential if we could discover genes and pathways that facilitated RGC survival after injury.
To get started, Si first repeated the experiments of Zou et al. (2013), observing exactly what they saw – RGCs did indeed survive, even after she completely severed the optic nerve! These are tough experiments and Si has amazing skills that enabled her to develop the surgical technique and rigorously reproduce the findings. With the model in hand, Si was then able to isolate RGCs and perform RNA-Seq to profile changes in gene expression after injury. This is a great hypothesis-generating experiment and indeed, she identified many genes and pathways that were altered by the injury. She decided to focus on Jak/Stat signaling as a proof-of-concept that this approach could yield interesting neuroprotective factors. Her results were striking; blocking Jak activity compromised RGC survival after injury. What was particularly exciting was how the project then dovetailed nicely with another interest in the lab – the role of immune responses during injury and regeneration. We had recently been studying innate immune responses during RPE regeneration (Leach et al., 2021) and leveraged some of what we had learned in that system to start to look at RGC survival. Si was able to show that blocking inflammation or depleting macrophages/microglia protected all RGCs after injury. The Jak/Stat pathway is activated in both RGCs and macrophages/microglia after injury, so we don’t yet know if activity is required in one cell type or both, and this is something for which we’d like to develop tools to answer.
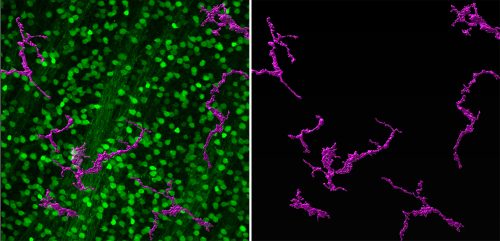
Zebrafish RGCs (labeled in green with isl2b:GFP) and macrophages/microglia (labeled in magenta with 4c4).
We’re excited to study some of the other genes and pathways identified in the screen and we’re also curious to see if there are more sensitive or resilient RGC subtypes. Our research will be assisted by the beautiful single cell atlas of zebrafish RGCs recently published by the Baier lab (Kölsch et al., 2021) and work from Nick Tran and colleagues, where they have identified resilient subtypes in mouse (Tran et al., 2019). A terrific new postdoc, Ashrifa Ali, has recently joined the lab and she plans to build off of these studies, using the zebrafish model to better understand RGC survival after injury. Hopefully, this will lead to the development of new therapeutics to treat glaucoma.
In the end, this was a really fun project that brought together a fantastic MD/Ph.D. student, Si Chen, with Taka and me to work together on something new for each of us. I love this part of basic science research; we have the best jobs in the world where we can come to work every day, ask exciting open-ended questions and then figure out creative ways to answer them. If you have a good idea and work hard, more likely than not, you will discover something new, and there aren’t many jobs that give this sort of freedom and reward. We have a lot more to do on this project, and certainly there will be a lot of ups and downs along the way, but I’m sure it will be a fun ride.
In our recently published paper1, we show that human stem cells self-organize into blastocyst-like structures, which we term blastoids based on 4 criteria. Because blastoids can be generated in large numbers, can be finely manipulated and recapitulate aspects of endometrial implantation in vitro, they are powerful models to investigate the principles of blastocyst development and implantation. Here I want to tell the backstory of our scientific journey and why I believe it is important for science and medicine.
Understanding human development is one of the new frontiers in developmental biology. Because of the intrinsic difficulties in studying human embryos, the majority of our knowledge has been inferred from model organisms such as mice, frogs, fish, or fruit flies. However, many developmental principles are not conserved and humans have evolved specific features2. Human embryos can be studied when generously donated by couples undergoing an IVF procedure, and this has proved extremely useful3. However, IVF embryos are scarce, not always of top quality, and considerably difficult to work with. On the contrary, embryo models formed from pluripotent stem cells (hPSCs), although they are simplified versions of the embryo, can be formed easily from established cell lines in the lab, generated in large numbers, and are easier to manipulate either physically or genetically. They thus represent an ethical and technical alternative complementing the use of IVF embryos.
