As I’m sure you’ve all heard, last week saw the 2020 Nobel Prize in Chemistry awarded to Emmanuelle Charpentier and Jennifer Doudna for their work on the CRISPR/Cas system. It’s hard to believe that it was only 8 or so years ago that they – along with their colleagues Martin Jinek, Krzysztof Chylinski and others – demonstrated the potential of the Streptococcus CRISPR/Cas9 adaptive defense system to be used for genome editing.
Whether you’re a fan of the Nobel or you think it’s over-rated, it’s hard to argue against the influence that CRISPR technology has had on the biomedical sciences, including developmental biology. Of course, it’s not that we couldn’t edit genomes before CRISPR came along – tools such as zinc-finger nucleases and TALENS were already proving useful to engineer mutations in a range of species – but the relative flexibility and ease of use of the CRISPR system opened up new possibilities for genome engineering both in traditional and non-traditional model systems.
Not only can targeted mutations now be made more quickly, efficiently and cheaply than we might have believed possible just a decade ago, but the precision with which specific edits to the genome can be made has also improved dramatically. Moreover, the CRISPR toolkit has been expanded to a wide range of other technologies – from genetic screening to manipulation of gene expression to barcoded lineage tracing.
We asked some of Development’s editors to sum up how CRISPR has impacted their research. Here are a few of their responses:
“CRISPR-based methods have become so ubiquitous that it is becoming difficult to remember how we were working in the pre-CRISPR era” (François Guillemot)
“Like many developmental biologists, CRISPR/Cas9 has revolutionised our research. Knocking-out a gene, easy-peasy; introducing a point mutation, no problem; deleting a regulatory element, piece of cake. It means we can design more creative and more precise experiments and we’re still only at the beginning of what is possible.” (James Briscoe)
“CRISPR/Cas9 has been a great addition to our tool box and substantially changed our life in the last 5-6 years. Genetic approaches in mice and cultured cells became so easy and became available for even non-standard organisms – the evo-devo field has perhaps been revolutionarily changed… Not only genetics, but also epigenetics: CRISPR/Cas9 technology is also applied to modify chromatin structures in highly targeted manner.” (Haruhiko Koseki)
“My postdoc knockout mouse took over two years to make. A student can make one in 3 weeks now.” (Benoit Bruneau)
To celebrate the Nobel award, Development has collected together just a few of the CRISPR papers published in the journal over the last 8 years. All these articles are free to read, and we invite you to browse the collection here. In it, you’ll find papers that apply CRISPR technologies in a wide range of model systems, that improve the efficiency with which mutations can be generated, and that use modified Cas9 proteins to manipulate gene expression or enhancer activity in a targeted manner. You’ll also find commentaries that touch on some of the thorny ethical questions that genome editing has thrown our way: how should we consider genome editing in the context of agriculture and genetically modified crops and – perhaps most difficult of all – what about human genome editing? We hope you enjoy this collection of articles.
