Location: The Francis Crick Institute, Midland Road, London
Contract: Fixed-term, 4 years, full time
Salary: Competitive with benefits, subject to skills and experience
Vacancy ID: 12287
Short summary
Dr Niakan’s laboratory focuses on understanding the mechanisms of lineage specification in human embryos and the derivation of novel human stem cells. Details of research projects currently being undertaken can be seen at: http://www.crick.ac.uk/kathy-niakan
Research techniques used in the laboratory include: molecular biology, advanced microscopy and image quantification, human and mouse preimplantation embryo culture and micromanipulation, genome modification, genome-wide techniques including single-cell RNA-sequencing, multi-omics analysis and human embryonic and induced pluripotent stem cell derivation.
We seek candidates who are energetic, focused, motivated, productive and collaborative with a desire to work in a congenial, dynamic, and collaborative research environment. Good organisational, analytical, and communication skills are essential.
Project scope
The aim of the project is to characterise early lineage specification in human preimplantation embryos. We aim to generate reporter embryos to perform in vivo lineage tracing to elucidate developmental trajectories of individual cells. The project will explore a range of single cell, imaging and genome editing techniques to understand early lineage specification mechanisms in human embryos. This knowledge will provide fundamental insights into human biology.
About us
The Francis Crick Institute is a biomedical discovery institute dedicated to understanding the fundamental biology underlying health and disease. Its work is helping to understand why disease develops and to translate discoveries into new ways to prevent, diagnose and treat illnesses such as cancer, heart disease, stroke, infections, and neurodegenerative diseases.
An independent organisation, its founding partners are the Medical Research Council (MRC), Cancer Research UK, Wellcome, UCL (University College London), Imperial College London and King’s College London.
The Crick was formed in 2015, and in 2016 it moved into a new state-of-the-art building in central London which brings together 1500 scientists and support staff working collaboratively across disciplines, making it the biggest biomedical research facility under in one building in Europe.
The Francis Crick Institute is world-class with a strong national role. Its distinctive vision for excellence includes commitments to collaboration; developing emerging talent and exporting it the rest of the UK; public engagement; and helping turn discoveries into treatments as quickly as possible to improve lives and strengthen the economy.
If you are interested in applying for this role, please apply via our website.
The closing date for applications is 01 November 2019 at 23:45.
All offers of employment are subject to successful security screening and continuous eligibility to work in the United Kingdom.
Dorsal view of the left hand of a 10-week old human embryo. The dorsometacarpales are highlighted: these muscles (like others described in this study) are present in adults of many other limbed animals, while in humans they normally disappear or become fused with other muscles before birth. CREDIT: Rui Diogo, Natalia Siomava and Yorick Gitton
A team of evolutionary biologists, led by Dr. Rui Diogo at Howard University, USA, and writing in the journal Development, have demonstrated that numerous atavistic limb muscles – known to be present in many limbed animals but usually absent in adult humans – are actually formed during early human development and then lost prior to birth. Strikingly, some of these muscles, such as the dorsometacarpales shown in the picture, disappeared from our adult ancestors more than 250 million years ago, during the transition from synapsid reptiles to mammals.
Also remarkably, in both the hand and the foot, of the 30 muscles formed at about 7 weeks of gestation one third will become fused or completely absent by about 13 weeks of gestation. This dramatic decrease parallels what happened in evolution and deconstructs the myth that in both our evolution and prenatal development we tend to become more complex, with more anatomical structures such as muscles being continuously formed by the splitting of earlier muscles. These findings offer new insights into how our arms and legs evolved from our ancestors’, and also about human variations and pathologies, as atavistic muscles are often found either as rare variations in the common human population or as anomalies found in humans born with congenital malformations.
Since Darwin proposed his evolutionary theory, scientists have argued that the occurrence of atavistic structures (anatomical structures lost in the evolution of a certain group of organisms that can be present in their embryos or reappear in adults as variations or anomalies) strongly supports the idea that species change over time from a common ancestor through “descent with modification”. For example, ostriches and other flightless birds have vestigial wings, while whales, dolphins and porpoises lack hind limbs but their embryos initiate and then abort hind limb development. Similarly, temporary small tail-like structures are found in human embryos and the remnant of the lost ancestral tail is retained as our coccyx. Researchers have also suggested that atavistic muscles and bones can also be seen in human embryos, but it has been difficult to visualize these structures clearly, and the images that appear in modern textbooks are mainly based on decades old analyses.
This is changing with development of new technology that provides high-quality 3D images of human embryos and fetuses. In the new study published in the journal Development the authors have used these images to produce the first detailed analysis of the development of human arm and leg muscles. The unprecedented resolution offered by the 3D images reveals the transient presence of several of such atavistic muscles. Dr. Diogo said: “It used to be that we had more understanding of the early development of fishes, frogs, chicken and mice than in our own species, but these new techniques allow us to see human development in much greater detail. What is fascinating is that we observed various muscles that have never been described in human prenatal development, and that some of these atavistic muscles were seen even in 11.5-weeks old fetuses, which is strikingly late for developmental atavisms “.
He further added: “Interestingly, some of the atavistic muscles are found on rare occasions in adults, either as anatomical variations without any noticeable effect for the healthy individual, or as the result of congenital malformations. This reinforces the idea that both muscle variations and pathologies can be related to delayed or arrested embryonic development, in this case perhaps a delay or decrease of muscle apoptosis, and helps to explain why these muscles are occasionally found in adult people. It provides a fascinating, powerful example of evolution at play.”
Cells are the building blocks of life. However, in multi-cellular organisms, millions of cells are subject to death due to injury, infection and ordinary cell turnover (Galluzzi et al., 2018). For example, epithelial cells in the small intestine rapidly renew every 2 to 6 days in most mammals, which is crucial to maintain proper function of villus epithelia (Mayhew et al., 1999). Moreover, during embryonic development, cell death serves as a crucial mechanism to remove unnecessary cells, adjust tissue size and shape, as well as correct developmental errors (Arya and White, 2015). In order to maintain homeostasis and avoid unwanted inflammatory responses, cellular debris is usually cleared rapidly by professional phagocytes, such as macrophages and microglia. However, during early neurogenesis when the neural tube develops and large numbers of neurons and glia undergo apoptosis, myeloid-derived professional phagocytes have not yet infiltrated the trunk of developing embryos (Herbomel, Thisse and Thisse, 1999, 2001; McGrath et al., 2003; Bertrand et al., 2013; Stremmel et al., 2018). How dead cells are removed from this region during early development remained largely unknown until recently.
