A postdoctoral position is available in the laboratory of Dr. Sophie Astrof at Thomas Jefferson University to study roles of cell-extracellular matrix (ECM) interactions in cardiovascular development and congenital heart disease. We have recently discovered that progenitors within the second heart field (SHF) give rise to endothelial cells composing pharyngeal arch arteries. Projects in the lab focus on the role of ECM in regulating the development of SHF-derived progenitors into endothelial cells and their morphogenesis into blood vessels. The successful candidate will combine genetic manipulation, embryology, cell biology, and confocal imaging to study molecular mechanisms of micro-environmental sensing during vascular development.
Astrof laboratory is a part of a modern and well-equipped Center for Translational Medicine at Jefferson Medical College (http://www.jefferson.edu/university/research/researcher/researcher-faculty/astrof-laboratory.html) located in the heart of Philadelphia.
To apply, please send a letter of interest detailing your expertise, CV and names and contact information of three references to sophie.astrof@gmail.com
A postdoctoral position is available in the lab of Thorold Theunissen at Washington University School of Medicine in St. Louis, Missouri, USA (theunissenlab.wustl.edu). Our research program is dedicated to exploring the molecular regulation of pluripotent stem cells and their applications in regenerative medicine. We have developed methods for inducing and maintaining human embryonic stem cells in a naive pluripotent state that shares defining transcriptional and epigenetic properties with the preimplantation embryo (Theunissen et al., Cell Stem Cell, 2014; Theunissen et al., Cell Stem Cell, 2016; Theunissen and Jaenisch, Development, 2017). We are seeking a highly motivated postdoctoral fellow who wishes to join a multidisciplinary research team investigating fundamental problems in stem cell biology. Projects are available in the following areas: (1) dissecting the signaling pathways and epigenetic mechanisms governing distinct human stem cell states, and (2) establishing novel stem cell models of early human development and disease. Assays used include RNA-seq, ChIP-seq, mass spectrometry, genome editing and flow cytometry.
Applicants should have a PhD and demonstrated relevant research experience. Excellent communication skills and the ability to work in collaboration are essential. A strong background in molecular biology, human ESC/iPSC culture, and differentiation assays is preferred. Experience in signal transduction, biochemistry and bioinformatics would be very beneficial.
Consistently ranked among the top 10 US medical schools, Washington University School of Medicine offers a highly interactive and stimulating academic environment for scientists in training. The candidate’s research will benefit from the highly collaborative environment within the Department of Developmental Biology and Center of Regenerative Medicine. We are located in the heart of the Central West End, a vibrant St. Louis neighborhood adjacent to major cultural institutions and one of the country’s largest urban parks. We offer competitive salary and benefit packages and candidates are eligible to apply for a Rita Levi-Montalcini Postdoctoral Fellowship offered by the Center of Regenerative Medicine.
To apply for this position please submit a CV, a cover letter describing research interests, and contact information for two references who can comment on your research to t.theunissen@wustl.edu. Applications will be reviewed promptly until the position is filled. Washington University is an equal opportunity employer and complies with applicable EEO and affirmative action regulations.
Hear from rodent and non-mammalian model organism users.
Learn about the highlight notice and applying for an NC3Rs grant.
Give a flash presentation: three minutes to speak about your research area, the specific expertise you offer and expertise you are seeking in a collaborator.
The topic: Segmentation is one of the most common features in animals and yet one of the most mysterious. The team led by Guillaume Balavoine explores the cellular and genetic bases of segment formation in the marine annelid Platynereis.
Segment formation in the annelid involves a special category of multipotent stem cells: teloblasts or posterior stem cells.
Platynereis is a very convenient model for microscopy and live-imaging. It is amenable to a number of genetic interference techniques such as morpholinos, RNAi and CRISPR-Cas9 knock-outs. Stable transgenesis can be performed using DNA transposition. We want to explore the biology of the teloblasts by developing a number of transgenic tools in Platynereis. We are performing clonal analysis using transgene mosaic insertions. We have developed a FUCCI tool to live-image the cell cycle transitions in individual cells.
Armed with these tools, we want to answer a number of questions concerning the stem cells: observe and characterize their lineage restrictions, observe asymmetric divisions and decipher the link between cell cycle and segment formation. Promising techniques such as light sheet microscopy, cell sorting and RNASeq analysis can be performed at the Institute.
The location: The Institut Jacques-Monod, funded jointly by the CNRS and the University Paris Diderot, is one of the main centers for basic research in biology in the Paris area. It is located near the center of Paris. The Institute provides an outstanding environment for research at the highest level and assembles around 30 groups working on genome and chromosome dynamics, cellular dynamics and signaling, development and evolution. The Institut Jacques Monod has developed a number of important core facilities, which all offer state-of-the art instrumentation and a high level of expertise. These include a large microscopy imaging platform and a genomics-transcriptomics facility.
Requirements: Candidate must have obtained PhD by the start date. Candidate will usefully have experience in developmental biology, including micro-injection, confocal imaging and image analysis. The position will remain open until filled.
Contact: To apply, please send your CV with a cover letter stating your motivation and contact details of three persons for references, to :
We are looking for a highly skilled and motivated candidate to join our group for a PostDoc position. In the Payer lab (http://www.crg.eu/bernhard_payer), we study epigenetic reprogramming in the mammalian germ line and the effects of ageing on fertility. In this project the prospective candidate will study the molecular links between ageing and oocyte quality decline in women. The work will be performed in collaboration with the fertility clinic Eugin in Barcelona (https://tinyurl.com/y9l834y5).
We are seeking applicants with a strong background in the fields of Mammalian Cell Culture, Embryology, Stem Cell Reprogramming and Differentiation, Tissue Engineering, Epigenetics and Reproduction. Excellent candidates from other related fields will also be considered.
Work Environment:
Our lab is part of the Gene Regulation, Stem Cells & Cancer Programme at the Centre for Genomic Regulation (CRG) in Barcelona, Spain (www.crg.eu). The CRG is a vibrant International Research Institute with Research Groups working in diverse fields such as Genomics, Cell and Developmental Biology, Systems Biology, Stem Cells, Cancer and Epigenetics. English is the working language.
Fellowship + Application Guidelines:
36 months by the INTREPiD Fellowship programme. Application guidelines including eligibility criteria can be found at: http://www.crg.eu/intrepid_fellowships
The application deadline is the 12th of October 2018, at 5:00pm (local time).
For informal questions regarding the position, please contact: academicoffice@crg.eu
It’s undoubtedly the middle of summer here in Saint Augustine, Florida. Daily temperatures are soaring into the 90s, and we’re grateful if the humidity dips below 70%. Thankfully, the Seaver lab doesn’t have to contend with much of this heat. Instead, our members are inside, comfortable though busier than ever, mentoring summer interns, piloting new experiments, and writing up papers. It’s summertime, and the University of Florida’s Whitney Laboratory for Marine Bioscience is bustling with research as a whole!
I am Alexis Lanza, a PhD candidate in Dr. Elaine Seaver’s Lab with a research interest in embryonic signaling events critical for dorsal ventral axis formation in annelids. Here, I share a bit about our lab and its daily activities.