Understanding implantation with blastoids
In our lab (Nicolas Rivron, IMBA-Institute for Molecular Biotechnology, Austrian Academy of Sciences), we are interested in understanding early mammalian development and how the embryo implants into the uterus. We are especially studying the formation of the blastocyst (day 4-7 in humans) that generates and pattern tissues to prepare the conceptus for implantation into the uterus. Implantation is the first physical contact between the mother and the embryo and an extremely crucial step towards the sustenance of the pregnancy. Implantation success marks the establishment of a clinical pregnancy as measured by levels of the hormone human chorionic gonadotropin. Of clinical importance, in humans, the steps of blastocyst development and implantation are prone to failure and are thought to account for as much as 75% of pregnancy loss4,5.
Despite its tremendous significance, understanding blastocyst development and implantation has proven to be a challenging endeavor. This is because embryos are available in limited numbers and their interactions with the uterus happens deep inside the womb, therefore is inaccessible and invisible. These limitations in working with IVF embryos and the uterus restrain the possibilities to perform, for example, complex genetic manipulations, biochemistry, as well as high throughput chemical or genetic screens that are at the heart of scientific and biomedical discoveries. To overcome these challenges, our lab has previously developed a model of mouse blastocyst generated solely using stem cells, which we termed blastoid6. Mouse blastoids formed through the combination of cells from lines of mouse embryonic stem cells7 and mouse trophoblast stem cells8. When treated with a defined cocktail of growth factors and/or their modulators, these two stem cell types interact, which triggers their self-organization into structures that are remarkably similar to mouse blastocysts6,9,10. The mouse blastoids was a first complete model of the conceptus and, because it modeled the pre-implantation stage, we were able to transfer them in utero to study implantation. No mice were formed from mouse blastoids until now but blastoids are a scalable and experimentally amenable platform to employ multimodal approaches such as complex genetic manipulations and high-throughput imaging in order to address scientific questions such as principles of embryonic/extraembryonic inductions and self-organization.
2021: Many attempts to model human blastocysts.
The mouse blastoids reported in 2018 triggered interest to form human blastoids and the potential clinical implications multiplied that enthusiasm. Till date, 6 groups independently proposed human blastocyst models, all of which were published in 2021. These 6 methods were developed using at least 4 different states of stem cells as a starting point, and 6 different culture conditions to tentatively trigger the formation of analogs of blastocyst lineages. The output with respect to the stage of the cells that were generated, the level of adequacy with developmental time and sequence, and the efficiency of the formation of structure widely varied. Overall, establishing ways of evaluating the weight of the initial experimental parameters and of defining criteria to assess the cells and structures that are formed will be essential. To our best of understanding, the initial cell state and the combination of molecules used to trigger blastoid formation are the two main parameters that, when combined, allow for modeling blastocyst development and forming the right cells reflecting the blastocyst stage.
Currently, the most powerful way to perform end-point assessments is to combine single cell RNA sequencing (scRNA seq) data of the model with a reference map generated from various staged human embryos (pre-blastocyst, blastocyst, post-implantation stages). Having multiple stages, including the ones before and after the blastocyst stage, is essential to detect potential off-target cells. Fortunately, the last few years have been fruitful in generating high-quality datasets from human embryos including IVF pre-blastocyst and blastocysts11,12, IVF blastocysts further cultured in vitro for up to 14 days13, that mimic aspects of early post-implantation development. Moreover, a very recent study has created an invaluable scSeq dataset from a gastrulation human embryo (Carnegie stage 7)14. Overall, this provides an unprecedented opportunity to generate a high-quality reference meta-map from human embryos in order to evaluate embryo models. In the future, additional layers of readouts (e.g., ATAC seq, Bi-sulfite seq, etc.) would be useful to complement the analysis of the transcriptome alone.
Our own attempt to form human blastoids.
In our study1, we report the generation of human blastoids that, to the best of our knowledge, faithfully and efficiently recapitulate key features of blastocyst development: the sequence and the pace of lineage specification and morphogenesis, which results in the formation of blastocyst-like cells. We measured 97% of blastocyst-like cells based on criterias that we think should be challenged, fine-tuned, and complemented. This reliability in modeling blastocyst development allowed us to subsequently model several aspects of implantation using endometrial organoids. Here we describe the initial cell state and combination of molecules triggering the formation of blastoids, and the end-point criteria that we think are valid to evaluate such models. the human blastocyst.