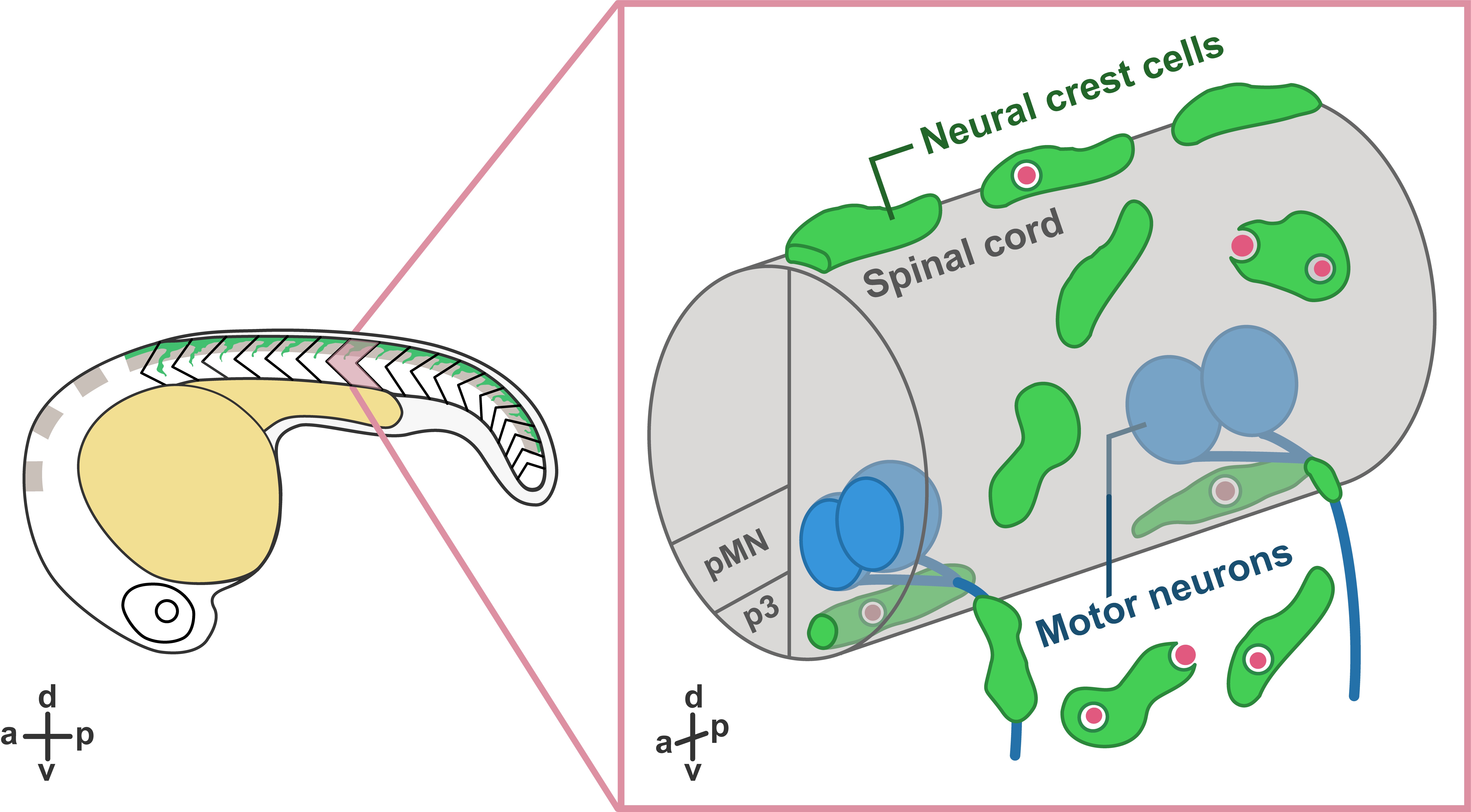
In our recent paper entitled “Migratory Neural Crest Cells Phagocytose Dead Cells in the Developing Nervous System”, we demonstrate an unexpected behavior of neural crest cells (NCCs), which have been studied for more than 150 years, in debris clearance before the colonization of professional phagocytes (Zhu et al., 2019) (Figure 1).
Figure 1. Schematic illustration showing neural crest cells (green) phagocytose dead cells (red) in zebrafish embryos. Right panel shows zoomed view of the trunk.
Interestingly, not only did we not expect this novel function of NCCs, but the discovery was also made by accident! Our original intent was to study how glial populations coordinated their behaviors during early spinal motor nerve development. But early in our studies, Yunlu started to notice NCCs behaving in a manner that didn’t match what the literature described. Some migrated away from their migratory streams, and others appeared to interact with cellular debris. Here, we would like to share the story behind this work.
Neural Crest Cells Clear Debris
What are NCCs and what do we know about them? During the earliest stages of vertebrate nervous system development, the ends of neural plate rise and fold into the neural tube, which further develops into the central nervous system (CNS), including the brain and the spinal cord. During or after the closure of the neural tube, NCCs located at the edge of the neural fold go through an epithelial-to-mesenchymal transition and delaminate from the dorsal neural tube. Multipotent NCCs then migrate through highly conserved pathways and give rise to a variety of cell types, including skeletal tissues, pigment cells, and neurons and glia of the peripheral nervous system (PNS) (Mayor and Theveneau, 2013). Given that NCCs are highly migratory and colonize the entire developing embryo before the infiltration of professional phagocytes, we hypothesized that they may have the ability to clear cellular debris at early developmental stages.
To examine whether NCCs are capable of clearing debris, we performed in vivo, time-lapse imaging in transgenic embryos expressing fluorescent proteins in NCCs. Excitingly, in these movies, we observed that some fluorescently-labeled NCCs migrated towards dead cells located far away from their innate migratory pathways. Then, they reached toward dead cells and engulfed them, resulting in the formation of large engulfment vesicles inside NCCs.
To confirm that the NCC engulfment process was similar to phagocytosis in professional phagocytes, we imaged transgenic lines labeling early endosomes (PI(3)P sensor) and lysosomes (Lamp1-GFP) (Rasmussen et al., 2015). In these movies, we found that, similar to the maturation of phagosomes, NCC engulfment vesicles fused with early endosomes and lysosomes, leading to progressive acidification inside the vacuoles. Interestingly, we observed NCCs phagocytosing a variety of dead cells, including dead NCCs, neuronal debris, and muscle cells.