Dr. Seaver and graduate student Alexis Lanza imaging samples using a compound microscope
Area of Research
Broadly speaking, the research being done in the Seaver lab aims at answering questions about the development and evolution of marine invertebrates. We’re curious about the cellular and molecular mechanisms that drive developmental processes from an unfertilized egg through to zygotic, larval, and juvenile stages, as well as during regeneration. We’re also interested in how molecular differences that exist across species may account for body plan differences across animals. In our lab we use a small marine worm called Capitella teleta as our model organism.
Capitella teleta as an ideal model
Adult Capitella teleta.
Capitella teleta is a polychaete annelid worm and member of the
Lophotrochozoa. It is a cosmopolitan species and can be found along intertidal zones living in organically rich marine sediments. Capitella can easily be kept as part of a lab colony in glass finger bowls with seawater and roughly a tablespoon of mud. Following fertilization, its embryos develop into pelagic non-feeding larvae that metamorphose into juvenile worms approximately 9 days post-fertilization. At around 8 weeks old, these juveniles mature into adults that are easily distinguishable by sex and are capable of reproducing year-round. Capitella’s ability to produce embryos regularly is one reason why it is an ideal candidate for early developmental studies.
Furthermore, its embryos undergo a stereotypic cleavage program called unequal spiral cleavage in which blastomere formation occurs according to a predictable order, size, and position. This predictable cleavage program is also shared by other taxa including mollusks and nemerteans, and allows for the identification of individual blastomeres that can then be microinjected, or deleted using a laser (Meyer and Seaver, 2010; Yamaguchi et al., 2016).
Early cleavage stage embryos of Capitella teleta
Another prominent area of study within our lab focuses on how regeneration occurs in Capitella (de Jong and Seaver, 2017). Capitella, like many of its annelid counterparts, possesses regenerative capabilities following transverse amputations, as it is capable of regenerating posterior segments. However, Capitella cannot regenerate anteriorly. The development of Capitella as an annelid model for developmental and regeneration studies was rooted most significantly in our ability to conduct experimental manipulations, functional genomics, as well as having a sequenced, annotated, and slowly evolving genome (Simakov et al., 2012). Altogether, the Seaver lab utilizes tools of experimental embryology, molecular and cellular biology, and functional genomics to answer evo/devo questions.
A typical day of animal care
Maintaining a Capitella colony requires several activities including feeding. While these tasks aren’t difficult in themselves, every member of the Seaver lab chips in throughout the week, tackling a different task during breaks in experiments.
We keep our Capitella lab colony in glass finger bowls in a 19°C incubator
Sifting
Every glass finger bowl initially contains 40 individual worms. As the worms become reproductive however, we need to make an effort to avoid overpopulation in each bowl. When “sifting,” worms are removed from their bowl of mud and carefully sorted under a dissecting microscope. Unhealthy worms and larvae (the product of spontaneous matings) are removed, and only healthy individuals are placed into a fresh bowl of mud to be restocked as part of the working colony used for experiments.
Worms are sorted under a dissecting microscope and healthy individuals are placed into a fresh bowl of mud.
The Seaver lab divides its colony of bowls into two groups, each of which is sifted on an alternating, bi-weekly schedule. At the start of each week, a table listing all the colony bowls that need to be sifted that week is made.
Every week, lab members sign up to sift a couple colony bowls.
Lab members will then sign up for particular bowls based on their experimental needs are for the week. For instance, colony bowls with worms that have only recently reached sexual maturity are less likely to have larvae than would older bowls. Thus, if aiming to collect larvae one might want to sift an “older bowl.” Alternatively, if the goal is to conduct embryological experiments one may want to collect mature male and female worms, which can be used as a mating pair to produce early stage embryos.
Setting up the next generation
To maintain our lab colony through generations, it’s crucial that we plan ahead. As our worms age, they become less healthy and produce less offspring so are gradually phased out via the sifting process. To account for losing these individuals we introduce young, sexually mature worms (approx. 8 weeks old), all of which were raised from larval stages.
During the sifting process, lab members collect late stage larvae (approx. 8-9 days old), which are then used to create the new generation of colony bowls. First, glass fingerbowls are filled with seawater plus a teaspoon sized scoop of mud. Next, late stage larvae from several different mating pairs are pipetted into said fingerbowls until the total number of larvae equals 40. The mud is used to induce larvae to metamorphose. These new bowls will then be raised at 16°C for 8 weeks before being incorporated into the working colony.
Feeding
Animals need to be fed fresh mud once a week. We collect, sieve and freeze large quantities of marine mud that can then be thawed and used for animal care over the course of several months. To feed the colony, seawater is first decanted from each bowl. One tablespoon of mud is then added to each bowl, followed by the addition of fresh seawater. This process takes anywhere from 10 minutes to 1 hour depending on how large the colony is at any given time.
Fingers crossed for embryos
Mating Bowls
Remember those mature male and female worms collected during the sifting process? To acquire early cleavage stage embryos, mature male and female worms are separated by sex into bowls with 4 or 5 individuals each for about 3 days. When ready to set up a mating we simply combine the individuals from a male bowl with those from a female bowl. In general, we’ve noticed that it takes about 8-10 hours for the worms to mate and the females to lay their broods. Since most of our embryology experiments begin in the morning we need to set up these matings around midnight… this often means taking your worms home with you and only giving vague answers to questions your housemates may ask.
In the morning, back at the lab, we sift our mating bowls with bated breath to check if any of the female worms laid a brood of fertilized embryos.
When female worms are ready to lay their eggs, they create an encasement called a brood tube. The worm remains inside the brood tube along with her offspring until they become larval stages.
Using our dissecting microscopes and a pair of forceps, we dissect open any brood tubes and gently pipette the embryos into a clean petri dish. From here we can stage the embryos and determine how many cleavage divisions into development they are. Capitella embryos are approximately 200 microns in size and each cleavage division takes approximately 45 minutes. These embryos are amenable to chemical perturbants, allowing us to manipulate various signaling pathways and determine its role in development (Lanza and Seaver, 2018). Furthermore, Capitella embryos can be microinjected. To do so, the egg membrane of the embryo is first softened using a sucrose: sodium citrate mixture. This treatment is typically performed one cleavage division before the desired cell stage you wish to inject. We can then microinject individual cells during early cleavage stages with aqueous or lipophilic lineage tracers, DNA, RNA, or morpholinos, and then raise these embryos in seawater to larval stages (Meyer et al., 2010; Seaver, 2016). Like in many other species, most of the difficulty about working with early cleavage stage embryos is timing related. Since we can’t predict exactly when the eggs are fertilized we often end up racing against the clock to get our experiments underway before our target cleavage cycle ends or wait for hours until our embryos finally make it to the target cell stage. In general, after experimental manipulation, embryos are raised for six days to larval stage before being fixed for phenotypic analysis.
By the day’s end, the afternoon showers have usually subsided and the temperature has waned. It’s around then that the members of the Seaver lab emerge – just in time to enjoy the last few hours of sunlight during another Florida summer.
Members of the Seaver lab Spring 2018. Left to right, Linlin Zhang, Elaine Seaver, Alexis Lanza, Stephanie Neal.
