Cell migration needs to be precisely regulated during development so that cells stop in the right position. A new paper in Development investigates the robustness of neuroblast migration in the C. elegans larva in the face of both genetic and environmental variation. To hear more about the story, we met the paper’s four authors: Clément Dubois and Shivam Gupta, and their respective supervisors Andrew Mugler (currently Assistant Professor at the Department of Physics and Astronomy at the University of Pittsburgh, where his lab recently moved from Purdue University) and Marie-Anne Félix (Principal Investigator at Institut de Biologie de l’Ecole Normale Supérieure in Paris and Research Director at CNRS).
Clément (top left), Shivam Gupta (top right), Marie-Anne (bottom left) and Andrew (bottom right).
Andrew and Marie-Anne, can you give us your scientific biographies and the questions your labs are trying to answer?
AM: After a PhD and postdoctoral research on theoretical biological physics, I began my own position investigating noise in biological systems. Cells contend with unavoidable noise from environmental fluctuations, small molecule numbers and many other sources. My group uses tools from theoretical physics to uncover strategies that cells use to precisely sense their environment, undergo controlled developmental or phenotypic changes, and execute collective behaviours.
M-AF: After a PhD in cell biology, I wanted to combine evolutionary biology with cell/developmental biology. In Paul Sternberg’s lab at Caltech and in my lab’s early years, we compared vulva development in different nematode species, giving flesh to the concept of developmental systems drift. Over the past 20 years, we moved to a microevolutionary scale: studying the sensitivities of developmental systems to various types of perturbation and comparing these with their evolutionary variation. The overall thread is to assess how much development biases phenotypic evolution, with some phenotypes being more easily reached than others upon random genetic variation or quantitative tuning of parameters.
Studying natural variation led us to seek C. elegans in nature, which was a lot of fun. With many colleagues, we have since been probing its population genetics structure, and the zoo of its associated microbes. Bringing real worms back to the lab also led us to discover natural variation in the duration of their multigenerational memory.
Clément and Shivam – how did you come to work in your respective labs and what drives your research today?
CD: I did an MSc in ecology and evolution, and was specifically interested in host-pathogen interactions. After a first internship in the Pasteur Institute working on mosquito-virus interactions, my former supervisor suggested I apply to Marie-Anne’s lab. I started working on the intraspecific evolution of C. elegans resistance to microsporidia, and then did a year as a technician working on the evolution of a peculiar cell in C. elegans called P3.p. This year was really formative. I gained a lot of scientific maturity, and realised how rich and friendly the C. elegans community is. Then Marie-Anne proposed I do a PhD on the evolution of QR neuroblast migration. Considering the different topics I worked on, I would say that my research is mostly driven by curiosity and the question of how much genotypes can shape phenotypes.
SG: I came to Purdue University as a grad student in 2015. I joined Dr Mugler’s lab after doing a lab rotation to get a feel for the research. The research work in the Mugler lab was very challenging, and it was intriguing to see how fundamental physics can explain the behaviours of microscopic organisms.
How has your research been affected by the COVID-19 pandemic?
M-AF: The lab closed for 3 months in the spring of 2020; since then it has been somewhat restricted, but we can do experiments. Of course, C. elegans is a great organism to work on because we do not lose strains: they can be kept frozen or can live for 3 months in diapause (a pre-adaptation to SARS-CoV2!). But some projects were affected for more than 3 months. To enhance social interactions, since the fall we decided to meet after lunch every day in a hybrid format (real life and video). This compensates for the usual lunch-time exchanges, be they scientific or not. On the positive side, the pandemic also gave us the impetus to meet more readily with colleagues worldwide, attending each other’s lab meetings or organising informal meetings.
AM: As a theory group, we could continue most of our work from home without interruption. However, developing ideas in any area benefits from true collective brainstorming, which has been slowed without the ability to physically meet at the whiteboard. Additionally, the pandemic has increased my childcare responsibilities, which has left less time to dedicate to students. On the positive side, I moved institutions in the middle of the pandemic, and I now hold hybrid group meetings with group members from both places, which is perfectly natural to do virtually in these times.
How did your labs come to collaborate on this project?
AM & M-AF: This was all started by our colleague Rik Korswagen, who has been working on C. elegans QR migration as a model. He contacted us both to write an application for a collaborative grant from the Human Frontier Science Program. We got the grant, and this has been a great experience. The modelling of body size variation was not in the grant, but the collaboration took place naturally once we were networking.
Can you give us the key results of the paper in a paragraph?
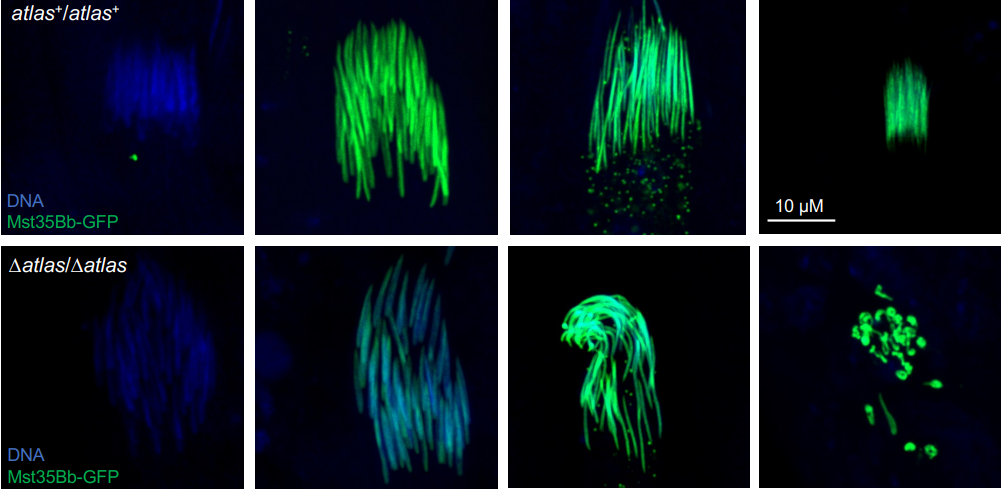
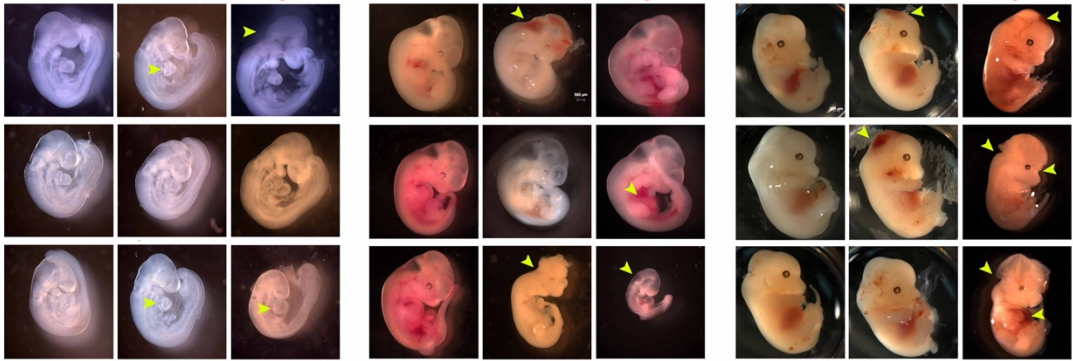
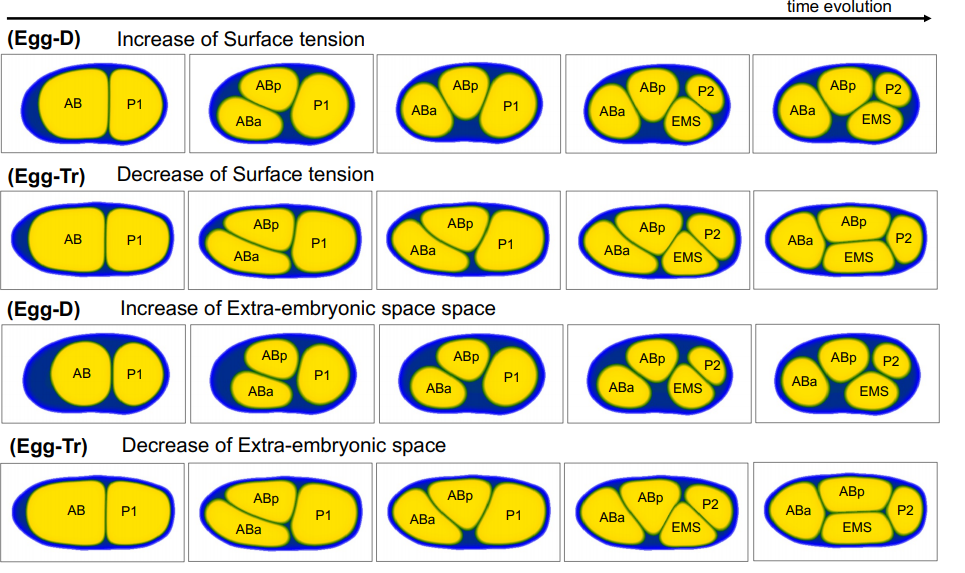
CD, SG, AM & M-AF: The goal was to measure the precision and natural variation in the final position of a cell that migrates during development. Other studies in various systems have examined the directionality of cell migration, but to our knowledge not the precision in the cell’s final position. Specifically, we studied two descendants of the QR neuroblast lineage that migrate a long way during C. elegans development. The Korswagen lab had shown that the end of their migration is specified by a temporal rather than a spatial mechanism. Given this temporal regulation, we hypothesised that a change in body size would affect the final position of the neurons relative to body landmarks. Indeed, we observed that a smaller body size resulted in the cell migrating further. Cell position even changed as a function of maternal age, a factor previously known to affect body size. We developed a mathematical model of the expectation, taking into account larval growth during the migration. The data did not fit quantitatively the simplest form of the model. Then, a model with partial compensation of body size, grounded on measurements of cell speed, fitted the experimental results without any free parameters. Finally, we revealed natural variation among wild isolates of C. elegans in the neurons’ final position, large enough that they result in a change in their neighbouring cells.