The right shade of pluripotent stem cells: a secret ingredient to faithful blastoids
In mice, the culture conditions to establish stem cell lines reflective of the blastocyst lineages are relatively well established. For example, stem cells can be captured in a state that reflects the blastocyst-stage epiblast, and that are referred to as 2i/Lif naive PSCs7. Similarly, the blastocyst-stage trophectoderm can be captured in a state that we refer to as trophectoderm stem cells (TESCs, unpublished manuscript)9. This allows these two developmentally matched stem cell types to efficiently self-organize and to form mouse blastoids. Altogether, to form a faithful and thus useful blastoid, it is crucial to begin with stem cells that reflect the blastocyst stage, and that are capable of efficiently forming and organizing analogs of the blastocyst lineages upon exposure to developmentally relevant cues.
In mice, the culture conditions to establish stem cell lines reflective of the blastocyst lineages are relatively well established. For example, stem cells can be captured in a state that reflects the blastocyst-stage epiblast, and that are referred to as 2i/Lif naive Pluripotent Stem Cells (PSCs)7. Similarly, the blastocyst-stage mouse trophectoderm can be captured in a constrained state that we refer to as trophectoderm stem cells (TESCs)9[now published in Cell Stem Cell]. This allows these two developmentally matched stem cell types to efficiently self-organize into mouse blastoids. Altogether, to form a faithful and thus useful blastoid, it is crucial to begin with stem cells that reflect the earliest stage possible, and that are capable of efficiently forming and organizing analogs of the blastocyst lineages upon exposure to developmentally relevant cues.
The human stem cells that best reflect the blastocyst stage have been difficult to capture. The firstly established and most commonly used hPSCs15 are referred to as primed hPSCs and are transcriptionally similar to the epiblast of post-implantation staged embryos (day 12-14) as compared to blastocysts (day 5-7)16. This is typically shown by measuring stage-specific molecular criteria (e.g., genome-wide DNA hypomethylation, X chromosome status17,18) and by comparing the cells transcriptome with reference monkey19 or human datasets11,13,14. However, in the last decade, multiple labs have shown that primed hPSCs cultured with specific signaling pathway inhibitors and activators are capable of resetting their transcriptome, decreasing the methylation of their DNA, and, to some extent, to reactivate silenced X chromosome17,18,20–22. These shades of naive pluripotency reflect the blastocyst epiblast (~ day 6). Other attempts to reset primed hPSCs allowed to capture other states, for example human extended potential stem cells (hEPSCs)23 but, upon re-analysis, these cells display a transcriptome more similar to the post-implantation stage (see analysis in Figure 724 and Figure 325).
The unrestricted potential of naive human pluripotent stem cells
The mouse and human blastocysts comprise 3 lineages: the trophectoderm that mediates implantation and then generates the placenta, the epiblast that forms the majority of the body, and the primitive endoderm that forms the yolk sac and part of the embryonic endoderm. In mice, the specification of the trophectoderm occurs during the transition from a morula to a blastocyst26. On the contrary, studies by the laboratories of Laurent David (INSERM and Université de Nantes, France) and Hilde van de Velde (Vrij Universiteit Brussel) have suggested that the early human blastocyst doesn’t comprise fully specified cells but rather cells that are not yet entirely restricted to a specific lineage1227. In correlation with this observation, the laboratories of Yasuhiro Takashima (CiRA) and of Austin Smith (Exeter University) have shown that naive hPSCs cultured in either PXGL20 or Ti2iLGö18 are capable of forming analogs of the blastocyst-stage trophectoderm28,29. Once again, this potential is in sharp contrast to the already determined potential of mouse PSCs that cannot efficiently form trophoblasts, as assessed in chimeric and in vitro assays30–32. Of note, although several studies have described the potential of primed hPSCs or hEPSCs to differentiate into cells with trophoblast-like properties, this process does not seem efficient, and the obtained cellular populations might be heterogeneous and reflecting a mix of post-implantation trophoblasts and amnion cells (a lineage thought to emerge during the post-implantation period) rather than the trophectoderm (that forms in the blastocysts). However, these attempts to efficiently form trophoblasts from primed hPSCs were very successful in order to learn about the signaling pathways33,34 regulating aspects of trophoblast specification and pinpointing the important plasticity of human cells as compared to mouse cells.