And if you have comments on how CRISPR technologies have impacted your research, we’d love to hear them – please leave a comment below!
Timothy Fulton, Vikas Trivedi, Andrea Attardi & Benjamin Steventon
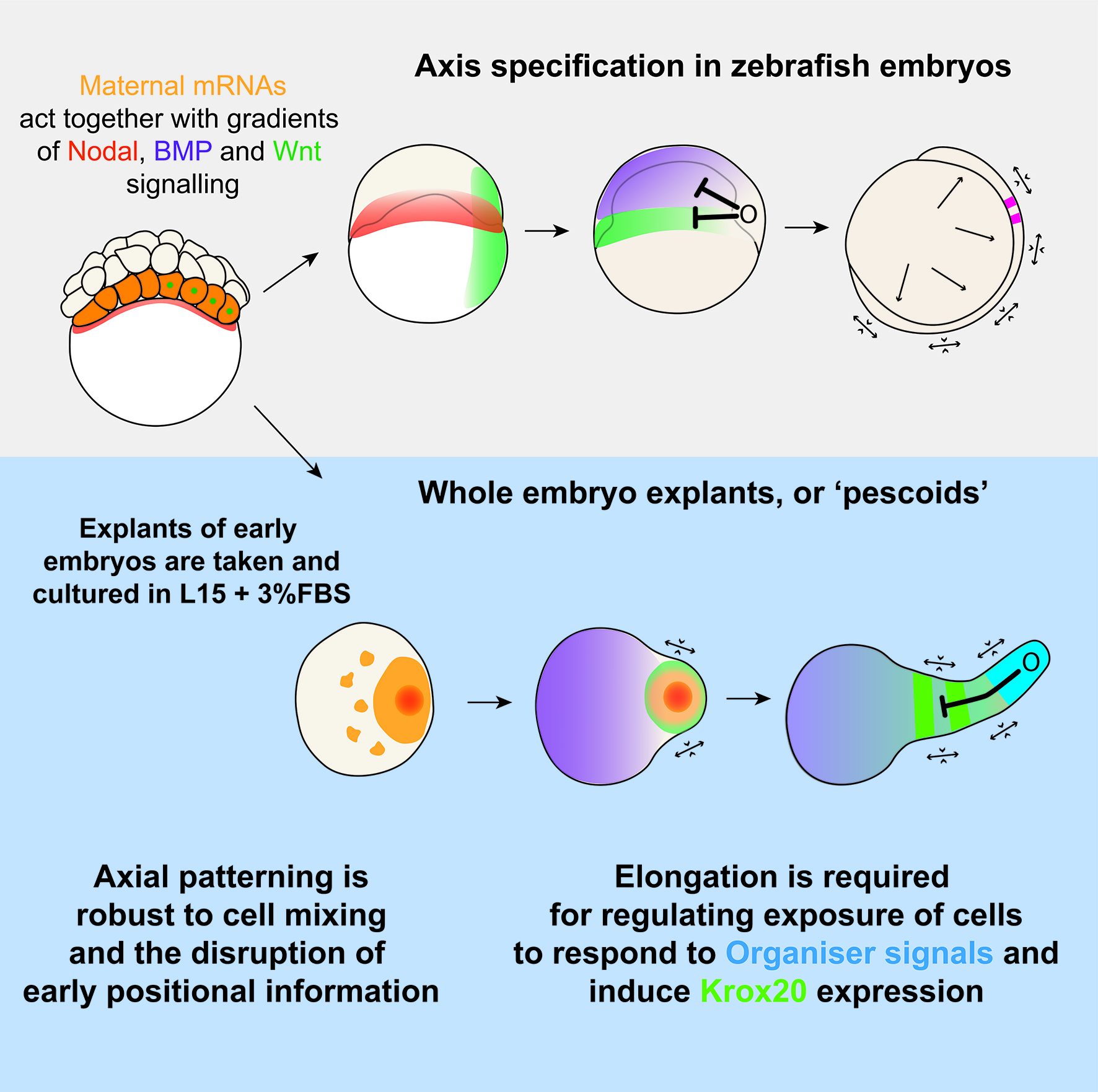
As developmental biologists, we often find ourselves carefully looking at developing embryos as they undertake a dramatic and fascinating task: making a complex organism out of a few initial embryonic cells. On the other hand, for engineers, building complex structures out of simple constituent parts under a defined set of rules is what creates excitement every day. Together, the bioengineer and the embryologist wonder how cells behave in a way that shapes the animal, whilst simultaneously deciding on their fate. Clearly, these two processes of morphogenesis and pattern formation must occur in concert.
We know that vertebrate embryos achieve this by integrating cues and signals from the extraembryonic environment. Such cues do not act alone though, as they are coupled to an intrinsic ability of early cells to drive morphogenesis and patterning. This latter feature of embryonic cells, known as self-organisation, is remarkably highlighted when cells are separated from their extra-embryonic constraint. An example of this is given by mouse and human embryonic stem cells (mESCs/iPSCs). Under controlled culturing conditions, these cells aggregate as ‘gastruloids’ (van den Brink et al., 2014; Moris et al., 2020), groups of cells that perform and recapitulate essential aspects of axis specification and pattern formation seen in intact mouse or human embryos.
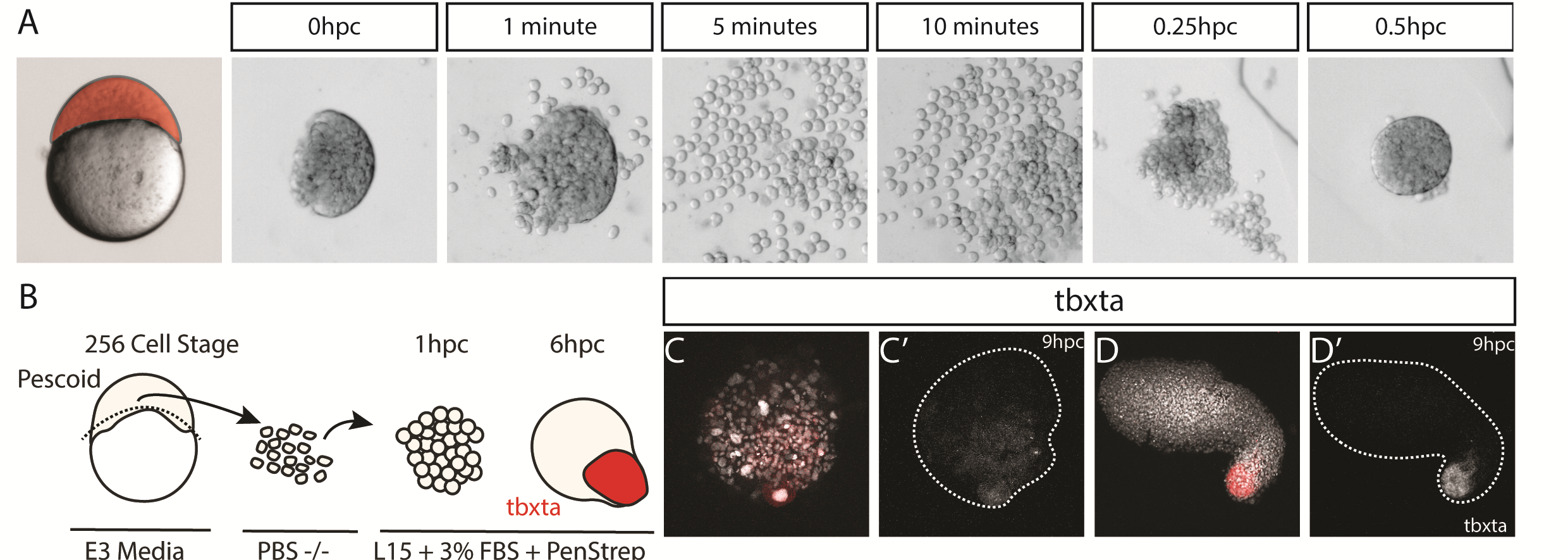
Such ideas about self-organization of embryonic cells were in the atmosphere during lab meetings and tea time discussions at the Department of Genetics. Vikas, who had recently joined the lab of Alfonso Martinez Arias as a postdoctoral fellow, came with the experience of having worked with zebrafish before and was wondering about a fish equivalent of gastruloids. At the same time, Andrea, a masters student in Ben Steventon’s lab, while working on a project to lineage trace Neuromesodermal Progenitors during gastrulation, was having similar thoughts (Attardi et al., 2018). The question arose quite spontaneously: could zebrafish embryonic cells tell us something new about morphogenesis and self-organisation, if we cultured them free of their embryonic constraints? That looked like a good opportunity for exploring what the collaboration of bioengineering and embryology could lead us to. We therefore decided to attempt culturing zebrafish embryonic cells at early stages of development, starting by explanting cells from the 256 cell stage embryo, prior to the midblastula transition. Strikingly, we observed that in doing so, these masses of cells elongated over a period of seven to ten hours. We observed this phenomenon in a range of media, with and without serum, including inert Ringer’s Solution. These results nicely echo those of Jane Oppenheimer (1936), who found similar results when explanting blastoderm from another teleost, Fundulus heteeroclitus.
