I was excited to be in Liverpool for the Biologists @ 100 meeting this past March. Not only is a meeting organised by The Company of Biologists always guaranteed to be friendly and exciting, this was also my first science-related activity back in the UK since I started a postdoc in Boston last November, so I was doubly happy to be there.
Sunny views on the River Mersey, Liverpool’s aquatic pride and joy.
The most intriguing aspect of the meeting for me was the fact that it drew together an unusually broad range of scientists – this wasn’t your average BSDB/BSCB joint meeting! It was a celebration of the 100 years since the founding of The Company of Biologists and the 5 journals that are published under its umbrella. Accordingly, in addition to the cell and developmental biology track, there were three more included in the meeting, each in reality a mini symposium or satellite meeting: sensory perception (the Journal of Experimental Biology’s symposium), interdisciplinary approaches to combatting antimicrobial resistance (Disease Models & Mechanisms’ symposium), and experimental biology and impact: solutions to climate change and biodiversity loss, organised by the Society of Experimental Biology.
The ACC, venue of the conference.Talks in the ACC.
Bringing together many different groups of scientists that might not typically encounter each other at meetings and pooling resources has some practical perks: firstly, the meeting was held in the ACC, a big, comfortable, and supremely well-located venue right on the River Mersey. We were also very close to the Museum of Liverpool, where a welcome reception was held at the end of the first day, and St George’s Hall, where the conference’s gala dinner was held on the penultimate day of the conference.
St George’s Hall, where the gala dinner was held.
However, what I appreciated most was getting to hear about many different types of science that I would not have normally expected to hear at a conference (and especially at the same conference!). The biodiversity and climate change plenary lectures by Jane Francis of the British Antarctic Survey, and Hans-Otto Pörtner from the Alfred Wegener Institute for Polar and Marine Research in Germany were on the one hand fascinating, and on the other hand greatly sobering. They highlighted the essential role every scientist (not just climate scientists) should play in disseminating accurate information about the climate catastrophe our planet is suffering, and what we should be doing to combat it.
The parallel session format of the conference meant that you could hop between tracks several times throughout the day and take in all the different types of science that converged at the meeting. For example, the sessions on sensory perception were very comprehensive, with talks on vision, light pollution, thermosensation, sound perception, magnetic and chemical senses. There were also talks and posters on insect behaviour and insect-plant parallel evolution – this is a very subjective opinion, but butterflies always make a conference more fun. I was also excited to attend one of the sessions on drug discovery and ‘omics. Talks ranged from large-scale efforts to combat the development of antimicrobial resistance, to systematically exploring the breadth of organic chemistry to discover new drugs. This might not be news to some readers, but I was not aware of the tendency of microbes that develop drug resistance to concomitantly become more sensitive to other classes of drugs. This is something that researchers are actively investigating in their efforts to combat antibiotic resistance.
There was also a handful of plant-themed posters and a couple of talks on plant biology. There I heard the best description of leaves from Dr Chris Whitewoods from the Sainsbury Lab in Cambridge: “kind of green, kind of flat, sort of pretty in the sunshine.” For someone who gave a talk on leaf air space development, it was hilarious to get such a fun, entry-level description of leaves. Speaking to the (relatively few) plant scientists that were present at the meeting, I wondered why there tend to be such few plant researchers at developmental and cell biology conferences. They shared with me that they tend to congregate at plant science conferences instead, so this is my personal plea to plant scientists and developmental biology conferences: please sign up for dev bio conferences / invite plant scientists to speak at them! It is so refreshing to hear about research in non-animal models (and this is coming from someone with a passion for animal development).
Overall, the Biologists @ 100 conference was a big success. It was fantastic to see old colleagues and former lecturers, meet new people and discuss potential collaborations, and above all be exposed to an unusually broad suite of scientific investigation. There is nothing quite as inspiring as sharing one’s excitement about research with others!
All the world’s a metabolic dance, early career scientists are leading the way!
Emerging perspectives in metabolism
Dr. Lautaro Gandara, Postdoctoral researcher, EMBL, Heidelberg.
This week we will get to know insights from Dr. Lautaro Gandara, who is a postdoctoral researcher in the Crocker lab at EMBL Heidelberg. Lautaro’s work delves into the profound interplay between metabolism, toxicology, and development. As he prepares to establish his own lab, Lautaro is driven by a deep curiosity about how life adapts to environmental challenges, and how metabolic shifts shape the very essence of biological resilience. From his studies of Drosophila melanogaster to the impact of environmental stressors on insect populations, his research questions the fundamental nature of life’s response to stress and transformation. For Lautaro, science is not just a pursuit of answers but a journey of discovery, where each question unfolds new dimensions of understanding. He believes that the study of metabolism and development is not merely academic—it’s a window into the intricate ways life connects, adapts, and evolves. Follow his journey as he continues to explore these deep questions and check out his work here.
Could you share your journey into studying metabolism and what inspired you to specialize in metabolic studies using Drosophila melanogaster.
When I took an introductory course in chemical biology as an undergrad, I remember being less than enthusiastic about the field. Metabolism was presented as a completed research program—a field in which human metabolic maps had already been established, flux control theory had provided all the relevant dynamical information, and the only open questions were clinical ones. It was only after I started my PhD work in Pablo Wappner’s lab that I got access to the then-new research showing how metabolism, far from being a housekeeping process, can actually transmit information by regulating gene expression, signaling pathways, and so on. At the time, I was studying the response to oxygen deprivation (hypoxia) in flies, and we soon realized that many metabolic facets of this process remained unexplored. Drosophila larvae can perform lactic acid fermentation in hypoxic environments, but at that time there was little information on the spatial and temporal properties of this metabolic switch. These questions became the focus of my work as a grad student.
How did you get interested in the field of toxicology and impact of chemicals on insect health and metabolism? How do you think toxicology and metabolism fields overlap and how they regulate each other or how they are connected?
I have always been fascinated by the way in which life actively responds to environmental change—its intrinsic ability to preserve itself by regulating its own activities and structures. When you look at these processes more closely, all the reductionist metaphors of “life as mere machines” start to crumble, right? This is where my interest in the hypoxia response originally came from. So as a postdoc in Justin Crocker’s lab at EMBL, I wanted to expand on the approaches I had used as an undergraduate to further explore these phenomena. Instead of focusing on just one environmental perturbation (hypoxia) and one phenotype (metabolism), we decided to test more than 1000 different chemical stressors and measure how the effects induced by these molecules propagate across the different scales of biological organization.
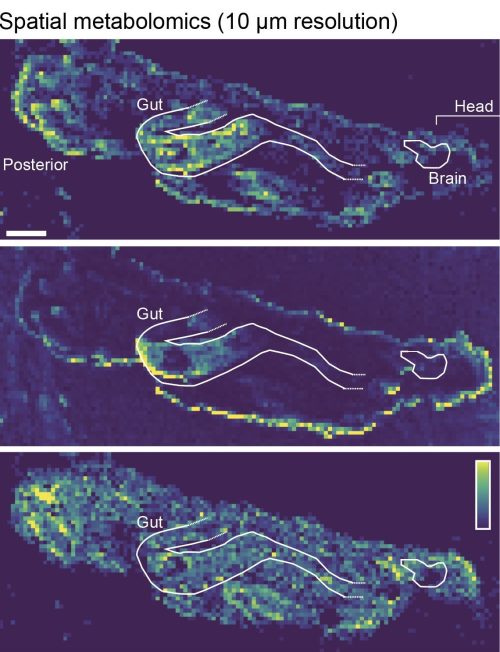
High spatial resolution MALDI-imaging analysis of a Drosophila larva (scale bar = 100 μm). The images show relative intensities of individual lipid species, each for an individual m/z value: upper panel = 544.3373 (C26H52NO7P); middle panel = 177.0158 (C7H6O4); lower panel =744.5537 (C41H78NO8P). Image source: LG.
Tell us about how you see the future of metabolism evolve with the new upcoming tools like the FRET biosensors you worked on. What changes have you seen in the community regarding exploring metabolic aspects of development ?
I consider the development of tools to be an essential driver of science, and I think that the interplay between development and metabolism is a paradigmatic case of this phenomenon. In the first half of the 20th century, embryologists such as Joseph Needham studied the metabolic facets of development using the technology available at the time (calorimetry, respirometry, etc.). However, as embryology was transformed into developmental biology, and the focus shifted from organismal-level processes to gene- and cell-level phenomena, this set of tools proved ill-suited for exploring metabolism at this scale. As a result, questions about the role of metabolism during development were put on hold. Recently, the combination of metabolic FRET sensors, spatial omics techniques (especially spatial metabolomics), and flux analysis by isotope tracing has reignited interest in these old questions and revitalized the field. Ongoing research by many different groups working on different model systems around the world is providing priceless information about the precise role of metabolism in the development of multicellular organisms.