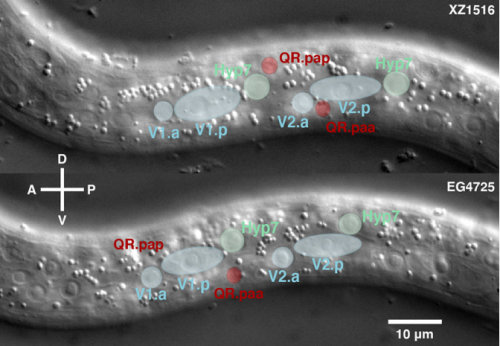
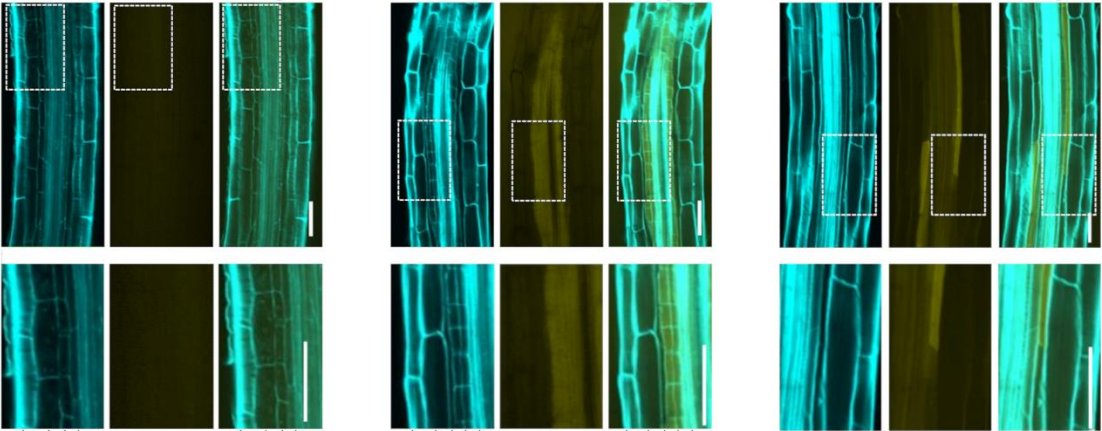
Late L1 larvae showing examples of the relative position of QR.pax in different strains: posterior (strain XZ1516, top) and anterior (strain EG4725, bottom).
Do you have any idea why cell migration speed is dependent on body size?
CD & M-AF: The first that comes to mind is that a larger body size means larger cells (at least for a species such as C. elegans, which has a quasi-invariant number of cells). But interestingly, we could not find much published on whether a larger cell would go faster or slower than a smaller one. Both appear possible, depending on specific motility mechanisms. Another possibility is that the external environment of the cell is altered as a function of body size; for example, a change in the Wnt gradient that influences QR migration, either its initial concentration or its decay length.
On the other hand, in the wild isolates, the eventual position of QR.pax did not correlate with body size: why do you think this is?
CD & M-AF: We did not detect a significant correlation, and we cannot rule there would be one with a larger set of experiments. In any case, this means that much of the variation is due to other factors that can go in another direction than the effect of body size. Variation in many genes and processes can affect a quantitative trait such as cell position.
Your paper was published in Development through the Review Commons route – how did you find the experience?
CD, SG, AM & M-AF: Great! This was our first time. With Review Commons, the manuscript is sent to reviewers without any submission to a specific journal. The reviewers do not have any specific journal in mind. In our case, the reviews were very constructive, especially by suggesting that we assessed the effect of maternal age. Once the reviews arrived, the authors can contact one of the participating journals, specifying what is planned for responding to reviews. Thanks to Development, this went very well.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
CD: The first eureka moment was to see the preliminary results on the natural variation of QR.pax final position. To be able to detect differences between wild isolates for the first time was really exciting and promising. From then on, I was excited at the end of each experiment after making plots from the data (and I spent a lot of time on them!).
SG: The first few months of my PhD work was mainly building mathematical models for temporal precision in gene expression. At first, I was a bit pessimistic that the rules of physics would explain the complex behaviour of organisms. After analysing experimental data from C. elegans, it was very satisfying to see that the behaviour of microscopic cells can be explained by our models.
And what about the flipside: any moments of frustration or despair?
CD: I did not have a particularly memorable moment of frustration or despair – even negative results were informative. Nonetheless, the most challenging part was probably the measurements of embryo and larva lengths at different time points, in parallel on four genotypes. It required a lot of organisation and efficacy, but it was worth it.
SG: To solve complex biological problems, we start with a simple model and slowly build up the model. After a point, our model became very complex and it took days to run the computer program. It was frustrating to wait days to see results, but in the end, when results were promising, it was very joyful.
What next for you two after this paper?
CD: In the short term I’m doing two things: writing my PhD manuscript and working on the intraspecific evolution of the final position of QR.pax. Using Recombinant Inbred Lines between CB4932 and JU1242 (two genotypes with an opposite phenotype), I found a QTL on chromosome IV. The goal now is to find the polymorphism associated with the difference in QR.pax final position.
SG: I have successfully defended my thesis and secured a job in the pharmaceutical industry. My ambition is to advance drug discovery and safety of drugs through developing pharmacokinetics/pharmacodynamics models and analysing clinical trial data.
Simple mathematical models can prove very useful for sharpening basic understanding of biological systems
Where will this story take the Mugler and Félix labs?
AM: This has been one of the most positive demonstrations that simple mathematical models can prove very useful for sharpening basic understanding of biological systems; I hope to continue in this vein with this and other collaborations.
M-AF: I hope we can find the polymorphism behind the divergent phenotypes of two wild C. elegans isolates. Beyond, I will keep vulva development as our main developmental model, but, as both are regulated by the same Wnt gradient, we will likely come back again to QR migration.
Finally, let’s move outside the lab – what do you like to do in your spare time in Paris and West Lafayette?
CD: Paris is full of small and hidden restaurants that are really good. I like to explore them with my partner. I also enjoy spending time with my friends playing board games, climbing in the gym or in Fontainebleau, a forest close to Paris known for its large boulders.
SG: I enjoy exploring new places around West Lafayette for hiking and biking.
AM: Biking, exploring new restaurants and travelling.
Hello! Welcome back to the #devbiolwriteclub! Over the last year or so, I’ve harangued you on Twitter and on The Node about practicing the craft of writing. I’ve ignored any practical advice on what you should actually put on a page and instead I have focused on how to build the habits of mind that allow you to grow as a writer. Yesterday, I launched a new Twitter project called #devbiolgrantclub, where I’ll be offering random bits of grantsmithing advice. Today, I’ll present a “crossover episode” of the two projects.
As academic scientists, we all know that we must master two types of writing: papers and grants. But I don’t think enough people really grasp the fact that the two represent entirely distinct disciplines within scientific writing. In fact, the best way to write a grant is totally, totally different from the best way to write a paper.
Ask anyone in my lab, and they’ll tell you that I love papers like druids love trees. Writing papers is one my life’s greatest joys. It’s also the ultimate goal of science, which is why papers are the hard currency of our field. In fact, call me sentimental, but I see something noble in our endeavor to elegantly reveal to the world the new knowledge that we have discovered.
Just the same, doing this noble work while simultaneously contemplating my next grant proposal brings to mind a great Melvern Taylor song, from which I borrowed this post’s title. That angel is perched there on your shoulder, egging you on as you do the good work of science. But let me tell you, successful grant writing absolutely requires that you ignore that angel and listen very carefully to the devil on your other shoulder.
The best grant writing advice I ever heard came from a clinician from UT, San Antonio at a grant writing workshop I attended when I was in Berkeley as a postdoc. Between solid practical advice and a riotously funny story about tequila and a big stack of grants to review, he said this:
“Listen. You don’t have to be proud of what you wrote. You have to get the money.”
Think about that. Let it sink in. It’s harsh, for sure. It may even be antithetical to how we see ourselves as academic writers. But it’s the ultimate truth of grant writing. You don’t have to be proud of what you wrote. You just have to get the money.
Now let me be clear: I am not talking about being sleezy or self-aggrandizing, and I’m definitely not talking about making stuff up. Rather, I am simply proposing that you be intentional about grant writing by ignoring what you want, learning exactly what your audience needs, and providing exactly that.
To understand what I mean, consider this: Every once in a while, I find myself with a totally free afternoon. So, what do I do? I go get a cup of coffee, sit somewhere pleasant, and read papers. Maybe I grab a few from the stack on my desk, maybe I hit Pubmed to look for something new, or maybe I click a link on Twitter. It’s a joy.