In these movies, we also observed that PNS NCCs migrated through motor exit point transition zones, where spinal motor neurons send their axons into the PNS, into the ventral neural tube, and phagocytosed CNS debris. Most of these CNS-located NCCs stayed inside the neural tube for 2 to 12 hours and returned to the PNS through motor exit point transition zones. To test whether PNS NCCs were recruited into the spinal cord by cellular debris, we induced CNS cell death by ablating radial glia using nitroreductase-mediated cell death (Smith et al., 2014; Johnson et al., 2016) and observed a significant increase in the number NCCs recruited into the CNS compared to control embryos. Therefore, we conclude that NCCs can migrate toward and phagocytose dead cells in both CNS and PNS.
Are There Subpopulations of Neural Crest Cells?
Interestingly, we noticed dead cells were not always cleared by the nearest NCC. Instead, under most circumstances, they were phagocytosed by NCCs that came from a distance. And that led us to wonder if phagocytic NCCs were a specific subgroup of the neural crest population. To examine whether phagocytic NCCs belonged to a specific lineage, we performed lineage tracing on phagocytic NCCs expressing a photoconvertible protein. Contrary to our expectations, we observed phagocytic NCCs differentiate into a variety of derivatives, including pigment cells, motor axon-associated cells, and dorsal root ganglia cells, suggesting that phagocytic NCCs are not lineage restricted.
Additionally, under physiological conditions, we only observed 5-10% of NCCs that were phagocytic. To examine whether more NCCs could be “activated”, we induced cell death using laser ablation. Surprisingly, immediately after the ablation, the majority of neighboring NCCs started to engulf cellular debris, resulting in the formation of massive numbers of phagocytic vesicles. These results demonstrated that not only could NCC phagocytic abilities be induced by acute cell death, but also that the majority of NCCs have the potential to phagocytose debris. Therefore, these data are consistent with the hypothesis that phagocytic NCCs are not a specialized subpopulation of the neural crest population.
Previous studies show that NCCs migrate along conserved, segmentally restricted pathways. However, we found that phagocytic NCCs move toward cellular debris and sometimes even crossed somite boundaries. So what is the mechanism that directs phagocytic NCCs toward dead cells? To answer this question, we designed an ablation assay to quantify NCC recruitment and inhibited a variety of signaling pathways. We found that NCC recruitment was compromised when we treated embryos with a Caspase-1 inhibitor or a interleukin (IL)-1 receptor antagonist, indicating NCC recruitment was mediated by the IL-1β signaling pathway. Interestingly, we observed IL-1β expression in both cellular debris and phagocytic NCCs. Moreover, the IL-1β level we observed after cell ablation was significantly lower than that in previous studies using spinal cord transection and bacterial infection (Bernut et al., 2014; Nguyen-Chi et al., 2014). Given that our cell ablation is more precise compared to manipulations performed in previous studies, the low level of Il-1β release supports the hypothesis that Il-1β secretion is tightly regulated and dependent upon the strength of the inflammatory stimulus (Lopez-Castejon and Brough, 2011).
Conclusion (Yunlu Zhu)
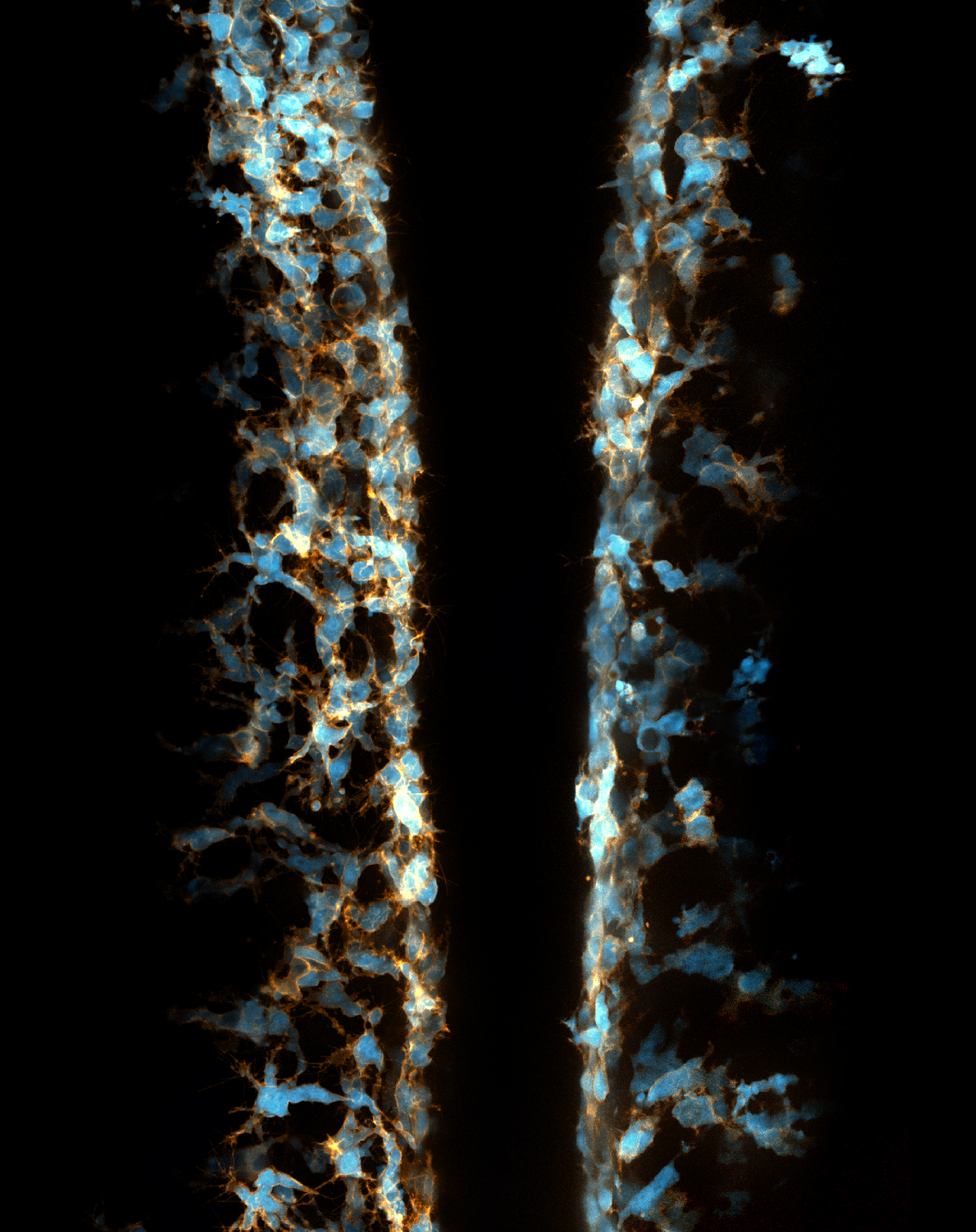
I joined the Kucenas lab at the University of Virginia in early 2014 because I was astonished by the beauty of live imaging in zebrafish embryos (Figure 2).