One of your reviews is titled “Metabo-Devo: A metabolic perspective of development”. Could you elaborate on the key findings and their implications for the field? How do you integrate different disciplines – metabolism, development, and evolution in your research, and what unique insights have emerged from this approach?
In that review article, we proposed a conceptual framework to discuss how metabolism interacts with developmental processes. We classified these interactions as either 1) bioenergetic functions, 2) regulation of gene expression through changes in the epigenome, or 3) signaling functions.
Bioenergetic processes are those that provide energy or building blocks to developing tissues. Many cell populations that proliferate at high rates acquire a particular metabolic state, called aerobic glycolysis, that allows them to synthesize macromolecules at the right pace. This metabolic switch was reported ~100 years ago in the context of cancer biology (i.e. the Warburg effect), but it is now becoming clear that aerobic glycolysis is also required for cell proliferation in developing organisms. Ongoing research efforts aim to elucidate the developmental processes during which this metabolic transition occurs, and how it is regulated.
In addition to this bioenergetic role, specific metabolic pathways can directly regulate gene expression. Certain metabolites have been shown to act in developmental contexts as rate-limiting substrates for histone and DNA modification. And this metabolic control of gene expression has been shown to play an essential role in key developmental processes, such as the zygotic genome activation. Similarly, many metabolites are directly involved in the post-translational modification of signaling-related proteins, while several metabolic enzymes have been reported to act as multifunctional “moonlighting” proteins that can perform alternative functions not necessarily related to metabolism. Thus, metabolites and metabolic enzymes have the potential to modulate signaling pathways that are essential for development.
Although we think that the classification described above provides a useful conceptual framework for designing and discussing experiments, dissecting the actual role of metabolism in specific processes remains challenging, especially because the same metabolites and enzymes may simultaneously play bioenergetic and signaling roles that affect the same developmental phenomenon. In any case, the emerging view is that metabolism and development are deeply intertwined processes that cannot be disentangled. Thus, this observation highlights the need for a discipline or research area—developmental metabolism or “metabo-devo”—that directly addresses these issues.
You are currently studying the impact of toxic substances from genotypic to phenotypic perspectives and your work suggests the use of agrochemicals is the root cause of insect decline. Can you briefly discuss this work ?
We decided to focus on agrochemicals (i.e. insecticides, fungicides, plant growth regulators, etc.). Increased use of pesticides has been proposed as a potential cause of the widespread declines in insect populations, but studies investigating the effects of these molecules on insects are often limited to a few chemicals and a single insect species. We started with a screen that tested the effects of 1024 agrochemicals on the behavior of Drosophila larvae. Behavioral changes often have a mechanistic basis at simpler phenotypic levels, and thus monitoring behavior can provide important information about the state of the biological system as a whole. Surprisingly, we found that ~60% of the molecules in our library significantly alter larval behavior! And these effects are not limited to flies—we also detected similar sublethal behavioral changes in mosquitoes and butterflies, suggesting a generalizable effect on insect populations.
By exploring some of the screen hits further, we found that the effects go well beyond behavior. Exposure to sublethal doses triggered widespread phosphoproteomic changes, revealing how these chemicals affect many different physiological processes. And chronic exposure led to delayed development and reduced reproductive output, potentially contributing to the decline in insect populations. Thus, the study showed that even non-insecticidal pesticides at field-realistic sublethal concentrations can have profound ecological consequences, highlighting the need for better safety assessments that take sublethal effects into account.
How difficult some of those experiments work – did you have to deal with midnight timepoints or require an army of undergrads/ long hours ?
The behavioral screen was very time consuming. We ended up testing 3072 different conditions (different molecules and concentrations), so including replicates and controls, we ran more than 10000 individual assays… It was a lot of work, but I got a lot of help from everyone in the lab, and fortunately we managed to get it done in just a few months. It was truly a collaborative effort!
Can you shed light on the big picture of the field, what are you most excited about and how does it all connect to impacting insect/human health.
Understanding how animal systems respond to stress is becoming increasingly important in the current context of human-induced global environmental change. Going back to my fascination with the resilience of biological systems, I think our previous work has opened up an exciting opportunity to test some of the open questions in the field of stress response. There is this idea that multicellular organisms need to activate a system-level stress response to restore homeostasis. This process would be based on the well-known “integrated stress response” pathway—a process that occurs at the cellular level—but would also involve organismal level stress defense systems involving cell differentiation processes, metabolic switches, physiological changes, and even behavioral effects. Testing this hypothesis, however, is not straightforward because it would require measuring the degree of interconnectedness among all these different processes across a wide range of environmental perturbations. By performing the behavioral screen I mentioned earlier, we have defined a panel of chemicals that induce widespread systemic changes in fly larvae, but at sublethal concentrations, meaning that these animals can orchestrate a successful response to these stressors and recover from these injuries. Thus, this panel of chemicals can be used next to explore the level of integration between the different stress defense systems operating at the organismal level. I hope to start my own group soon, and this is one of the first problems I’d like to tackle.
What advice would you offer to students and early-career scientists interested in exploring the intersections of metabolism, development, and evolution?
I’d tell them to have fun! These are indeed exciting times to be doing metabolic research in developmental systems. New technologies are allowing us to explore the various ways in which metabolism transfers information not only across space (inter-organ communication, metabolic coupling between cell types, etc.), but also across time (developmental and cell differentiation processes). I think this is the time to be bold and creative in finding ways to make the most of this technological advantage.
What role does curiosity play in your life, both within and outside of science? How important is it for you to answer basic science questions about behavioral and metabolic aspects of toxicology and how are you planning to use insect models to bridge basic science and applied research ?
Curiosity does play an essential role in my life and in my research. The project I mentioned earlier, in which we studied the sublethal effects of agrochemicals on insects, has some facets that are obviously relevant to the community as a whole. But I strongly disagree with the idea that it’s only worth studying certain natural phenomena if they affect us directly, or if we can use them for our direct benefit in the short term. Basic and applied research aren’t in opposition to each other—on the contrary, they are involved in a dialectical feedback in which the former feeds the latter with information about how fundamental processes work, while the latter not only highlights which questions are most pressing, but also drives the development of new tools and methods that can then be applied to basic research. Hypoxia research is a good example. The oxygen sensor—the molecule that allows cells to determine oxygen availability and trigger an appropriate response when oxygen levels become too low—was identified more than 20 years ago in fundamental work on C. elegans. Years later, this molecular machinery was found to have enormous clinical relevance, as it could be manipulated to induce angiogenesis and treat the symptoms of cardiovascular disease or prevent it and limit tumor growth. But this useful knowledge first came from curiosity-driven research on basic genetics.
Were there any pivotal moments that shaped your career path? What’s an unexpected place you’ve found inspiration for your work?
It may sound simple, but for me the most important source of scientific inspiration is talking to other people. I can think about some problems for hours, but in my case, ideas really take shape when I express them, either by writing them down or by discussing them with someone else. It’s as if, by trying to communicate my thoughts, I organize them into a coherent narrative—a logical structure—from which new ideas can occasionally emerge. And then different people, with different backgrounds and different opinions, will steer your train of thought in completely different directions, certainly leading to unexpected places… But I’m not talking about big meetings full of people here—it’s the one-on-one discussions that force you to interact longer with a single concept and exhaust all its multiple possibilities that are often more productive for me.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
I don’t really see myself doing anything other than biological research. But if it weren’t for biology, I think I would have pursued a career in the social sciences. Human societies are incredibly complex entities, but understanding how they work is not just a matter of academic curiosity. Especially now, looking at the current times, explaining how society, economics, or history actually works has become a pressing issue. I think that on an individual level—and as citizens —we can’t afford to ignore these problems any longer.
Anything you’d want to highlight for the future.
I’m currently looking for a place to set up my own lab. Aside from the global uncertainty we discussed earlier, I think these are exciting times to start a group. New technologies are making old questions experimentally tractable, new species are being proposed all the time as novel model systems, and AI promises to change the way we approach data analysis. There’s no doubt that the way we do science is going to change dramatically in the coming years, and that’s an idea I find particularly appealing.
Last week we learnt about how males and females are metabolically differently rewired – from the perspective of lipid storage and utilization – The Fat of the Matter (Lianna Wat)
In our recently published paper ‘Ectoderm barcoding reveals neural and cochlear compartmentalization‘, we utilized ultrasound-guided in utero nano injections to deliver heritable DNA barcodes to cells exposed to the amniotic fluid, performing the first high-throughput single cell lineage tracing study of the developing nervous system and inner ear. Our results led to the reclassification of cell lineages in the cochlea and provided a comprehensive single-cell atlas of neural and cochlear clonal relationships.