The right signalling activity combination- TGF-β, ERK and Hippo inhibition: other secret ingredient
Therefore, considering the similarities of naive PXGL hPSCs to the blastocyst epiblast and the enhanced plasticity of human cells as compared to mice, we tested the potential of PXGL hPSCs to form human blastocyst-like structures. To do so, we formed a small aggregate of cells (~80-100 um) and provided molecules that were previously found to support either the formation of human trophoblasts (inhibition of TGF-β with A83-0133, ERK with PD0328,29,33,35) or to participate in the maintenance of mouse TESCs (inhibition of Hippo pathways with LPA9), and to maintain naive hPSCs (LIF)21, along with a ROCK inhibitor to enhance cell survival and aggregation. To our surprise, within 96 hours, this culture condition was sufficient to induce the very efficient self-organization of the aggregates into structures that bear remarkable morphological resemblance to the human blastocyst. This protocol is robust and, with minor tuning of concentration of inhibitors, multiple hPSC,both ESC and iPSC, lines formed such structures with remarkably high efficiency (70-90% for every cell line we tested).
Fantastic 4 features: Rise of the human blastoids
For embryo models to be valuable, they must possess a good level of analogy with the embryo. Therefore, here we benchmarked our blastocyst-like structures using several known features of the human blastocyst. Here, we propose 4 cardinal features that define a human blastoid.
1. Size, morphology, and cell numbers: A fully expanded human blastocyst consists of a trophectoderm cyst of diameter between 150-250 μm that surrounds a single fluid-filled cavity and encompasses a single cluster of inner cells. We used the threshold of 150-250um diameter to define a blastoid and thus their yield of formation. These blastoids showed remarkable morphological resemblance to the human blastocysts and surprisingly, almost always form only one inner cell cluster of Inner Cell Mass (ICM)-like cells. Moreover, the total number of cells as well as ratios between various compartments are conserved as well (Figure 1h1). However, in order to better benchmark blastocyst-like structures, a more detailed and more extensive reference dataset of cell numbers within each lineages and over blastocyst development time (e.g., day 4-7), would be highly valuable.
2. Timing and sequence of development: The human blastocyst forms towards the end of the 4th day after fertilization and continues to develop for the next 3 to 4 days, at which time (~days 7-8) it undergoes implantation. Thus, blastocyst development from its formation to implantation takes 3 to 4 days. During that time, a precise sequence of events happens: First, the trophectoderm and ICM form. Then, in a second step, the ICM generates GATA4+ primitive endoderm cells and the trophectoderm matures to form NR2F2+ polar trophectoderm. This developmental dynamics of lineage specification was spontaneously recapitulated during blastoid formation: Similar to human blastocysts, trophectoderm and ICM analogs were the first lineages to form (24-60 hours), while primitive endoderm and polar trophectoderm analogs only formed in the second stage (60-96 hours). Of note, analogs of primitive endoderm and of the polar trophectoderm formed autonomously, solely based on the initial conditions, and probably due to intrinsic cell interactions and positional information originating from the overall geometry of the structures. No external stimulations are necessary during intermediate stages.. This pace matching is important in order to ultimately form cells that reflect the blastocyst stage. The necessity to culture cells for a longer time in order for morphogenesis to occur properly would also lead to the formation of cells reflecting the post-implantation rather than pre-implantation stage.