Figure 2. Two transgenic zebrafish embryos at 22 hour-post-fertilization expressing fluorescent protein in neural crest cells mounted back to back.
My previous focus was the development of perineurial glia and the role of cell-cell interactions in the development of spinal motor nerves. However, while imaging transgenic embryos labeling NCCs, I accidentally observed that some NCCs migrated ectopically away from their innate pathways and had spherical vacuoles with diameters of 3 to 10 µm. I discovered that these weird behaviors of NCCs had never before been described and was deeply attracted by this phenomenon. Therefore, I shifted my focus toward this NCC project.
This novel role of NCCs in debris clearance was quite unexpected but perfectly reasonable. However, the nature of these phagocytic NCCs is not well understood and we have many remaining questions. Given that the majority of NCCs are capable of phagocytosing debris, why don’t they respond to cell death under physiological conditions? What is special about those active NCCs that migrate towards dead cells from a distance? And what happens when professional phagocytes like macrophages colonize the trunk region of the developing embryo? Are their immune-NCC interactions? And what happens if one of these two populations is unable to clear debris? There are so many new questions that have come from this work, and I’m excited to see which ones the next students decide to pursue.
I would like to end this post with a quote from Yogi Berra, which, I think, precisely describes the beauty of live imaging and my experience in this project: “You can observe a lot by just watching”.
References:
Arya, R. and White, K. (2015) ‘Cell death in development: Signaling pathways and core mechanisms’, Seminars in Cell & Developmental Biology. Academic Press, 39, pp. 12–19. doi: 10.1016/J.SEMCDB.2015.02.001.
Bernut, A. et al. (2014) ‘Mycobacterium abscessus cording prevents phagocytosis and promotes abscess formation.’, Proceedings of the National Academy of Sciences of the United States of America. National Academy of Sciences, 111(10), pp. E943-52. doi: 10.1073/pnas.1321390111.
Bertrand, J. Y. et al. (2013) ‘Three pathways to mature macrophages in the early mouse yolk sac Three pathways to mature macrophages in the early mouse yolk sac’, Blood, 106(9), pp. 3004–3011. doi: 10.1182/blood-2005-02-0461.
Galluzzi, L. et al. (2018) ‘Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018’, Cell Death & Differentiation. Nature Publishing Group, 25(3), pp. 486–541. doi: 10.1038/s41418-017-0012-4.
Herbomel, P., Thisse, B. and Thisse, C. (1999) ‘Ontogeny and behaviour of early macrophages in the zebrafish embryo.’, Development (Cambridge, England), 126(17), pp. 3735–45.
Herbomel, P., Thisse, B. and Thisse, C. (2001) ‘Zebrafish early macrophages colonize cephalic mesenchyme and developing brain, retina, and epidermis through a M-CSF receptor-dependent invasive process.’, Developmental biology, 238(2), pp. 274–88. doi: 10.1006/dbio.2001.0393.
Johnson, K. et al. (2016) ‘Gfap-positive radial glial cells are an essential progenitor population for later-born neurons and glia in the zebrafish spinal cord’, Glia, 64(7), pp. 1170–1189. doi: 10.1002/glia.22990.
Lopez-Castejon, G. and Brough, D. (2011) ‘Understanding the mechanism of IL-1β secretion’, Cytokine and Growth Factor Reviews. Elsevier Ltd, 22(4), pp. 189–195. doi: 10.1016/j.cytogfr.2011.10.001.
Mayhew, T. M. et al. (1999) ‘Epithelial integrity, cell death and cell loss in mammalian small intestine.’, Histology and histopathology, 14(1), pp. 257–67. doi: 10.14670/HH-14.257.
Mayor, R. and Theveneau, E. (2013) ‘The neural crest’, Development, 140(11), pp. 2247–2251. doi: 10.1242/dev.091751.
McGrath, K. E. et al. (2003) ‘Circulation is established in a stepwise pattern in the mammalian embryo.’, Blood. American Society of Hematology, 101(5), pp. 1669–76. doi: 10.1182/blood-2002-08-2531.
Nguyen-Chi, M. et al. (2014) ‘Transient infection of the zebrafish notochord with E. coli induces chronic inflammation’, Disease Models & Mechanisms, 7(7), pp. 871–882. doi: 10.1242/dmm.014498.
Rasmussen, J. P. et al. (2015) ‘Vertebrate epidermal cells are broad-specificity phagocytes that clear sensory axon debris.’, The Journal of neuroscience : the official journal of the Society for Neuroscience. Society for Neuroscience, 35(2), pp. 559–70. doi: 10.1523/JNEUROSCI.3613-14.2015.
Smith, C. J. et al. (2014) ‘Contact-Mediated Inhibition Between Oligodendrocyte Progenitor Cells and Motor Exit Point Glia Establishes the Spinal Cord Transition Zone’, PLoS Biology. Edited by B. A. Barres, 12(9), p. e1001961. doi: 10.1371/journal.pbio.1001961.
Stremmel, C. et al. (2018) ‘Yolk sac macrophage progenitors traffic to the embryo during defined stages of development’, Nature Communications. Nature Publishing Group, 9(1), p. 75. doi: 10.1038/s41467-017-02492-2.
Zhu, Y. et al. (2019) ‘Migratory Neural Crest Cells Phagocytose Dead Cells in the Developing Nervous System’, Cell, 179(1), pp. 74-89.e10. doi: 10.1016/j.cell.2019.08.001.