Sandra’s perspective: Work related to this publication already started a long time before I (Sandra) joined Emma R. Andersson’s lab back in 2019 to pursue my PhD studies. Work done by Katrin Mangold and Jingyan He, PhD students in the lab at the time, laid the foundation for successful in utero injections into the amniotic cavity to target the neural plate (Mangold et al, 2021). Like the neural plate, the progenitors of the inner ear are exposed to the amniotic fluid during an early time window of development (~E7 – E9) and we therefore hypothesized that we would be able to target the otic placode and manipulate progenitors of the inner ear using this technique. In fact, preliminary data from a Master’s student, Sanne Stokman, showed some targeting of the vestibular system of the inner ear. This data hinted that we should be able to target the cochlea as well – and laid the foundation to further explore this during my PhD studies. Drawing on expertise from two laboratories—the Andersson lab at the Karolinska Institute, specializing in developmental biology, Notch signaling, and in utero injection techniques, and Matt Kelley’s lab at the National Institutes of Health, experts in inner ear developmental biology—the first experiments targeting the cochlea were conducted in 2019. I vividly recall witnessing the targeting of the cochlea, for the first time, using low-titer H2B-GFP lentivirus injections performed by Jingyan. The mosaic-like targeting of hair cells along the cochlear spiral was truly remarkable (Fig. 1). I remember sharing these initial positive results with Emma via text while at the confocal microscope. Together with Jingyan, who truly mastered the injection technique, and together with the Infinigene core facility (established by Emma), we optimized our injection strategy to target the inner ear, including the injection volume, embryonic stage and viral titer. This journey was, of course, not without its challenges. At times, we faced difficulties with mouse breeding, low viral titers, and, not to forget, a pandemic that occurred along the way. Finally, in 2022, injections using high-titer virus yielded high-targeting efficiencies – of over 90% of the hair cells and supporting cells in the Organ of Corti. I remember observing the high efficiency targeting, quantifying targeting efficiencies late at night the same day, and presenting the results the next day over Zoom during the Kelley lab meeting – excited to share the new results.
Fig1: First cochlear targeting results using low-titer H2B-GFP lentivirus. Injections performed at embryonic day 8, collection at postnatal day 0. Hair cells shown in magenta, GFP-labelled cells shown in green. DAPI (nuclei) in blue
Using viral delivery of heritable DNA barcodes, we were able to perform high-throughput lineage trace studies in the inner ear for the first time, answering fundamental questions about inner ear development. I am still thrilled that we could apply this advanced method to explore lineage relationships within the inner ear. Although we initially focused on the divergence of medial and lateral cochlear cells, we soon realized that the most intriguing findings came from cells outside the Organ of Corti, including Hensen’s cells and populations within the stria vascularis. One of my favorite insights is the classification of Hensen’s cells. At the time, more research was performed on this specific population of cells in the cochlea, but no consensus existed on whether this cell type should be considered a supporting cell subtype or grouped with cells lateral to the Organ of Corti. Our data indicated that Hensen’s cells should be classified as lateral to the Organ of Corti, rather than being a supporting cell subtype of the Organ of Corti – if basing this definition on lineages. This classification might be relevant for future strategies focusing on regeneration and differentiation of cells within the cochlea. These findings also nicely aligned with results from another project of my PhD studies, in which we showed that Hensen’s cells respond differently to the loss of Jag1-mediated Notch activation compared to lateral supporting cells (De Haan et al 2024, Development).
As dissociation of cochlear cells relies on physical dislocation through microdissections, we inadvertently included cell types in our analysis that were not initially the focus of the study, including spiral ganglion neurons and glia. The analysis of barcode sharing between these populations proved to be quite complex. Contamination between spiral ganglion neurons and glia cells often occurs in single-cell preparations, so we investigated the barcode sharing between these populations to determine whether it was due to contamination or if subtypes of neurons might share a common origin with glia cells, which would challenge the current view. Ultimately, this experience taught me the importance of remaining open to new discoveries while ensuring that data collection and experimental design are well-suited to address the research questions. It highlighted the need for careful experimental planning, robust data collection and validation to draw accurate conclusions from complex datasets.
Jingyan’s perspective: While contributing to the inner ear lineage tracing project, I (Jingyan) was also focused on the overarching goal of my PhD projects: advancing the in utero injection technique to label non-ectodermal cells, building upon previous success in ectoderm targeting. By exploring different injection approaches, I successfully established a technique to label diverse cell types with other embryonic origins (ongoing work). As part of this highly ambitious project, we lineage traced tall ectoderm-derived cells using amniotic cavity injections at E7.5. We collected whole embryos at E9.5 and E10.5 after barcode labeling at E7.5 with amniotic cavity injection. This approach allowed us to study the lineage relations of central nervous system, neural crest-derived cells, as well as various epithelial lineages, including the otic lineage. This part of data was later incorporated into the inner ear lineage tracing paper to add a more comprehensive understanding of neurodevelopment and illustrate the potential clonal relations between otic epithelial lineages and other cell types.
One of the key challenges in this work was balancing the viral transduction efficiencies across different collection time points to ensure an optimal number of labeled cells for the single cell RNA sequencing and clonal analyses. Although E9.5 and E10.5 are just one day apart, the difference in total cell number is substantial. We had to pool a few E9.5 embryos to obtain sufficient cell numbers for clonal analyses. However, for E10.5, if we used the same amount of viral particle and reached the same transduction efficiency as E9.5 collection, a single E10.5 embryo yielded so many labeled cells that we needed to split them into multiple reactions when preparing the sequencing libraries.
Viral transduction efficiency itself was influenced by a variety of factors, such as the subtle differences of the embryos’ stages when injecting, viral storage time, freeze-thaw cycles and the variability between different virus production batches, making it difficult to consistently control the number of transduced cells recovered from each injection.
Collection days were always highly intense and stressful, involving a full workflow from the setting up of the cell sorter, embryo collection, dissection, dissociation, cell sorting, to library preparation, all within a tight window to preserve cell viability and RNA quality. Sandra and I always teamed up to streamline the workflow, assisting each other with reagent preparation, cell counting, and other time-sensitive steps.
Joint perspective: The manuscript, initially focused on cochlea only, was submitted shortly before Sandra’s successful PhD defense in August 2024. The E9.5/E10.5 ectodermal lineage tracing data was initially intended for a separate publication, but in response to reviewer and editorial feedback, we decided to incorporate it into the current paper during the revision process. This addition significantly enriched the manuscript and provided a more comprehensive understanding of neurodevelopment. As a result, Jingyan and Sandra shared the first authorship of the paper, highlighting the collaborative nature of our research and the importance of these findings.
Now that the paper has been published and new projects are underway in the Andersson lab applying this technique to different tissue systems, we are both excited to see how the technology will be used, further developed, and what biological insights it will uncover. We’re proud that our work also laid the foundation for an ERC Consolidator Grant to the Andersson lab, who will continue to push the frontiers of the technology – and the lab is looking for post docs!
An important new resource for researchers, reviewers, and funders has been developed with major contributions from the Mary Lyon Centre at MRC Harwell.