On the other hand, no one in the history of science ever said to themselves: “Gee, I’d really like to read and carefully review ten randomly assigned grants in my free time today.” This gives us the First Principle of grant writing:
1. Assume your readers do NOT want to read your grant.
This is possibly the biggest concept that grant writers fail to grasp. When you write a paper, you can reasonably assume that whoever reads it wants to read it. They very likely share your interest in the subject. You might even say they want to know how the story ends. Thus, they are actually quite likely to overlook a confusing paragraph, or power through a difficult passage. At the very least, they’ll give it a read and likely learn something.
But your reviewer did not choose to read your grant. Your reviewer was assigned your grant by a grant officer. The cynical (and effective) grant writer therefore assumes that the reviewer does not care how the story ends, but does know exactly when it will end: When the review is written and submitted to that grant officer.
Presented with this hard fact, you might feel compelled to try to write the grant in such way as to MAKE the reviewer want to read the grant. Don’t. It’s too risky. There are just too many variables. (E.g., Sorry, you cannot ever make me want to read a grant about… well, lots of things, but I won’t name them.)
On the other hand, there is one thing that will make all reviewers happy, and that is making the grant EASY for the reviewer to read and absorb. More thoughts on how to do this later, but for now, let me drive home why this strategy works by presenting the Second Principle of grant writing:
2. Accept that your readers MUST compare your grant to other grants, mentally ranking them in real time.
This is another critical distinction between grants and papers that people usually don’t consider when writing. If I am doing my work as a scientist, I am judging the quality of every single paper. Am I convinced by the data? Do I care about the conclusions? Just the same, I am not sitting there thinking hard about whether your paper is better or worse than the last one I read. And I am absolutely not trying to mentally rank the last ten papers I read. On the other hand, if I am reading your grant, this is exactly what I am doing.
This simple fact creates a very different workflow for grant readers, as compared to paper readers. The grant reader won’t just start reading ten grants in the order they were assigned. Instead, most reviewers will at least glance through the grants they are assigned, but then will pretty quickly decide which ones to read first – or last. So, in very short order, a mental ranking of some sort is already starting to emerge. This bears directly on our work as grant writers.
Here’s another thought exercise. You are a reviewer with a stack of nine grants. Two are in your specific sub-field and are asking questions that you find interesting. One grant has the minimum-allowed 0.5-inch margins, long paragraphs taking up nearly half a page, no white space breaking up the text, tables filled with huge amounts of data in small fonts, and no color anywhere. The other grant has larger margins and bullet points and colorful diagrams to summarize key points; these are well separated by white space from the main text, and that text is presented in short, easy-to-digest paragraphs. This second grant stops several inches short of the bottom of that last page. Tell me, which of those two grants -that you have to read today- will you choose to read first?
This brings us to the Third Principle.
3. Help your readers; they have to WRITE a careful review of your grant, AND ALSO several other grants.
This is the last major distinction between paper and grant readers. When I am reading a paper, I may or may not need to write about it, but if I do, I will usually end up doing so in a pretty indirect way. I’m very unlikely to do that writing the same day I read the paper. But consider the beleaguered grant reviewer. Just a few weeks to read an entire stack of grants, choose which ones are better and worse, and write reviews for all of them, justifying their decisions.
So, as a writer, this is where I am least proud of my grant writing: In my quest to make my grants easy for Reviewers, I consider that there are sections of NIH grant review forms titled Significance and Innovation. So, my grants include prominent, underlined sentences that read: “This grant is significant because…” or “This grant is innovative because…”. It’s clunky, unsubtle language, and I hate it. I hate bullet points, too. I also despise witless diagrams that I know are oversimplified. But if I provide these things, and do so with care and intent, it will help me get the money.
So those are three Principles to keep in mind when writing grants. Next, let’s see how they relate to the Rules of #devbiolwriteclub.
Rules #1 and #2 tell us to do the work. Make grant writing a specific craft you practice with intent throughout your career, not just this thing you need to do when you’re out of money.
Rule #3 tells us to revise and edit, again and again. When you are doing this, though, keep the Principles foremost in your mind. Revise and edit to make the grant EASIER to read, EASIER to rank, and EASIER to review.
Rule #4 tells us to read with intent. So, read grants with intent. Ask you PI and your peers for grants they wrote. Read the ones that got funded, but also read the ones that DID NOT get funded. Read the reviews! Here’s an idea I just came up with: Instead of journal club, have grant reviews club. Read a grant and the reviews together as a group. Try to figure it out.
Rule #5 says you can’t do it alone. So, when you beg your friends to read your grant, don’t ask them if they found typos. Ask them if it was easy to read. Also, recall that grants are assigned to reviewers, frequently outside their core area. So, ask people WHO ARE NOT IN YOUR lab to read your grants.
Finally, leave waxing lyrical and fighting the good fight for your papers.
Write grants with a singularity of purpose: Get the Money.
Naa12 compensates for Naa10 in mice in the amino-terminal acetylation pathway Hyae Yon Kweon, Mi-Ni Lee, Max Dörfel, Seungwoon Seo, Leah Gottlieb, Thomas Papazyan, Nina McTiernan, Rasmus Ree, David Bolton, Andrew Garcia, Michael Flory, Jonathan Crain, Alison Sebold, Scott Lyons, Ahmed Ismail, Elaine Marchi, Seong-keun Sonn, Se-Jin Jeong, Sejin Jeon, Shinyeong Ju, Simon J. Conway, TaeSoo Kim, Hyun-Seok Kim, Cheolju Lee, Tae-Young Roh, Thomas Arnesen, Ronen Marmorstein, Goo Taeg Oh, Gholson J. Lyon
Preterm birth alters the development of cortical microstructure and morphology at term-equivalent age Ralica Dimitrova, Maximilian Pietsch, Judit Ciarrusta, Sean P. Fitzgibbon, Logan Z. J. Williams, Daan Christiaens, Lucilio Cordero-Grande, Dafnis Batalle, Antonios Makropoulos, Andreas Schuh, Anthony N. Price, Jana Hutter, Rui PAG Teixeira, Emer Hughes, Andrew Chew, Shona Falconer, Olivia Carney, Alexia Egloff, J-Donald Tournier, Grainne McAlonan, Mary A. Rutherford, Serena J. Counsell, Emma C. Robinson, Joseph V. Hajnal, Daniel Rueckert, A. David Edwards, Jonathan O’Muircheartaigh
Connectomes across development reveal principles of brain maturation Daniel Witvliet, Ben Mulcahy, James K. Mitchell, Yaron Meirovitch, Daniel R. Berger, Yuelong Wu, Yufang Liu, Wan Xian Koh, Rajeev Parvathala, Douglas Holmyard, Richard L. Schalek, Nir Shavit, Andrew D. Chisholm, Jeff W. Lichtman, Aravinthan D.T. Samuel, Mei Zhen
A comprehensive series of temporal transcription factors in the fly visual system Nikolaos Konstantinides, Anthony M. Rossi, Aristides Escobar, Liébaut Dudragne, Yen-Chung Chen, Thinh Tran, Azalia Martinez Jaimes, Mehmet Neset Özel, Félix Simon, Zhiping Shao, Nadejda M. Tsankova, John F. Fullard, Uwe Walldorf, Panos Roussos, Claude Desplan
A multimodal iPSC platform for cystic fibrosis drug testing Andrew Berical, Rhianna E. Lee, Junjie Lu, Mary Lou Beermann, Jake A. LeSeur, Aditya Mithal, Dylan Thomas, Nicole Ranallo, Megan Peasley, Alex Stuffer, Jan Harrington, Kevin Coote, Killian Hurley, Paul McNally, Gustavo Mostovslavsky, John Mahoney, Scott H. Randell, Finn J. Hawkins
Stem cell-free therapy for glaucoma to preserve vision Ajay Kumar, Xiong Siqi, Minwen Zhou, Wen Chen, Enzhi Yang, Andrew Price, Liang Le, Ying Zhang, Laurence Florens, Michael Washburn, Akshay Kumar, Yunshu Li, Yi Xu, Kira Lathrop, Katherine Davoli, Yuanyuan Chen, Joel S. Schuman, Ting Xie, Yiqin Du
Zebrafish pigment cells develop directly from persistent highly multipotent progenitors Masataka Nikaido, Tatiana Subkhankulova, Leonid A. Uroshlev, Artem J. Kasianov, Karen Camargo Sosa, Gemma Bavister, Xueyan Yang, Frederico S. L. M. Rodrigues, Thomas J. Carney, Hartmut Schwetlick, Jonathan H.P. Dawes, Andrea Rocco, Vsevelod Makeev, Robert N. Kelsh