Pescoid Elongating – Wildtype pescoid elongating in normal L15 + 3% FBS Serum imaged on a widefield microscope
After identifying optimal incubation conditions, Tim set about understanding which cell types formed within these elongating explants. After making many hundreds of pescoids, and using multiplex in situ Hybridisation Chain Reaction (Choi et al., 2018), he demonstrated that these explants produce all three germ layers in their elongated form. We observed the mesodermal marker tbxta expressed in the elongating pole, with sox17 marking the endoderm in the core of the explants. In the non-elongating end, we observed sox2 expressed marking the ectoderm. In addition to these marker genes, we also observed expression of germ cell markers nanos and vasa, as well as expression of the hindbrain marker gene krox20 which, as in the embryo, was expressed in two clearly defined stripes of expression in the non-elongating end. Interestingly, we did not observe expression of otx2 positive forebrain cell.
Figure 1 – In Situ Hybridisation Chain Reaction demonstrates all three germ layers form in 256 cell stage embryonic explants, cultured for 7-10 hours. (A) Schematic of the experimental protocol. (B-F) Tbx16::GFP live reporter demonstrates the elongation end is comprised of mesoderm, which is confirmed by (C-K) HCR for tbxta. (L-L’) Sox2, representing ectoderm, is expressed in the non-elongating end and (M) Sox32, representing endoderm is expressed centrally though the core of the pescoid. (N) pesoids also demonstrate some level of mesoderm patterning with gata4 in the non-elongating end, and noto expressed in the elongating end. (O) Germ cells, expressing nanos and vasa are found deep inside the central core of the pescoid. (P-Q) the two ends of the pescoid after elongation are marked by activity of BMP in the non-elongating end, and TCF/Wnt in the elongating end. The region low in both BMP and Wnt activity (R) express two clear stripes of krox20.
We wanted to test exactly how robust the germ layer formation was to mixing of the cells. In order to test this, we cut and totally dissociated and reaggregated the explants immediately after explanting using two eyelashes glued to capillary tubes. Tim and Ben swept the cells together gently under a microscope in order to break down and reassemble the explant. A very delicate procedure, made harder by Ben’s insistence that we listened to Test Match Special, in order to keep up to date with the Cricket World Cup! When we reaggregated pescoids, we observed that the explants rarely elongated, however the majority still expressed tbxta. We hypothesised that the initial size of the explant is important for elongation potential, as quartered explants rarely elongate, however half sized explants retain this potential.