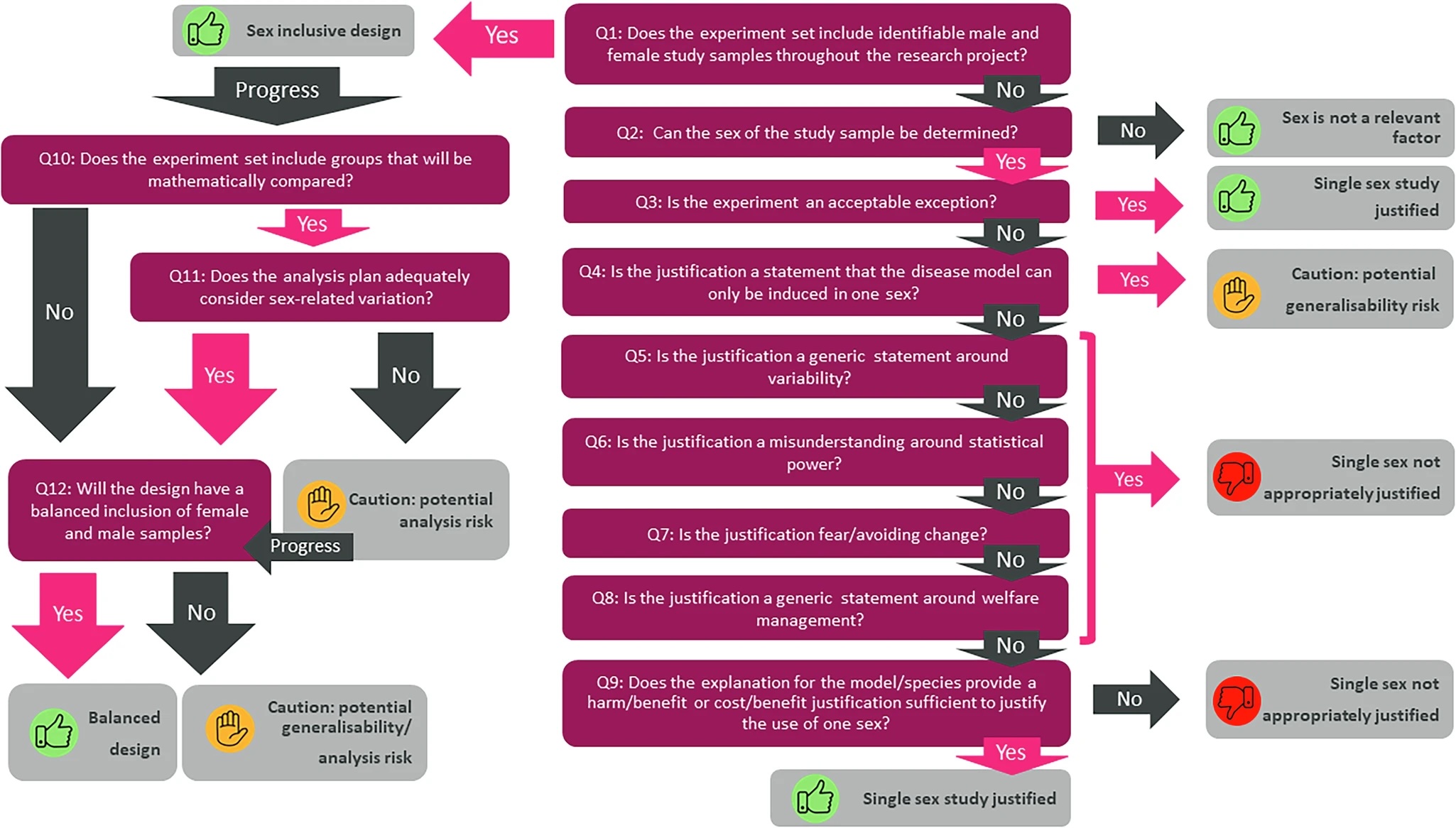
A group of researchers and policy leaders from organisations involved in in vivo and ex vivo research have unveiled a transformative tool to assist scientists, policy makers, funders and reviewers in eliminating persistent sex bias in biomedical research. Published today in Nature Communications, the Sex Inclusive Research Framework (SIRF) introduces a structured and interactive approach to ensure equitable inclusion of male and female samples in preclinical studies.
Preclinical research has long favoured male animals and cell lines, leading to a skewed knowledge base that does not represent the human population. Consequently, it leads to non-optimal use of animals and resources as it ultimately results in less reliable data and less successful therapeutic interventions. Despite mandates by funding bodies to include female and male samples (funders such as the Medical Research Council have introduced this requirement since 2022), many research proposals fall short due to ingrained misconceptions and inconsistent evaluation standards. Recent media attention on this subject was reflected by this Guardian article https://www.theguardian.com/science/article/2024/jul/22/sex-bias-labs-women-losers-research-ageing?CMP=Share_iOSApp_Other, highlighting some of the negative impacts of sex-biased research.
SIRF addresses these gaps with an intuitive, traffic light–based decision tree that evaluates the appropriateness of sex inclusion in experimental design. Developed through collaboration across academia, industry, funding agencies, and animal welfare organisations, the framework assesses whether proposals include balanced sex representation and appropriate analysis plans, and whether single-sex studies are scientifically justified.
“Sex bias in research isn’t just a scientific issue, it’s an equity issue,” said Natasha Karp, lead author, “SIRF provides the structure, rationale, and accessibility needed to make inclusive design the standard, not the exception.”
SIRF offers:
A clear, reproducible evaluation method for use by researchers, ethics boards, and funders.
Detailed guidance to debunk myths that hinder sex-inclusive practices.
An open-access web interface with interactive features and educational resources.
The framework complements and improves upon prior initiatives, such as NIH and SAGER guidelines, by emphasising transparency, scientific justification, and practical implementation.
Available now as an interactive tool or downloadable resource, SIRF is set to reshape how research proposals are developed, reviewed, and funded—ultimately enhancing the validity and impact of biomedical discoveries for all sexes.
Our Director, Sara Wells, was part of the original working group that set out the scope and direction of this work and contributed with the wealth of data and experience from years of in vivo preclinical work at the Mary Lyon Centre. Commenting on this milestone and the resource now available to the community, she said: “The translatability of in vivo experiments is a key element of the success of preclinical studies. The analysis of every variable affecting experimental outcomes is a major objective as we work towards achieving this goal. Sex has now been shown to be a fundamental piece of the data variability puzzle, and the SIRF framework supports the community in assessing where its inclusion is essential, as well as providing resources for adequate data analysis. The implementation of the framework will contribute to the community’s drive for more relevant, translatable data and effective uses of both animals and financial resources as we strive for more successful therapeutics.”
Lilian Hunt, Lead Advisor to the Executive Director of Equity, Wellcome said:
“We’re pleased to see the publication of this vital tool for research. Ensuring sex inclusion in in vivo and ex vivo research is key to ensuring excellent, reproducible, and translatable research that benefits health equity. Wellcome will be exploring how best to integrate this guidance into our expectations of inclusive research.”
As you may have seen in an earlier post, I’m moving on from Development after 13+ years as the journal’s Executive Editor. The good news (for me at least!) is that I’m not going far – I’m taking up a new position here at The Company of Biologists as the Publishing Director, overseeing our editorial programme and supporting the activities of all five of our journals. This means I’ll still be very involved with Development, though much more on the publishing side of things than the science – and this is definitely a real wrench for me. I love developmental biology, I love the community and I love working with such a fantastic and dedicated team of academic editors and in-house staff. But it was time for me to move on – after all this time, I’m ready for a new challenge. And I’m excited to announce that we’ve appointed Alex Eve, who’s been with the journal as a Reviews and then Senior Editor since late 2018, as the new Executive Editor. His knowledge of, enthusiasm for and dedication to the field mean that the journal’s going to be in great hands. You can expect to hear a bit more from Alex and his plans in the coming weeks.
As I turn my attention to my new role, I’ve also been thinking back over the past 13 years and thought I’d share some (perhaps a slightly random selection of!) particularly memorable activities and moments (big and small) from my time here at Development…
When I first joined the journal in late 2011, Olivier Pourquié had been in place as Editor-in-Chief for almost two years, and one of his major focusses was on attracting stem cell scientists to Development as a journal. Olivier, earlier than most, recognised the potential in the synergy between the established field of in vivo embryology and the burgeoning in vitro stem cell field, but also saw a divide between the two communities. I’ll admit that – at first – I was sceptical about the hype surrounding stem cell biology, but Olivier’s viewpoint was persuasive and he rapidly won me around. One of my early tasks was therefore to think about ways in which we could bring the stem cell and developmental biology communities closer together – to benefit both the journal and the fields more broadly. One major initiative in this area was the ‘From stem cells to human development’ meeting – which Olivier and I initially conceived in 2013, and which first ran in September 2014. I’ll be writing more extensively about these meetings for Development later in the year, but suffice to say that I’m super-proud of how this first meeting, and the biennial series it spawned, panned out in supporting and promoting the growing field of human developmental biology. The meeting is still going strong, and I’m delighted that – next year – the journal is partnering with the Wellcome-funded Human Developmental Biology Initiative (HDBI) to run the next edition of this conference.
Another highlight from my early years was visiting the Woods Hole Embryology course back in 2013. Nipam Patel, then the course director and an Editor at Development (and now Director of the Marine Biological Laboratory; MBL), invited me to the MBL to give a talk to the students and find out more about the course – which The Company of Biologists has been supporting for many years (look out for more on the relationship between the Company and the course in an upcoming issue of the journal). I was able to tag this visit on to an already-planned trip to Boston for the International Society for Stem Cell Research (ISSCR) Annual Meeting (incidentally leading to a hotel booking error that left me homeless for a night in Boston!), and visiting Woods Hole – albeit for just 24 hours – reinforced my regret at never having taken the course myself. As well as giving a talk, Nipam roped me in to helping the students with fly imaginal disc dissections – something I’d not done for around a decade at the time but somehow still retained the muscle memory for – and with some imaging experiments. While I’ve never regretted leaving lab science behind, it’s the dissections, injections, and transplantations that I missed – so having just a brief opportunity to do some of this again was a real pleasure for me! As well as the ISSCR meeting, that trip also took in the International Society for Developmental Biology (ISDB) conference in Cancun (and the added bonus of watching a turtle crawling up on to the beach at night to lay eggs) – making it a really memorable, if hectic, couple of weeks.
