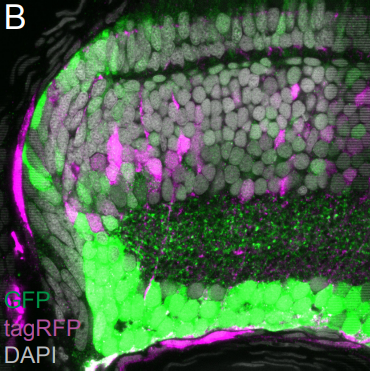
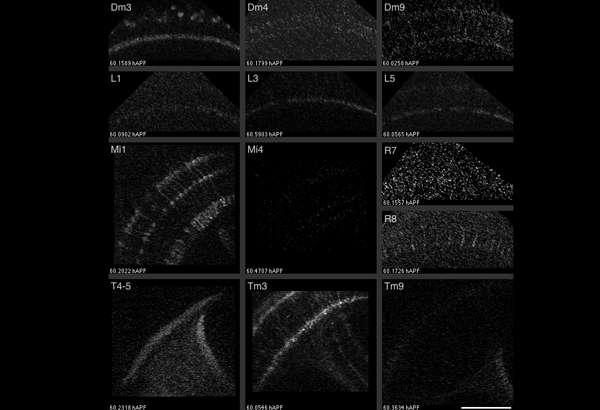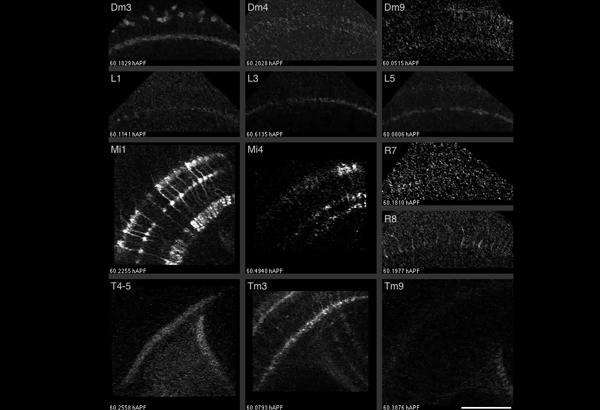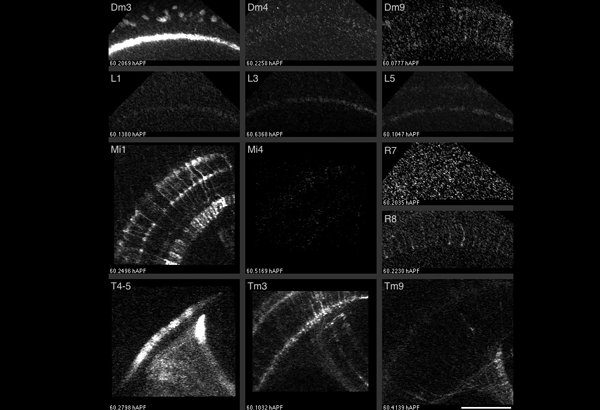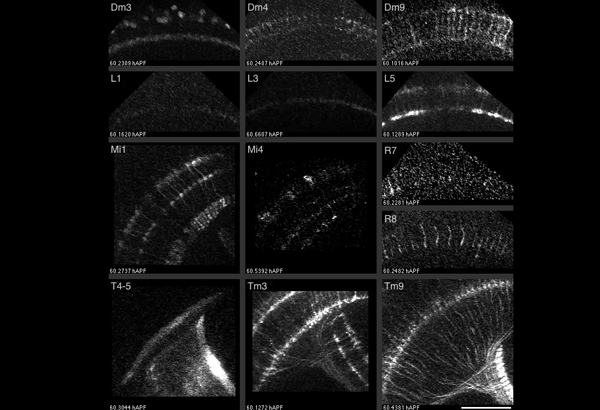
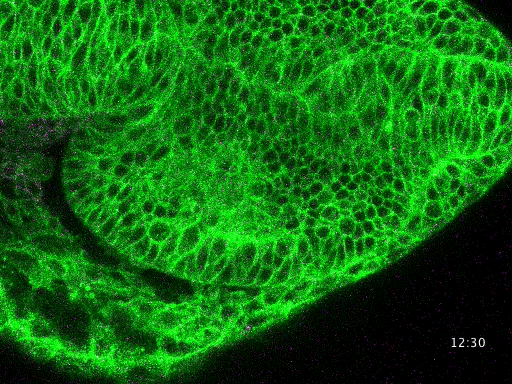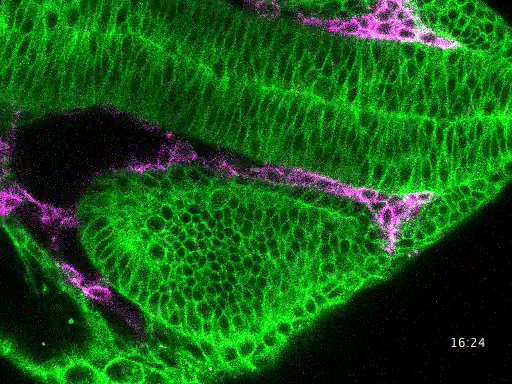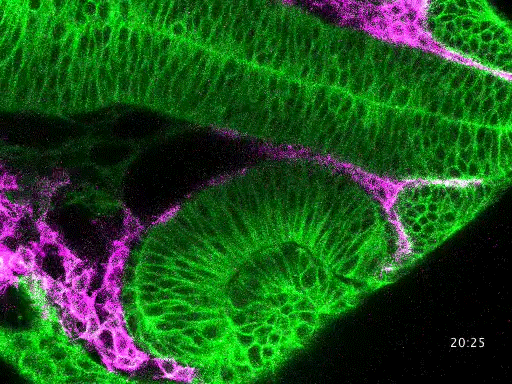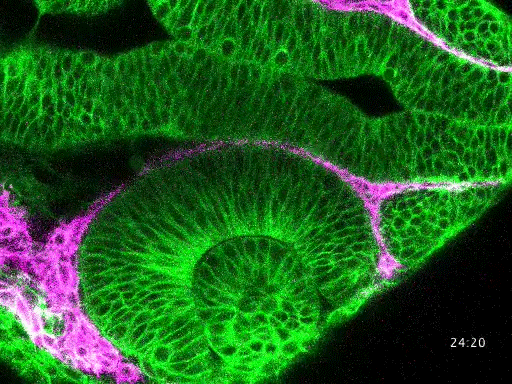
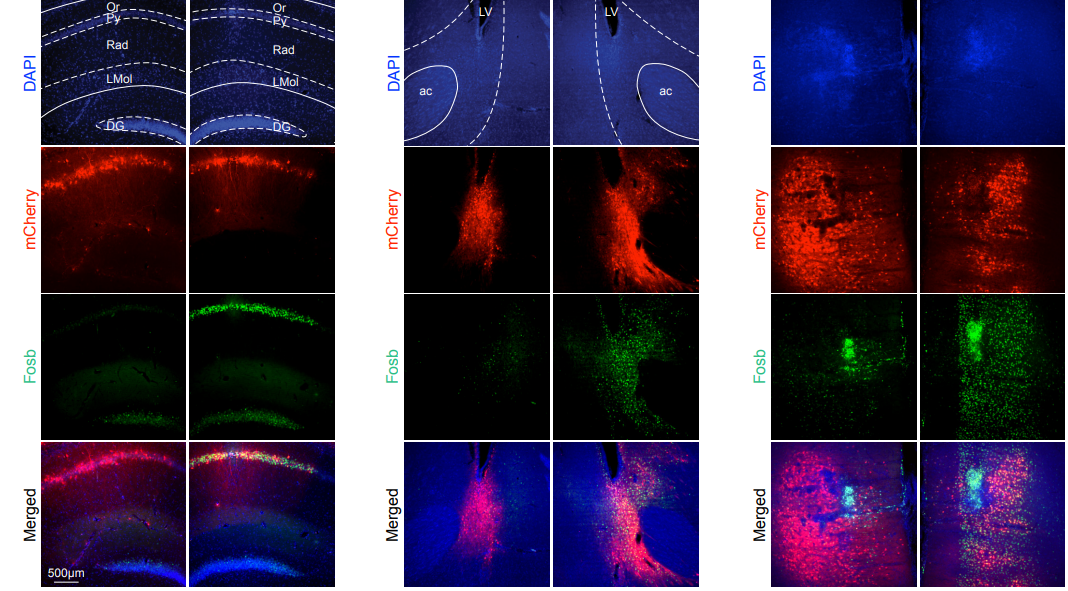
Studying mechanisms of genetic renal disease in Drosophila and mouse
The Renal Division of the University Medical Center Freiburg is offering a PhD position (DFG-funded). The position is available for three years and should be filled as soon as possible (starting in fall 2018).