3. Transcriptomic states and stage: Human blastocysts ultimately consist of only three cell types, epiblast, trophectoderm and primitive endoderm that are each characterized by a distinct transcriptome. Importantly, these transcriptomes significantly differ from that of their derivatives in the post-implantation stages. It is thus possible to qualitatively infer the stage equivalent of the cells based on their transcriptome. However, methods to put hard numbers on this remain insufficiently developed. It is crucial for blastoids to be composed of analogs of only the blastocyst cell types and devoid of their post-implantation derivatives or of any other ‘off-target’ cell types. Several recent studies have reported high quality single cell transcriptomic datasets from human embryos of various stages ranging from morula to gastrula11,13,14. The latest one was produced by the laboratory of Laurent David (INSERM and Université de Nantes, France) who combined a metadataset and developed a pseudotime trajectory that appeared extremely useful36. An efficient and relaxed collaboration with this laboratory allowed us to have precise reference maps in order to determine the stage of the cells formed within the blastocyst-like structures and the precise cell states composing them. Ultimately, the blastocyst-like structures formed contained only three main, transcriptionally distinct clusters and each individual cluster showed a gene signature of epiblast, trophectoderm and primitive endoderm respectively. However, the decisive experiment came when we projected these cells on the human embryonic reference map consisting of cells isolated from pre-blastocyst, blastocyst, in vitro cultured blastocyst (equivalent of day 7 to day 14), and gastrulating embryos (CS7). The cells isolated from the blastocyst-like structures coincided quite precisely with the transcriptome of the cells of the blastocyst but not with pre-blastocyst stages or any post-implantation stages. Specifically, the transcriptome of the cells that formed overtime matched with day 5 blastocysts after 24 hours, and with day 7 blastocysts after 96 hours thus revealing the stages that are modeled. We found a minor population of ‘off-target’ cells which accounted for less than 3% of the cells analyzed. Thus, with morphological resemblance to human blastocyst, efficient formation of analogs of blastocyst cells according to the sequence and pace of blastocyst, and absence of off-target cells, we now felt confident to refer to these structures as blastoids.
4. Implantation and post-implantation developmental potential: Between day 4 and day 7 after fertilization, the blastocyst forms a cavity, which expands. During this, the blastocyst prepares itself to interact with the uterus so as to implant in the right direction and facilitate subsequent embryonic development. For example, the trophectoderm patterns into 2 distinct zones: the trophectoderm cells closer to the epiblast form the polar trophectoderm while the cells on the opposite end form the mural trophectoderm. In human blastocysts, such patterning is key for implantation as the polar trophectoderm forms the sticky cells that mediate the initial interactions with the endometrial cells of the uterus. Strikingly the trophectoderm analogs of the majority of blastoids (>60%) were patterned into polar and mural-like trophectoderm cells as seen by their protein expression profile (NR2F2High/CDX2Low). This indicated that the blastoids may possess the ability to undergo directional implantation. However, it is of course unethical and now forbidden by the ISSCR to transfer human blastoids into the uterus of humans or any other model organisms for any purpose including for scientific experiments37,38.
Therefore, we developed, through a fruitful and fun collaboration with Nina Maenhoudt and Hugo Vankelekom (KU Leuven, Belgium), an in vitro implantation assay using the cells from the endometrial organoids. We were extremely fascinated to see that, when blastoids were deposited on endometrial cells previously stimulated with hormones, blastoids attached to the endometrial cells thus mimicking the first step of implantation. However, this interaction did not occur if endoTherefore, we developed, through a fruitful and fun collaboration with Nina Maenhoudt and Hugo Vankelekom (KU Leuven, Belgium), an in vitro implantation assay using the cells from the endometrial organoids. We were extremely fascinated to see that, when blastoids were deposited on endometrial cells previously stimulated with hormones, they attached to the endometrial cells thus mimicking the first step of implantation. However, this interaction did not occur if endometrial cells were not stimulated with hormones. In accordance with clinical knowledge, we concluded that the hormones changed the state of the endometrial cells and made them receptive for the blastoid. Even more surprisingly, upon live imaging, we realized that this initial interaction was reproducibly mediated by the cells on the polar side of the trophectoderm. This was an exciting moment for us because replicating this directional attachment to hormonally stimulated endometrial cells gives confidence that we can mimic important aspects of the initiation of implantation. Importantly, the exposure of blastoids to a STAT inhibitor (SC144) and to a strong Hippo inhibitor (XMU-MP01) prevented the formation/maintenance of the inner cell cluster and thus to the formation of trophospheres. These trophospheres that transcriptionally appear to most resemble mid-stage blastocysts were incapable of attaching to hormonally stimulated endometrial cells. These additional controls reinforced our conviction that this assay demonstrated the specificity and reliability of blastoids to mimic aspects of implantation. It also opens up new possibilities to understand the molecular basis of embryo-uterus cross talk during implantation. Finally, blastoids can also be cultured for an extended period of time after their attachment to endometrial cells, during which they consistently expand and attain several features of post-implantation human embryos. For example, some trophectoderm cells differentiate into syncytiotrophoblast and extravillous trophoblast; the epiblast begins to express CD24 and appear more epithelial. However, since the methods to culture human blastocysts to post implantation stages are currently suboptimal, the extended blastoid cultures cannot be appropriately benchmarked. We do not think that we have properly modeled a day 13 stage. Nevertheless, blastoids cultured to extended periods augurs well for using these to study peri-implantation embryonic development.