Another ISSCR meeting makes my list of conferences never to be forgotten, though for a very different reason. 23 June 2016 – I was in San Francisco while, back here in the UK, the country was voting in the Brexit referendum. As votes were being counted, I was at the President’s Reception at the ISSCR meeting, where I should have been chatting about the latest stem cell research, but was actually watching my phone, incredulous, as the ‘leave’ vote mounted – with a similarly shocked group of academics around me. I remember going to bed with the radio on, listening to David Cameron resign and thinking that I might just stay in the US (we were still in the Obama days back then…!). The UK is still feeling the negative ramifications of that vote, though I’d definitely rather be here than Stateside right now…
Back in the office, one of the contributions I feel I’ve personally made to the Company and its journals has been in pushing the preprint agenda. Again, Olivier was prescient on this front and James Briscoe – the journal’s current Editor-in-Chief – has also been very active in this area. When bioRxiv launched in late 2013, Olivier and I really felt this was a game-changer in the publishing ecosystem, so we rapidly changed journal policy to allow preprinting and initiated discussions with bioRxiv to facilitate co-submission to the journal and the preprint server. Since then, I’ve been very involved in various initiatives related to preprints – from the launch of preLights to cross-publisher discussions around preprint peer review – and it’s great to see how the uptake of preprint posting has grown in our community over the years. Moving forwards, this is something that I’ll continue to work on: the value in early sharing of manuscripts, both for the individual researcher and the broader research community, is undeniable and I see the preprint ecosystem as a benefit not a threat to journals.
Over the past couple of years, I’ve also really enjoyed setting up the Pathway to Independence programme. Kudos for this one goes to James – the idea of setting up a scheme to support postdocs going on the job market was all his, but I’m delighted to have played my part in putting this into action. We’ve just selected our third cohort of PI Fellows, and we’re looking forward to starting to work with them. It’s a privilege to help support the next generation of leaders of our field, and I do hope that – in a small way at least – we’ve helped some of them get their foot on the ladder. I hope that, going forwards, we can continue to grow a supportive network of PIs from across the breadth of our field.
From a scientific perspective, the past decade-and-a-bit has been hugely exciting too. It seems bizarre, but when I started at Development, we had no CRISPR-based genome editing, no single-cell RNAseq, and organoids were very much in their infancy (one of the earliest Review articles I comissioned and edited at the journal on was this piece from Yoshiki Sasai and colleagues; his early work on recapitulating optic cup development in vitro will forever be one of the most mindblowing papers I’ve read). To name just a few areas, we’ve also seen tremendous advances in 4D imaging of developmental processes, in our appreciation of the contribution of biophysical forces to development and in our understanding of how genomic elements interact to direct the complex and dynamic patterns of gene expression required to orchestrate development. But we’ve still got so much more to learn – and while I know that there has been a lot of angst in the developmental biology community regarding its place in the broader scientific enterprise and how the field is prioritised for funding, I maintain that (current political circumstances aside) there is no more exciting time to be a developmental biologist than now.
Finally, though, what I’m perhaps most proud of from my time at Development is the people I’ve worked with – both the academic editors, whose dedication to this role never fails to impress me, and the in-house team. When I first arrived at the Company – with zero management experience, limited knowledge of how publishing works and feeling very apprehensive about the new challenge – I was told by the then Company Secretary “you’ll be fine – Development’s got a great team”. They were right, and this continues to be true to this day. I’ve been lucky to work with a wonderful group of in-house staff, including four individuals who were part of my team when I joined and still work for the Company to this day, and several who’ve left and gone on to do fabulous things elsewhere. I hope that I’ve helped them succeed with their career aspirations, whatever they may have been, and I celebrate their ongoing successes. I’m super-lucky that two of them – Seema Grewal (now Executive Editor of Journal of Cell Science) and Alex Eve – are a part of my new team so I can continue to work with them in the months and years to come.
Right now, I’m still figuring out exactly what my new job involves – it’s a new position here at the Company – and for the time being, I’ll be focussing on learning about the other journals and their communities and really getting my head around what matters most to our authors, reviewers and readers. These are interesting times in academic publishing: there’s a fair bit of (understandable) discontent out there about how the whole process works which, combined with things like changing business models and the rise of AI-based technologies, means that there’s an awful lot for me to think about!
Join us to celebrate the finalists of Development’s 2024 outstanding paper prize, where three first authors will present their work. The webinar will be chaired by Development’s Executive Editor, Alex Eve.
Wednesday 28 May – 15:00 BST
Bridget LaMonica Ostrem (University of California, San Francisco) ‘Microglial memory: Shaping the fetal brain response to maternal inflammation’
Thamarai Athilingam (University of Warwick) ‘Concentration dependent diffusion dynamics of Bicoid in the early Drosophila embryo’