Project:
The lab is interested in studying mechanisms of genetic renal disease with a focus on monogenic causes of nephrotic syndrome.
To this end we are utilizing the Drosophila and mouse animal models in conjunction with approaches in vitro.
The podocyte-like Drosophila nephrocytes will be employed to characterize disease genes functionally and for whole-animal drug screening to develop novel therapeutic strategies.
The candidate will work in diverse scientific environment with intensive supervision and support.
Requirements:
We are looking for a highly motivated and ambitious candidate with a strong interest in basic research with clinical relevance.
The applicant should hold a master’s degree in Biology, Molecular Medicine or an equivalent discipline.
Experience in cell biology, mice and/or Drosophila is desirable but not mandatory.
Application: Interested candidates should send an application including a CV, a brief motivation letter and two references to
Dr. Tobias Hermle (tobias.hermle[at]uniklinik-freiburg[dot]de).
Selected References:
1. Hermle, T, Schneider, R, Schapiro, D, Braun, DA, van der Ven, AT, Warejko, JK, et al.: GAPVD1 and ANKFY1 Mutations Implicate RAB5 Regulation in Nephrotic Syndrome. J Am Soc Nephrol, 2018.
2. Helmstadter, M, Huber, TB, Hermle, T: Using the Drosophila Nephrocyte to Model Podocyte Function and Disease. Front Pediatr, 5: 262, 2017.
3. Hermle, T, Braun, DA, Helmstadter, M, Huber, TB, Hildebrandt, F: Modeling Monogenic Human Nephrotic Syndrome in the Drosophila Garland Cell Nephrocyte. J Am Soc Nephrol,28: 1521-1533, 2017.
https://commons.wikimedia.org/wiki/File:Freiburg_Schlossbergturm_Panorama_2010.jpg (No Ratings Yet) Loading...
A postdoc position is available in the Heemskerk lab, Department of Cell and Developmental Biology at the University of Michigan Medical School.
We are looking for a postdoc that shares our excitement about using stem cells to discover the developmental mechanisms underlying embryogenesis and organogenesis. The lab is highly interdisciplinary and combines experimental and theoretical methods from biology, physics, and engineering, focusing on quantitative live-cell measurements to study spatial organization and cell fate determination in human pluripotent stem cells.
The broader institutional environment of the University of Michigan – the public university with the highest research spending in the United States – provides exceptional resources for professional development and trailblazing scientific exploration. The university is located in Ann Arbor, which is consistently named among cities with the highest quality of life in the US.
The successful candidate is a rigorous thinker with either a relevant biology background and strong interest in quantitative methods, or a quantitative background (e.g. physics, engineering, math) and strong interest in wet lab biology.
For more information see http://idseheemskerk.com.
To apply, please send a CV with publication list and three references, as well as a cover letter stating your motivation to iheemske@umich.edu.
Welcome to our monthly selection of developmental biology (and related) preLights
preLights, the preprint highlighting service supported by The Company of Biologists, has now reached over 150 posts since launching in late February. To further promote this platform, we are now featuring preLights content in three of The Company of Biologist’s journals – including Development. In addition, complementing the Node’s monthly trawl for preprints, we are starting a monthly series where we highlight some of the exciting developmental biology (and related) preLights articles. So, on to our first post!
Flies, fish and chicks
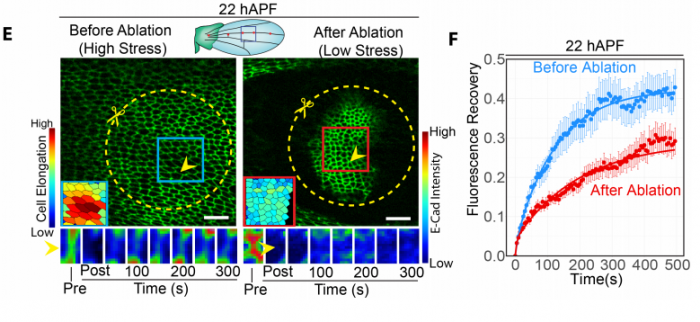
preLighters featured several exciting preprints using Drosophila. A modern imaging approach to study transcriptional dynamics, combined with mathematical modelling shed new light on the generation of the famous even-skipped stripes, and was covered by Erik Clark. Mathematical modelling was also key in getting at the mechanisms of dorsal closure in a preprint that Yara Sanchez and Arnaud Monnard reviewed together. Ivana Viktorinova’s preLight discussed how the wing epithelium responds to mechanical stress during morphogenesis, in which the mechanosensitive binding of the endocytic regulator p120 plays a major role. Drosophila embryos also have to tolerate other kind of stresses during their development, such as hypoxia; Sarah Bowling’s preLight featured a study that showed how the fat body of larvae senses low oxygen and modulates TORC1 in response.
Mechanical stress regulates E-cad turnover; reproduced from Figure 3. of the preprint by Iyer et al. and discussed by Ivana Viktorinova
There were plenty of exciting preprints coming from the zebrafish community, and the preLighters did their best to cover them. James Gagnon highlighted a fascinating study showing the essential role of a novel small protein for species-specific fertilization. Genes and phenotypes were studied at a much larger scale in a story – covered by Daniel Grimes – about an impressive effort to link functions to human schizophrenia-associated loci. The strength of zebrafish in modelling complex human diseases was also nicely featured in preLights posts by Hannah Brunsdon and Giuliana Clemente. Hannah highlighted how a putative human CHD gene variant enhances the phenotype of a known CHD-associated gene, and called attention to the importance of investigating the contribution of risk alleles to existing disease-associated phenotypes. Giuliana wrote about the introduction of elements of the human immune system into zebrafish, which could allow future research to investigate how immune cells contribute to cancer progression and metastasis. It turns out that immune cells, and specifically the spatial-temporal dynamics of cytokine induction, are crucial for spinal cord regeneration, as explored in Shikha Nayar’s preLight.
Wouter Masselink and Ashrifia Adomako-Ankomah both highlighted preprints that combined chick embryology methods with modern molecular approaches to gain new insights into one of developmental biology’s ‘classical’ problems: limb development. Wouter covered a study showing that the collinear activation of Hox genes is important for controlling the position of the forelimb. Ashrifia’s post dealt with the question of how the final size of the limb is determined and a preprint showing BMP signalling-dependent regulation of proliferation rates in the limb bud.
Chromatin and genomics
The chromatin biology and genomics fields were well represented on preLights this month. Claire Simon and Sophie Morgani discussed how Sox3 and Pou5f3 act as pioneer transcription factors to open chromatin for genome activation in Xenopus, while the organization of inactive chromatin in senescent cells was the focus of Carmen Adriaens’ post. Lauren Neves’ preLight reported on a remarkable study, which showed that the histone H3-H4 tetramer – a main component of the building blocks of chromatin – also has copper reductase activity. This enzymatic function of histones may have been important for the emergence of eukaryotes. Several preLights featured the ever-increasing power of RNA-seq to answer interesting biological questions. A transcriptomic study in plants, highlighted by Martin Balczerowicz, investigated the level of transcriptional noise among genetically identical species. Rob Hynds’ preLight discussed the use of single-cell transcriptomics combined with bulk proteomics to study healthy ageing in mouse lungs. Finally, an RNA-seq study also gave insight into how a remodelled chloroplast is maintained in a non-photosynthetic alga, preLighted by Ellis O’Neill.
Tools & Technologies
Novel technologies featured heavily this month, for instance Rebekah Tillotson’s preLight on CRISPR gene drives in mouse, which could help scientists create disease models with multiple mutations. Satish Bodakuntla covered a novel expansion microscopy technique that goes beyond the limits of super-resolution. The most popular type of method to highlight turned out to be optogenetics. Mahesh Karnani reflected on two preprints that made major advances in achieving optogenetic inhibition of neural activity by soma targeting, and the preLight includes insightful comments from both research teams. Patricia’s preLight discussed optogenetic manipulation in locust brains, while moving a bit away from neuronal function, Srivats Venkataramanan preLighted a study that used an optogenetic tool to control the formation of stress granules.