Figure 2 – Mesoderm formation is robust to experimentally introduced cell mixing. (A-B) by dissociating and reaggregating pescoids, mesoderm is still observed being formed, measured through expression of tbxta. (C) The majority of pescoids do not elongate but still express tbxta (D) however if sufficient cells are reaggregated, elongation and mesoderm formation is observed.
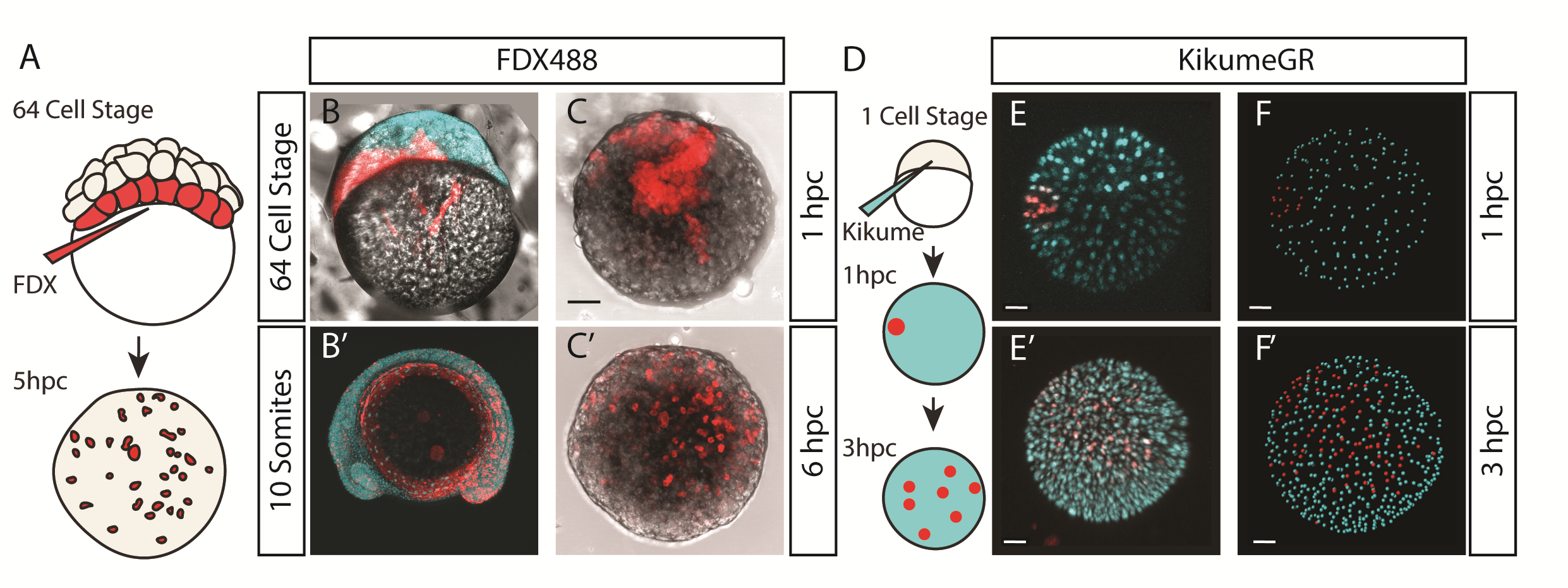
We were also interested to discover how much cell mixing was occurring in these explants naturally. Using two different methods, Chaitanya labelled either the edge of the explant, or the marginal cells of the embryo prior to explanting, and then tracked these cells post explant. In all situations, we observed that the labelled cells mix significantly with the non-labelled cells prior to elongation. Schauer et al. (2020) recently conducted very similar experiments to us, and demonstrated that the prepatterns provided to the embryonic cells by the yolk prior to the stage at which we take explants are essential for the formation of the elongation. We also found that the whole process is dependent on early Nodal activity, a result that fitted nicely with the work of (Williams and Solnica-Krezel, 2020).
Nodal in Explants – Activin Response Element::GFP reporter for Nodal activity demonstrating correlation of Nodal signalling with the elongation
We had some excellent discussions with Alexandra Schauer at the EDBC meeting in Alicante in 2019 and really enjoyed being able to share and discuss our findings with one another. However, the extensive mixing we observe here demonstrates that whatever the mechanism between prepattern and pattern formation is, it must also be highly robust to cell mixing over long timescales of 3 to 5 hours. Overall the individual cells regularly swap positions with one another, however, at the organism level, the explants form reproducible domains of stable gene expression.