Kevin Emmerich (NIH/NHLBI Translational Vascular Medicine Branch) ‘A large-scale CRISPR screen reveals context-specific genetic regulation of retinal ganglion cell regeneration’
At the speakers’ discretion, the webinar will be recorded to view on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
Sophie Wiszniak, Dimuthu Alankarage, Iman Lohraseb, Ceilidh Marchant, Genevieve Secker, Wendy Parker, John Toubia, Melissa White, Sandra Piltz, Markus Tondl, Eleni Giannoulatou, David Winlaw, Gillian M. Blue, Congenital Heart Disease Synergy Group, Patrick P. L. Tam, Paul Thomas, Natasha Harvey, Sally L. Dunwoodie, Quenten Schwarz
Jens Bager Christensen, Alex P.A. Donovan, Marzieh Moradi, Giada Vanacore, Mohab Helmy, Adam J. Reid, Jimmy Tsz Hang Lee, Omer Ali Bayraktar, Andrea H. Brand, N. Sumru Bayin
Jade A. Phillips, Jessica Perochon, Cai T. Johnson, Matthew Walker, Colin Nixon, Mark Hughes, André Barros-Carvalho, Yachuan Yu, Louise Mitchell, Karen Blyth, Massimo Vassalli, Julia B. Cordero
Gabriel Baonza, Tatiana Alfonso-Pérez, Gonzalo Herranz, Carlos Quintana-Quintana, Carmen Gordillo-Vázquez, Yara El Mazjoub, L.M. Escudero, David G. Míguez, Elisa Martí, Nuria Martínez-Martín, Fernando Martín-Belmonte
Aishwarya Ramamurthy, Masha D Bandouil, Likhita Aluru, Esther Yoon, Nicholas Bodkin, Jennifer Z Cheng, Carina G Biar, Jeffrey D Calhoun, Gemma L Carvill
Ruben I Calderon, Nirvay Sah, Molly Huang, Ryan H. Kittle, Walee Shaik, Jennifer N Chousal, Sampada Kallol, Tony Bui, Robert Morey, Alejandra Mitre, Norah M.E. Fogarty, Claudia Gerri, Claire Zheng, Peter ME DeHoff, Pratik Home, Kathy Niakan, Heidi Cook-Andersen, Kathleen M. Fisch, Soumen Paul, Francesca Soncin
Rashi Agarwal, Joergen Benjaminsen, Katharina Lust, Clara Becker, Natalia Fuchs, Eva Hasel de Carvalho, Fanny Eggeler, Omnia El Said Ibrahim, Narges Aghaallaei, Baubak Bajoghli, Joachim Wittbrodt
Damián García-García, Dunja Knapp, Minjoo Kim, Katelyn Jamwal, Heath Fuqua, Ryan P. Seaman, Riley E. Grindle, Sergej Nowoshilow, Maria Novatchkova, Fred W. Kolling, Joel H. Graber, Prayag Murawala
Adrien Franchet, Yuhong Jin, Clare L. Newell, Victor Girard, Gérard Manière, Yaël Grosjean, Christopher Barrington, James I. MacRae, Ian S. Gilmore, Alex P. Gould
Jennifer Falconi, Miriam Rodríguez-Vázquez, Katrin Strobel, Céline Jahannault-Talignani, Lisa Heron-Milhavet, Patrice Lassus, Charles Géminard, Alexandre Djiane
Zhenzhen Peng, Thitinee Vanichapol, Phong Dang Nguyen, Hao-Han George Chang, Katrinka M. Kocha, Lori L. O’Brien, Peter D. Currie, Peng Huang, Alan J. Davidson
Declan L. Turner, Hannah Baric, Katelyn Patatsos, Sahel Amoozadeh, Michael See, Kathleen A. Strumila, Jack T. Murphy, Liam Gubbels, Elizabeth Ng, Andrew Elefanty, Melanie Neeland, Shivanthan Shanthikumar, Sarah L. Londrigan, Mirana Ramialison, Fernando J. Rossello, Ed Stanley, Rhiannon B. Werder
Alexander Walker, Paula Olaizola, Euan Brennan, Edward J Jarman, Yuelin Yao, Elizabeth Carmichael, Andreea Gradinaru, Alexander EP Loftus, David H Wilson, Anabel Martinez Lyons, Laura Charlton, Kimberley Ober-Vliegen, Wunan Mi, Amy Broeders, Kyle Davies, Neil O. Carragher, Asier Unciti-Broceta, Timothy J Kendall, Luc van der Laan, Monique MA Verstegen, Margaret C Frame, Scott H Waddell, Luke Boulter
Esra Sengul, Helen G. Potts, William T. Stockdale, Ryan D. Carter, Laura Bevan, Maria Nozdrina, Rita Alonaizan, Zhilian Hu, Abigail Goodship, Jun Ying, Konstantinos Lekkos, Lucy O’Byrne, Madeleine E. Lemieux, Rebecca Richardson, Mathilda T.M. Mommersteeg
Jennifer Saile, Hannah Walter, Moritz Denecke, Patrick Lederer, Laura Schütz, Andreas Hiltbrunner, Katharina Lepp, Sofia Lobato-Gil, Petra Beli, Andreas Wachter
Patrycja Sokolowska, Matthias Jost, Wolfram Buss, Brett Ford, Peter Michael Chandler, Wolfgang Spielmeyer, Andy Phillips, Alison K. Huttly, Danuše Tarkowská, Rocío Alarcón-Reverte, Suzanne J. Clark, Stephen Pearce, Peter Hedden, Stephen G. Thomas
Juan Zapata-Muñoz, Juan Ignacio Jiménez-Loygorri, Michael Stumpe, Beatriz Villarejo-Zori, Sandra Alonso-Gil, Petra Terešak, Benan J. Mathai, Ian G Ganley, Anne Simonsen, Jörn Dengjel, Patricia Boya
Cerys E. Currie, Aleksandra Byrska, Deborah M. Taylor, Muriel Erent, Daniela Bakalova, Xuhui Sun, Constandina Koki, Nigel J. Burroughs, Richard. A. Anderson, Adele L. Marston, Geraldine M. Hartshorne, Andrew D. McAinsh
Marina Venero Galanternik, Daniel Castranova, Ryan D. Gober, Tuyet Nguyen, Madeleine Kenton, Gennady Margolin, Aurora Kraus, Abhinav Sur, Louis E. Dye 3rd, Van Pham, Adilenne Maese, Melanie Holmgren, Aniket V. Gore, Bakary Samasa, Allison Goldstein, Andrew E. Davis, Avery A. Swearer, James Iben, Tianwei Li, Steven L. Coon, Ryan K. Dale, Jeffrey A. Farrell, Brant M. Weinstein
From Galanternik et al. This image is made available for use under a CC0 license.
Leanne E. Iannucci, Velanganni Selvaraj Maria Thomas, Micaela R. Murphy, Caitlin E.T. Donahue, William K. Anderson, Catherine E. Rogers, Allison J. Saul, Katherine W. Rogers
Dominic W.H. Shayler, Kevin Stachelek, Linda Cambier, Sunhye Lee, Jinlun Bai, Bhavana Bhat, Mark W. Reid, Daniel J. Weisenberger, Jennifer G. Aparicio, Yeha Kim, Mitali Singh, Maxwell Bay, Matthew E. Thornton, Eamon K. Doyle, Zachary Fouladian, Stephan G. Erberich, Brendan H. Grubbs, Michael A. Bonaguidi, Cheryl Mae Craft, Hardeep P. Singh, David Cobrinik
In this ‘Featured image’ post, we find out more about the story behind Özge Özgüç’s image, which was the winner of the competition.
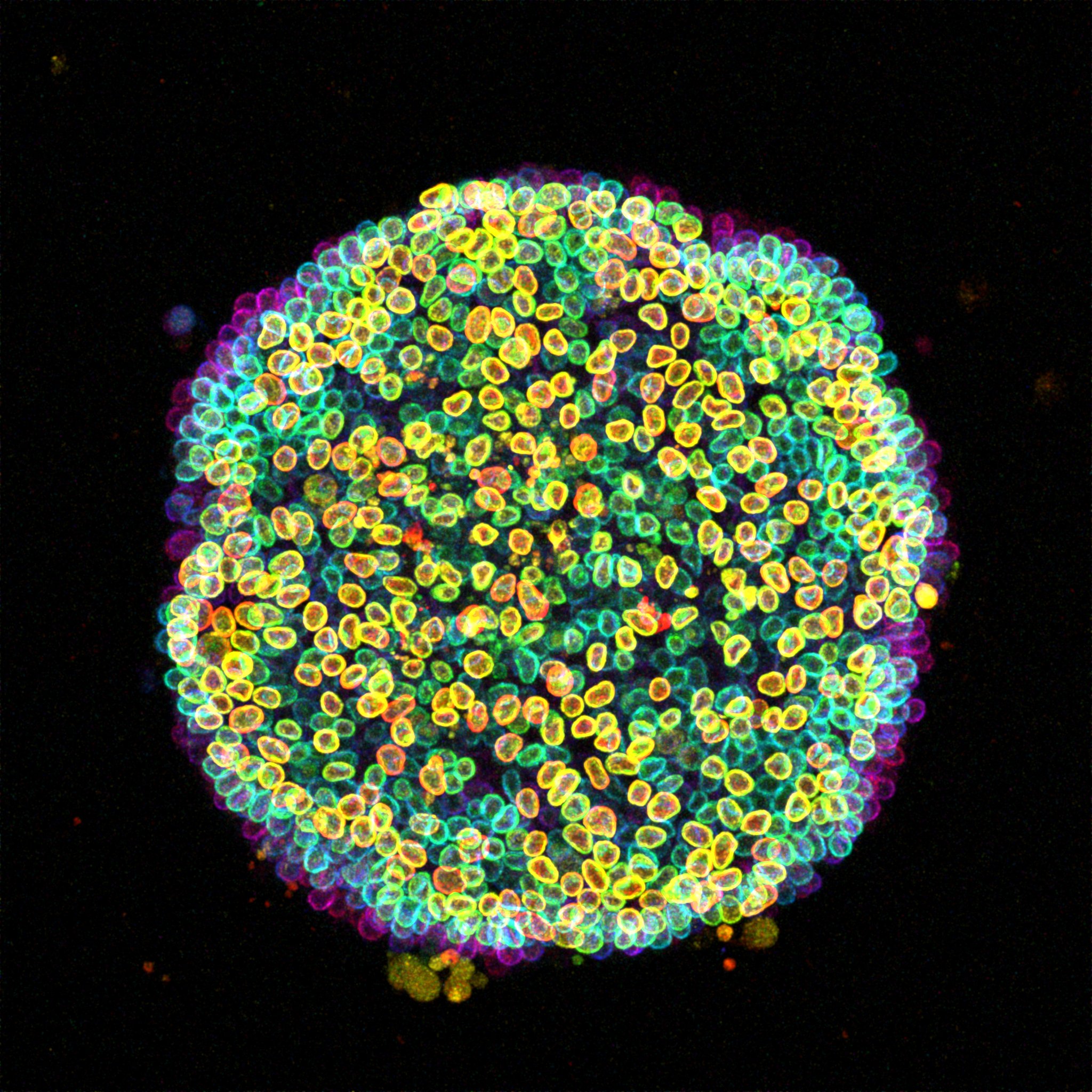
Cell-estial bloom A ‘Cell-estial Bloom’ of human induced pluripotent stem cells (hiPSCs) flourishes on a micropatterned island. This image presents a colony of live hiPSCs, with fluorescently labeled Lamin B delineating the nuclear lamina within each cell. Acquired with a Zeiss LSM 880 Airyscan microscope, this maximum intensity projection is enhanced with depth-coded coloring to reveal the captivating three-dimensional landscape.
What is your background?