One of the biggest open questions in biology is how organisms can form complex patterns (limbs, organs, entire body plans) from initially disordered or very simple states. Every animal does this at the beginning of its life, forming its full complexity from a single cell. Some are capable of similar feats even after their bodies are fully constructed: starfish and flatworms survive when cut in pieces, the zebrafish restores lost fins, the axolotl regrows limbs and organs alike. How does either case work? How are those patterns established so accurately, and how is it that things so rarely go wrong?
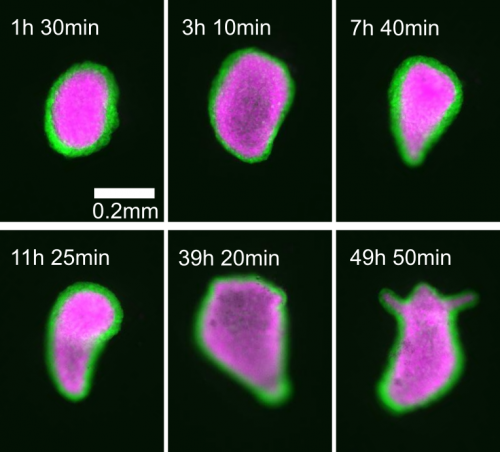
I’m trying to approach this question with the help of one of the most impressive regenerators of all: Hydra. As a 1 cm long freshwater cnidarian it’s a lot less impressive than the monster whose name it bears, but what it lacks in scales and fangs it makes up for in raw regenerative ability. The mythological beast grew two heads for every one it lost. The real thing can regenerate its head, can regenerate from a ball of tissue as small as 150 microns across, and can come back from being dissociated down to single cells if enough of those cells are placed together. On top of this, the small size and simplicity of the creature make it very easy to study and manipulate compared to other model organisms. In fact, several theoretical models purporting to describe the patterning process have already been developed.
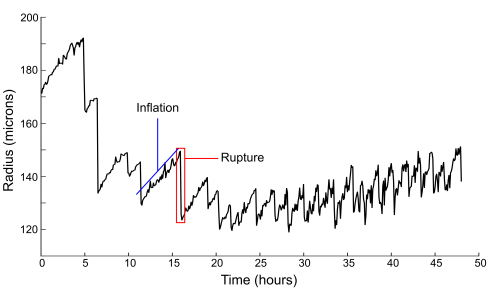
Of particular interest is the fact that these models propose mechanical forces as drivers of pattern formation. Mechanical forces are well known to be important in embryonic development but their role in regeneration is less clear. As a Hydra regenerates it first forms a hollow sphere, which then undergoes osmotically driven cycles of swelling and rupture. According to the literature, there is a characteristic switch in these oscillations from large amplitude and low frequency to small amplitude and high frequency, and this shift is linked to when the animal sets its body axis. Thus the pattern shift was proposed to represent a link between mechanics and biochemistry. I wanted to determine what the mechanism linking mechanical forces to biochemical axis specification was.
On observing many regenerating animals, one of the first things we noticed was that nearly half of the animals we imaged regenerated without a clear shift in oscillation pattern. As this called into question essentially all previous assumptions about the nature and relevance of the pattern shift, we turned to trying to figure out its exact cause.
It was previously proposed that the change in oscillation behavior might be due to the beginnings of a regenerated mouth. This idea makes logical sense, as adult Hydras open their mouths to relieve internal water pressure as well as to feed, but nobody had experimentally tested it. By using injected fluorescent beads to track the location of successive rupture sites, we determined that the spot where rupture occurs is random during large oscillations but conserved during small ones. These data would be consistent with the theory that the large oscillations are due to the tissue tearing under internal pressure, while small oscillations occur when the mouth is established to act as a vent. As confirmation, a tissue piece containing the mouth of the original animal also produces conserved rupture sites and only shows small oscillations.
To this point we had only shown that oscillation behavior and the mouth are somehow correlated. These experiments do not explain how the mouth might affect oscillation behavior. Thankfully, Hydra’s ease of manipulation offers a direct way to establish causality. It’s possible to eliminate all nerve cells from a Hydra, producing a nerve-free animal that is structurally normal but cannot actively move. The critical experiment was to use mouth-containing tissue pieces cut from such nerve-free animals. These contain a mouth that is fully formed and complete, but cannot open on its own. If the deciding factor for small oscillations to occur is simply that the mouth acts as a weak point and tears more easily, nerve-free samples with a mouth should show small oscillations exactly like their normal counterparts. If on the other hand mouth function is the key, they should show large oscillations.
We find that nerve-free mouth pieces show only large oscillations, indicating a causal link between active control of mouth function and a decrease in oscillation amplitude. This provides a concrete explanation for the oscillation pattern shift in cases where it occurs: it is caused by the animal’s ability to open its mouth at will.
Our study does not answer why some pieces are delayed in developing mouth function or provide exact developmental checkpoints. What it does give us is further constraints and parameters that can be used to improve existing models, and put us one step closer to understanding how Hydra regenerates. As Hydra shares many key biochemical pathways with more familiar animals despite its alien appearance, figuring out patterning here could one day be the basis of a similar understanding in humans.
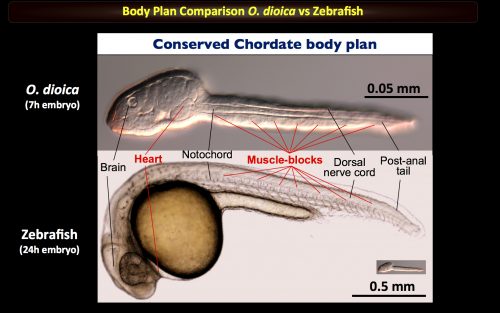
We are seeking a PhD candidate to join our EvoDevo lab in the University of Barcelona to study our favorite chordate model Oikopleura dioica, in which we are currently interested in heart development, 4D imaging of early embryo cleaving, and early developmental responses to environmental challenges. To meet our unique Oikopleura model -> Click here for a tour “A day in our lab” posted in The Node.
We have also engaged a new EcoEvoDevo line investigating if the developmental mechanisms of marine embryos are ready to respond to climate change, including the effects of biotoxins derived from algal blooms. Click here for a tour on this new EcoEvoDevo adventure.