Figure 3 – Mesoderm formation is robust to extensive cell mixing. (A-C) Through labelling the marginal blastomeres at the 64 cell stage using fluorescent dextran, and then cutting the explant as normal at the 256 cell stage, labelled cells were observed spread across the entire explant by 6 hours of culture, prior to elongation. (D) This result was repeated by labelling the edge of a pescoid immediately after cutting using photoconvertible kikume protein. These labelled cells were observed scattered across the entire pescoid within 3 hours.
How these different domains of gene expression came about remained unclear, and to investigate this, we looked at the signalling events taking place within the explant. We observed that initially the activity of Wnt and BMP signalling completely overlapped. During elongation however, these two activity domains separate, with Wnt activity being restricted to the elongation, and BMP to the non-elongating end. We hypothesized that the elongation may be the driving force behind the separation of these two domains, and by inhibiting morphogenesis both genetically and pharmacologically, we demonstrated that this is indeed the case; without morphogenesis, the two domains remain overlapping and two krox20 stripes were never observed. The explants however are unable to produce an environment with very low levels of both Wnt and BMP, and therefore this may explain why Oxt2 positive cells are not produced.
Pescoids Elongating with TCF::GFP and BRE::RFP. TCF::GFP reporter for Wnt activity, demonstrating Wnt signalling occurs across the entire explant their localises to just the elongation (Green to White). BMP Response Element fusion of RFP demonstrates BMP signalling is active across the entire explant then localises to just the non-elongating end.
We therefore had a situation where our explants are displaying a very high level of cell mixing, with cells regularly changing positions with one another, whilst also maintaining distinct regions of gene expression. Whilst our work here begins to answer how these domains form, it does not answer how these domains are then maintained. This opens an intriguing question: how do cells, via signalling and gene regulatory networks, update the cell’s positional information whilst constantly moving?
Traditionally, this type of question would invoke ideas of the French Flag Model, which describes a situation where morphogen gradients inform cells of their position and, therefore, of their fate decision. However, as Wolpert described in the French Flag Problem (Sharpe, 2019; Wolpert, 1969), this idea becomes difficult to apply when cells also regularly change positions with one another and therefore scramble the pattern as it is being produced. This is something which Tim is continuing to investigate as part of his PhD. Using live imaging data and tracking of individual cells, we are overlaying dynamical systems models of a GRN onto in vivo tracks in order to model cell fate decisions in a tissue undergoing active morphogenesis. The Steventon Lab is also interested in investigating how pattern formation may be an emergent property of gene regulatory networks and morphogenesis working together. Using the idea of tectonic plates, the term tissue tectonics (Busby and Steventon, 2020) describes how morphogenetic movements regulate the timings of interactions between different tissues as they slide together or apart, similar to as we see in these pescoid explants, and other embryonic systems. The Trivedi lab is pursuing studies on understanding the biophysics of self-organization that leads to patterning in early embryonic systems including pescoids.
Figure 4 – Gastrulation in zebrafish occurs through interaction of cell movements with signalling gradients. These gradients themselves are a product of maternally derived mRNA and Nodal ligands. The entire gastrulation process occurs around the yolk, resulting in convergence and extension movements separating different domains, thereby patterning the embryo. Without the yolk, these events all still occur, however convergence and extension now pulls the elongation away from the main pescoid body, thereby producing a rudimentary patterned axis.
Overall, this work, and the ideas of tissue tectonics propose a model where pattern formation is as much the output of genes and local cell rearrangements, as it is the more global tissue movements which shift cells into and out of range of one another. By considering patterns as an emergent property of morphogenesis and GRNs, we can begin to ask questions about how these components are brought together, and what role cell-cell signalling plays in this.
Faculty of Health and Medical Sciences
University of Copenhagen
Institute: The Novo Nordisk Foundation Center for Stem Cell Biology – DanStem is located at the University of Copenhagen. DanStem addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new strategies and targets for the development of new therapies for cancer and chronic diseases such as diabetes and liver failure. Find more information about the Center at https://danstem.ku.dk/.
Job description
The Serup group is looking for a talented postdoc with experience in stem cell biology, NGS-based methods and bioinformatics. Recently, we found that gene regulatory networks downstream of Notch signaling that regulate pancreatic cell fate decisions were highly dynamic and more complex than previously anticipated (Seymour et al., Developmental Cell 2020). Importantly, we found that the core Notch effector, HES1 is oscillating and regulates cell fate by inhibiting entire gene regulatory networks downstream of bHLH master regulators, and this project continues our ongoing efforts to understand the molecular basis for these observations. We identify transcription factor target genes and explore target gene regulation by ChIP-seq and mutagenesis followed by RNA-seq, using a combination of different model systems for pluripotent stem cell culture, in vitro organ/organoid culture as well as in vivo mouse models. This project will involve human ES cell differentiation, NGS-based methods and bioinformatics analysis.