The School of Natural Sciences invites applications for three full-time, permanent Lectureships in the broad area of Biology encompassing the full spectrum of genes, organisms and ecosystems. As part of the School of Natural Sciences, the biology and zoology subject areas are dynamic and growing elements with strong research records and successful recruitment onto our undergraduate and postgraduate degrees. Currently we are looking to expand our research base and our teaching and outreach capabilities. Bangor University is committed to excellence in research, teaching & scholarship and offers attractive career prospects for staff with such expertise.
Details of current staff research interests are available online. The School benefits from modern research laboratories and microscopy suites, a Natural History Museum, freshwater and marine aquaria, fly room, reptile and rodent facilities, plus a number of temperature controlled rooms, a botanical garden located on the fringes of the Menai Strait, a herbarium, and avian housing facility. Further research opportunities are facilitated by close links to the Centre for Ecology and Hydrology (CEH), the Centre for Environmental Biotechnology (CEB), the Schools of Ocean Sciences, Computer Sciences, and Psychology and access to the Supercomputing Wales High Performance Computing cluster.
Candidates should have a PhD in biological science, be able to develop research programmes of their own, contribute to the School’s Research Excellence Framework (REF) submission, show an enthusiasm for teaching, be willing to engage with outreach activities and also have excellent interpersonal skills.
Interviews will be held in the week commencing 3rd September. The successful candidates will be expected to start on 1st October 2018 or as soon as possible thereafter.
Informal enquiries may be made to Professor Chris Freeman, email c.freeman@bangor.ac.uk; and copied to Dr Nia Whiteley, email n.m.whiteley@bangor.ac.uk.
The transience of flowers is proverbial. Degeneration of flowers is elicited after successful pollination by the onset of seed and fruit development. However, also unpollinated flowers do not last forever – on the contrary, the life span of unpollinated flowers is a tightly regulated trait that differs greatly among plant species. Some plant species like orchids of the genus Phalenopsis have very long-lived flowers that can stay receptive for weeks, even months. Most plant species, however, sustain unpollinated flowers for much shorter time spans. In the most extreme cases mere hours pass between flower opening and withering.
In the Programmed Cell Death lab at the VIB Center for Plant Systems Biologywe are using the model plant Arabidopsis thaliana to study the longevity and senescence of unpollinated flowers. Although Arabidopsis is the most advanced model species for plant developmental biology, it is actually not particularly convenient to investigate the biology of flower senescence. Arabidopsis flowers are tiny, and moreover they are self-pollinated, so that the male floral parts (anthers) have to be either removed manually by emasculation, or male-sterile mutants have to be used.
A typical Arabidopsis flower aligned next to a pin. The floral stigma is at the level of the pointed end of the needle.
Every Arabidopsis researcher knows how to perform emasculation and pollination in order to cross different mutants, marker lines, or accessions. And many of us know that a flower emasculated on Friday might already have started to senesce by Monday, hence making successful pollination and seed set inefficient or impossible.
In order to observe and stage flower withering in Arabidopsis, we set up a macroscopic imaging system using a single lens reflex camera equipped with a macro objective. Taking images every 10 minutes under constant light enabled us to monitor flower senescence in detail. We could observe that concomitant with the withering of petals and sepals, also the floral stigma was degenerating.
The stigma of flowers is the primary receptive surface for pollen grains. In Arabidopsis the stigma consists of over 200 elongated fingerlike cells, the so-called stigmatic papillae, specialized epidermal cells that serve to intercept the pollen. In Arabidopsis, hundreds of pollen grains compete to fertilize about 50 ovules located inside the pistil of each flower. Once in contact with the stigma cells, pollen grains will hydrate, and germinate to form a single tip growing cell, the pollen tube. The pollen tube penetrates the papilla cell wall, growing in between the cell wall and the plasma membrane to the base of the papilla cell. From there on, guided by chemical cues, the pollen tube grows through the style and the transmitting tract to the ovules. There, it will release its two sperm cells to fertilize the egg cell and the central cell of the female gametophyte, thus initiating embryo and seed development.
The journey of the pollen tube during plant reproduction in Arabidopsis. Pollen grains (orange) are shed from the anthers and adhere on the stigmatic papilla cells. During the preovular guidance phase, pollen grains hydrate and germinate, and pollen tubes (red) penetrate in the papilla cells and grow through the style and transmitting tract. After that, pollen tubes are attracted by the ovules, enter the ovules with a near one-to-one relationship during the ovular guidance phase. Finally, pollen tube growth stops, and the sperm cells are released into the female gametophyte to perform double fertilization. Picture is a courtesy of Zhen Gao.
The structure of the intact Arabidopsis stigma appears a bit like a little hedgehog due to the hundreds of erect elongated papilla cells. Once flower senescence sets in, individual papilla cells start to break down, leading first to a ragged, and finally to a completely collapsed appearance of the stigma.
Pollination assays at different time points during this process showed that stigma degeneration coincided with a sharp decline in seed set, suggesting that viable papilla cells are necessary for successful reproduction. To test this hypothesis, we specifically ablated papilla cells by expressing diphtheria toxin chain A (DT-A) under a stigma-specific promoter. As we had assumed, DT-A induced stigma degeneration likewise caused a strong reduction of seed set.
In order to quantify stigma longevity, we faced a considerable problem – the macro setup was only able to follow a single flower at a time, and a price tag of about 1400 Euro per unit made the parallel acquisition of time courses impracticable. Coincidence came to our aid: Setting up a skype conference call we noticed that a cheap webcam without autofocus can be brought very close to an object (in that particular case a certain PhD student’s uvula) and amazing magnification can be achieved. After some experimentation with different webcams, we rigged up a phenotyping system with 20 webcams. We used a custom-made script to operate the open source camera surveillance software so that each camera acquires one picture frame every 10 minutes. Although the image quality was of course not comparable to the conventional macro lens setup, in the end of the day we had established a phenotyping platform with 20 imaging units for mere 250 Euros.
Webcam imaging platform to follow flower senescence in Arabidopsis
Analysis of the time lapse images revealed a remarkable reproducibility of stigma collapse on average around 56 hours after emasculation of a flower at the developmental stage 12c1.
The apparently precisely timed collapse of papilla cells suggested that an active cell death process might be occurring in Arabidopsis stigmata. Based on a large-scale meta-analysis of mRNA transcriptome profiles, our lab has established promoter-reporter lines of genes transcriptionally upregulated prior to a number of developmentally regulated programmed cell death (PCD) events2. Microscopic analyses revealed that most promoter-reporters come up during stigma senescence, thus linking stigma degeneration with established forms of developmentally controlled PCD processes3. Microscopic imaging of papilla cells is not trivial; although the individual cells are thankful objects for cell biological analyses, the stigma as a whole is a rather large and very sensitive three-dimensional structure. In order to investigate cell biological hallmarks of PCD in living stigmata, we could not use conventional slide-and-coverslip setups lest we create injuries and mounting artefacts in papilla cells. After testing diverse setups, we found that mounting the entire flower in an agar block and viewing the stigma from top with a long working-distance water dipping lens on an upright Zeiss 710 confocal microscope allowed a minimally invasive way to perform live-cell imaging of papilla cell death. In order to visualize cell death, we used a live/death stain in which fluorescein-diacetate (FDA) fluorescence indicates living cells, while nuclear staining of the membrane impermeable stain propidium iodide (PI) reveals plasma-membrane permeation as a committing step of cell death.