Intrinsic and extrinsic regulation of human fetal bone marrow haematopoiesis and perturbations in Down syndrome Laura Jardine, Simone Webb, Issac Goh, Mariana Quiroga Londoño, Gary Reynolds, Michael Mather, Bayanne Olabi, Emily Stephenson, Rachel A. Botting, Dave Horsfall, Justin Engelbert, Daniel Maunder, Nicole Mende, Caitlin Murnane, Emma Dann, Jim McGrath, Hamish King, Iwo Kucinski, Rachel Queen, Christopher D Carey, Caroline Shrubsole, Elizabeth Poyner, Meghan Acres, Claire Jones, Thomas Ness, Rowan Coulthard, Natalina Elliott, Sorcha O’Byrne, Myriam L. R. Haltalli, John E Lawrence, Steven Lisgo, Petra Balogh, Kerstin B Meyer, Elena Prigmore, Kirsty Ambridge, Mika Sarkin Jain, Mirjana Efremova, Keir Pickard, Thomas Creasey, Jaume Bacardit, Deborah Henderson, Jonathan Coxhead, Andrew Filby, Rafiqul Hussain, David Dixon, David McDonald, Dorin-Mirel Popescu, Monika S. Kowalczyk, Bo Li, Orr Ashenberg, Marcin Tabaka, Danielle Dionne, Timothy L. Tickle, Michal Slyper, Orit Rozenblatt-Rosen, Aviv Regev, Sam Behjati, Elisa Laurenti, Nicola K. Wilson, Anindita Roy, Berthold Göttgens, Irene Roberts, Sarah A. Teichmann, Muzlifah Haniffa
Inflammatory blockade prevents injury to the developing pulmonary gas exchange surface in preterm primates Andrea Toth, Shelby Steinmeyer, Paranthaman Kannan, Jerilyn Gray, Courtney M. Jackson, Shibabrata Mukherjee, Martin Demmert, Joshua R. Sheak, Daniel Benson, Joe Kitzmiller, Joseph A. Wayman, Pietro Presicce, Christopher Cates, Rhea Rubin, Kashish Chetal, Yina Du, Yifei Miao, Mingxia Gu, Minzhe Guo, Vladimir V. Kalinichenko, Suhas G. Kallapur, Emily R. Miraldi, Yan Xu, Daniel Swarr, Ian Lewkowich, Nathan Salomonis, Lisa Miller, Jennifer S. Sucre, Jeffrey A. Whitsett, Claire A. Chougnet, Alan H. Jobe, Hitesh Deshmukh, William J. Zacharias
A mechano-osmotic feedback couples cell volume to the rate of cell deformation Larisa Venkova, Amit Singh Vishen, Sergio Lembo, Nishit Srivastava, Baptiste Duchamp, Artur Ruppel, Stéphane Vassilopoulos, Alexandre Deslys, Juan Manuel Garcia Arcos, Alba Diz-Muñoz, Martial Balland, Jean-François Joanny, Damien Cuvelier, Pierre Sens, Matthieu Piel
A ciliopathy complex builds distal appendages to initiate ciliogenesis Dhivya Kumar, Addison Rains, Vicente Herranz-Pérez, Quanlong Lu, Xiaoyu Shi, Danielle L. Swaney, Erica Stevenson, Nevan J. Krogan, Bo Huang, Christopher Westlake, Jose Manuel Garcia-Verdugo, Bradley Yoder, Jeremy F. Reiter
Mouse oocytes do not contain a Balbiani body Laasya Dhandapani, Marion C. Salzer, Juan M. Duran, Gabriele Zaffagnini, Cristian De Guirior, Maria Angeles Martínez-Zamora, Elvan Böke
Genome editing in animals with minimal PAM CRISPR-Cas9 enzymes Jeremy Vicencio, Carlos Sánchez-Bolaños, Ismael Moreno-Sánchez, David Brena, Dmytro Kukhtar, Miguel Ruiz-López, Mariona Cots-Ponjoan, Charles E. Vejnar, Alejandro Rubio, Natalia Rodrigo Melero, Carlo Carolis, Antonio J. Pérez-Pulido, Antonio J. Giráldez, Benjamin P. Kleinstiver, Julián Cerón, Miguel A. Moreno-Mateos
Anatomical Structures, Cell Types, and Biomarkers Tables Plus 3D Reference Organs in Support of a Human Reference Atlas Katy Börner, Sarah A. Teichmann, Ellen M. Quardokus, James Gee, Kristen Browne, David Osumi-Sutherland, Bruce W. Herr II, Andreas Bueckle, Hrishikesh Paul, Muzlifah A. Haniffa, Laura Jardine, Amy Bernard, Song-Lin Ding, Jeremy A. Miller, Shin Lin, Marc Halushka, Avinash Boppana, Teri A. Longacre, John Hickey, Yiing Lin, M. Todd Valerius, Yongqun He, Gloria Pryhuber, Xin Sun, Marda Jorgensen, Andrea J. Radtke, Clive Wasserfall, Fiona Ginty, Jonhan Ho, Joel Sunshine, Rebecca T. Beuschel, Maigan Brusko, Sujin Lee, Rajeev Malhotra, Sanjay Jain, Griffin Weber
A dynamic pattern of histone methylation and demethylation controls gene expression during development, with some processes such as formation of the zygote involving large-scale reprogramming of methylation states. A new paper in Development investigates how inherited histone methylation regulates developmental timing and the germline/soma distinction in Caenorhabditis elegans. To hear more about the story we caught up with first author and postdoctoral researcher Brandon Carpenter, and his supervisor David Katz, Associate Professor in the Department of Cell Biology at Emory University School of Medicine in Atlanta, Georgia.
Brandon (L) and David (R)
David, can you give us your scientific biography and the questions your lab is trying to answer?
DK: As a graduate student, I worked with Dr Shirley Tilghman at Princeton on the regulation of genomic imprinting. We provided in vivo evidence for the first chromatin boundary formed by CTCF at the H19 locus in mouse. As a postdoc, I worked for Dr Bill Kelly at Emory University on the regulation of histone methylation in the germline of C. elegans. We provided the first evidence of a transgenerational phenotype (sterility) caused by the build-up of histone methylation, when the H3K4me1/2 demethylase Lsd1 is mutated. In my own lab at the Emory University School of Medicine, we have worked on both C. elegans and mouse model systems to study the mechanisms that regulate histone methylation and how inappropriately inherited histone methylation gives rise to phenotypes. We have also implicated LSD1 as a crucial molecule that may contribute to Alzheimer’s disease: our data suggest it is being inhibited by pathological aggregates in dementia patients. We are currently trying to develop a therapeutic intervention based on what we have learned about the function of LSD1 in the Alzheimer’s disease pathway.
Brandon – how did you come to work in David’s lab and what drives your research today?
BC: After obtaining my doctoral degree, I knew I wanted to continue studying development, and that I wanted to focus on a model system that would allow undergraduates to develop projects related to my research. As a graduate student, I fell in love with mentoring students and wanted to find opportunities to inspire students in the classroom as well as at the bench. Thus, my passion for mentoring and studying developmental biology led me to the Katz lab, where I could work with the beautiful model system, C. elegans, to study how epigenetic inheritance affects developmental cell fates.
At Emory University, I joined the Katz lab as a Fellowship In Research and Science Teaching (FIRST) Fellow (part of the National Institutes of Health-funded IRACDA programme) where, in parallel with my research, I was able to develop my teaching and mentorship skills. The most exciting part of joining the Katz lab was being able to bring my research into the classroom at Oglethorpe University, a nearby small liberal arts college. The Katz lab has a longstanding collaboration with Dr Karen Schmeichel from the biology department at Oglethorpe, integrating C. elegans experiments into the entire curriculum. As part of this, I was able to teach a semester-long Course-based Undergraduate Research Experience (CURE) based on the research I was conducting in the Katz lab. As part of this semester-long CURE, Oglethorpe students became inspired by the science we are doing in the Katz Lab and generated data for this manuscript. Jovan Brockett, an undergraduate student, is an author on this manuscript for research he did in the classroom! My passion for understanding how an organism develops from a single cell drives my research, and the feeling I get when I see my mentees succeed while studying these mechanisms provides the fuel that keeps me going.
How has your research been affected by the COVID-19 pandemic?
BC: For me, COVID-19 hit right as I was finishing the experiments for two of my postdoctoral projects. During the ∼2-3 month lab shut down I was able to stay productive by submitting and revising manuscripts, but being away from the lab made it hard to advance interesting new ideas. The thing I miss most about not being able to go into lab is my ability to bounce crazy ideas off my talented Katz lab colleagues.
Before your work, what was known about the role of inherited histone methylation in the germline/soma distinction?
BC, DK: This paper is really about two major lines of research coming together to create a new story. We had been working on how two histone modifying enzymes, the H3K4me1/2 demethylase LSD1 (SPR-5 in C. elegans) and the H3K9 methyltransferase MET-2 cooperate to reprogramme histone methylation at fertilization to prevent the inappropriate chromatin environment from being passed on from one generation to the next. We had found that a failure to reprogramme histone methylation in spr-5; met-2 double mutants causes a maternal effect developmental delay and sterility phenotype. We were interested in how the inappropriate inheritance of histone methylation causes the developmental delay. Dr Susan Strome, with some help from Dr Bill Kelly, had performed some beautiful work showing how maternal deposition of the H3K6 methyltransferase is required transgenerationally to help specify the germline in progeny. Brandon had noticed some similarities between the developmental delay that we were observing and some high temperature phenotypes that Susan Strome had shown and were continuing to be worked on by Dr Lisa Patrella in her own lab. As detailed below, Brandon was able to show that the MES-4 system and the SPR-5; MET-2 reprogramming mechanism antagonize one another. It is also important to note that several labs have identified somatic repression mechanisms that antagonize the MES-4 system. We are interested in seeing how these systems interface with SPR-5; MET-2 reprogramming, so stay tuned!
Can you give us the key results of the paper in a paragraph?