Our approaches include Single-cell transcriptomics,RNAseq, RNAi Knockdowns, CRISPR and Fluorescent-Microscopy
PhD fellowship Call OPEN (FI Catalan program): October 1st-14th 2019 (contact for enquiries as soon as possible canestro@ub.edu)
REQUIREMENT: to have finished a Master degree
CONTACT: please send an email to Cristian Cañestro (canestro@ub.edu), including a brief letter of interest, and the final scores for the degree and Master (indicating the scale), all together in ONE single pdf file.
Dolly the Sheep via Flickr, Toni Barros (CC BY-SA 2.0)
In this episode from our centenary series exploring 100 ideas in genetics, we’re looking at mergers and acquisitions – but in a biological rather than a financial sense. We find out what happens when two cells decide to move in together, unpack the history of genetic engineering and bleat on about the story of Dolly the Sheep.
If you enjoy the show, please do rate and review and spread the word. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com
The Department of Biological Sciences at the University of South Carolina (UofSC) invites applications for two tenure-track Assistant Professor positions in neurobiology. The successful candidates will be expected to establish an independent, extramurally funded research programs in: 1) Molecular or Cellular Neurobiology relevant to neural development and/or disease focused on cell-cell interactions; or 2)Molecular or Cellular Neurobiology using animal models to understand the pathophysiology of Autism or neurodevelopmental disorders (see below). Minimum qualifications include a Ph.D. or M.D. and post-doctoral training in a relevant area. The successful candidates will be responsible for teaching courses relevant to their area of expertise, as well as mentoring research training for graduate and undergraduate students.
1) Molecular or Cellular Neurobiology focused on Cell-Cell Interactions (position # 67536): We are interested in applicants focusing on how interactions between different cells, including cellular interactions with axons, contribute to the biology of nervous system development, disease, or response to injury. This individual will closely interact with research groups in the SmartState Center for Childhood Neurotherapeutics, which includes neurobiologists focused on molecular mechanisms of axon growth in development and after neural injury, and the broader UofSC Neuroscience Community. Applications are made online at http://uscjobs.sc.edu/postings/67536. For questions or further information, please contact Dr. Fabienne Poulain (fpoulain@mailbox.sc.edu).
2) Neurobiology of Autism or Neurodevelopmental Disorders (position # 67556): This position is part of a university-wide initiative to enhance research on Autism and Neurodevelopmental Disorders and establish a Center of Excellence (USCAND) to accelerate interdisciplinary efforts in neuroscience. There is a parallel faculty search in the Department of Psychology for this initiative, and several additional USCAND faculty hires planned over the next few years in complimentary disciplines. This individual will closely interact with research groups in USCAND, as well as the SmartState Center for Childhood Neurotherapeutics, Institute for Mind and Brain,andResearch Consortium on Children and Families. Applications are made online at http://uscjobs.sc.edu/postings/67556. For questions or further information, please contact Dr. Jeff Twiss (twiss@mailbox.sc.edu).
Review of applications will begin by November 1, 2019. The review process will continue until the positions are filled. Qualified individuals should submit a curriculum vita, research statement (3 pages), teaching philosophy (1 page), and the names, email addresses and phone numbers of at least three references to http://USCjobs.sc.edu/postings (with position # and links as above).
The Department of Biological Sciences is a multidisciplinary unit of approximately 1,600 undergraduate students, 50 graduate students, and 35 tenure-line faculty representing a broad range of research areas (www.biol.sc.edu). UofSC has a highly interactive neuroscience research community that encourages and precipitates collaborations. UofSC in Columbia (www.sc.edu) is the state’s flagship university (founded in 1801 and currently one of the top 50 “Best Colleges” according to U.S. News and World Report).
Columbia, SC enjoys more than 300 days of sunshine annually and has ready access to pristine beaches, lakes, rivers and mountains.The city hosts historical and cultural attractions, festivals, performing arts and sporting events, parks and outdoor recreation including Congaree National Park and 50,000-acre Lake Murray.
The University of South Carolina is an affirmative action, equal opportunity employer. Minorities and women are encouraged to apply. The University of South Carolina does not discriminate in educational or employment opportunities on the basis of race, gender, age, color, religion, national origin, disability, sexual orientation, genetics, veteran status, pregnancy, childbirth or related medical conditions.
We are seeking to recruit an outstanding group leader who aims to explore bold new areas of biological inquiry and carry out interdisciplinary research to investigate multicellular development at all scales. How organisms respond and adapt to their environment, during development and throughout their lifetime, is of central interest.
The Developmental Biology Unit seeks to understand the general principles and mechanisms underlying the development of multicellular organisms. Researchers in the unit combine the power of genetic model organisms with quantitative imaging and -omics technologies, synthetic biology, reduced (in vitro) systems and theoretical modelling, to create a cross-cutting approach to modern developmental biology.
Research in the Developmental Biology Unit is firmly embedded within the overall EMBL environment, with extensive in-house collaborations, access to outstanding graduate students and postdoctoral fellows, and support from cutting-edge facilities, including genomics, transgenesis, metabolomics, mass-spectrometry, and microscopy.
Your role
You will lead a research group to pursue highly ambitious and original research at the frontier of developmental biology. In general, EMBL appoints group leaders early in their career and provides them with a very supportive, collaborative environment and generous work package for their first independent position. Significant core funding and limited teaching responsibilities allow you undertake a farsighted research program.
You have
The successful candidate will present a highly original and ambitious research plan that concisely describes the background and status of the questions that will be addressed, the experimental strategies and methods that will be employed, and the ultimate goals. A PhD degree in the Natural Sciences is expected. Candidates with a background in physics and modelling of developmental processes are also encouraged to apply.
Why join us
EMBL is an inclusive, equal opportunity employer offering attractive conditions and benefits appropriate to an international research organization with a very collegial and family friendly working environment. EMBL is committed to achieving gender balance and strongly encourages applications from women. Appointment will be based on merit alone. The remuneration package comprises a competitive salary, a comprehensive pension scheme, medical, educational and other social benefits, as well as financial support for relocation and installation, including your family, and the availability of an excellent child care facility on campus.
What else you need to know
We are Europe’s flagship research laboratory for the life sciences – an intergovernmental organisation performing scientific research in disciplines including molecular biology, physics, chemistry and computer science. We are an international, innovative and interdisciplinary laboratory with more than 1700 employees from many nations, operating across six sites, in Heidelberg (HQ), Barcelona, Hinxton near Cambridge, Hamburg, Grenoble and Rome.