Cell death/ viability staining of stigmatic papilla cells. Fluorescein-diacetate (FDA) fluorescence indicates living cells, while nuclear staining of the membrane impermeable stain propidium iodide (PI) reveals plasma-membrane permeation as a committing step of cell death. Picture is a courtesy of Yulia Salanenka.
Alternatively, we used a tool developed in our lab, a tonoplast integrity marker (ToIM) line4. This plant line expresses a cytoplasmic green fluorescent protein (cGFP) and a polycistronically produced vacuolar red fluorescent protein (vRFP). Living cells display clearly separated GFP and RFP domains indicating an intact vacuolar membrane (tonoplast). Collapse of the large central vacuole in mature plant cells, another hallmark of plant programmed cell death5, is visible as merging of red and green fluorescence signals.
A confocal maximum intensity projection of a representative stigma from a Tonoplast Integrity Marker line (ToIM). Green labels cytosolic GFP, red shows RFP localized to the vacuole. The fusion of both signals appears yellow and indicates a loss of cellular compartmentalization symptomatic of cell death.
Our analyses revealed that as in the macroscopic setup, stigma degeneration occurred over a time span of 12 hours. Fascinatingly, the death of individual papilla cells followed a pattern from the periphery towards the center of the stigma, and often small clusters of individual cells underwent vacuolar collapse almost synchronously. On the level of individual cells, the PCD program occurred surprisingly fast; vacuolar collapse, plasma membrane permeation, nuclear fragmentation, and finally cellular collapse occurred within about one hour. The reproducibility of these successive events confirmed our hypothesis of a tightly regulated cell death program.
In order to discover regulators of this cell death process, we set out to perform an RNA-seq analysis monitoring the transcriptional changes occurring in the stigma over time. We manually dissected the stigmata from flowers at 1, 2, and 3 days after emasculation, corresponding to young, mature, and senescent flower stages. In total we had to stage, emasculate, and dissect way over individual 2000 flowers, a feat that we only managed with lots of training and a great team effort.
Emasculation session in the PCD lab. The combined efforts of at least three people were required during the laborious process of Arabidopsis flower emasculation. The stigmata of about 2000 emasculated flowers was collected and subjected to RNA-Seq analysis.
Illumina RNA-seq revealed a large number of differentiallty regulated genes (1180 genes out of more than 25 000 predicted genes in Arabidopsis) over the course of stigma senescence. Interestingly far more genes were upregulated (897) than downregulated (283), again indicating an actively controlled program controlling stigma senescence and papilla cell death.
RNA sequencing also confirmed the strong upregulation of developmental PCD-associated genes. In order to identify key regulators of this process, we investigated the expression profiles of transcription factors (TFs). Interestingly, the plant specific NAC TF family was strongly overrepresented in senescence-associated TFs. Among the most strongly upregulated NAC TFs was the well-established leaf senescence regulator ORESARA1/ANAC092 (ORE1, Korean for “long-living”), and the flower-specific ANAC074, a previously uncharacterized NAC TF that we dubbed KIRA1 (KIR1) after the killer “Kira” in the Japanese manga “Death Note”.
Using our webcam-phenotyping platform, we investigated an established knock-out mutant of ORE1 and found a slight, but not significant delay of stigma collapse. Interestingly, a newly established KIR1 knock-out mutant showed modest, but significant extension of stigma life span. We crossed the mutants and the resulting kir1 ore1 double mutant showed a clear synergistic effect which led to a doubled life span in comparison to the wild type. We also generated lines overexpressing dominant repressive mutant versions of KIR1 and ORE1, and some of these lines had an even stronger effect with some stigmata only collapsing at 11 days after emasculation. Intriguingly, both recessive kir1 ore1 loss-of-function mutations, as well as expression of dominant repressive alleles showed an uncoupling of stigma life span from floral organ longevity. Although the stigma life span was considerably increased, sepals and petals senesced similar to the wild type.
In a complementary approach, we investigated KIR1 gain-of-function mutants. Estradiol-inducible systemic misexpression of KIR1 in seedlings caused a rapid growth arrest caused by a widespread ectopic cell death. This demonstrates that the transcriptional program controlled by KIR1 is sufficient to elicit cell death outside of the stigma context. When inducing precipitate KIR1 expression specifically in the stigma, the papilla cell death and loss of receptivity occurred significantly earlier than in estradiol treated wild types or mock treated mutant lines. These results demonstrated that KIR1 functions to actively terminate the receptive life span of the flower by promoting a cell death program in the stigma.
This picture shows an emasculated Arabidopsis flower undergoing localized estradiol treatment in order to induce KIR1-GFP overexpression in the stigma. The floral stigma is imbibed into an estradiol-containing droplet for 6 hours and pollinated after that.
A central question arising from this investigation was of course whether aged but viable kir1 ore1 loss-of-function mutants could be successfully pollinated. When performing a pollination time series, we discovered that in dominant repressive KIR1 and ORE1 mutants, there was as significant, but rather modest extension of flower receptivity. In the recessive kir1 ore1 double mutant, there was no extension of floral receptivity at all. We used pollen from a transgenic Arabidopsis line that expresses β-glucuronidase (GUS) under a pollen-tube specific promoter to visualize pollen tube growth on pollinated stigmata after addition of the GUS substrate X-Gluc6. Microscopic analyses showed that pollen hydrated and germinated, but that pollen tube growth on aged mutant stigmata was strongly reduced in comparison to younger wild type or mutant stigmata. No pollen tubes could be detected to enter the style or the transmitting tract of the aged mutant flowers, suggesting that KIR1 / ORE1 loss of function is sufficient to suppress age-induced stigma PCD, but not sufficient to maintain stigma function in a corresponding fashion.
In summary, our research on stigma senescence suggests that a KIR1-ORE1 dependent cell death program actively terminates stigma and flower receptivity. However, as suppression of this cell death program is not sufficient to extend stigma function, we assume there must be additional, KIR1 – ORE1 independent pathways that either passively or actively terminate stigma function in the absence of cell death. With research going on in our lab we attempt to research these pathways in order to effectively modulate flower receptivity.
While Arabidopsis serves as a model system to discover these pathways, our research might also be applicable to outcrossing crop species. An extension of floral receptivity, especially under spells of environmental stresses, which are deleterious to plant reproduction, might be a key strategy to stabilize the yield of seed and fruit bearing crops7.
I recently attended the biennial Development, Cell Biology and Gene Expression C. elegans Meeting, this time in combination with the 2018 European Worm Meeting, in Barcelona. C. elegans meetings are always pretty special, defined more than anything else by the strong sense of community between researchers, or, as well like to call ourselves, ‘Worm People’. Worm culture has extended from a magazine (The Worm Breeder’s Gazette) to comedy shows (The Worm Show), and worm-inspired art collections (The Worm Art Show) curated by Ahna Skop, designer of this year’s meeting logo. This shared love of the worm translates into an atmosphere of supportiveness and openness within the community, making worm meetings ideal places to share new data and ideas – and at the same time being a huge amount of fun.