Activation of a light-gated chloride channel causes excitatory and inhibitory neurons to release neurotransmitter. Reproduced from Figure 1. of the preprint by Messier et al. and discussed by Mahesh Karnani
Finally, preLighters’ coverage of reproducibility (see Carmen’s earlier post and Reid Alderson’s preLight from this month) and flaws in experiments (see Fabio Liberante’s post on false detection of circular RNAs from RNA-seq) signalled the importance of preprints in rapid dissemination of studies which are extremely valuable for the community, but often more difficult to get published in journals.
Considering reading something more out of your scope? Then explore the preLights website, where you can also find out about microbes in a Mars-analogue environment, Mycobacteria on your showerheads, or drug repurposing to combat brain-eating amoebae.
Postdoctoral positions are available in the laboratory of Daniel T. Grimes, Institute of Molecular Biology and Department of Biology at the University of Oregon. The laboratory focuses on symmetry in the biological world.
We are a new lab opening in January 2019. We want to understand how some features of vertebrate bodies (like limbs and the skeleton) develop with left-right symmetry, while others (like the heart, liver, and gut) develop with striking left-right asymmetries.
We use zebrafish to address these fundamental questions of developmental biology. Our main techniques include genome editing, confocal and micro-CT imaging, genetics and genomics, single-cell sequencing, and more. Current projects include: understanding how mechanical flow signals are generated and sensed by cilia in left-right patterning, investigating Polycystin signal transduction, elucidating mechanisms by which the spine remains straight during growth. We also model human diseases of aberrant (a)symmetry including heterotaxia, scoliosis, and primary ciliary dyskinesia.
We offer the opportunity to work closely with the PI, and to help build and shape lab culture. Your career and manuscripts are important; we are committed to the mentorship and long-term success of our lab members. We also offer a high quality training in zebrafish development and genetics. Zebrafish research was founded at the University of Oregon. As such, we share a large state-of-the-art aquarium with labs that use the zebrafish for diverse science. There is no better place to work with zebrafish!
The Biology Department at the University of Oregon offers an exceptional environment and a broad range of research, great core facilities and close ties to the Knight Campus, a new $1-billion initiative that brings together engineers, biologists and computational scientists to address pressing biological questions.
We are looking for ambitious candidates who hold (or be due to complete) a relevant PhD and have evidence of excellent scholarship. Experience with confocal imaging is a bonus but previous experience with zebrafish is not essential. Candidates should be able to work independently, to implement new technologies, and be productive and collaborative.
The position is fully funded but the candidate will be expected to seek external funding opportunities during their tenure.
The candidate should email (dtgrimes@princeton.edu) with a cover letter describing their career goals, previous experience, scientific interests and reasons for applying, along with a CV. Informal inquiries are welcome (see grimes-lab.com/join-us-1).
The Novo Nordisk Foundation Centers for Stem Cell Biology (DanStem) and Protein Research (CPR) at the University of Copenhagen are jointly seeking applications for a 2-year position of Bioinformatics Specialist.
The Novo Nordisk Foundation Center for Stem Cell Biology (DanStem) addresses basic research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new therapeutic strategies for cancer and chronic diseases. Learn more about DanStem at https://danstem.ku.dk/.
The Novo Nordisk Foundation Center for Protein Research (CPR) promotes basic and applied research on proteins of medical relevance. Spanning in silico biology, proteomics, biochemistry and cell biology, and the unifying research theme is the identification and functional exploration of proteins and protein pathways involved in disease. Learn more about CPR at www.cpr.ku.dk.
Background
Both DanStem and CPR are vibrant, internationally diverse and ambitious research centers housing modern laboratories with state-of- the-art facilities located in close proximity to one another at the Faculty of Health and Medical Sciences, University of Copenhagen. The setting is ideally suited for seamless collaboration and exchange of expertise, material and lively scientific communication. DanStem and CPR collaboratively operate three research platforms in imaging, flow cytometry and genomics. These platforms are staffed with experts who provide advanced training and tailored project planning and assistance. The Genomics Platform provides high-throughput sequencing services with personalized support, covering applications from classical genomics to single-cell transcriptomics and has generated a demand for bioinformatics support. Therefore, the Centers intend to develop a new informatics platform to full fill the need of bioinformatics and computational approaches to data analysis enhancing the scientific synergies between the two centers.
We are now seeking a highly motivated and qualified bioinformatician with specialty in genomics to work within the Informatics Platform at DanStem and CPR.
Responsibilities
The Bioinformatics Specialist will provide bioinformatics support, develop new pipelines, and organize practical training to assist DanStem and CPR researchers with analysis and interpretation of primarily genomics data. The bioinformatician will work within the newly developing Informatics Platform, as well as with the dedicated sequencing specialist to provide tailored service and support for multiple research projects of broad biomedical scope throughout the two research centers.
This Bioinformatics Specialist is expected to:
Develop tailored bioinformatics approaches and provide advice and assistance to DanStem and CPR researchers in their analyses, focusing in particular on the planning, processing, analysis and publication of transcriptomic and epigenomic next-generation sequencing data.
Prepare pipelines and scripts for use by DanStem and CPR researchers.
Provide training and design new approaches to analysis.
Cooperate and collaborate with DanStem and CPR researchers.
Cooperate with platform personnel and contribute to platform management.
Network actively with the global bioinformatics community and stay up-to-date with current trends in the field.
Similar to our other platform specialists, the Bioinformatics Specialist will sit at the nexus of the two centers. To ensure good support and guidance, the NGS Bioinformatics Specialist will work closely with the Big Data Management Platform at CPR, which handles data from other technology platforms, as well as among research groups that cover many systems-level aspects of biology and medicine, including the integration of molecular level and healthcare data.
Qualifications, competences and experience
Candidates are expected to have a Master’s or PhD degree in bioinformatics, computational genomics or biology, computer science and data analysis, or a biological or natural sciences discipline, with at least four years of experience in bioinformatics for genomics. In addition, we are seeking a candidate with the following competences and experiences:
Fluent programming skills in R/Bioconductor.
Experience with next generation sequencing data.
Strong statistics skills.
Knowledge of UNIX-like operating system.
Experience in working with biologists and an understanding of molecular biology and genomics.
Excellent oral and written communication skills and fluency in English.
Successful experiences in project management, including multi-tasking.
A service-minded and team player attitude.
Employment conditions
We offer
State-of-the-art computational infrastructure for data analysis, including access to cluster computing at the Danish National Supercomputer (Computerome) and excellent IT and systems administration support.
Stimulating and multifaceted research environment of high scientific and societal impact.
Possibility for continued education and training, especially to develop further as an expert in single cell bioinformatics.
With an initial duration until the end of 2020 with possibility of extension, employment can begin in November 2018 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. The position, with formal employment title of Senior consultant at the University of Copenhagen, will be in accordance with provisions of the collective agreement between the Danish Confederation of Professional Associations (AC/PROSA) and the Danish Ministry of Finance. To the salary is added a monthly contribution to a pension fund according to the collective agreement. A supplement could be negotiated, depending on the candidate’s experiences and qualification.
Questions
Informal inquiries can be directed to Chief Operating Officer Gretchen Repasky.
Your online application must be submitted in English by clicking ‘Apply now’ below.
Please note that only online applications will be accepted.
The application must include a statement of motivation, curriculum vitae, and copies of relevant diplomas.
Application deadline: September 2, 2018.
Only applications received in time and consisting of the documents stated above will be considered.
University of Copenhagen and The Centers highly value diversity and welcome applications from all qualified candidates, regardless of personal background and status.