I completed my undergraduate education in the Department of Molecular Biology and Genetics at Middle East Technical University (METU) in Ankara, Turkey. During this time, I participated in research across different fields of biology through various internships to discover what truly excited me. It was during one of these internships that I realized I was most interested in developmental biology. To follow this interest, I pursued a master’s in Developmental Biology at Sorbonne University in Paris/France, followed by a PhD in Cellular and Developmental Biology at Institut Curie in Paris/France in the lab of Jean-Léon Maître. During my PhD, I focused on the physical forces that shape the preimplantation mouse embryo, particularly how actomyosin contractility prepare itself for morphogenesis by transitioning from an egg to an embryo state before it becomes the driving machinery behind the morphogenetic events of preimplantation development. Currently, I’m a postdoctoral researcher at the Institute for Bioengineering of Catalonia (IBEC) in Barcelona/Spain in the lab of Xavier Trepat. My work has grown increasingly interdisciplinary, bringing together developmental biology, biophysics, and bioengineering, to explore how mechanical forces influence early developmental processes across different model systems.
What are you currently working on?
Currently, I’m working on building experimental models that allow us to study the mechanical aspects of early human development. Human post-implantation stages are notoriously difficult to access and study in vivo, so we’re developing in vitro systems that recreate aspects of this development in a controlled and mechanically accessible way. With these tools, I aim to understand how physical forces, like pressure and tissue tension, influence key cell fate decisions and morphogenetic events, such as the symmetry breaking and start of gastrulation.
Can you tell us more about the story behind the image that you submitted to the image competition?
This image comes from a live-imaging session of micropatterned human induced pluripotent stem cells (hiPSCs), with fluorescent Lamin B marking the nuclear envelope. I was curious about how the cells were packing their nuclei into such a confined space, so I applied color code for the depth. Seeing nuclei at different height with a different color revealed the layered organization which was both informative and eye-catching. I first used the image as a cover slide for my lab meeting and got very nice comments about it, so I decided to submit it to the competition. The name “Cell-estial Bloom” actually came up while chatting with colleagues because we couldn’t decide whether it looked more like a flower or a galaxy.
What is your favourite technique?
I really enjoy live imaging. Something is very captivating about watching cells move and change shape in front of your own eyes and I find it incredibly satisfying to capture dynamic processes as they unfold. But I also love techniques that let you physically interact with cells and tissues. For example, during my PhD, I used various methods to change the cell size and shape, like aspirating them into micropipettes, fragmenting, fusing, or placing them into molds. These kinds of manipulations gave me a very hands-on understanding of how cells respond to mechanical cues. So, overall, I think I’m mostly excited by techniques that combine observation with gentle intervention, where you’re not just watching biology happen, but actively nudging it to reveal how it works.
What excites you the most in the field of developmental and stem cell biology?
What excites me the most is how the development of new tools and techniques open doors to explore developmental processes that once seemed out of reach. I love how these innovations often bring together ideas from completely different fields and invite you to look at the developmental processes from a fresh angle.
The Node is turning 15 this year. If you’ve ever visited the Node to read, write and interact with the global developmental and stem cell biology community – thank you for your support!
We would really appreciate any feedback you have about the Node, to make sure the community site is still relevant and useful for you, our community members.
Please spare around 5 minutes to fill in our survey. Participants of the survey can enter a prize draw for a chance to win a bag of goodies from The Company of Biologists.
All the world’s a metabolic dance, early career scientists are leading the way!
Emerging perspectives in metabolism
Dr. Lianna W Wat, Post doctoral scholar, Stanford University
This week, we delve into the story of Dr. Lianna W. Watt, a Leading Edge fellow and a postdoctoral researcher at Stanford University, who is passionate about unraveling the intricacies of metabolism and sex differences—one fly and mouse at a time. Driven by curiosity and a deep respect for basic science, Lianna has explored how diet can rewire the way male and female bodies store and break down fat. She’s worked across model systems—from Drosophila to mammals—always with an eye toward understanding how sex-specific metabolic regulation shapes health and disease. Keep reading to discover how mentorship, curiosity, and a few bags of mini eggs helped shape Lianna’s career—and why she believes that studying both sexes is fundamental biology, essential not only for understanding disease and metabolism, but also for uncovering evolutionary principles. Check out all her work here .
What was your first introduction to the field of metabolism – what’s is your first memory?
It was actually a conversation with my future undergraduate thesis supervisor, Dr. Ian Dworkin at McMaster University. I was interviewing to join his lab as a summer research student and that was when I learned that changing the diet of flies can reduce how different male and female wing shape and size are. The idea that changing the diet could have such drastic effects on metabolism to the point that organ shape and size are altered is what first drew me into metabolic research.
Could you share your journey into studying metabolism and what inspired you to specialize in metabolic studies using Drosophila melanogaster?
My research journey in metabolism began in flies, and it was truly just luck. I was in a joint-major undergraduate program and part of the requirement was an undergraduate thesis project. I had always planned on going into medical school, so I was late to the game looking for a lab. But a new professor had just joined McMaster’s biology department (Ian), and he took a chance on me. I worked with Ian on understanding how the ratio of macronutrients, or nutritional geometry, affected how different male and female shape and size are using Drosophila wings as a model system. This summer research projected turned into an undergraduate thesis and is what made me fall in love with research. I ended up forgoing applying to medical schools and instead applied for graduate research programs. From my time with Ian, I knew I wanted to do research in sex differences, continue using Drosophila as my model, and transition to a more biomedical research question. At the time, very few labs focused on investigating sex differences but there was a new lab at the University of British Columbia (UBC) that studied sex differences in metabolism and physiology in Drosophila. This was Dr. Elizabeth Rideout’s lab, and it was the perfect fit for what I wanted to do and is ultimately where I completed my PhD.
How has your transition from working in Drosophila to working in the mammalian system been?
After my PhD, my career goal was to open my own lab that used multiple model systems to bridge the gap between basic science and clinical research. This motivation was why I transitioned to a mammalian lab for my postdoc. The transition for me was fairly smooth as I had ~1yr experience with the Kieffer and Clee labs at UBC using mice. The main differences between using flies and mice for me was how you plan experiments. In flies, you can decide to do an experiment and have the flies ready to go in 1-2 weeks and you can simply do one experiment per cohort. However, with the mice, I would need to have experiments planned over a month in advance (quarantine, breeding, weaning etc) and because it took so much time to have the correct mice for an experiment, you had to maximize what experiments you would perform on each cohort. However, after joining a mouse lab, I quickly realized that I much preferred working with flies to mice. It turns out, I am a geneticist at heart and many of the genetic tools I was used to having in my arsenal in a fly lab did not exist in the mouse world yet. Additionally, while vertebrate model systems are incredibly important for basic research, there is an emotional toll associated with solely using mammalian models. My time in a mammalian lab also helped me realize that I was more interested in understanding the basic science underlying the regulation of metabolism rather than the discovery of new therapeutics to treat metabolic disease. This together with the development of an anaphylactic allergy to mice is what solidified my return to a Drosophila model system.
Tell us how you got interested in the field of nutritional and metabolic aspects of sex differences? How do you think the fields of studying sex differences and metabolism overlap – tell us about your interests in these areas? How have both the sexes evolved to respond to nutrition and metabolic stresses?
One of my motivators for wanting to study metabolism is that my family has a history of type 2 diabetes and obesity – I recently found out that I have a genetic variant that predisposes me to obesity. While starting in the sex differences world was by luck, I decided to stay in this field because I realized just how widespread yet understudied sex differences are (almost every phenotype has a sex difference). Historically, females were omitted from studies because they didn’t show the same phenotypes as males and there was this belief that sex hormones just complicated the data. We can learn so much new biology if we were to include both sexes since males and females form naturally dichotomous groups. In the case of metabolism, sex differences can be found everywhere from the risk and prevalence of metabolic disease, the response to therapeutics, basal metabolic phenotypes (ie. fat accumulation, blood glucose levels), and the regulation of major metabolic signaling pathways such as insulin and GLP1 (Glucagon-Like Peptide-1). In the metabolism field, is it widely accepted that males and females are phenotypically very different but many studies still only investigate males because females tend to have much weaker responses to metabolic challenges such as high fat diet. To me, this is actually an extremely exciting phenotype. Why are females more protected from developing metabolic dysfunction in response to metabolic challenges? If we could figure out the mechanisms that allow females to be protected, these may be promising avenues for new therapeutics to reverse or alleviate metabolic disease.
Why do you find the basic science aspects exciting ?
I find basic science so exciting because it is the foundation of discovery. We first need to understand normal regulatory processes to understand how these processes become dysfunctional and lead to disease. By investigating how metabolism is regulated in healthy individuals and how these processes can go wrong form the foundation for the development of novel therapeutics to treat metabolic disease. Without basic science, the development of new therapeutics would be significantly hampered.