The meeting logo was designed by Ahna Skop to represent the extensive work that has been carried out investigating the neurobiology of the worm. C. elegans remains the only animal to have its entire connectome mapped.
A person who exemplified the ethos of the worm research community, and the memory of whom permeated the meeting, was John Sulston, who sadly passed away earlier this year. John Sulston was one of the first scientists working on C. elegans with Sydney Brenner at the LMB in Cambridge during the ’70s. He is known most famously within the worm field as the person who gave us the complete C. elegans cell lineage, a fundamental cornerstone of C. elegans research. This lineage represents years of work, as he monitored in real time every cell division of the larval – and following this, embryonic – worm, in order to build a remarkably accurate map of wild-type development from zygote to the adult. It is only very recently (40 years later) that any additional cell divisions have been identified. The concept of lineage is so fundamental to the ‘worm world’ that lab members also identify themselves according to their ‘lineage’ – tracing themselves back to that first ‘zygotic’ lab in Cambridge through a network of supervisors – in fact, many of the conference speakers introduced themselves in this way at the meeting.
However, John Sulston’s scientific legacy extends beyond the relatively small sphere of C. elegans biologists, as he was also the scientist who lead the British side of the Human Genome Sequencing Project. A first consequence of this was that the C. elegans genome was actually completed first, cementing it’s place in history as the first sequenced genome of a multicellular organism (John was apparently just as excited – if not more so – to resolve the worm genome as to complete the human genome). But more importantly, John Sulston was also responsible for fighting to make the human genome sequence freely accessible to all, not patented and the property of a company. The contributions that John Sulston made to the scientific community during his time with the Human Genome Sequencing project, and how his work with C. elegans shaped his scientific ideology, can be read in a book called The Common Thread, which he wrote with the science writer Georgina Ferry. His attitude towards open-access science and the free sharing of data and resources remains a defining property of the C. elegans field.
The meeting: attendees and talks
The meeting, held at the World Trade Centre on the waterfront in Barcelona and organised by Sander van den Heuval, Sophie Jarriault and Alex Hajnal, opened to an audience of 465 people, representing research from 33 different countries. As well as the talks, the meeting also involved two lively poster sessions, and a fabulous evening of food, wine and dancing by the sea – and where better to party than in Spain? I certainly came away from the meeting with more worm-sympathetic friends than I’d started with.
As there were unfortunately too many presentations to talk about them all here, I have summarised a selection of my favourites. However, a complete list of the abstracts can be found here.
Meeting attendees, representing 33 countries.
The World Trade Centre (arrow) in Barcelona, where the meeting was held. Photo by the author.
Worm behaviour and neurogenesis
When Sydney Brenner first selected Caenorhabditis for research, his ultimate goal was to solve the genetics and cell biology of behaviour. To this end, the electron microscopist Nichol Thompson created serial sections (up to 5000, uninterrupted) of the worm in order to map each synapse, and subsequently a draft of ‘the Mind of a Worm’ was published in 1986. Given that the worm is the only organism to date to have it’s entire connectome mapped, the field of neurobiology – and more recently behaviour – is an intensely studied area of C. elegans research.
Upon completion of the C. elegans post-embryonic lineage, it was established that the adult hermaphrodite worm had 302 neurones, and the male 385. However, as I have already mentioned, the first post-Sulston cells to be identified added two neurones to the male count, discovered by Arantza Barrios of UCL, London, and published in 2015. At this meeting, Arantza Barrios revealed how her lab, together with the lab of Richard Poole, also at UCL, have since gone on to update the number of male neurones yet again (to 389) although this time the cells in question were always known to be there. However, they have now been identified as undergoing a fate and morphology change during larval development, producing a pair of putative proprioceptive neurones, the PHDs, from a pair of glial cells. The Barrios and Poole labs, with the help of Scott Emmons and Steven Cook at the Albert Einstein College of Medicine in New York, who through electron microscopy established the connectivity of the newly discovered neurones, have identified that the PHDs are essential for efficient male mating behaviour. As Arantza described, males are usually lead by the tail (in which their hermphrodite-sensing organ and spicule for sperm-delivery are located) during mating. In contrast to this known mating behaviour, a newly-described behaviour (termed the Molina Manoeuvre after the post-doc who discovered it) involves an unusual forward sweep along the hermaphrodite followed by a reversal in direction in order to relocate the vulva. This manoeuvre is only performed in the case of the male being unsuccessful in its first attempt to mate. Without the PHDs, the Molina Manoeuvre cannot take place successfully and the male is likely to lose its potential mate.
Wiring diagram of the newly-discovered PHD neurones, shown to be involved in male mating behaviour. L. Molina-Garcia et. al. “A direct glia-to-neuron natural transdifferentiation ensures nimble sensory-motor coordination of male mating behaviour” BioRxiv https://doi.org/10.1101/285320
Many mutant animals, or wild-type animals subjected to different environments, exhibit changed behavioural patterns. Systematic quantification of this behaviour can allow both detection of the behavioural differences and analysis of the specific change. Andre Brown of Imperial College London spoke about how his lab have broken down the 2-dimensional behaviour of C. elegans in a petri dish into a series of distinct positions, and uses the analogy of language to compare these positions to letters in an alphabet. He showed, using automated imaging of free-moving worm populations followed by extraction of relevant features, how wild-type worms are more likely to use a certain subset of ‘letters’ compared to mutant animals. His newly developed algorithms and imaging systems are likely to be useful for a wide range of applications, from behavioural genetics to agrochemical analysis.
Oliver Hobert from Columbia University in New York spoke about his lab’s recent feat of creating a comprehensive neurotransmitter map which overlays the anatomical map of the worm connectome, completed for both hermaphrodites and males. Most neurones in the worm fall into one of the main neurotransmitter classes: glutamatergic, GABAergic, or cholinergic. Upon completion of the neurotransmitter map, the Hobert lab has been able to identify 86 different classes of neurone within these classes, and through mining of the Wormbase database of gene expression, further show how a group of only 33 different transcription factors are able control these identities, acting as ‘terminal selectors’. (Remarkably, Oliver points out, the Wormbase collection of C. elegans gene expression patterns allows between 5 and 141 cell markers to be identified for each neurone – that’s an average of 32 markers per cell.) The Hobert lab deduced that there must, therefore, be a combinational code for neurone identity, and Oliver described experiments where the loss of one of a pair of transcription factors can have varying consequences on the identity of differing neurones. A second observation was that these terminal selector transcription factors are predominantly homeobox genes (c.f. only 10% of all transcription factors in the worm are homeobox genes). The lab went on to develop a worm strain that could be used to identify gene expression in any neurone based on co-expression (NeuroPal – Neuronal Polychromatic Atlas of Landmarks) and characterised gene expression of all homeobox genes in all neurones. They found that every neurone expresses at least one homeobox gene, with 113 out of 118 neurone classes expressing a unique homeobox gene code. This means the same homeobox gene gets used again and again to specify different neurones. A striking observation was made when looking for commonalities between the neurones that express the same transcription factor: they are all synaptically connected. This has lead to speculation in the lab of how evolution may first have resulted in individual networks of neurones, each based on the presence of a founding homeobox gene specifying a single neurone that subsequently duplicated, before acquiring additional specialisation across the network to encode multiple neuronal types. These networks, Oliver proposes, may have later linked up to form the connectome as we know it.