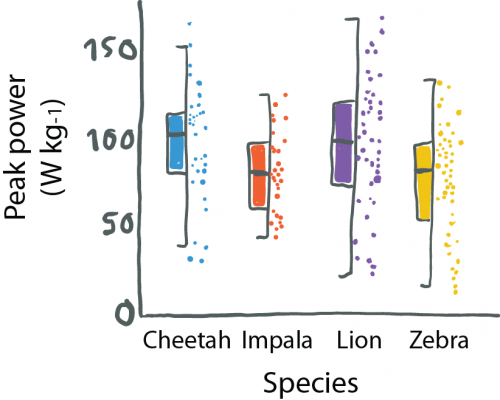
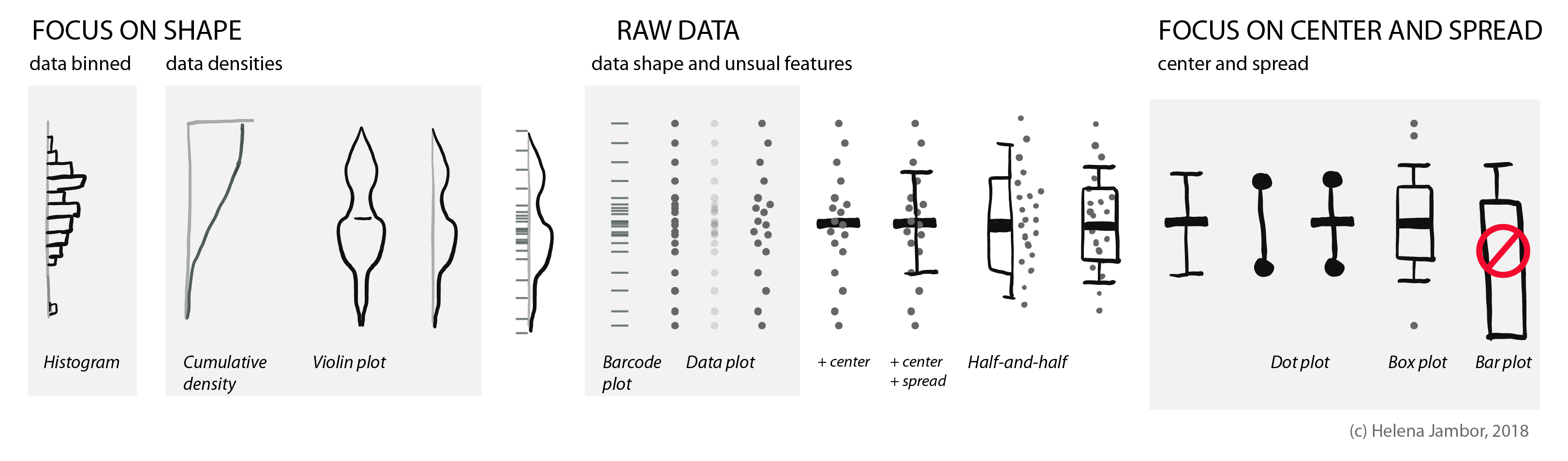
When reading about co-evolution of prey and predators, I stumbled across a cute new plot type: a half boxplot, half dot plot to show data distributions.
Half boxplot, half data plot. Figure re-drawn from Wilson et al. 2018 (doi: 10.1038/nature25479).
Wilson used this plot to simultaneously visualize summaries about their data (center, spread) and the actual data points. This allows us, the audience, to learn a lot about their results. That cheetahs are maybe binomially distributed and have outliers, or that zebras show a curious clustering.
Your quick guide to distribution plots:
The half-and-half, aka dox-plot (a friend), led me to explore which visuals are commonly used for showing distributions.
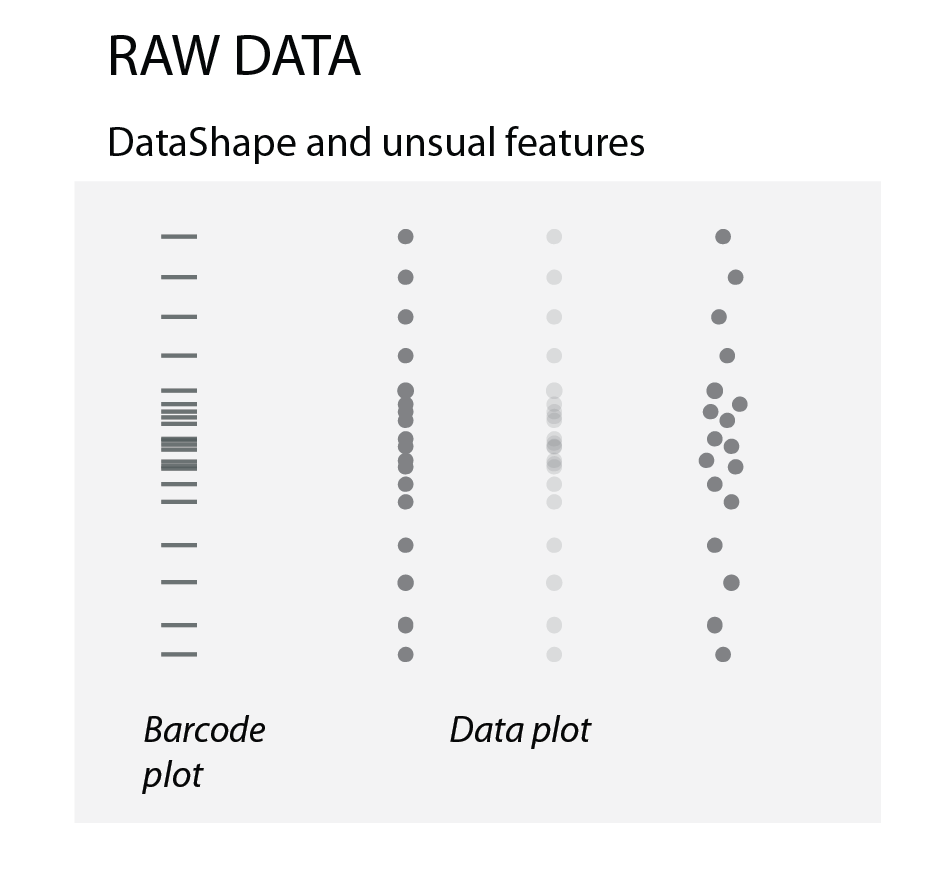
Raw data
To show how the raw data is distributed, we simply use dots or bars (as in barcode plots). When there are overlapping data points, we can use transparency or “jittering”. Jittering is distributing the data points in a given area, for increased clarity: the y-position remains the same, the x- position becomes random.
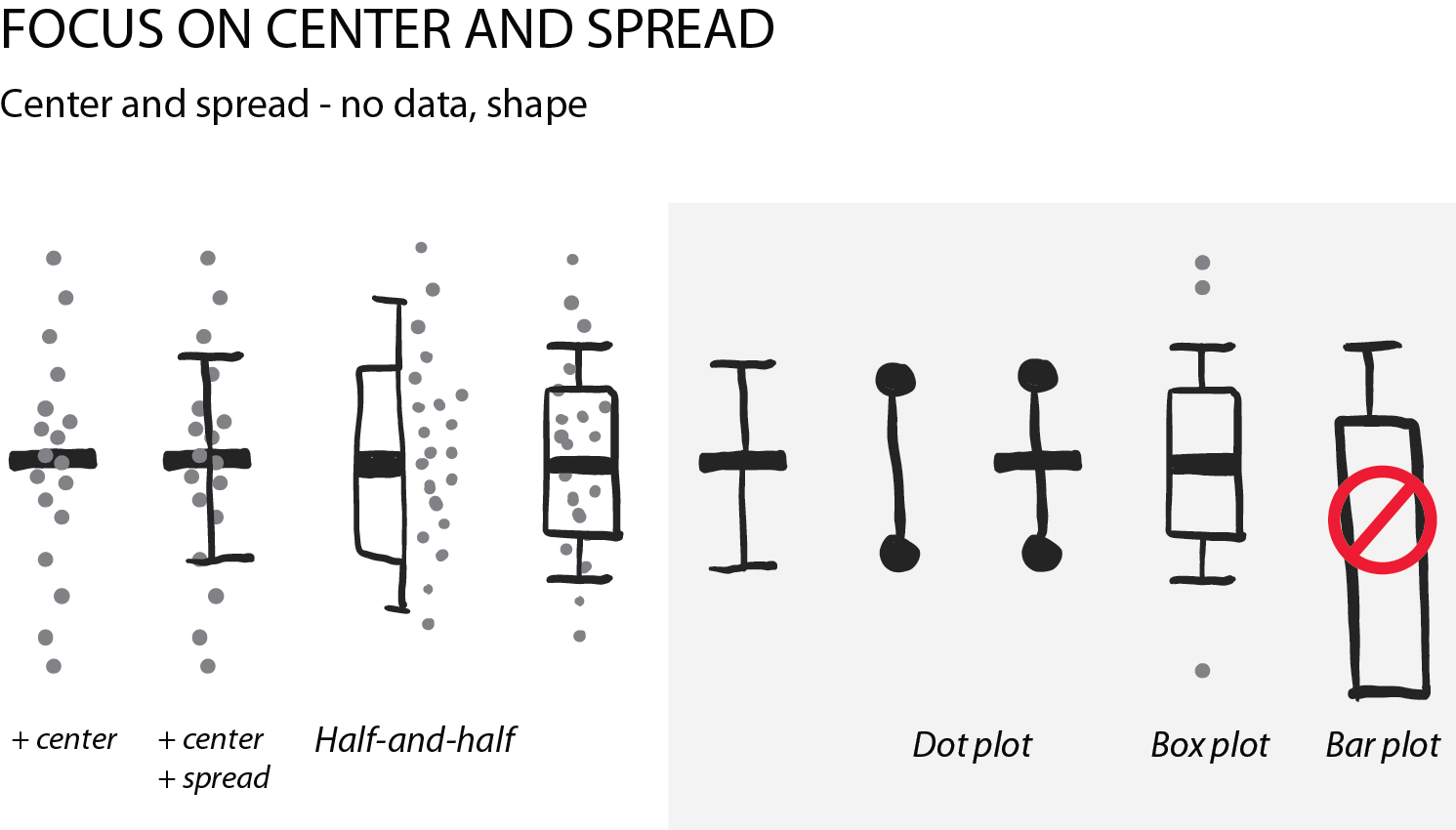
Summarizing the data: center and spread
Often, we are interested in summarizing statistics to judge and compare data. By convention the center (median) is indicated with a horizontal bar and the spread (variance, standard deviation) with vertical whiskers. Common plot types for this are the “star-wars rebel fighter”, the dot plot and boxplot. Bar plot used to be widely used, but are now banned by most journals for concealing most relevant information, so they are here only for completeness (see previous post). Very often these days I see boxplots that are overlaid with the data points – this works really well for up to 100 data points and is easy to implement with most software.