Our mission is to offer vital services in training scientists, students and visitors at all levels; to develop new instruments and methods in the life sciences and actively engage in technology transfer activities, and to integrate European life science research. The working language of the institute is English.
In your online application, you will be asked to include a cover letter, your CV, the names and contact details of 3 referees and a concise description of research interests & future research plans, typically not exceeding five pages.
Further information about the position can be obtained from the Head of Unit, Anne Ephrussi (anne.ephrussi@embl.de).
Information on Group Leader appointments can be found here http://www.embl.org/gl_faq.
Interviews are planned for 18, 19 and 20 December 2019.
An initial contract of 5 years will be offered to the successful candidate. This is foreseen to be extended to a maximum of 9 years, subject to an external review.
Rapid turn over of sex determination mechanisms provides biologists with an elegant study system connecting sexual selection to molecular evolution. Striking examples of this turnover are found in African cichlids, where multiple sex determination signals exist not only within the same genera, but sometimes within the same species [1]; in the common house fly, where the primary sex determiner varies along a longitudinal gradient [2], and in the Japanese frog Rana rugosa, a species in which sex chromosomes segregate geographically but whose signals can be overridden by environmental (hormonal) intervention [3].
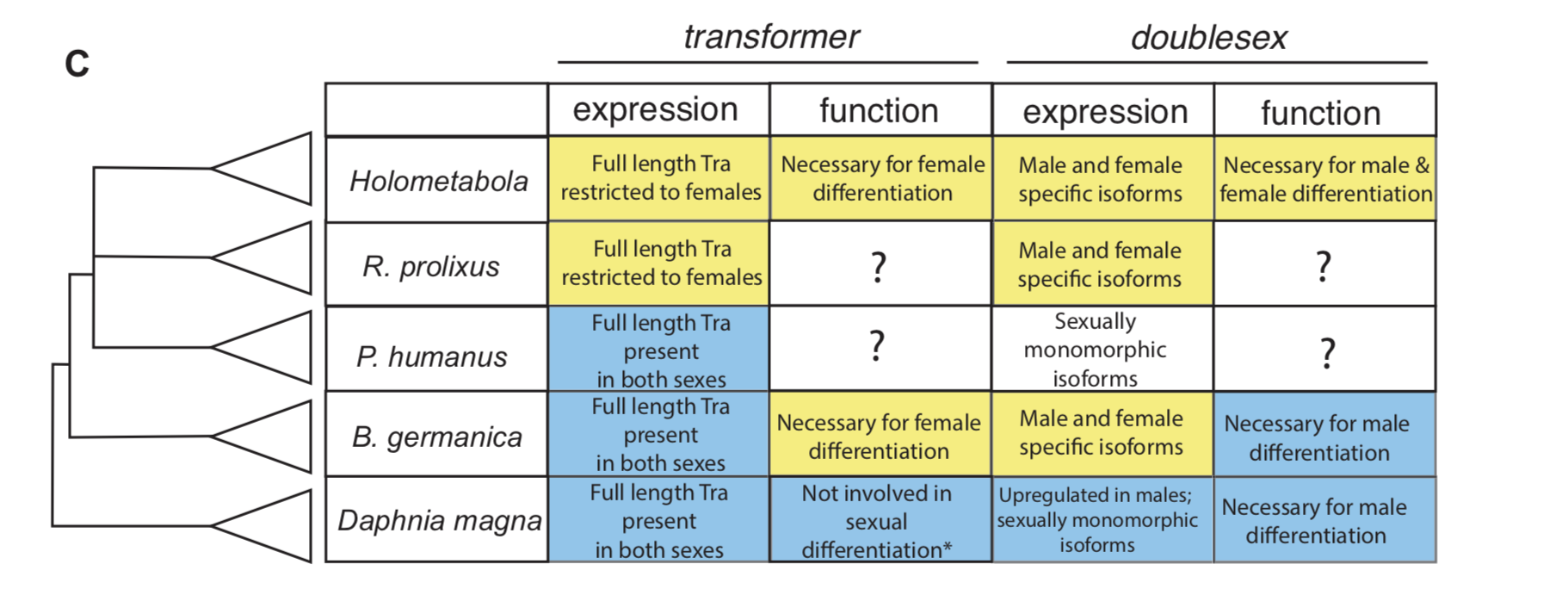
Downstream of the primary sex determination signal is a developmental pathway that also presents an intriguing puzzle for evolutionary biologists. While the sexual differentiation pathways of mice and humans are fundamentally similar, and while the downstream elements of this pathway are conserved between bees and flies, there are no shared genes in the sex determination pathways of mammals, arthropods, and nematodes with the striking exception of the Doublesex Mab-3 Related Transcription factor (DMRT) family. Furthermore, the fundamental biochemistry of sexual differentiation in these taxa is distinct. In mammals, signaling molecules establish feedback loops that promote one sexual identity and repress the opposite one [4]. In insects, sex-specific RNA splicing determines sexual identity [5]. And in nematodes, a cast of genes including a phosphatase and protease control the sexual fate of a cell [6]. Thus the puzzle: how does a pathway which appears stable on the order of hundreds of millions of years diverge so distinctly between different animal taxa? The Kopp lab was interested in how it is that the sexual differentiation pathway changes on a macro-evolutionary time scale.
In insect developmental biology Drosophila is the point of reference for all other species. Sex differentiation is no different, where two genes at the bottom of this pathway — transformer (tra) and doublesex (dsx) — were first described in the fruit fly. doublesex (a member of the eponymous Doublesex Mab-3 Related Transcription factor family) is a transcription factor with male- and female-specific isoforms. Both isoforms contain the same DNA binding domain, while alternative, sex-specific splicing at the 3’ end of dsx transcripts changes the effect the male and female dsx isoforms have on their targets [7]. In Drosophila, the male isoform of dsx is produced by default. Active intervention by Tra changes the pattern of dsx splicing from a male to a female isoform. tra itself is also sex specifically spliced, such that a premature stop codon appears in the male, but not the female, transcript [8]. The outgroup to insects with the best studied developmental model is the class Branchiopoda, containing the model crustacean Daphnia magna. In Daphnia, dsx is upregulated in males and lacks male and female isoforms. It is required for male, but not female differentiation [9]. tra is not sex-specifically spliced in this species [10].