Tissue-specific expression analyses
Tissue-specific RNA-seq in the worm has provided a challenge for the field, especially for those interested in understanding the factors that drive differentiation. Current analyses include using known gene expression profiles coupled with machine learning algorithms to sort sequencing profiles obtained from FACS-sorted single cells back to their tissue of origin, such as the efforts of John Murray’s laboratory at the University of Pennsylvania. John presented his lab’s work on dissecting the cell-specific embryonic expression profile over time, using a ‘whole-organism shotgun’ approach. This approach involves using the RNA-seq data obtained from individual, separated cells from mixed stage embryos. He then profiles each cell in order to map it back to the cell lineage, using known gene expression ‘hallmarks’ obtained from expression patterns of individual genes. Using this data, it is possible to start to answer questions such as which genes are required for acquiring cell identity versus those that are required for maintaining cell identity. An interactive database of this data is under development and should be available soon (keep an eye on BioRxiv). In contrast, Annabel Ebbing, from Hendrik Korswagen’s lab at the Hubrecht Institute in the Netherlands, described how RNA tomography can contribute to our understanding of spatially-resolved gene expression patterns without any prior knowledge of gene expression – rather, this approach relies on the highly invariable anatomy of the worm. RNA tomography involves cryo-sectioning young adult animals at 20µm intervals along the anteroposterior axis, before using CEL-seq to sequence each section. The data were aligned to provide a comprehensive map of gene expression along the anteroposterior axis of the worm, at single cell resolution. This map has already revealed sex-specific differences in the expression of previously-unidentified genes required for male fertility in the male reproductive tract, and has been made available here.
Outline of the RNA tomography method. Worms are sectioned at 20µm intervals before RNA from individual sections is sequenced. The relatively simple anatomy of the worm allows for single-cell and tissue resolution of sequencing data using this section width. Ebbing, A.*, Vertesy, A.*, Betist, M., Spanjaard, B., Junker, J.P., Berezikov, E., van Oudenaarden, A., and Korswagen, H.C. Spatially-resolved transcriptomics of C. elegans males and hermaphrodites identifies novel fertility genes. Submitted.
Cell fate and plasticity
The understanding of cell fate determination, and its opposite, fate plasticity, is important due to the potential implications of reprogramming in cancers and regenerative medicine. C. elegans provides two models with which to study in vivo reprogramming: cells which undergo natural transdifferentiation, and induced conversion of cells by ectopically expressed transcription factors.
The most famous natural transdifferentiation event in the worm is the Y to PDA transition in the tail. Y is a fully differentiated rectal epithelial cell that, partway through larval development, detaches from the rectum, migrates and converts into the PDA neurone. This process was first detected by John White and noted in his lineage, and later confirmed to be a bona fide transdifferentiation event by Sophie Jarriault. Sophie’s lab have continued to contribute to the field of reprogramming, and more recently her work has expanded to encompass other natural fate changes in C. elegans. Claudia Riva from the Jarriault lab talked about her work on these other cells. Although direct conversion of a cell from one fate to another is rare (the two known examples are the changing fate of Y, and also the phasmid socket PHso1, which produces the PHD neurone required for the Molina Manoeuvre), there are also cases of seemingly differentiated cells dividing to produce a cell of a different fate. These include the K cell in the rectum, which divides to produce the DVB neurone, and the G1 excretory pore cell, which gives rise to two RMH neurones. Claudia showed that even though these cells undergo a cell division, the ‘plasticity cassette’ comprising SEM-4, EGL-27, SOX-2 and CEH-6, identified to be required for the Y to PDA transition, is also required for the production of DVB from K. For example, in sem-4 mutant animals K goes on to divide but K.p remains epithelial. However, upon halting the cell cycle using a temperature sensitive allele of lin-5, she demonstrated that cell division is also essential for the production of DVB. The asymmetric division of K requires Wnt signalling, like many other asymmetric pathways in C. elegans, leading the lab to investigate the interplay of Wnt (during asymmetric cell division) and plasticity reprogramming factors (for the acquisition of neuronal cell fate) in the process of K dividing to produce the DVB.
Anna Reid, who works in the Tursun lab at the Berlin Institute for Medical Systems Biology in Germany, talked about her work on the forced reprogramming of mesodermal coelomocytes. There are three pairs of coelomocytes located in the pseudocoelom of the worm. The lab has found that the ectopic expression of the GATA factor ELT-2 is sufficient to convert these cells to the intestinal fate, and upon closer inspection found that they even produced an intracellular lumen lined with microvilli. Testing the limits of forced reprogramming, she found that the zinc transcription factor CHE-1, which specifies the fate of the ASE neurones, was also found to be able to reprogram the coelomocytes, with a 70% success rate for one of the three pairs of cells. Intriguingly, since the coelomocyte fate is lost at the time of conversion, so too is the expression of the coelomocyte-specific transgene inducing the forced transition event (in this case, the unc-22 promoter is used to drive ectopic che-1 expression in the coelomocytes). Thus only a pulse of expression must be sufficient to convert fate. Anna finds that this pulse of ectopic CHE-1 expression leads to the endogenous che-1 locus to be switched on in these cells; the fate-converting experiment in a che-1 mutant background is much less efficient. Lastly, Anna explained how the coelomocytes, now shown to be amenable to either intestinal or neuronal reprogramming, are much more resistant to a muscle fate change. Anna suggests that this is surprising, since the muscle lineage is much closer to that of the coelomocytes than the neuronal lineages. This raises the question of how lineage proximity can affect fate plasticity: if two lineages are closer, does this make it easier or harder to reprogram between them?
Transcriptional metabolic network rewiring
Marian Walhout, from the University of Massachusetts Medical School, USA, delivered a very elegant talk on the work her lab are carrying out in the field of systems biology. She explained how the worm has two metabolic pathways for breaking down proprionate, one of which is dependent on vitamin B12, and the other not. The B12-dependent pathway is preferable to the worm as it does not risk the build up of toxic intermediates, but the pathways are interchangeable in the event of no B12 being available and subsequent build up of proprionate. (Marian also points out that the standard laboratory culture conditions of growing C. elegans on the OP50 strain of E. coli means that the worms are always B12-deficient in the lab.) To convert from a B12-dependent pathway to a B12-independent pathway requires the transcriptional machinery to switch to expressing the enzymes required for the new pathway. However, in order to achieve this only when there is a sustained deficiency of B12, and not just in the case of a temporary peak in proprionate, requires a delay to be built into the switch. Marian explains how this delay has been modelled in bacterial systems by a coherent type 1 feed-forward loop with an AND-logic gate, providing persistence detection in response to a stimulus. However, this type of transcriptional circuitry has never been observed in an animal system, until now. To turn on the B12-independent pathway, Marian’s lab have found that two genes, nhr-10 and nhr-68, together act in such a persistence switch to turn on five genes of the B12-independent metabolic pathway, and in this way prevent this potentially hazardous pathway being turned on during a transient peak in proprionate.