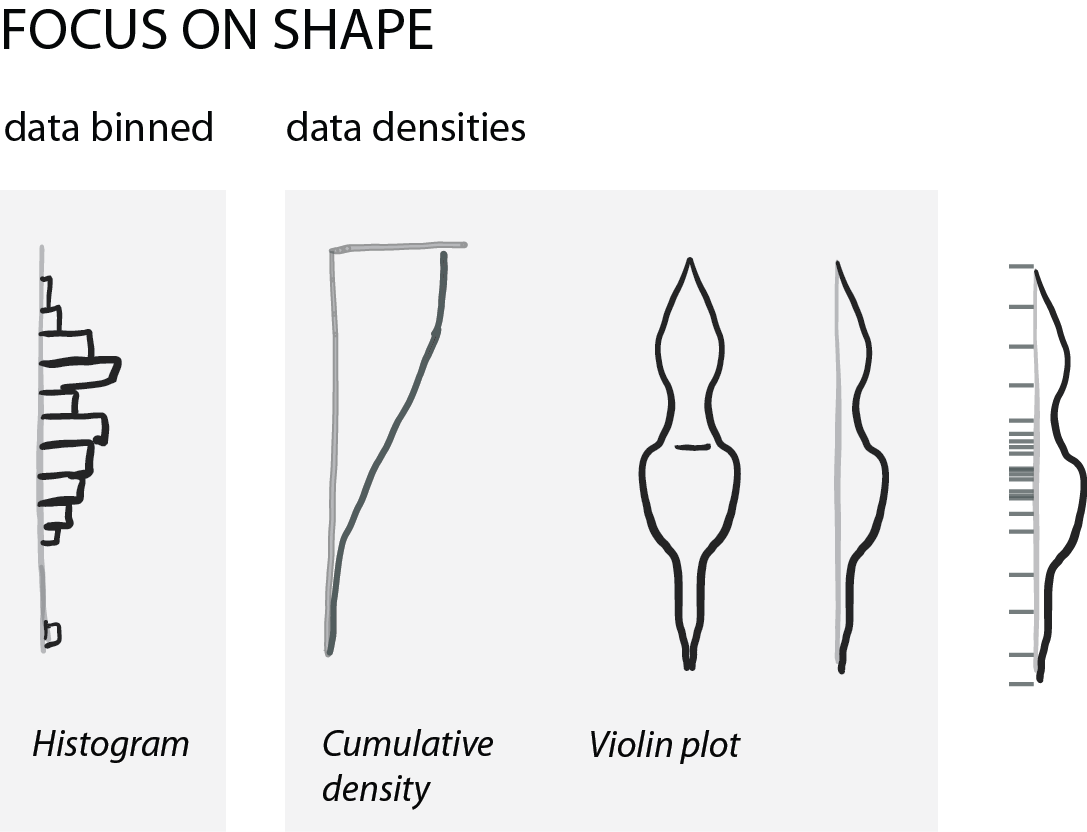
Show shape and unusual features
For normally distributed data, the center and the spread are highly informative. However, in life science we often have bimodal distributions, clusters, or gaps. Then boxplots become very insufficient and might even conceal interesting aspects (if not outright be misleading).
For faithfully showing distributions, histograms have a long history. Here, one has to be very careful with choosing bin sizes: too large or too small bins can greatly distort the histogram shape and result in a misleading chart. Choosing bin sizes is a science in itself, for details see wikipedia – but basically, it again depends on the data shape and sampling depth.
An alternative to histograms are density plots. Density plots show how data are distributed. They become very useful for large data sets. For large data sets individual points can’t be visualized anymore and the eye can’t anymore judge spread intuitively. A rather recent but so far “happy marriage” is the violin plot. Violin plots are a fusion of the boxplot and its summary statistics, with the density/shape of the data (Hintze and Nelson, 1998 doi: 10.1080/00031305.1998.10480559).
In LM Escudero´s group, we like developmental biology, mathematical biology and computational biology. We try to be imaginative and get inspiration from simple things… such as a toilet paper roll. Using this tool (and some computers), we claim that we have described a novel geometrical shape… You will be wondering… how do you do that??? You are going to find here: the story behind the discovery of “scutoids”.
First, you need a multidisciplinary group distributed between two buildings: “the wet lab”, with flies and their epithelial tissues, at Institute of Biomedicine of Seville, IBiS (Fig. 1); and the “computer lab” with… well… with a bit of everything (it does not fit in a simple picture… so, see Video 1) at the Faculty of Biology of University of Seville. It is ideal if the two buildings are separated by 1 km, so you can exercise everyday walking between the two labs to interact with the members of each branch.
Fig. 1. A. Tagua (at right, masterful segmenter and drosophilist) and C. Gordillo looking for more scutoids in the fly lab.
Video 1. The computational lab (that is wet if you open the tap) showing P. Gómez-Gálvez and P. Vicente-Munuera, the two first authors, working hard and listening good music.
Very important! You need also an office where both computational and Drosophila worlds can fit together (Fig. 2).
Fig. 2. Very trendy office. Please note the a/c controller, essential if you are based in Seville.
Then, you need a problem to think about while you are walking between the two labs… As animals develop, the initial simple planar epithelia of embryos must be sculpted into complex three-dimensional tissues. These cells pack together tightly. To accommodate the curving that occurs during embryonic development, it has been assumed that epithelial cells adopt either columnar or bottle-like shapes.
We thought that a very logic way to approach the problem of how curved epithelia pack in 3D was to design a computer model with the shape of a toilet paper roll (Fig. 3). The results we saw were weird. Our model predicted that, as the curvature of the tissue increases, columns and bottle-shapes were not the only forms that cells may developed. To our surprise, the discovered geometric solid didn’t even have a name in math! We were happy. Really happy. One does not normally have the opportunity to name a shape. We chose the name scutoid.
Fig. 3. The toilet paper: reality versus model.