Why do you think understanding both males and female systems from a metabolic perspective is important? How is it relevant in today’s human health dynamic? Your work is focused on uncovering mechanisms explaining how sex differences in fat metabolism arise, identifying novel functions for metabolic genes and pathways that contribute to how males and females store and break down fat differently. Could you elaborate on the key findings and their implications for the field?
For many years, the metabolism field has known that males and females store and distribute fat differently, and that many metabolic diseases associated with abnormal fat storage hare a male-biased risk and prevalence. While there is a beautiful body of work investigating how sex determination factors (ie. sex chromosomes and sex hormones) establish these sex differences, we lack an understanding of the metabolic genes and metabolic pathways that act downstream of sex determination factors to contribute to the regulation of sex differences in fat metabolism. My major findings during my PhD were 1) majority of lipid metabolism genes are sex-biasedly regulated, 2) the triglyceride lipase brummer (mammalian ATGL) acts in the somatic cells of the gonad and the neurons to regulate sex differences in fat storage and fat breakdown, 3) lipid droplets are normally present in the neurons (not just diseased states) and may be sex-biasedly regulated by brummer, and 4) the sex determination factor Transformer establishes sex differences in fat metabolism in flies via the sex-biased regulation of the adipokinetic hormone (Akh) signaling pathway. These findings represent novel functions of metabolic effectors and open the doors for interesting questions such as how lipid droplet dynamics in neurons are regulated and how does this impact whole-body fat metabolism, how sex determination factors regulate downstream metabolic effectors like brummer (bmm) and Akh. Also, ATGL inhibition is being investigated in mammals and humans as a potential therapeutic but my data suggests that inhibiting bmm/ATGL function will have greater effects in males than females, thus indicating that ATGL inhibition studies need to be performed in both sexes.
Your work intersects sex differences, metabolism and aging. How do you integrate these disciplines in your research, and what unique insights have emerged from this approach?
I tend to think of sex differences as a tool to understand metabolism. For example, my broad question is how does our brain respond to a high fat diet? Are there certain regions/neuronal populations that become more or less active? How are these high fat diet-induced changes different between males and females? In this way, studying sex differences sheds light on understanding the metabolic phenotype.
You work on sex specific hormonal regulation of lipid metabolism. How difficult were those experiments? Did you have to deal with midnight timepoints or require an army of undergrads/ long hours etc.?
I think the difficulty of any experiment or technique really varies from person to person. For example, molecular techniques such as colorimetric assays and qPCRs came easily to me but I always found imaging more challenging. Having more hands on deck was always a huge bonus because it meant larger or more experiments could be done. For example, if it was just me, I could maybe screen ~5 RNAi lines simultaneously. But if I had 2-3 trainees helping me, that could easily go up to 15-20 RNAi lines. Training and mentoring the next generation of scientists has always been very important to me and I’m really grateful that I had the opportunity to work with so many amazing budding scientists – many of which are recognized as authors on my publications. As for late night timepoints – this only happened for specific experiments, namely whether circadian rhythm affected the sex difference in fat storage. For this set of experiments, I had a timepoint every 4 hours for a 24 hour period. My philosophy is that I would never have my trainees do something that I wouldn’t do myself so for these experiments, I collected all the samples. While napping on a desk wasn’t the most comfortable, I didn’t mind because I knew this data was important and it wasn’t a regularly occurring experience. I also had the added benefit of Liz (my PhD supervisor) buying me a huge bag of mini eggs to help me make it through the night haha
Building upon your findings in sex-specific fat metabolism and hormonal regulation, what are your upcoming plans? Are there particular metabolic pathways or hormonal regulators you aim to investigate further?
My plans going forward are actually to take a broader look at metabolic function. I mentioned earlier that one well-known sex difference in mammalian metabolism research is that females do not develop metabolic dysfunction to the same degree as males in response to metabolic challenges such as high fat diet. For example, in response to HFD, male mice will develop glucose intolerance and gain more body weight/fat mass than females, and male mice will also have worse cognitive defects after chronic high fat diet than female mice. This together with my previous work suggests that the brain plays a major role in regulating the sex-biased response to HFD. Thus, one major question of my postdoctoral work is what are the brain-wide effects of HFD on neuronal metabolic function? My goal is to use live, volumetric 2-photon imaging in conjunction with genetically-encoded metabolite sensors to investigate how HFD alters neuronal metabolic flux and function in male and female brains.
How are you planning to integrate insect and mammalian models to bridge basic science and therapeutic research?
My current plan for the future is to establish a lab that integrates neurobiology and molecular biology to study how the brain responds to external metabolic stressors (such as chronic diet perturbations or fasting) to regulate whole-body energy homeostasis. My primary model system will likely be Drosophila and any findings that are particularly exciting, I will also investigate in mammalian models, thus allowing me to bridge the gap between invertebrate and vertebrate systems.
What changes have you seen in the research community in regard to studying sex differences ? How do you think scientific paradigms around studying both sexes will evolve in the coming decades? Are we moving toward a more nuanced understanding, or do you see potential pitfalls?
When I started my PhD, I felt that the community acknowledged that sex differences exist but did not think they were important enough to dedicate an entire research project to. In the last decade, I have definitely seen this mentality shift to more appreciation for studies that uncover the mechanisms by which sex differences are established and controlled. We’ve also seen changes in regulations where studies need to justify why they only study one sex and more acknowledgement that what we learn from studying males may not necessarily apply to females. Studies are now also more transparent regarding which sexes are used for specific experiments. This shift towards more studies including both sexes or detailing which sex is used can only be a good thing as it provides us with more data and thus a better understanding of the normal regulatory processes of metabolism. However, even sex is a spectrum with many variations in sex chromosomes. As the field of sex differences evolve, I believe it will become increasingly nuanced until the whole spectrum of sex can be studied to the best of our ability.
How you see the future of metabolism evolve with the new upcoming tools – what techniques have you used and which tools are you most excited about ?
One roadblock that has hampered the discover of new signaling pathways that control metabolism is the identification of ligand-receptor pairs. With the advent of AI-assisted protein structure prediction (eg. AlphaFold, AlphaLigand), the ability to predict receptors for a known ligand or vice versa significantly speeds up our ability to identify metabolic molecular mechanisms. Recent advances have even been able to use AI to predict new drug therapies for example. I think AI will be a really strong tool in a basic scientist’s arsenal.
What role does curiosity play in your life, both within and outside of science?
Curiosity is a huge part of being a scientist – the desire to know more can really motivate your work. There’s this misconception that scientists know all there is to know about a subject, but if you maintain a child-like sense of wonder or curiosity, you’ll see that there is so much left to learn. When I spend time with my nieces and nephews, my favorite part is hearing their questions because really, every question can lead to a research project. I recently told my niece that our hair and our nails are made of the same thing. She asked me why and I didn’t know. But that could be a budding scientist’s first foray into research.
Were there any pivotal moments that shaped your career path? What advice would you offer to students and early-career scientists interested in exploring the intersections of metabolism and inter-organ communication?
My pivotal moment was joining the Dworkin lab for my undergraduate thesis project – if I hadn’t, I very likely wouldn’t have fallen in love with research and would have gone to medical school. For anyone interested in research, I would suggest that you think broadly and approach your research question from many angles. While my main focus is on energy metabolism, you can study this from many different points of view such as a neuroscientist or a mathematician.
How do you maintain a balance between your rigorous research activities and personal life? Are there hobbies or practices you find particularly rejuvenating?
I learned the hard way that if you don’t make time for things outside of research, you will burn out. My life outside the lab is equally as important as my time in the lab so I put more effort into planning my work week/month and experiments to maximize the likelihood that I won’t need to be in lab on the weekends or late into the night. Sometimes, that’s just impossible and I work the occasional weekend/late night. Outside the lab, I’m a huge book lover and spend a lot of time reading. I also love to cook and bake. I’ve also been an avid yogi since my undergraduate days so I try to maintain this hobby by going to yoga practice first thing in the morning – I find that waking up early is more reliable than leaving lab at the same time every day.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
I’d love to open a cozy bookstore/café hybrid! Somewhere people could get lost among the shelves with a mug of tea. Or maybe that’s just what I want to do haha !
Anything you’d want to highlight ?
I was just selected as one of 2025’s Leading Edge fellows. This is a group of women and non-binary early career scientists that support one another in obtaining R1 faculty positions and tenureship. I’m really proud to be a part of this community to elevate women and non-binary individuals in science.
Last week we learnt about how nutrient dependent signaling shapes cell fate decisions and developmental plasticity in aquatic organisms like sea anemones and planarians. Check out – Of Tor and Tide (Eudald Pascual-Carreras)




 (2 votes)
(2 votes)
 (No Ratings Yet)
(No Ratings Yet)