Model for a persistence detector in the form of a coherent type 1 feed-forward loop with an AND-logic gate. Transcription factors A and B are both required to turn on their targets. In the case of a transient stimulus, however, gene A will not be expressed long enough to make sufficient amounts of gene B, in order to turn on the switch. Yet if the stimulus is sustained, transcription factor A will activate the expression of gene B, and the target genes will be transcribed.
Meiosis
Abby Dernburg from the University of California, Berkeley shared her work on how crossovers are regulated during meiosis. In C. elegans, each pair of homologous chromosomes undergoes just one crossover event, and all other double-stranded breaks are repaired without the transfer of genetic information. Little is known about how chromosomes convey this crossover information along their lengths. Through fluorescent analysis, allowing direct visualisation of the synaptonemal complex assembly which forms between homologous chromosomes, the Dernburg lab have been able to quantify the kinetics of synapsis. This has lead to the conclusion that the synaptonemal complex in fact acts as a liquid crystal – an ordered assembly of molecules that show liquid-like mobility – rather than a static complex. This rapid diffusion within the complex could be the mechanism by which information is rapidly transported down the length of the homologous chromosomes. The lab is now testing how each of the previously-identified factors for regulating crossover events, including the RING finger proteins ZHP-1-4, CDK-2 and COSA-1, can act with the synaptonemal complex to regulate and limit the number of crossovers to one per chromosome pair.
Membranes in biology
David Sherwood of Duke University, USA, delivered a wonderful talk on his work studying basement membranes in C. elegans. I had first come across David’s work as a Masters student, reading about how the anchor cell invades through the basement membrane that separates the gonad from the uterine seam during vulval formation. During his talk, David described how his lab have created a toolkit of fluorescently labelled endogenous basement membrane components (17 components and 13 receptors) in order to study the composition and dynamics of this ancient extracellular matrix. One of the striking results from this work has been the discovery of the huge range in the rates of turnover and mobility between these tagged components: FRAP and FLIP experiments show that whereas some components take hours to recover, others recover in a matter of milliseconds.
Meera Sundaram from the University of Pennsylvania, USA, is more interested in the apical membranes of cells – during this meeting she shared some of the work from her lab on how the worm builds tiny, seamless tubes from a single cell. There are three possible ways for a single cell to form a tube: through endocytosis and internal vesicle fusion, through membrane invagination followed by exocytosis, or through auto-fusion after the cell has rolled around the lumen. In the excretory system of C. elegans, there are two cells which each form a tube. These are the duct cell, which forms a seamless tube, and the pore cell, which remains seamed. Both tubes are formed by an initial rolling of the cell, but the duct cell then auto-fuses in an EGF-Ras-ERK dependent manner. When looking for targets of this pathway, Meera’s lab discovered that this fusion is dependent on the activation of the fusogen aff-1, which has already been shown to drive fusion in the uterine seam and between seam cells. Auto-fusion of the duct cell occurs early in development, and is followed by morphogenesis and elongation of the tubular cell into its final shape. Using the degron system to deplete aff-1 after the fusion event but before remodelling, Meera showed that surprisingly, aff-1 was also required for the elongation of the duct cell. In the absence of aff-1, she showed using STED microscopy that the cell accumulates vesicles containing apical cargoes, and also had nucleus-sized ‘hair-balls’ of membrane attached to the basement membrane, suggesting that AFF-1 is required for apically-directed trafficking. AFF-1 is present at endocytosing membranes, so may be required for scission. Vesicle trafficking is known to be required for remodelling of the duct cell, as Rab11, required for the recycling of endocytic vesicles, is essential for duct cell elongation. Thus Meera’s lab has discovered a novel role for fusogens during vesicular trafficking.
Model of the role of AFF-1 during endocytic scission in the duct cell of the excretory system.Soulavie F, Hall DH, Sundaram MV. The AFF-1 exoplasmic fusogen is required for endocytic scission and seamless tube elongation. Nature Communications. 2018;9:1741. doi:10.1038/s41467-018-04091-1
This was not the only talk at the meeting that presented a novel role for fusogens, however. Piya Ghose from the Shaham laboratory in the Rockefeller University, New York presented her work on how the fusogen EFF-1 was required for phagosome sealing during the programmed cell death of the tail spike cell. The role of fusogens in cell biology appears to be broader than was previously thought.
The ageing worm
The striking genetic control of worm lifespan has been demonstrated many times over the last few decades, and this together with the worm’s relatively short natural lifespan makes it an attractive model for ageing research. Adam Antebi of the Max Plank Institute for Biology of Ageing in Cologne, Germany, presented his lab’s work on how nucleolar size in the young worm can be used as a predictor of the lifespan of that worm – with worms with smaller nucleoli living the longest. Not only is this trend true for worms, but he showed how it also holds true for fruit flies, mice and humans, suggesting that the pathways of ageing that converge on nucleolar size are conserved. Adam’s lab found that nucleolar size is controlled by the NCL-1 tumour suppressor, with long-lived animals representing different longevity pathways exhibiting small nucleoli, decreased rRNA and ribosomal protein expression, and decreased fibrillarin, a nucleolar protein. Indeed, knockdown of fibrillarin is sufficient to reduce nucleoli size and increase lifespan in worms.
Nucleolar size inversely correlates with lifespan, and as such is a cellular hallmark of longevity. Tiku V, Jain C, Raz Y, et al. Small nucleoli are a cellular hallmark of longevity. Nature Communications. 2017;8:16083. doi:10.1038/ncomms16083.
Wild C. elegans
C. elegans research has primarily been performed on a single, Bristol isolate of C. elegans, called ‘N2’. However, there are myriad natural isolates of C. elegans around the globe. Resources for wild isolate research include those of Mark Blaxters lab in Edinburgh, and Erik Anderson, who manages the association mapping resource CeNDR. Jan Kemmenga has also produced a library of sequenced RILs between two highly polymorphic strains, the Bristol and Hawaiian isolates, that can be used to map differences in any phenotypic differences between these strains.