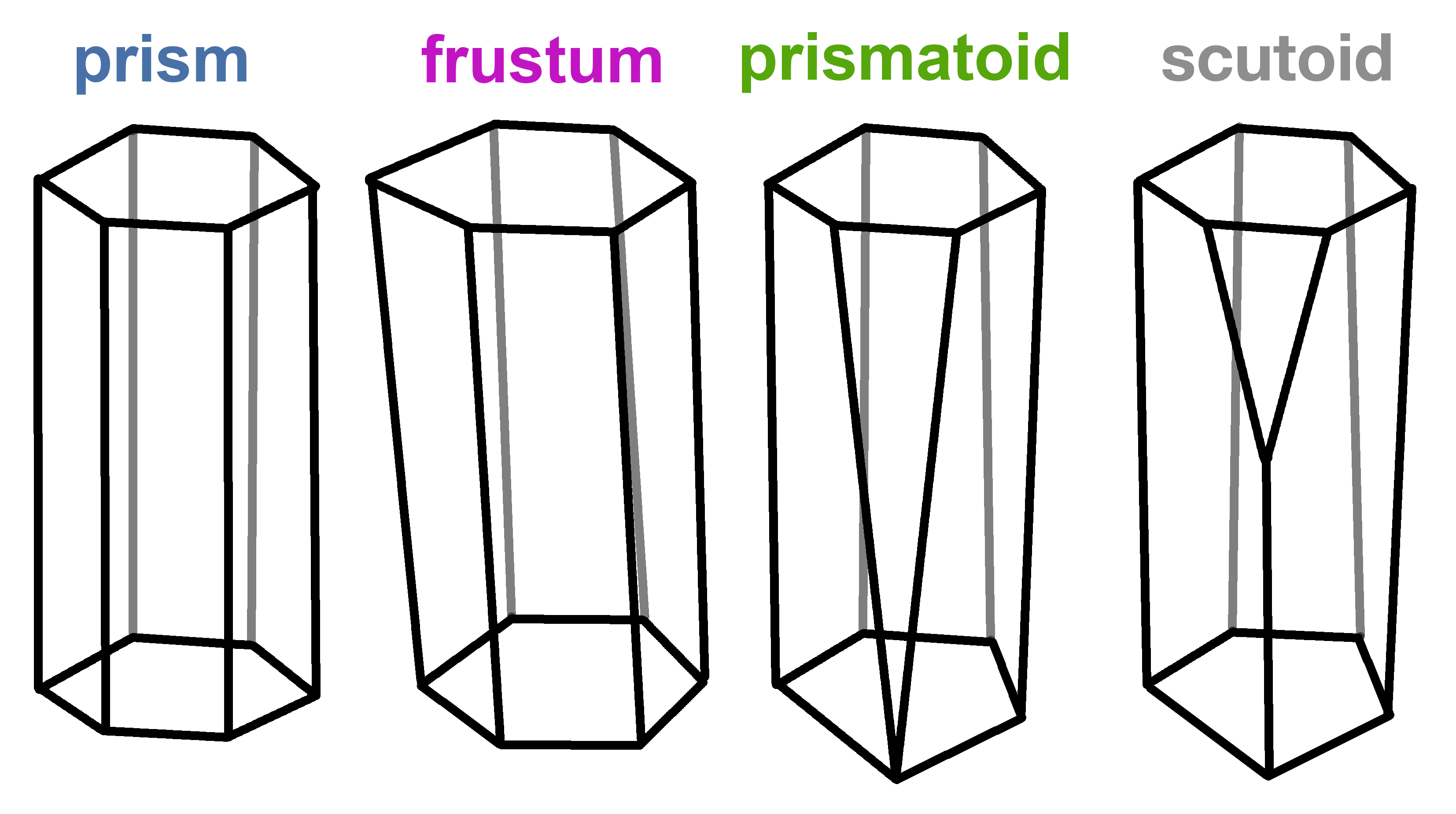
The undescribed shape was characterised by having at least a vertex in the lateral surface (Fig. 4). This vertex confers an interesting property to the scutoid: when cells adopt this shape, they can have different neighbours in the upper and bottom surfaces (apical and basal in biology). This is exclusive of scutoids and cannot be done with the “prism”, “trunk” (frusta) or “prismatoid” shapes.
Fig. 4. The brotherhood of the cellular geometry: evolution from prisms to the undescribed scutoid geometrical shapes.
All this was amazing… but we needed help to completely understand the problem. We needed lots of collaborators (we are 16 authors), starting with Dr Grima and Dr Márquez, real mathematicians that helped us with the formal aspects of the toilet paper model.
To verify the model’s predictions, the group investigated the three-dimensional packing of different tissues in different animals. The experimental data, using Drosophila salivary glands, confirmed that epithelial cells adopted shapes and three-dimensional packing motifs similar to the ones predicted by the computational model. This important validation required more collaborators: Dr Sotillos and Dr Martín-Bermudo groups (CABD, CSIC/JA/UPO, Seville) and Dr Cavodeassi (St. George’s University of London) all with biological background but working in other types of epithelia.
And then, the biophysics. We joined efforts with the lab of Dr Javier Buceta (Lehigh University), to study the role of scutoids in tissue architecture from a mechanical point of view. We argue that the scutoids stabilise the three-dimensional packing and make it energetically efficient. Our conclusion was that we have uncovered nature’s solution to achieving efficient epithelial bending. This was really cool… but not so much as the summary that Dr Buceta has prepared for you if you do not have time to read the whole paper… this is art and science in a single sheet (Fig. 5, click to see at full resolution).
Thanks for reading!!!
If you need more information, you can check our paper: Scutoids are a geometrical solution to three-dimensional packing of epithelia. Gómez-Gálvez P, Vicente-Munuera P, Tagua A, Forja C, Castro AM, Letrán M, Valencia-Expósito A, Grima C, Bermúdez-Gallardo M, Serrano-Pérez-Higueras Ó, Cavodeassi F, Sotillos S, Martín-Bermudo MD, Márquez A, Buceta J, Escudero LM. Nat Commun. 2018 Jul 27;9(1):2960. doi: 10.1038/s41467-018-05376-1.
An opportunity is available for a Postdoctoral position in the Cox Lab at the Peter MacCallum Cancer Centre in Melbourne, Australia. The position requires a highly motivated and enthusiastic postdoctoral scientist to investigate how metabolic reprogramming contributes to liver regeneration and cancer using zebrafish (Danio rerio) as a model organism. The successful candidate should hold a PhD in biochemistry, molecular biology, developmental biology or a related discipline. The person will be expected to conduct rigorous, valid and ethical research both independently and as part of the research team. The person will be expected to supervise undergraduate and postgraduate students, and technical staff. For more information on recent publications and projects running in the Cox laboratory refer to: https://www.petermac.org/research/labs/andrew-cox
Welcome to our monthly trawl for developmental biology (and other related/just plain cool) preprints.
In one of the most contentious (at least on Twitter!) pieces of preprint news in July, Tom Sheldon, Senior Press Manager at the Science Media Centerin London, voiced his concerns about the impact of preprints on public understanding of science in his ‘World View’ in Nature. Sheldon, building on an earlier SMC blog post, pictured the harm that could be done if bad science were to be deposited on preprint servers and picked up by journalists, and wondered how rigorous science would fare in a journalistic ecosystem which prioritises breaking the story first (i.e. giving credence to the preprint, not the peer reviewed article). Here’s a completely unscientific selection of the mainly negative Twitter responses to the piece, from Michael Eisen(who in fact has 3 preprints in this month’s haul!), James Fraser, Leslie Vosshall and Alejandro Sanchez Alvarado. The arguments swirled around the legitimacy of peer review, the responsibility of journalists to vet their stories properly, and the responsibility of scientists and university press offices not to oversell their results.
Away from the tumult, there was so much beautiful research deposited as preprints in July, from the molecular drivers of neurogenesis to the derivation of platypus pluripotent stem cells, cephalopod limb patterning to fern shoot development (and, right at the bottom, a truly humungous humdinger of a fungus!).