The above criteria canThe above criteria can be used to determine the reliability and predictive power of any in vitro blastocyst model. We propose that these initial criteria must be thoroughly evaluated in order to define the in vitro structures as blastoids. However, we also acknowledge that these criteria are currently minimal. With further research more features such as the proteomic and epigenetic status, the mechanisms of lineage specification, the X chromosome status, etc., must be benchmarked to the human blastocysts and such features measured to evaluate the quality and utility of the model.
Publishing human blastoids: A long roller coaster ride
The journey between the generation of the first human blastoid in our lab to the first submission of our manuscript was an intense one. Several members of the team took responsibility for different aspects of the study and worked in a highly coordinated manner. Though intense, the team made this journey an exciting and delightful one. We submitted our manuscript with a completely optimized blastoid to the journal Nature on the 12th of February 2021. At that time, no other protocol to form models of the human blastocyst had been published. The editor received our manuscript with a lot of enthusiasm but there was definitely a shared sense of urgency to publish it. As a consequence, the first round of the peer review process was accelerated as well. We are extremely thankful to our reviewers Jan Bronsens, Jianping Fu, Insoo Hyun (open reviewers) and 2 other anonymous reviewers for the fast yet mostly constructive criticisms and the suggested experiments. However, the editor gave us a hard deadline of only 4 months to revise the manuscript and only one chance for re-submitting a manuscript that would address all the reviewer’s comments.
This brought back the intense phase, but this time with a deadline. Our synergistic work and the help of Maria Novatchkova (an excellent bioinformatician at the Vienna Biocenter) and collaborators, especially in that revision phase, from the lab of Laurent David, made it possible to submit a revised manuscript after approximately 3 months, about one month before the deadline. However, during this phase several reports were published reporting blastocyst-like structures, albeit with low efficiency. Though we were disappointed, we were rejoiced by the interest that the scientific community has towards this field.
Within weeks after the first attempts were released, a consortium of independent stem cell biologists and embryologists led by Fredrik Lanner (Karolinska Institute) and Sophie Petropoulos (Montreal University) gathered to evaluate the state and the stage of the cells within these models. They used an extensive reference map formed by parametrizing and merging multiple datasets of cells harvested a pre-blastocyst, blastocyst, and post-implantation stages. The initial report from this consortium was very rapidly published on Biorxiv and reported the abundance of ‘non-blastocyst’ like cells that closely resemble several post-implantation cell types present in the amnion, mesoderm, and primitive streak (~day 14)39. The presence of such ‘off-target’ cells is probably due to the sub-optimal initial cell state and/or suboptimal combination of molecules, which results in slow development (6 days or 9 days) and in a process of directed differentiation that led to the formation of cells similar to the gastrulation (day 14) and germ layer (mesoderm) stages. We suspect this, along with the low efficiency, can greatly reduce the utility of the model. It reignited our enthusiasm to finalize our proposition to form a model of human blastocyst-stage embryo.