We are seeking a highly motivated and ambitious candidate with experience in NGS-based methods and bioinformatics analysis with a professional profile that closely matches the qualifications below:
The candidate is required to hold a PhD degree in stem cell/developmental biology or molecular biology.
The candidate should have extensive experience in NGS-based methods and bioinformatics.
Techniques such as human ES cell differentiation protocols, 3D culturing and flow cytometry is an advantage.
A relevant publication record is essential.
Terms of employment
The fulltime employment is for 2 years with a possibility of extension and scheduled to start 1 February 2021 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen.
Salary, pension and terms of employment will be in accordance with the agreement between the Ministry of Finance and AC (Danish Confederation of Professional Associations). Currently, the monthly salary starts at 34,650 DKK/ approx. 4,650 Euro (April 2020-level). Depending on qualifications, a supplement may be negotiated. The employer will pay an additional 17.1 % to your pension fund.
Non-Danish and Danish applicants may be eligible for tax reductions, if they hold a PhD degree and have not lived in Denmark the last 10 years.
The position is covered by the “Memorandum on Job Structure for Academic Staff at the Universities” of 19 December 2020.
Questions
For further information, please contact Professor Palle Serup, palle.serup@sund.ku.dk.
Foreign applicants may find this link useful: www.ism.ku.dk (International Staff Mobility).
Application procedure
Your online application must be submitted in English by clicking ‘Apply now’ below. Furthermore your application must include the following documents/attachments – all in PDF format:
Motivated letter of application (max. one page).
CV incl. education, work/research experience, language skills and other skills relevant for the position.
A certified/signed copy of a) PhD certificate and b) Master of Science certificate. If the PhD is not completed, a written statement from the supervisor will do.
List of publications.
Application deadline: 15 November 2020, 23.59pm CET
We reserve the right not to consider material received after the deadline, and not to consider applications that do not live up to the abovementioned requirements.
The applicant will be assessed according to the Ministerial Order no. 242 of 13 March 2012 on the Appointment of Academic Staff at Universities.
The further process
After the expiry of the deadline for applications, the authorized recruitment manager selects applicants for assessment on the advice of the hiring committee. All applicants are then immediately notified whether their application has been passed for assessment by an unbiased assessor. Once the assessment work has been completed each applicant has the opportunity to comment on the part of the assessment that relates to the applicant him/herself.
The applicant will be assessed according to the Ministerial Order no. 242 of 13 March 2012 on the Appointment of Academic Staff at Universities.
Interviews are expected to be held in week 49-50.
University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates regardless of age, disability, gender, nationality, race, religion or sexual orientation. Appointment will be based on merit alone.
Tenure-track Assistant or Associate Professor of Genetics or Genomics
In anticipation of substantial growth over the next five years, the Department of Human Genetics at the University of Utah School of Medicine (www.genetics.utah.edu) seeks outstanding applicants for one or more tenure-track positions at the level of Assistant or Associate Professor. We seek highly creative scientists who use genetics to investigate fundamental biological problems. We encourage applicants whose research focuses on evolutionary and functional genetics and genomics; human and medical genetics; computational genomics; and research programs using established model or unconventional organisms. As part of a vibrant community of faculty with a strong track record of collaborative mentorship, research, and funding, the Department of Human Genetics lies at the interface between basic and clinical sciences. This creates ample opportunities for interdisciplinary research (e.g., our Center for Genetic Discovery, Transformative Excellence Program in Evolutionary Genetics and Genomics, and Center for Genomic Medicine). As a department, we value diversity and equity, and believe that the best science is done when researchers of diverse backgrounds are integrated and supported in an inclusive manner. We seek faculty who share these values. Our institution is set in a unique geographical landscape that attracts a heterogeneous and productive scientific community. Successful candidates will receive a generous startup package and enjoy a stimulating research environment that places a strong emphasis on innovation and interaction.
Curriculum Vitae – CV and 3 most relevant reprints or preprints
Research statement – Describe your most significant scientific accomplishments, your goals for research as a faculty member, and the qualifications and experience that have prepared you to achieve these goals.
Teaching statement – Describe your commitment to education, your teaching philosophy, and the courses you might potentially teach.
Diversity, equity, and inclusion statement — Describe your past and future contributions to diversity, equity, and inclusion through research, teaching, and service.
The University of Utah is an Affirmative Action/Equal Opportunity employer and does not discriminate based upon race, national origin, color, religion, sex, age, sexual orientation, gender identity/expression, status as a person with a disability, genetic information, or Protected Veteran status. Individuals from historically underrepresented groups, such as minorities, women, qualified persons with disabilities and protected veterans are encouraged to apply. Veterans’ preference is extended to qualified applicants, upon request and consistent with University policy and Utah state law. Upon request, reasonable accommodations in the application process will be provided to individuals with disabilities. To inquire about the University’s nondiscrimination or affirmative action policies or to request disability accommodation, please contact: Director, Office of Equal Opportunity and Affirmative Action, 201 S. Presidents Circle, Rm 135, (801) 581-8365.