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Let us know if we missed anything, and use these links to get to the section you want:
Selective auxin agonists induce specific AUX/IAA protein degradation to modulate plant development
Thomas Vain, Sara Raggi, Noel Ferro, Deepak Kumar Barange, Martin Kieffer, Qian Ma, Siamsa Melina Doyle, Mattias Thelander, Barbora Pařízková, Ondřej Novák, Alexandre Ismail, Per Anders Enquist, Adeline Rigal, Małgorzata Łangowska, Sigurd Ramans Harborough, Yi Zhang, Karin Ljung, Judy Callis, Fredrik Almqvist, Stefan Kepinski, Mark Estelle, Laurens Pauwels, Stéphanie Robert
CrLFY1 promoter expression in fern tissues, from Langdale, et al.’s preprint
Nashwa Araby, Soha Soliman, Eman Abdel Raheem, Yasser Ahmed
Radial F-actin Organization During Early Neuronal Development
Durga Praveen Meka, Robin Scharrenberg, Bing Zhao, Theresa Koenig, Irina Schaefer, Birgit Schwanke, Oliver Kobler, Sergei Klykov, Melanie Richter, Dennis Eggert, Sabine Windhorst, Carlos G. Dotti, Michael R. Kreutz, Marina Mikhaylova, Froylan Calderon de Anda
The Nucleome of Developing Murine Rod Photoreceptors
Issam Al Diri, Marc Valentine, Beisi Xu, Daniel Putnam, Lyra Griffiths, Marybeth Lupo, Jackie Norrie, Jiakun Zhang, Dianna Johnson, John Easton, Abbas Shirinifard, Ying Shao, Victoria Honnell, Sharon Frase, Shondra Miller, Valerie Stewart, Xiang Chen, Michael Dyer
Role of Cnot6l in maternal mRNA turnover
Filip Horvat, Helena Fulka, Radek Jankele, Radek Malik, Jun Ma, Katerina Solcova, Radislav Sedlacek, Kristian Vlahovicek, Richard M Schultz, Petr Svoboda
Signalling pathways drive heterogeneity of ground state pluripotency
Kirsten R McEwen, Sarah Linnett, Harry G Leitch, Prashant Srivastava, Lara Al-Zouabi, Tien-Chi Huang, Maxime Rotival, Alex Sardini, Thalia E Chan, Sarah Filippi, Michael Stumpf, Enrico Petretto, Petra Hajkova
Human intestinal organoids in Capeling, et al.’s preprint
Towards an autologous iPSC-derived patient-on-a-chip
Anja Patricia Ramme, Leopold Koenig, Tobias Hasenberg, Christine Schwenk, Corinna Magauer, Daniel Faust, Alexandra K. Lorenz, Anna-Catharina Krebs, Christopher Drewell, Kerstin Schirrmann, Alexandra Vladetic, Grace-Chiaen Lin, Stephan Pabinger, Winfried Neuhaus, Frederic Bois, Roland Lauster, Uwe Marx, Eva-Maria Dehne
Need for high-resolution Genetic Analysis in iPSC: Results and Lessons from the ForIPS Consortium
Bernt Popp, Mandy Krumbiegel, Janina Grosch, Annika Sommer, Steffen Uebe, Zacharias Kohl, Sonja Ploetz, Michaela Farrell, Udo Trautmann, Cornelia Kraus, Arif B Ekici, Reza Asadollahi, Martin Regensburger, Katharina Guenther, Anita Rauch, Frank Edenhofer, Juergen Winkler, Beate Winner, Andre Reis
Differentiating hIPSCs from Burke, et al.’s preprint
Dissecting transcriptomic signatures of neuronal differentiation and maturation using iPSCs
Emily E Burke, Joshua G Chenoweth, Joo Heon Shin, Leonardo Collado-Torres, Suel Kee Kim, Nicola Micali, Yanhong Wang, Richard E Straub, Daniel J Hoeppner, Huei-Ying Chen, Alana Lescure, Kamel Shibbani, Gregory R Hamersky, BaDoi N Phan, William S Ulrich, Cristian Valencia, Amritha Jaishankar, Amanda J Price, Anandita Rajpurohit, Stephen A Semick, Roland Bürli, James C Barrow, Daniel J Hiler, Stephanie Cerceo Page, Keri Martinowich, Thomas M Hyde, Joel E Kleinman, Karen F Berman, José A Apud, Alan J Cross, Nick J Brandon, Daniel R Weinberger, Brady J Maher, Ronald DG McKay, Andrew E Jaffe
Sexual Dichromatism Drives Diversification Within a Major Radiation of African Amphibians
Daniel M Portik, Rayna C Bell, David C Blackburn, Aaron M Bauer, Christopher D Barratt, William R Branch, Marius Burger, Alan Channing, Timothy J Colston, Werner Conradie, J. Maximillian Dehling, Robert C Drewes, Raffael Ernst, Eli Greenbaum, Václav Gvoždík, James Harvey, Annika Hillers, Mareike Hirschfeld, Gregory Jongsma, Jos Kielgast, Marcel T Kouete, Lucinda P Lawson, Adam D Leaché, Simon P Loader, Stefan Lötters, Arie van der Meijden, Michele Menegon, Susanne Müller, Zoltán T Nagy, Caleb Ofori-Boateng, Annemarie Ohler, Theodore J Papenfuss, Daniela Rößler, Ulrich Sinsch, Mark-Oliver Rödel, Michael Veith, Jens Vindum, Ange-Ghislain Zassi-Boulou, Jimmy A McGuire