While setting up rapid and good standards for the field, this independent consortium report probably caused an unprecedented scrutiny over our manuscript. We received very positive comments from 2 out of 3 of the reviewers which hinted our optimistic minds towards the acceptance of our manuscript. In the meantime, an ethicist, who turned out to be Insoo Hyun (open reviewer no. 4) gave us an ethical green light based on compliance with ISSCR guidelines and on the assessed oversight by the Austrian Academy of Sciences. However, the editor decided to add an additional reviewer (Reviewer no. 5) specifically to ensure the validity of the generation and analysis of the single cell transcriptomic data used to claim the formation of cells reflecting the blastocyst stage. Although disappointed by that unexpected move that consumed more time than we anticipated, we were excited to learn a bit more than one month later, at the end of August, that our scRNA seq analyses had passed this additional test. In addition, an update of the meta-analysis led by Fredrik Lanner and Sophie Petropoulos also included our model along with the one formed in the lab of Ge Guo (Exeter University) and showed that these models reflect the blastocyst stage. Forming the right cells is crucial because only faithful embryo models will be capable of reliably predicting human development. This will ensure that we can trustfully use embryo models to infer the (epi)genetic mechanisms, molecular pathways and cellular behaviors of blastocysts. While this first report from the consortium is important, more detailed analysis (such as quantitative assessment of the resemblance of a specific cell type of a blastocyst model to the one of the blastocyst) would be extremely beneficial to assess degrees of similarity. Such an understanding of the initial cell state and of the combination of molecules that allow for the formation of human blastoids also call for unbiased re-analysis of single cell RNA sequencing (scSeq) by independent bioinformaticians. It would support valid scientific and medical discoveries, and assist ethical evaluations.
The editor then sent us a ‘provisional’ acceptance (provided it clears the editorial criteria) on September 3rd, 2021. However, for reasons that we do not understand, we had to anxiously wait for more than 75 days for our manuscript to become formally accepted on the 18th of November, 2021. Finally, the study was published on the 2nd of December 2021, 3 months after provisional acceptance, through an accelerated track gladly proposed by the editor.
We hope that the scientific community has read our study with as much excitement as we had when first realizing that these blastoids form reliably according to the cardinal features of human blastocysts (size/morphology/cell numbers, timing/sequence of development, transcriptomic states/stage, functional implantation, and post-implantation developmental potential). Our data points to the possibility that the proposed blastoid is valuable to study early human development. Overall, this scientific journey shows that the initial cell state (PXGL naïve hPSCs) and the combination of molecules (triple inhibition of the Hippo/ERK/TGF-β pathways) allow for the formation of human blastoids that accurately form blastocyst-stage cells. However, we strongly encourage independent groups and bioinformaticians to critically work with our sequencing data sets to compare with that of human embryos to verify or challenge our analysis. Afterall, self-correction can only allow for the field to progress toward more stringent criteria and models predictive of human blastocyst development and implantation.
Blastoid team: great companions in a gratifying journey
Working on the project to develop human blastoids was a rare opportunity that I was extremely privileged to experience. I am extremely thankful to my supervisor Nicolas Rivron for providing me this opportunity. The most striking aspect to me was his broad vision behind the development of the project and the tremendous planning to execute the ideas in a highly coordinated manner. This was made possible by the extremely supportive and helpful atmosphere in the lab. When I started working in this lab, in January 2020, I joined the efforts of an extremely talented postdoc, Harunobu Kagawa, in characterizing naive PSCs and human trophoblast stem cells. Meanwhile, in parallel, another postdoc, Heider Hedari Khoei, who joined the lab just 2 days after me, used his clinical expertise on endometrial organoids. Through collaborative work with Nina Maenhoudt and Hugo Vankelekom (KU Leuven, Belgium), he developed the in vitro implantation assay. Thus, when we successfully developed our first human blastoid, we had already established a system to assess their functionality by the implantation assay. This synergistic team work within the lab (Theresa Sommer and Yvonne Scholte Op Reimer performed several crucial experiments and Giovanni Sestini led all the analysis of all the sequencing data) as well as collaborations (Labs of Laurent David, Hugo Vankelekom and Thomas Freour) allowed for the realization of this exciting study. This has been a learning experience and a journey that I will cherish. Blastoids have now opened up a new opportunity to dive deep into exploring mechanisms underlying early embryogenesis in humans. Thus, the exciting journey continues.

 (No Ratings Yet)
(No Ratings Yet)
 (2 votes)
(2 votes)