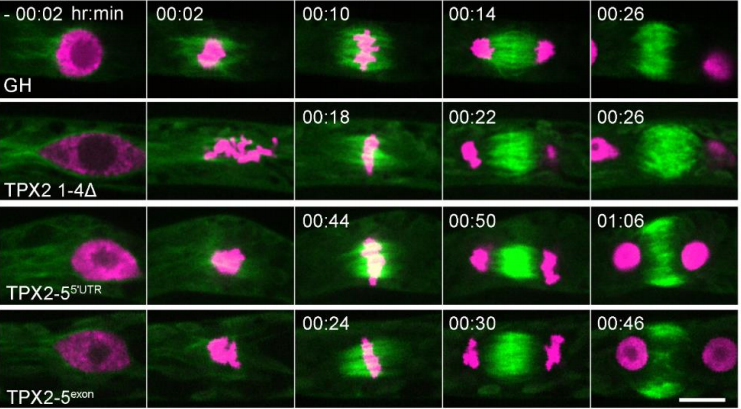
BC, DK: We had previously shown that SPR-5 and MET-2 act together to repress germline genes at fertilization. In this paper, we found that H3K36 methylation antagonizes this repression to prevent these germline genes from being completely shut down. Without inherited H3K36 methylation, the germline is not properly specified. In contrast, without SPR-5; MET-2 repression, H3K36 is inappropriately propagated to the soma, resulting in germline genes being inappropriately expressed there. The inappropriate expression of germline genes in the soma results in a developmental delay. Thus, neither SPR-5; MET-2 reprogramming nor the MES-4 germline inheritance system can properly function without each other. Instead SPR-5, MET-2 and MES-4 coordinately balance three difference histone modifications (H3K4, H3K9 and H3K36 methylation) to ensure that germline versus soma is properly specified.
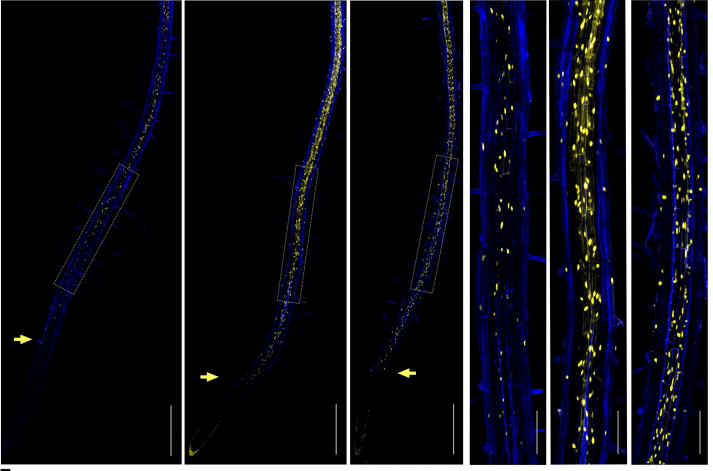
Single molecule fluorescence in situ hybridization image of a C. elegans L1 larvae ectopically expressing a germline-specific gene, htp-1, in somatic tissues (grey, htp-1mRNA; blue; DAPI).
Why do you think inappropriate somatic expression of germline genes causes developmental delay?
BC, DK: As discussed in the paper, we think that there are two possible mechanisms for how the ectopic expression of germline genes causes developmental delay in spr-5; met-2 mutants. One possibility is that transcription of the germline programme itself causes the developmental delay. For example, if germline transcription factors are competing with somatic transcription factors to turn on genes, it is possible that the mix of proteins generated is simply too confusing for the cell to commit to its proper cell fate. The alternative is that a part of the germline function interferes with somatic development; for example, the germline precursors undergo a cell cycle arrest. It is possible that the proteins involved in this germline cell cycle checkpoint slow the progression of somatic cells via cell cycle regulation. Consistent with this latter possibility, we show that spr-5; met-2 mutants can silence an extrachromosomal array in the soma. This function, which is normally confined to the germline, suggests that somatic tissues in spr-5; met-2 mutants make proteins that can perform some germline functions. Thus, it is possible that a germline function acting in the soma prevents somatic cells from quickly adopting their proper cell fate.
What relevance do your data have for human patients harbouring mutations in histone-modifying enzymes?
BC, DK: Recent genome sequencing has revealed that several neurodevelopmental disorders are caused by mutations in histone-modifying enzymes. These include mutations in: (1) the H3K36 methyltransferase Setd2, the H3K27 demethylase Kdm6a and the H3K4 methyltransferase Kmt2d, which cause Kabuki Syndrome; (2) the human orthologue of spr-5, LSD1, which causes a Kabuki-like Syndrome; and (3) the H3K36 methyltransferase Nsd1, which causes Sotos Syndrome. Similar to what we observed in spr-5; met-2 mutant progeny, many of the human patients with mutations in these histone-modifying enzymes suffer from global developmental delay. Based on our model, it is possible that the developmental delay in these patients may be caused by the failure to properly regulate histone methylation during essential developmental transitions. Consistent with this, we have recapitulated some phenotypes in a maternal hypomorphic mutant of Lsd1 in mice that are reminiscent of Kabuki Syndrome. We hope that by continuing to study how mutations in histone-modifying enzymes in C. elegans and mice give rise to developmental defects, we will shed light on the human diseases caused by defects in histone-modifying enzymes.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
BC: As detailed above, I had made the observation that the developmental delay we were observing had some similarities to some high-temperature phenotypes that Susan Strome had shown and were continuing to be worked on by Lisa Patrella in her own lab. This raised the possibility of a connection between SPR-5; MET-2 reprogramming and the MES-4 inheritance system. The crucial test of this potential connection was to knock down mes-4 via RNAi and see if it rescued the developmental delay, and we were very excited to find that it did rescue it, and even more excited when our RNA-seq subsequently showed that MES-4 germline genes are expressed in the somatic tissues of spr-5; met-2 mutants. After this independent confirmation of the connection between the two systems, we were confident that we had figured it out.
And what about the flipside: any moments of frustration or despair?
BC: The most frustrating part of this project was trying to gather enough L1 larvae to perform the initial genomic experiments. When we first started the project, there were no strains available that could balance the spr-5 mutant allele. I had to genotype every single hermaphrodite parent! At one point, I thought I would never get enough larvae to perform the genomic experiments. But David saw on science Twitter that the Caenorhabditis Genetics Center (CGC), which houses C. elegans strains, was developing new balancer strains. I contacted them and was able to get the FX30208 tmC27 [unc-75(tmls1239)](I) balancer even before they made it available to the broader C. elegans community. By reporting back that it worked well, I was also able to give back to the C. elegans community.
What next for you after this paper?
BC: I am officially on the academic job market searching for a tenure-track position and developing exciting new projects of my own that stem from this paper. We have mounting evidence that mutations in highly conserved histone-modifying enzymes may give rise to developmental phenotypes in vertebrates that are similar to what we observe in C. elegans. I want to take advantage of C. elegans mutants that fail to properly inherit histone methylation to further investigate how inherited chromatin states affect complex developmental processes like cell-to-cell communication and cell migration. I am also interested in potentially introducing the human version of Lsd1 into C. elegans to humanize the worm so that I can generate mutations that have been found in the human LSD1 patients. This type of approach is on-going through the NIH-funded Undiagnosed Diseases Network (UDN).
Where will this story take the Katz lab?
DK: We believe that spr-5; met-2 double mutants provide an excellent model for understanding how cells respond to inappropriately inherited histone methylation. We are taking advantage of the invariant embryonic cell lineage in C. elegans by performing automated lineage tracing experiments in spr-5; met-2 mutants. This will enable us to understand cell by cell how inappropriately inherited histone methylation affects processes such as cell division timing, cell migration, programmed cell death, etc. We hope to combine this with single cell RNA-seq to ask how each cell responds transcriptionally to this inappropriately inherited histone methylation. So stay tuned.
Remarkably, trying to understand the regulation of histone methylation in the germline has also taken us into the Alzheimer’s disease field. While trying to understand whether SPR-5; MET-2 reprogramming is conserved in mice, we serendipitously discovered that LSD1 is continually required for the survival of hippocampus and cortex neurons. We are interested in the possibility that terminally differentiated cells continually employ histone-modifying enzymes, such as LSD1, to maintain their cell fate. In the meantime, we have gone on to provide evidence that LSD1 is inhibited by pathological aggregates of tau in mice and human Alzheimer’s disease patients. We believe that this inhibition is a crucial part of how pathological tau induces neurodegeneration. So, you never know where developmental biology will lead!
You never know where developmental biology will lead!
Finally, let’s move outside the lab – what do you like to do in your spare time in Atlanta?
BC: I like to go hiking with my 5-year-old twin daughters, play golf, and find cool breweries who push the edge on brewing delicious stouts and IPAs!
DK: I enjoy soccer with my 13-year-old twins and co-host a popular Atlanta United podcast. I also co-founded a very small vineyard just outside Atlanta at a friend’s house, and we have just produced our first successful vintage of a Norton/Cabernet Sauvignon blend. But I am also happy to drink a local beer with my outstanding postdoc Brandon.
From six-toed cats to cyclops lambs – and, of course, it’s fabulous name – the Sonic Hedgehog gene has a fascinating history, as well as a whole bunch of interesting developmental biology behind it.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
By Koichiro Uriu, Bo-Kai Liao, Andrew C. Oates and Luis G. Morelli
How local cell-cell communication can generate a global tissue pattern is one of the fundamental questions in developmental biology. Yet, studying this remains challenging, because developing tissues involve complexities such as cell rearrangement, heterogeneity along the body axis, and massive tissue shape changes. In our recent paper in eLife (Uriu, Liao et al. 2021), we addressed this question using the zebrafish segmentation clock as a model system. Our integration of experimental and theoretical approaches revealed that after desynchronization, the recovery of the iconic synchronized wave pattern in the segmentation clock is influenced by two distinct spatial and temporal scales. Firstly, there is the faster and local communication directly between the cells, and secondly, there is a slower and much longer-distance movement of the cells and the tissues in the embryo as the tissues change their overall shapes.