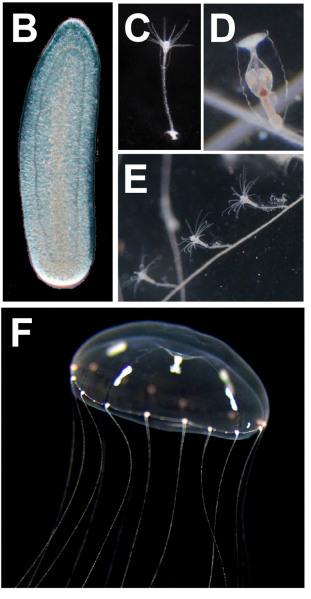
The Clytia life cycle, from Leclère, et al.’s preprint
The genome of the jellyfish Clytia hemisphaerica and the evolution of the cnidarian life-cycle
Lucas Leclère, Coralie Horin, Sandra Chevalier, Pascal Lapébie, Philippe Dru, Sophie Peron, Muriel Jager, Thomas Condamine, Karen Pottin, Séverine Romano, Julia Steger, Chiara Sinigaglia, Carine Barreau, Gonzalo Quiroga-Artigas, Antonella Ruggiero, Cécile Fourrage, Johanna Kraus, Julie Poulain, Jean-Marc Aury, Patrick Wincker, Eric Quéinnec, Ulrich Technau, Michaël Manuel, Tsuyoshi Momose, Evelyn Houliston, Richard Copley
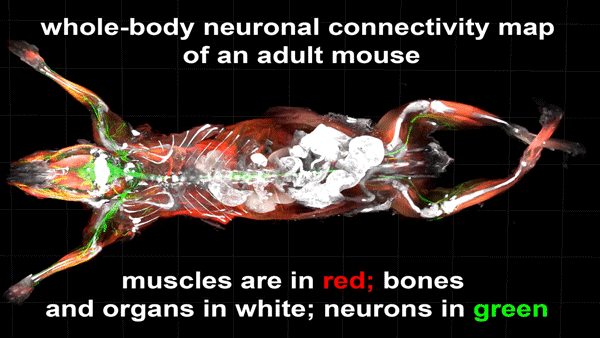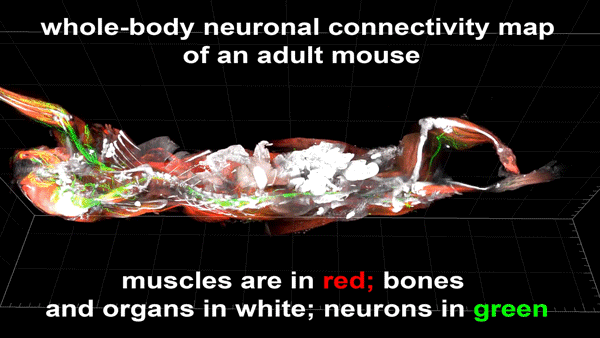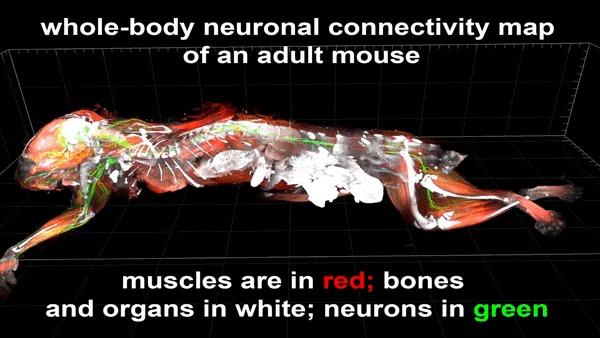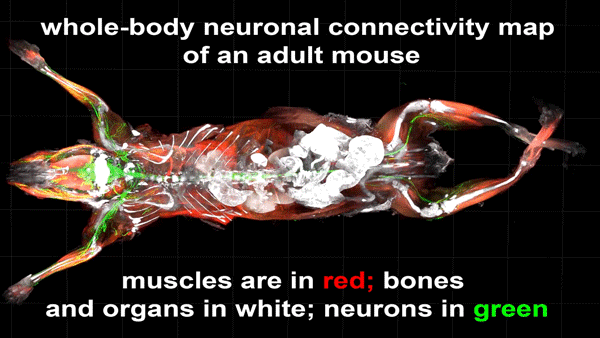
Cortical Column and Whole Brain Imaging of Neural Circuits with Molecular Contrast and Nanoscale Resolution
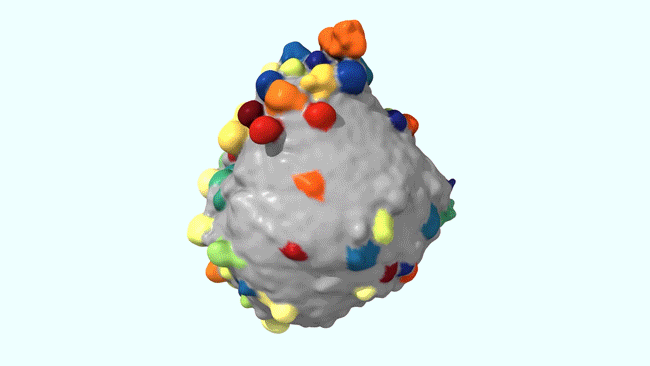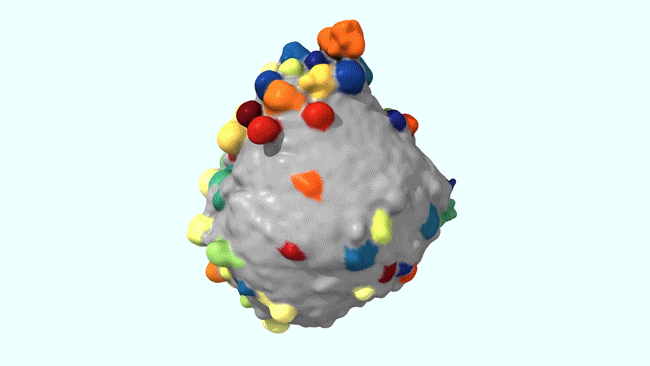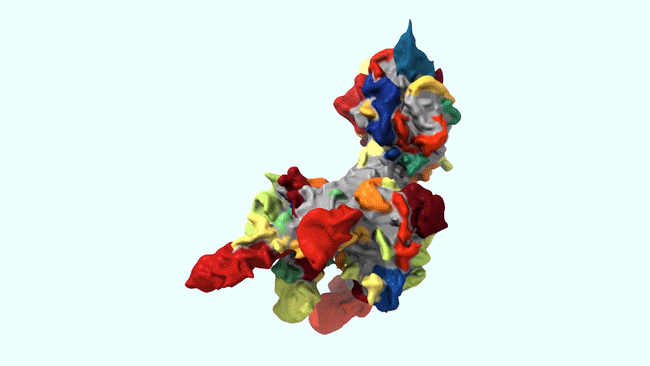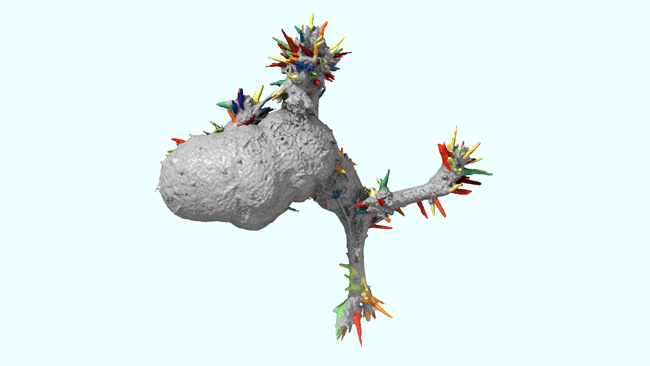
Ruixuan Gao, Shoh M Asano, Srigokul Upadhyayula, Igor Pisarev, Daniel E Milkie, Tsung-Li Liu, Ved Singh, Austin Graves, Grace H Huynh, Yongxin Zhao, John Bogovic, Jennifer Colonell, Carolyn M Ott, Christopher Zugates, Susan Tappan, Alfredo Rodriguez, Kishore R Mosaliganti, Sean G Megason, Jennifer Lippincott-Schwartz, Adam Hantman, Gerald M Rubin, Tom Kirchhausen, Stephan Saalfeld, Yoshinori Aso, Edward S Boyden, Eric Betzig
Content-Aware Image Restoration: Pushing the Limits of Fluorescence Microscopy
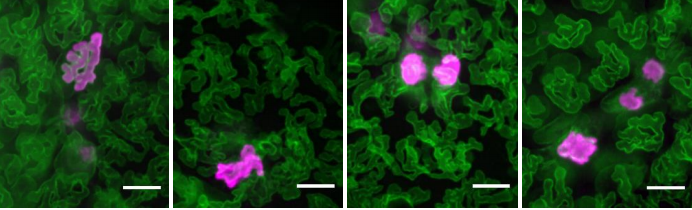
Martin Weigert, Uwe Schmidt, Tobias Boothe, Andreas Müller, Alexandr Dibrov, Akanksha Jain, Benjamin Wilhelm, Deborah Schmidt, Coleman Broaddus, Siân Culley, Maurício Rocha-Martins, Fabián Segovia-Miranda, Caren Norden, Ricardo Henriques, Marino Zerial, Michele Solimena, Jochen Rink, Pavel Tomancak, Loic Royer, Florian Jug, Eugene W. Myers
Unsupervised correction of gene-independent cell responses to CRISPR-Cas9 targeting
Francesco Iorio, Fiona M Behan, Emanuel Goncalves, Shriram Bhosle, Elisabeth Chen, Rebecca Shepherd, Charlotte Beaver, Rizwan Ansari, Rachel Pooley, Piers Wilkinson, Sarah Harper, Adam P Butler, Euan Stronach, Julio Saez-Rodriguez, Kosuke Yusa, Mathew J Garnett
Trafimow D, Amrhein V, Areshenkoff CN, Barrera-Causil C, Beh EJ, Bilgiç Y, Bono R, Bradley MT, Briggs WM, Cepeda-Freyre HA, Chaigneau SE, Ciocca DR, Carlos Correa J, Cousineau D, de Boer MR, Dhar SS, Dolgov I, Gómez-Benito J, Grendar M, Grice J, Guerrero-Gimenez ME, Gutiérrez A, Huedo-Medina TB, Jaffe K, Janyan A, Karimnezhad A, Korner-Nievergelt F, Kosugi K, Lachmair M, Ledesma R, Limongi R, Liuzza MT, Lombardo R, Marks M, Meinlschmidt G, Nalborczyk L, Nguyen HT, Ospina R, Perezgonzalez JD, Pfister R, Rahona JJ, Rodríguez-Medina DA, Romão X, Ruiz-Fernández S, Suarez I, Tegethoff M, Tejo M, van de Schoot R, Vankov I, Velasco-Forero S, Wang T, Yamada Y, Zoppino FC, Marmolejo-Ramos F
laure Bally-Cuif & Claude Desplan, conference organizers
Invited speakers:
Geneviève Almouzni Alexander Aulehla Allison Bardin Dominique Bergmann Florence Besse Sarah Bray Claire Chazaud Enrico Coen Michèle Crozatier Alain Goriely Thomas Gregor Laura Johnston Frank Jülicher Ryoichiro Kageyama Bill Keyes Thomas Lecuit Andrew Oates Patrick O’Farrell Ewa Paluch Nancy Papalopulu Catherine Rabouille Jody Rosenblatt François Schweisguth Benjamin Simons Claudio Stern Julien Vermot Jean-Paul Vincent Magdalena Zernicka-Goetz


 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)