To investigate how this insect-specific pathway of sexual differentiation based on sex-specific RNA splicing evolved from an ancestral state without male and female isoforms, we had to look beyond the Kopp lab’s traditional Drosophila domain. Way beyond it. Drosophila-like splicing and function have been documented in Coleopterans and in Hymenopterans, suggesting that the tra–dsx axis of sexual differentiation was present in the common ancestor of Holometabola [11,12,13,14]. The phylogenetic interval between crustaceans and Holometabola is filled in large part by the paraphyletic group of hemimetabolous insects — or those that go through a partial metamorphosis. And because insect pests are more likely to have sequenced genomes, we settled on the following delightful trio of hemimetabolous insects: the kissing bug Rhodnius prolixus (order: Hemiptera), the louse Pediculus humanus (order: Pthiraptera), and the German cockroach Blattella germanica (order: Blattodea). Because R. prolixus and P. humanus are both obligate blood feeders, they cannot be cultured in a lab without strict regulations. Thus, we were reliant on the kindness of Ian Orchard’s lab in Toronto for R. prolixus tissue shipped in RNALater. For P. humanus, our tissue collection was somewhat more exciting — we struck up a relationship with a local louse collector, who is hired to comb nits and lice from the heads of infested humans. In contrast, my experience getting to know German roaches was much more civilized. I was lucky to receive an NSF EDEN grant to study in Barcelona for two months with Xavier Belles, where I learned how to knock down genes in B. germanica. The Belles lab has done extensive work developing B. germanica into an evo-devo model.
Double stranded RNAi targeting transformer masculinizes females. From Wexler et al, eLife 2019;8:e47490.
For all three species, we investigated the isoforms of tra and dsx present in males and females. In the German cockroach, we knocked down both genes to probe their functions. The phylogenetic breadth of our samples allowed us to observe the evolutionary trajectory of dsx and tra across a large evolutionary interval. We discovered that the holometabolous pattern of tra splicing, with a male-specific premature stop codon, evolved after tra evolved a functional role as a regulator of female development. Our results from R. prolixus, where we isolated tra with a male specific stop codon, showed that the “canonical” pattern of sex-specific splicing of this gene did evolve before the common ancestor of Holometabola. In contrast, it appears that the familiar holometabolous pattern of male- and female-specific dsx isoforms differing at their 3’ end evolved before dsx became a key regulator of female differentiation. So for each gene, we observed a different story: with dsx, the “holometabolous” splicing pattern evolved before the “holometabolous” functional role; for tra, this pattern was reversed.
Summary of tra and dsx expression and function in the three hemimetabolous insect orders in comparison to the canonical holometabolous mechanism and to Daphnia. From Wexler et al, eLife 2019;8:e47490.
The knockdown experiments lead to some very fun collaborations as we attempted to probe the resulting phenotypes. After I returned from Spain and set up my own B. germanica colony at UC Davis, I was reliant on the generosity of Coby Schal, who shared a few roaches from his colony, allowing me to set up my own. The Schal lab specializes, among other things, in the study of insect behavior. When I attended the conference of the International Congress of Entomology with Coby a year later, I excitedly sat down to show him images of the masculinized females we observed upon knocking down tra. Coby then offered to quantify the degree to which these roaches’ behavior was masculinized. The experiments he did with his lab technician Ayako Wada-Katsumata revealed that like in Drosophila, sex-specific behavior of the German roach is under the control of tra. Separately, I began chatting with my teammate from the UC Davis cycling team about the work. This teammate and friend, Matt Amicucci, also happened to be a graduate student in the chemistry department, who then offered to help investigate the oligosaccharides produced by masculinized females in their tergal glands. Thanks to Matt’s expertise, we learned that tra also controls the chemical physiology of German roaches.
The breadth of the questions motivating this project and the number of questions that came out of it; the diversity of skills among the paper’s authors, and the generous support of everyone involved really made this work a wonderful and memorable PhD.
Above: Wild type males respond to the stimulus of a wild type female antenna by raising their wings and presenting their tergal gland.
Above: Females treated with dstra respond as wild type males do to the stimulus of a wild type female antenna. These treated females raise their wings in a stereotypical male courtship display.
The research group headed by Werend Boesmans and Veerle Melotte is looking for a highly motivated and talented PhD student to investigate the role of the intrinsic innervation of the gut in cancer. The enteric nervous system, also known as the second brain, is a mostly ignored member of the tumor microenvironment but is linked to the development and progression of colorectal cancer. In the current project, the involvement of specific neural cell types in colorectal tumorigenesis will be investigated in both in vivo and in vitro models, and combined with advanced optical microscopy, genetic lineage tracing, marker gene expression analysis and single cell transcriptomics.
This PhD project takes place within the School for Life Sciences (SLS) of the Transnational University of Limburg (tUL), a unique collaboration between Hasselt University (UHasselt, Belgium) and Maastricht University (UM, The Netherlands). The student will perform research at the Biomedical Research Institute (BIOMED, https://www.uhasselt.be/biomed) and the School for Oncology and Developmental Biology (GROW, https://www.maastrichtuniversity.nl/research/school-oncology-and-developmental-biology) embedded in the Department of Pathology of the Maastricht University Medical Center (MUMC).
You can only apply online.
The selection procedure consists of a preselection based on the application file and an interview.
Please provide the contact details of two referents in your application.
 (No Ratings Yet)
(No Ratings Yet)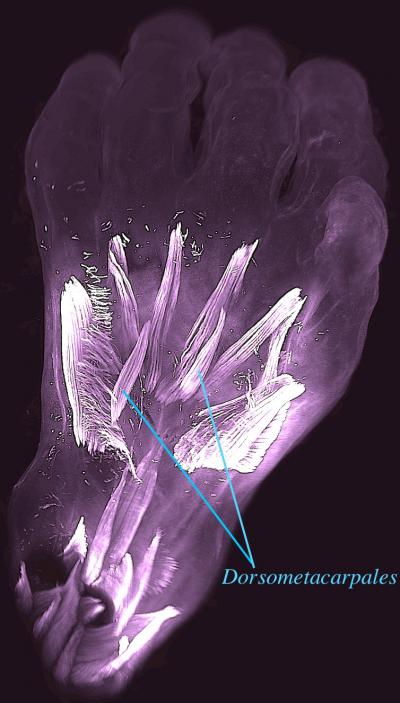
 (1 votes)
(1 votes)