This work started about 10 years ago, when the four of us were all at the Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG) in Dresden, Germany. In fact, we all worked in the same research group, so local communication was relatively easy. This local discussion was triggered when one of us, Bo-Kai Liao, noticed an unusual pattern of disrupted body segments in zebrafish embryos as they attempted to re-synchronize their cellular clocks after perturbation. The thing that struck us was that the pattern really couldn’t be explained by the current models of re-synchronization.
We are sure you all recognize the situation: “Hang on a minute, that should not be happening…” and you just know you are going to find out something cool. Well, in this case, the journey from initial observation to an explanation would take us a decade, during which time all of us moved from the MPI-CBG. We each moved country at least once, and we changed jobs and titles and had families and even got some grey hair. The scientific story mirrored the personal: how local communication is affected by long-distance movements. To understand this story better, we need to go back to the beginning and start with the biology.
The precursors of the body segments of vertebrates, called somites, are formed rhythmically during embryonic development. Each segment buds off from the unsegmented tissue, presomitic mesoderm (PSM) one by one. The rhythm of segment formation is determined by an oscillatory spatial pattern of gene expression in the PSM and tailbud, termed the segmentation clock. In these tissues, the peaks of gene expression travel across the anterior-posterior axis. Cells in the tissue synchronize their gene expression rhythms with neighbors through Delta-Notch signaling, providing local integrity of gene expression patterns. Understanding this synchronization, and it’s role in forming body segments is what brought the four of us together in the first place.
Previous studies had shown that the treatment of an inhibitor of Delta-Notch signaling, called DAPT, led to formation of defective segments in zebrafish embryos. Some of these studies had also observed the recovery of normal segments after the washout of DAPT (Riedel-Kruse et al. 2007; Liao et al. 2016). These results had been interpreted in terms of desynchronization and resynchronization of oscillators: treatment of DAPT desynchronizes oscillators by inhibiting intercellular coupling through Delta-Notch signaling, and the hallmark spatial wave pattern of gene expression in the PSM is abolished due to the noise in the individual cellular oscillators. However, its washout restores coupling, letting cells gradually resynchronize their oscillators. When the synchrony level reaches a threshold, a normal segment reappears. This desynchronization hypothesis has quantitatively explained several experimental data, but there was a remaining gap in our understanding: how is the tissue-scale pattern reorganized through local coupling?
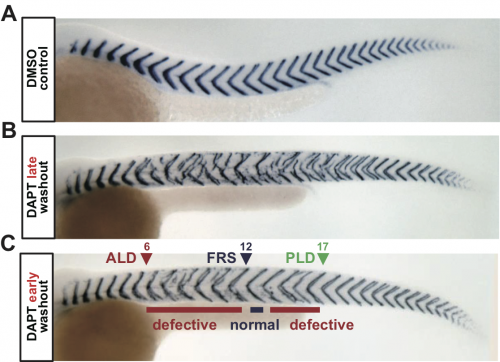
Figure 1. Intermingled segment boundary defects after DAPT washout. (A) Control embryo. (B) Embryo with late DAPT washout. (C) Embryo with early washout. Defective segments appear even after first recovered segment (FRS). ALD: anterior limit of defect. PLD: posterior limit of defect.
To address this, we analyzed segment recovery processes with different DAPT washout timing in zebrafish embryos. We unexpectedly found that washing out DAPT at earlier developmental stages caused intermingled segment boundary defects: a defective segment boundary was formed even after some normal segment boundaries were already formed (Fig. 1). This result suggests that the synchrony level fluctuates around the threshold, for some reason. We were very surprised by this result because we expected a monotonic recovery of normal segments based on previous theoretical works on coupled oscillators. Usually, once a population of oscillators gets synchronized, they remain synchronized and a large fluctuation of synchrony level hardly ever occurs. Thus, pattern recovery in the segmenting tissue seemed not so simple.
At this time, there were no transgenic markers of the zebrafish segmentation clock, nor were the microscopes, image processing, and data analysis developed to follow all these oscillating cells during the entire re-synch process (in fact, we are still working on this…).
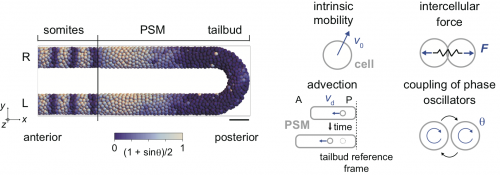
Therefore, we decided to use a physical model for the segmenting tissue to analyze this phenomenon. We have previously proposed models for the entire presomitic mesoderm and tailbud tissues in lower spatial dimension (1D or 2D; Morelli et al. 2009) or in 3D space but only a part of the tissue (Uriu et al. 2017). This time, we chose to describe the entire tissue in 3D and tried to integrate some of the previous modeling efforts by us and others in this framework (Fig. 2).
Figure 2 Physical model of the PSM and tailbud. Cells are represented as particles and rendered as spheres. Color represents the phase of oscillation. R: right. L: left. The model includes four key aspects in the tissue shown in right schematics.
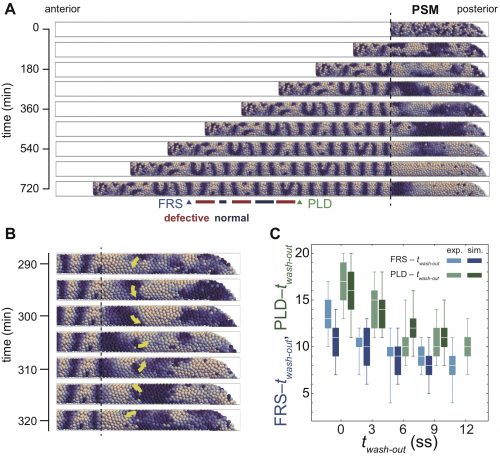
In numerical simulations of the model, we found a rotating phase pattern of oscillators, termed a phase vortex, in some situations of resynchronization (Fig. 3A, B). A phase vortex emerges in the tissue by local interactions of oscillators, moves through the tissue along the anterior-posterior axis by cell advection, and generates a defective boundary when it arrives at the anterior part of the PSM. A phase vortex can be formed posterior to the well-synchronized domain in simulation, so it can cause intermingled defective segments, as observed in the experiments.
Figure 3 Phase vortices and intermingled defects in simulations. (A) Snapshots of segments and phase patterns in the PSM. (B) Phase vortex in the PSM. Yellow arrow indicates the direction of rotation of the vortex. (C) Comparisons of time to first recovered segment (FRS) and time to posterior limit of defect (PLD) between embryonic experiment (exp.) and simulation (sim). twash-out: DAPT washout time. ss: somite stage.
It turns out that although the formation of a phase vortex is driven by the local interactions, its kinematics is determined by the larger-scale tissue properties. A phase vortex moves from posterior to anterior by cell advection caused by embryonic axis extension. We reasoned that the global tissue properties, such as cell advection and tissue length could affect the pattern recovery in the PSM. Moreover, these tissue properties change with developmental stages. Hence, our physical model predicted that the time to complete recovery, that is, when we do not see any further phase vortices, would depend on the developmental stage at which DAPT washed out. In fact, we observed good agreement between simulations and experiment for the time to complete recovery (Fig. 3C). So, are we done? No. The next key task is to test the theory by observing phase vortices in living tissues. This should be possible by the live imaging of reporters of both oscillatory protein and segment boundaries simultaneously, which requires the long-term imaging, tracking and analysis of the cells in the segmentation clock.
In summary, our study indicated that pattern recovery in the zebrafish segmentation clock occurs at two spatial and temporal scales: quick local synchronization and transport of local patterns through slow tissue shape changes. There’s a nice symmetry to the slow, long-range drift of the four authors, which certainly changed the dynamics of our local communication. Although we were already quite good at video conferencing when the pandemic struck, the available time interval when everyone on the team was awake and alert simultaneously in Japan, Taiwan, Switzerland and Argentina was rather short. Thus, a high signaling strength was an important asset. Indeed, working together on the project over such a long time and through so many life changes was a rewarding experience, and we hope you will enjoy the pattern that self-organized from the process as much as we do.
Uriu K., Liao BK., Oates A.C., Morelli L.G. (2021) From local resynchronization to global pattern recovery in the zebrafish segmentation clock. Elife 10:e61358. doi: 10.7554/eLife.61358.
References
Liao BK., Jörg D.J., Oates A.C. (2016) Faster embryonic segmentation through elevated Delta-Notch signalling. Nat Commun. 7:11861. doi: 10.1038/ncomms11861.
Riedel-Kruse I.H., Müller C., Oates A.C. (2007) Synchrony dynamics during initiation, failure, and rescue of the segmentation clock. Science 317(5846):1911-5. doi: 10.1126/science.1142538.
Morelli L.G., Ares S., Herrgen L., Schröter C., Jülicher F., Oates A.C. (2009) Delayed coupling theory of vertebrate segmentation. HFSP J. 3(1):55-66. doi: 10.2976/1.3027088.