The University of Utah values candidates who have experience working in settings with students from diverse backgrounds, and possess a strong commitment to improving access to higher education for historically underrepresented students.
Yesterday over two hundred people from all over the wall tuned in to the first instalment of Development presents…, Development’s new webinar series. For those who missed it, here you can watch the talks, plus their following Q&A sessions moderated by Development Editor-in-Chief and webinar chair James Briscoe.
The Dunn School of Pathology at the University of Oxford seeks an outstanding candidate for the position of Associate Professor of Cell and Molecular Biology.
The Dunn School has an outstanding track record in biomedical research, with a strong tradition in studying molecular and cellular mechanisms that underlie disease. The Associate Professor will be expected to develop a research programme in any aspect of fundamental or translational cell and molecular biology, although preference will be given to candidates whose research complements existing strengths at the Dunn School.
In this episode we’re taking a road trip from Philadelphia to Baltimore, exploring stories of chromosomal cut-and-paste, cancer cures and Henrietta Lacks’ incredible cancer cells.
Studying human cells in the lab allows us to understand more about how they work, investigate the causes of disease, and design new treatments. But growing cells in the lab isn’t as easy as you might think. We explore the story of how Henrietta Lacks’s immortal cells (known today as HeLa cells) became the go-to human cell line for biomedical research, and the impact they’ve had as a result.
We also tell the story of the ‘Philadelphia chromosome’ (a key cause of chronic myeloid leukaemia), how it was discovered and how it’s discovery influenced the search for a cure.
Journal of Cell Science, published by The Company of Biologists, is seeking enthusiastic and motivated applicants for the role of Features & Reviews Editor.
Joining an experienced and successful team, including Editor-in-Chief Michael Way, this is an exciting opportunity for a talented scientific editor to make a significant contribution to an important journal in the field of cell biology. Journal of Cell Science publishes outstanding primary research articles, reviews and topical comment, and continues to expand its services to authors and readers.
Applicants should have a PhD, and ideally postdoctoral experience, in cell biology or a relevant scientific field, and a broad knowledge of cell biology. The successful candidate will have strong communication, networking and interpersonal skills. We are also looking for enthusiasm, motivation, commitment, and a broad interest in science and the scientific community. Editorial experience is preferable but not essential.
Core responsibilities include:
Commissioning, handling peer review and developmental editing of review-type content
Maintaining and developing the ‘Cell Scientists to Watch’ interviews section of the journal
Writing content for the Research Highlights section of the journal
Representation of the journal at local and international conferences and within the wider scientific community
Creative involvement in the journal’s development and marketing activities
Additional responsibilities may be available for the right candidate. The Features & Reviews Editor will work alongside both an experienced Executive Editor and Senior Editor. This is a permanent, full-time position, and is based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open. We take great pride in the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should be eligible to work in the UK and are requested to send to recruitment@biologists.com: a CV; a 400-word summary of a recent ground-breaking development in cell biology; and a cover letter explaining their interest in the post. Initial application deadline is 2 November 2020, but we will consider applications on a rolling basis so encourage candidates to apply as soon as possible.
Journal of Cell Science and its publisher The Company of Biologists are seeking to appoint a new Community Manager to run our newly launched microscopy community website, FocalPlane. This is initially offered as a two-year fixed term contract with the expectation that as the project develops the role would become permanent in the future.
Launched in 2020, FocalPlane is a curated and centralised platform for the microscopy community to share news and techniques, discuss issues relevant to the field and read about the latest research and events. We are now looking for an enthusiastic and motivated person with fresh ideas and a willingness to learn to join us to develop and maintain this site.
Core responsibilities of the position include:
Creating and commissioning content for FocalPlane, including writing posts and soliciting content from the academic community, societies, companies and other organisations
Providing user support and ensuring site functionality on a day-to-day basis
Providing creative and practical input into the development of the site
Maintaining and developing the site’s presence on social networking sites such as Facebook and Twitter
Representing Journal of Cell Science and FocalPlane at international conferences
Essential skills:
PhD in a relevant scientific field, ideally with experience of microscopy
Willingness to grow and develop knowledge of microscopy
Demonstrable ability to write for an online audience and/or produce social media content
Clear understanding of the online environment as it applies to scientists
Excellent interpersonal and communication skills
Strong networking abilities online and in person
Desirable:
Experience with additional media (e.g. video or podcasting)
Experience with WordPress
Contacts within the microscopy community
This is an exciting opportunity to develop a hub for the microscopy community – in a similar vein to the Company’s established community site for developmental biologists, the Node – and to engage with relevant people at all levels: academics, developers, facilities, institutes and companies. The Community Manager will work alongside an experienced in-house team, including the Executive Editor of Journal of Cell Science. Additional responsibilities may be provided for the right candidate.