Uriu K., Bhavna R., Oates A.C., Morelli L.G. (2017) A framework for quantification and physical modeling of cell mixing applied to oscillator synchronization in vertebrate somitogenesis. Biol Open. 6(8):1235-1244. doi: 10.1242/bio.025148.
The plant epidermis is a single layer of cells that forms a crucial barrier to the outside world, but the mechanisms that control epidermal differentiation – in particular the relative importance of position and lineage – remain incompletely understood. A new paper in Development tackles this question in Arabidopsis. To find out more about the story, we caught up with first author Kenji Nagata and his supervisor Mitsutomo Abe, Associate Professor at the University of Tokyo.
Mitsutomo (L) and Kenji (R).
Mitsutomo, can you give us your scientific biography and the questions your lab is trying to answer?
MA: As a PhD student I researched molecular genetics in Arabidopsis in the lab of Yoshibumi Komeda at Hokkaido University in Sapporo, Japan. I was excited to go to the lab every day, and so fascinated by the beautiful expression patterns of ATML1 and PDF2, twin genes that enabled me to get my PhD in 2001. After my PhD, I joined the lab of Takashi Araki at Kyoto University as an Assistant Professor and started the ‘florigen quest’ with great colleagues. I was fortunate to make a fundamental discovery regarding florigen in 2005, and I am very pleased that FT is now well known as an important component of florigen signalling. After this, I had a great experience working in the lab of Richard Amasino at the University of Wisconsin for two years, and very much enjoyed American life. I then moved to the University of Tokyo and started my own research group in 2009, and keep working on plant molecular genetics.
Our research group has a broad interest in plant development. Some members are involved in elucidating florigen function in Arabidopsis, others (like Kenji) are working on epidermal cell differentiation, and others are interested in the interaction between meristem identity and plant architecture. All have one thing in common: we are focusing on phenomena and molecules that are unique to plants.
Kenji – how did you come to work in Mitsutomo’s lab and what drives your research today?
KN: When I was an undergraduate student, I was very interested in two aspects of sessile plants: the phenotypic plasticity they show in morphology and physiology, and their developmental robustness in terms of patterning. So I looked for a lab that would allow me to explore these issues. I was fortunate to attend one of Mitsutomo’s lectures, where he described his lab’s work on flowering, which is mediated by signals from the external environment, and robust protodermal cell differentiation, which is not influenced by the external environment. I felt that working with him would provide me exciting research opportunities, and decided to join his lab. Curiosity about how the traits unique to sessile plants work and why they developed in an evolutionary context drives my research today.
How has your research been affected by the COVID-19 pandemic?
KN: Due to entry restrictions to the university, we were forced to suspend our experiments and stay at home. Fortunately, none of the members of our lab have caught the virus and all have stayed healthy so far. Although now there are still some restrictions, it is slowly getting back to (a new) normal.
MA: Like most universities and research institutes, only the minimum number of staff necessary to maintain plants and equipment was permitted to enter the lab from spring to summer. Since mid-July, research activities are allowed with the utmost care to prevent the spread of infection. But the number of infected patients in Tokyo is increasing recently, so I’m worried that the lab will be closed again.
Before your work, what was known about the relative roles of lineage and position in plant epidermis differentiation?
KN: It is widely accepted that cell position rather than cell lineage is important for plant cell fate decisions. For example, the inner cells, which derive from occasional periclinal divisions of epidermal cells, develop according to their position rather than their epidermal cell lineage. On the other hand, it is also known that, in shoots, the inner cells never adapt their epidermal identity even if they occupy the outermost position, suggesting that cell lineage is involved in the epidermal cell differentiation. However, it was recently shown that when cells in the inner cell lineage are displaced to the outermost position through laser ablation, they appear to acquire root epidermal cell fate. Thus, it is controversial whether cell lineage indeed affects epidermal cell fate decisions.
Can you give us the key results of the paper in a paragraph?
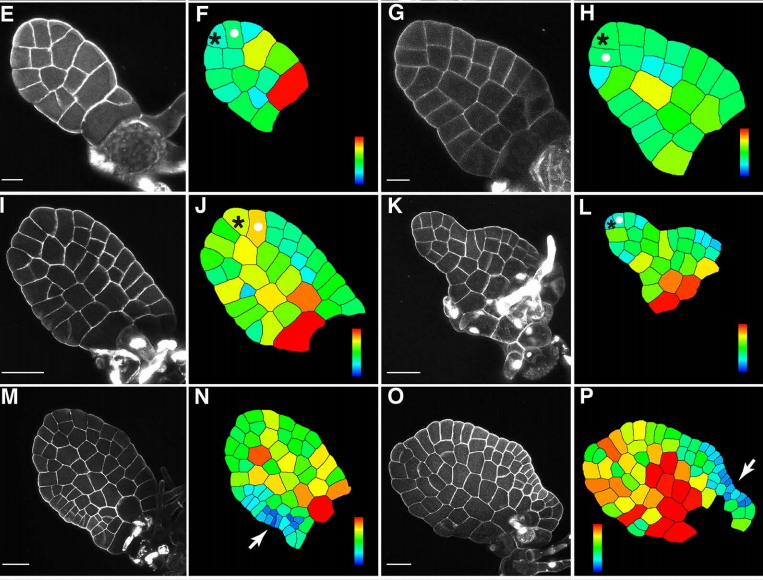
KN: In this paper, we found that ATML1, a master regulator of protoderm/epidermis differentiation, is only stabilized in the outermost cells derived from the outermost cell lineage. Furthermore, the stability of ATML1 in these cells is conferred by the interaction with its lipid ligand VLCFA-Cers. VLCFA-Cers appears to be polarly localized to peripheral domains in epidermal cells, and passed on to the outermost epidermal cells in a cell position- and lineage-dependent manner. Based on these results, we have proposed a novel model in which ATML1-VLCFA-Cers interaction is restricted to the outermost epidermal cells and consequently restricts protoderm/epidermis differentiation to the appropriate position.
MA: I think for me the key experiment in our paper is the transient induction of ATML1 by a heat shock treatment – I was very excited when Kenji first came to show me the GFP images.
Expression of gATML1-EGFP in two 16-cell embryos.
When outermost cells divide asymmetrically, the inner cells inherit ATML1 protein: what then stops them from differentiating as protoderm?
KN: This is because the ATML1 protein in inner cells is rapidly broken down, as shown in the careful observation in the 16-cell-stage embryo of gATML1-EGFP plants, or in our transient ectopic expression assay using HSP::NLS-mCherry; HSP::ATML1-EGFP plants in which the inner cells are not able to differentiate into protoderm. The rapid breakdown of ATML1 protein is due to the absence of VLCFA-Cers, in inner cells. Thus, we propose that VLCFA-Cers act as a landmark of the outermost cell position and lineage, and act as a post-translational signal that mediates positional information.
MA: Epidermis-specific expression of ATML1 and PDF2 is strictly regulated. Therefore, in addition to the mechanism we reported here, I believe that several other regulatory mechanisms are involved in epidermal cell differentiation. I’m really looking forward to the seeing this research progress in the future.
I was very excited when Kenji first came to show me the GFP images
Do lipid-transcription factor complexes mediate positional signals elsewhere in plant development?
KN: We would assume so. Kathrin Schrick and colleagues have shown that START domains from plant HD-Zip transcription factors bind lipid ligands to regulate transcription factor activity in a yeast system. Together with our findings, this suggests that lipids may mediate positional signals and modulate HD-Zip transcription factor activity elsewhere in plant development.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
KN: I think the most memorable moment was when I observed the outermost cell-specific ATML1-EGFP signal after heat-pulse treatment. This was an important moment when I was certain that our hypothesis was right.
And what about the flipside: any moments of frustration or despair?
KN: The purification of the START domain as a soluble form was challenging. I first tried to purify the full-length ATML1 protein as a soluble form, but I couldn’t obtain it, and ultimately it took over a year to obtain the soluble START domain.
What next for you after this paper?
KN: I am interested in how the ATML1 protein is broken down when VLCFA-Cers is absent – this will deepen our insights about lipid-mediated modulation of transcription factor activity in plants. On the other hand, from an evolutionary perspective, it is important to know whether lipid-transcription factor-based developmental mechanisms also work in the basal land plants or algae.
Where will this story take the Abe lab?
MA: As I mentioned in my biography, since my PhD I’ve been very interested in the molecular mechanism of epidermal cell differentiation. For the past 10 years or so, my lab has been focusing on florigen function and regulation, which involves FT, FD and FE. But I am very pleased that Kenji was interested in and restarted this project 5 years ago. In the future, Kenji and I hope to make exciting discoveries on epidermal cell differentiation.
Finally, let’s move outside the lab – what do you like to do in your spare time in Tokyo?
KN: I love to spend my spare time at the Onsen (a Japanese hot spring). After Onsen, I always drink a beer!
MA: In Spring 2019, our lab moved from Hongo to Komaba, which is a 20-min walk from Shibuya city. Shibuya city is a centre for modern culture and entertainment in Japan. I hope I will get to properly enjoy Shibuya life after the COVID-19 pandemic has settled down.
Development and Disease Models & Mechanisms welcome you to apply for our joint virtual meeting ‘Developmental Disorders: From Mechanism to Treatment‘, which aims to bring together developmental biologists, human geneticists and clinical researchers who are united in the goal of understanding and treating developmental disorders. The underlying causes of developmental disorders – genetic or environmental – are often not understood. Moreover, there is a disconnect between researchers working on animal models of developmental disorders, geneticists trying to identify the genomic lesion responsible, and clinicians hoping to treat affected patients. Given the resulting urgent need to improve communication between these groups, to promote basic research into congenital anomalies and to invest in translating this research to the clinic, this Meeting will focus specifically on building bridges from bench to clinic.