The Company of Biologists exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV along with a covering letter that summarises their relevant experience, and in particular any specific microscopy/image analysis skills, and includes links to any online activities, salary expectations, and details about why they are enthusiastic about this opportunity.
Applications and informal queries should be sent by email to recruitment@biologists.com. We may request written tests in advance of any interview.
The Childs lab studies developmental angiogenesis and vascular stabilization using the zebrafish model. We are looking for an outstanding postdoc to probe the mechanism of genetic vascular malformation development. We are located at the University of Calgary, Canada. Applicants should be within 3 years of their PhD degree and have a demonstrated track record of success in publications. Please send a letter of interest, your CV and the names of 3 potential referees to schilds@ucalgary.ca.
 (3 votes)
(3 votes)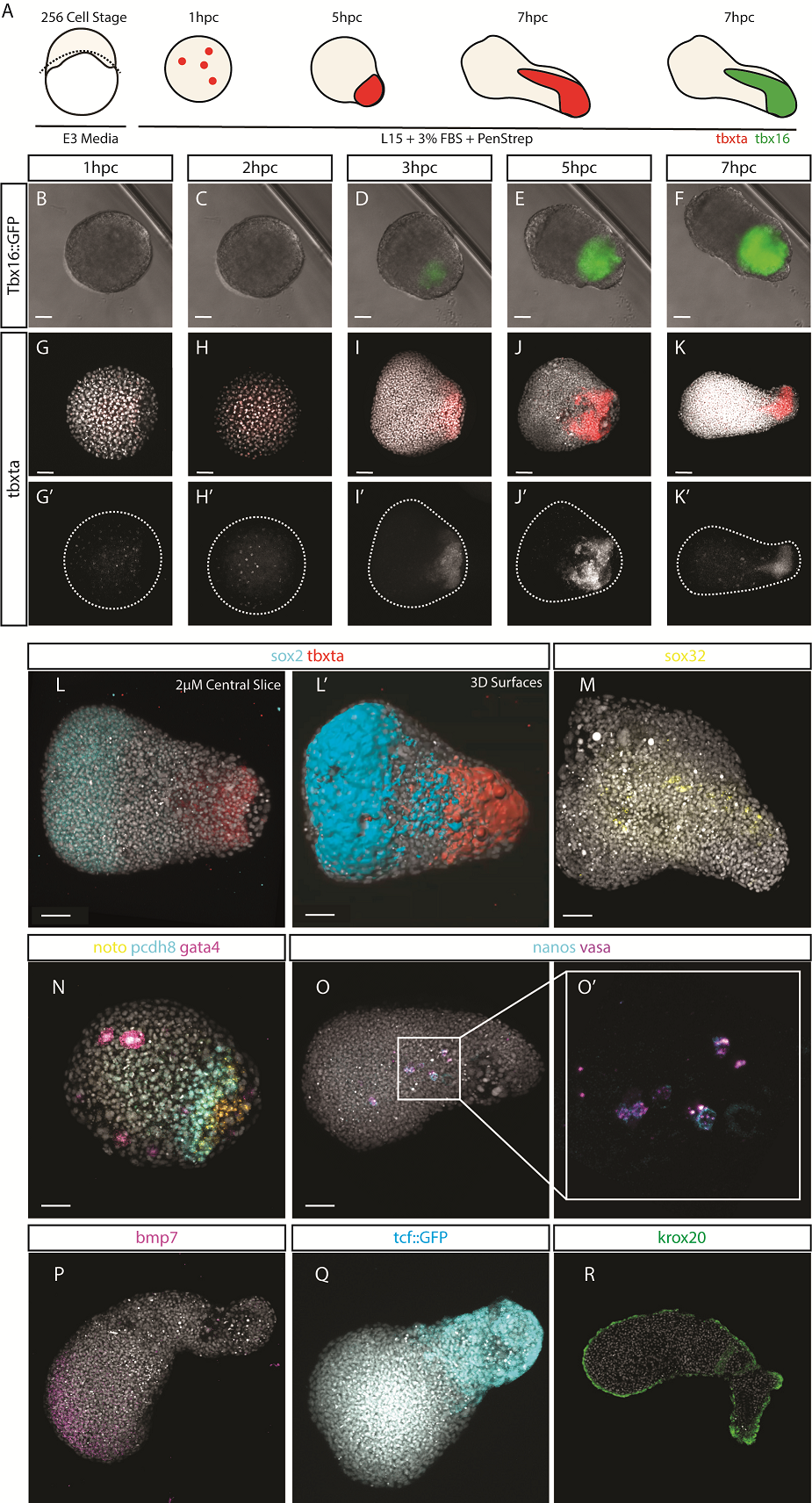
 (2 votes)
(2 votes)