Find out more about the meeting from the organisers Phil Beales, James Briscoe, Monica J. Justice and Lee Niswander in the video below.
Speakers
Jeanne Amiel Institut Imagine, France Han Brunner Maastricht University Medical Center, The Netherlands Brian Ciruna The Hospital for Sick Children, Toronto, Canada Dagan Jenkins University College London, UK Nicholas Katsanis Rescindo Therapeutics Inc., USA Karen Liu King’s College London, UK Stefan Mundlos The Max Planck Institute for Molecular Genetics, Germany Emily Noël University of Sheffield, UK Eric Olson UT Southwestern Medical Center, USA Álvaro Rada-Iglesias IBBTEC, Spain David Rowitch University of Cambridge, UK Ian Smyth Monash Biomedicine Discovery Institute, Australia Lilianna Solnica-Krezel Washington University School of Medicine in St. Louis, USA Xin Sun University of California San Diego, USA Lori Sussel University of Colorado, USA
Organisers: Phil Beales, James Briscoe, Monica J. Justice and Lee Niswander Date: 14-17 September 2021 Abstract deadline: 16 July 2021 Location: Online
The Beddington Medal is the British Society for Developmental Biology’s major commendation to promising young biologists, awarded for the best PhD thesis in Developmental Biology that was defended in the year before the award.
The design of the medal, mice on a stylised DNA helix, is from artwork by Rosa Beddington herself.
This year, the Beddington Medal was awarded to Kristina Stapornwongkul, who did her PhD with Jean Paul Vincent at the Francis Crick Institute. Kristina presented her work at the BSDB/Genetics Society 2021 meeting and we caught up with her after the meeting to find out more about her life in science. Be sure to also check out the profile of Kristina – including a letter from JP Vincent and a list of Kristina’s selected publications – over on the BSDB site.
Where were you born and where did you grow up?
I was born in Giessen and grew up in Weil am Rhein. It’s a small town in Southwest Germany, directly at the border of France and Switzerland. It’s one of the sunniest places in Germany and I love to have a stroll through the vineyards whenever I go back there.
When did you first get interested in science?
I was always fascinated by technology and science, but I only developed a real passion for it in the last two years of high school. The realisation that each of our cells contains the information necessary to build an entire human, really blew my mind and made me want to learn more about how cells work.
How did you come to do a PhD in the lab of JP Vincent?
I think it all started with a zebrafish embryo and a stereo microscope. A few years later, at the end of my Masters studies, I knew that I wanted to do a PhD in the field of developmental biology. The UK has an outstanding developmental biology community and so I applied for the Wellcome Trust PhD programme in Developmental and Stem Cell Biology at UCL. As part of the programme, the students get the opportunity to rotate in three different labs. During my rotation, I realised that the Vincent lab was the perfect fit for me, both scientifically and personally.
Tell us about your PhD project: what were the main questions you were trying to answer?
The concept of morphogen gradient-mediated patterning has always fascinated me with its elegant simplicity: a single signalling molecule that can induce multiple cell fates depending on its concentration. How morphogen gradients form and what determines their shape are therefore important questions in order to understand how robust patterning is achieved in tissues. Several mechanisms by which morphogens might spread have been suggested over the years, with passive diffusion being the most parsimonious one. If such extracellular protein gradients form by simple diffusion, it shouldn’t be that difficult to engineer a morphogen gradient, no? At least that was the idea. So instead of further dissecting how natural morphogen gradients are generated, I wanted to test if an inert protein, such as GFP, could be transformed into a gradient-forming morphogen. Apart from probing whether diffusion is sufficiently reliable as a morphogen-transport mechanism, I was hoping that this synthetic approach would also help to uncover general principles and constraints that shape extracellular gradients. To do this work in vivo, I used the Drosophila wing pouch, one of the best studied model systems for morphogen gradient formation.
In your 2020 Science paper you describe your efforts to engineer a morphogen gradient, replacing Dpp with GFP. What did this technique reveal about how morphogens work?
The thing with engineering a synthetic morphogen gradient is that, even if it works, there is no guarantee that natural morphogens work exactly the same way. Nevertheless, our approach enabled us to show that protein gradients can, in principle, form by passive diffusion and that such gradients are reliable enough to pattern a tissue in vivo. It also made it feasible to specifically manipulate properties, such as binding affinities or expression levels, and test their effect on GFP gradient shape. Combining this with a modelling approach, we were therefore able to gain a good understanding of what each component was doing in our synthetic system.
Of course, we encountered several difficulties while building the GFP morphogen system and these were probably the most informative, because natural morphogen gradients that form by diffusion will encounter them as well. For instance, it became clear that secreted GFP can be lost from the tissue and end up in the larval blood, the hemolymph. This was really a big issue for the patterning performance of the GFP gradient. In fact, all secreted morphogens interact to a with components in the extracellar matrix and this is probably one important mechanism to regulate morphogenetic retention in epithelia.
If I had to summarise our findings in a sentence, I would probably say that a combination of high-affinity signalling receptors and low-affinity non-signalling receptors is sufficient to allow diffusing GFP to mimic the organising activity of a natural morphogen.
If you took one abiding memory with you from your PhD, what would it be?
As you can imagine this project involved many ‘trial and error’ experiments, and of course a good amount of luck. When you try to engineer something, it might not work for so many reasons. Even if your general design is good, expression levels might be too high or too low, or your synthetic receptor pair is not recycled efficiently (yes, that was an issue). So I think one of the most abiding memories of my PhD was when I saw for the first time that GFP in combination with GFP-responsive Dpp receptors was able to rescue growth and patterning of the fly wing pretty well. I expected a bit of a rescue, but I never thought that a two-component system could substitute that successfully an endogenous extracellular morphogen system, which not only consists of ligands and receptors but also of many extracellular regulators. My first thought was, ‘I must have messed up the genetics. The rescue is too good’. So, after checking everything three times, I went to JP and showed him the wing. His first response was, ‘Are you sure, you didn’t mess up the genetics?’.
You recently published a review making ‘the case for diffusion’. Why did you need to make this case?
Morphogen-mediated patterning has been studied extensively in a variety of model systems. However, the question of how morphogens spread in a tissue has remained quite controversial, especially in epithelial tissues. For instance, it has been suggested that diffusion is difficult to regulate and not reliable enough to generate robust extracellular gradients. As an alternative, active transport mechanisms, such as planar transcytosis or specialised filopodia (cytonemes), have been proposed. In our review, we try to give a comprehensive overview of the existing evidence from different model systems and conclude that there is strong evidence that morphogens disperse by diffusion-based mechanisms. In particular, we highlight how the tissue architecture and the ligand’s biochemical properties impose constraints on diffusion-based gradient formation and how components of the extracellular matrix help to overcome them.
So after your PhD you’ve recently moved to Barcelona: what are you doing there and how are you finding the city compared to London? I started as a postdoctoral fellow in Vikas Trivedi’s and Miki Ebisuya’s lab at EMBL Barcelona. We use aggregates of mouse embryonic stem cells as minimal model systems to study symmetry breaking and germ layer specification. London is amazing and definitely has a special place in my heart, but I have to admit that I am really in love with Barcelona. Being able to go for a swim after work and having tons of herbs that happily grow in the sun, is really amazing!
Longer term, do you know if you plan to stay in science?
Working in science is a huge privilege and I really appreciate the chance to interact with so many bright and inspiring people. Currently, I can’t imagine a more enjoyable job. However, being able to stay in science depends on many different factors and so I always try to stay open-minded.
Where do you think developmental biology will be in ten years?
I expect that we will have a much better understanding of the molecular mechanisms of human development. Already now, stem cell-based in vitro systems give us first insights into human organogenesis – a developmental stage in which functional studies were basically impossible before. As a consequence, we will probably also see a much stronger engagement of developmental biologists with the field of disease modelling.
Similarly, our research will depend less and less on the classical model systems. With CRISPR and stem cells, we will probably be able to widen our perspective on development by investigating anything from small insects to large mammals. I think it will be very exciting to see the differences and similarities we can find!
But I’m sure that’s not the only exciting direction developmental biology will take! I think developmental biology will be even more interdisciplinary (if that’s even possible) in ten years. Personally, I’m quite interested in the role of metabolism in development, but I’m sure there are also many more interesting intersections that we will further explore.
When you’re not in the lab, what do you do for fun?
I like to go climbing, swimming, hiking… pretty much all kinds of outdoor activities. Travelling is also a big passion of mine and I hope it will soon be possible again.
Some readers of the Node might already be familiar with preLights, another community site run by The Company of Biologists which aims to highlight new preprints from across the biological sciences. Most of the preLights community members are early-career researchers (PhD students and postdocs), but recently, preLights has found a new role as a teaching aid. Both the NYU Peer Review and Utrecht Protein Folding and Assembly courses have started using group preprint review projects as a tool to learn about critical reading and peer review. To learn more about how preLights has helped them teach these courses, preLights spoke to Gira Bhabha at NYU, and Tessa Sinnige at Utrecht University.

 (No Ratings Yet)
(No Ratings Yet)






{kind=link}