Marianne Félix, of the Marie Curie Institute in Paris, has been sampling wild C. elegans for many years, studying natural variation and using polymorphic genomes for quantitive genetic approaches to C. elegans research. Marianne described at the meeting a recently accepted (Lise Frézal et. al., Current Biology, in press) story investigating germline mortality in a C. elegans wild isolate. She described how the project started by leaving wild worms in a petri dish on her bench – resulting after a few generations in a sterile population. Luckily the lab was able to recover the population before it was too late, and realised they were observing a temperature-sensitive Mrt phenotype (mortal germline). Mrt phenotypes had already been observed in screens by Jonathan Hodgkin, which isolated mutants in DNA repair. The lab found that many of the wild isolates exhibited Mrt phenotypes to differing degrees of severity – the strain QX1211, for example, exhibits sterility in one generation. Many of these strains were isolated from hot countries, raising the question of how these strain survive in their natural habitats. Marianne observed that infection of the worms by natural pathogens was able to suppress the Mrt phenotype, indicating that the phenotype was in fact an artefact of the worms’ sterile environment in the lab petri dish. In order to understand the genetics behind why some strains exhibit the Mrt phenotype and not others, RILs were created between two strains, one Mrt and one not, JU1395 and MY10. Through phenotyping followed by whole genome sequencing, the Mrt phenotype was mapped to the set-24 locus on chromosome II. However, upon inspection of set-24 alleles across various wild isolates, Marianne’s lab found that the set-24 Mrt allele was, in fact, a rare allele in the wild. Thus to understand the common factor shared between the Mrt wild isolates, GWAS (genome-wide association mapping) was performed across 95 wild isolate genomes. This approach identified the more commonly present Mrt factor as morc-1, on chromosome III, a gene which helps to silence repetitive elements. Thus two different quantitive genetic approaches to find the cause of the same phenotype resulted in the identification of two different Mrt alleles, one of which was relevant for a particular strain (set-24), and one of which helped describe the differences between many wild isolates (morc-1).
Family, friends and enemies of Caenorhabditis
Although Caenorhabditis elegans took centre stage at the meeting, there were a few friends and even enemies of our favourite worm present. Marie Delattre from the Université de Lyon in France told us about one such worm, Mesorhabditis belari, which has a curious mechanism of genetic reproduction. For comparison, C. elegans has two sexes: hermaphrodite (of genotype XX) and male (of genotype X0). Populations are predominantly hermaphrodite, although the loss of one X chromosome at a rate of 1:1000 means that males do occur at a very low incidence. If a male mates with a hermaphrodite, mendelian genetics dictates that 50% of the resulting cross progeny, fertilised by sperm not carrying an X chromosome, are male. By contrast, Mesorhabditis belari is present in populations comprised of female and male individuals (so far so good). However, although the male is required by the female to fertilise her oocytes and to produce viable offspring, the male animal only rarely passes any of its DNA to the next generation, and progeny are predominantly clones of the female parent. So what is the point of producing males at all? Marie discovered that females produce two different kinds of embryos upon oocyte fertilisation. To understand the difference between them, it should first be mentioned that C. elegans oocytes, as well as those of Mesorhabditis belari, undergo only meiosis I before fertilisation by the sperm; meiosis II occurs shortly after fertilisation and it is only after expulsion of the sister gamete that the male pronucleus joins the female pronucleus to form the zygote. So, to go back to the oocytes of Mesorhabditis belari, it was found that 10% of oocytes carry out meiosis II after fertilisation, as expected, followed by fusion of the male and female pronuclei and subsequent development of the embryo – into male worms only. The remaining 90% of fertilised oocytes of Mesorhabditis belari do not complete meiosis II at all. Instead, the male pronucleus is prevented from expanding and fusing with the female nucleus, which enters mitosis to produce a female embryo – containing only the DNA of the female parent. In this scenario, it would seem that there is no evolutionary advantage to producing males. Marie turned to evolutionary game theory to provide some answers. She modelled two genetically distinct populations of Mesorhabditis belari, using factors to describe population size (i.e. how many progeny are produced) and how many worms will migrate away and towards each population. Using this model, she was able to identify a scenario in which males of Mesorhabditis belari presented an advantage to the population: if the males were to preferentially mate with their sisters. Indeed, Marie realised that the very low mating efficiency of these worms in the lab may also be explained by this preference, and upon performing the experiment, she found this to be the case. Thus, males provide their genetically similar sisters with sperm for the activation of oocytes, initiation of polarisation and for providing centrosomes – just not for their DNA.
A more nefarious creature (at least from the point of view of C. elegans researchers) is Pristionchus Pacificus. This is a dimorphous worm that in one of its forms is carnivorous – enjoying munching on C. elegans, amongst its other prey.
If one can overcome the macabre, however, an interesting question presents itself: how does Pristionchus Pacificus avoid becoming cannibalistic? James Lightfoot, from the lab of Ralf Sommer at the Max Planck Institute for Developmental Biology in Germany, explained how self-recognition (and thus avoidance of eating its own species) was achieved. In order to test the limits of self-recognition, James mixed populations of four different carnivorous species (including the sister species of Pristionchus pacificus) together on a plate, and observed that all four species recognised and avoided eating only themselves. If larvae from their own species were allowed to populate a plate and were later removed and replaced by the larvae of an alternative species, the new larvae were not protected from the predatory worms. Together these experiments suggested that self-recognition involved a surface-bound molecule. During his investigation, James found that individual isolates of Pristionchus pacificus, such as those isolated from Washington and those from California, will happily eat each other (even though they are members of the same species) but avoid worms from their own genetic pool. This observation allowed him to cross together the two strains to produce recombinant inbred lines (RILs), followed by phenotyping and sequencing the resulting strains. Impressively, the lab had to overcome a significant incompatibility region that obscured the data, and eventually managed this by inducing double stranded breaks along the chromosomes using CRISPR. This resulted in the identification of the locus ‘self-1’. Deletion of this locus resulted in a loss of self-recognition, and it was found to encode a 63 amino acid short peptide. An alignment of 38 strains revealed that although most of the self-1 sequence was highly conserved, it contained a highly variable terminal sequence. In addition, the copy number of this gene varied extensively between strains. James and his colleagues found that changing this variable region between strains was enough to protect the opposite strain, and that even a single amino acid change in this region was enough to affect self-recognition. However, he has identified some strains that carry identical self-1 sequences, but do not eat each other, suggesting that there are as yet still undiscovered mechanisms that regulate self-recognition in Pristionchus pacificus.
EMBO Young Investigator lecture
Luisa Cochella of the Research Institute of Molecular Pathology (IMP) in Vienna, delivered the EMBO Young Investigator lecture. Luisa studies the role of miRNAs (short, approximately 22nt long non-coding RNAs that repress gene expression) during development, and started her own lab after working as a postdoc in Oliver Hobert’s lab at Columbia University.
Luisa Cochella presents the EMBO Young Investigator lecture at the European Worm Meeting. Her slide is showing John Sulston’s lineage of the worm.
Luisa described how it has been previously proposed that miRNAs can act as ‘de-noisers’ of transcription, expressed in domains where their target genes are not normally transcribed, to reinforce the transcriptional control of the gene. Alternatively (and she suggests, perhaps more interestingly), miRNAs can act on their own to change cell identity. miRNAs begin their existence as miRNA precursor molecules (a self-annealing loop of RNA) which must be processed by first the RNase III Drosha, together with Pasha (partner of Drosha), and then again by Dicer to produce the mature miRNA which can be recognised by Argonaute. Embryos of a strain containing a temperature sensitive allele of Pasha, pash-1(mj100), fail to elongate during embryogenesis at the restrictive temperature, showing that miRNAs are essential for embryogenesis in the worm. Individual knockouts of 90 of a total 150 miRNAs, performed by Eric Miska and Bob Horwitz, showed no developmental arrest. However, upon deletion of entire miRNA families, they found that two families in particular were important for development. These were the mir-35 family (comprising 8 members) and the mir-51 family (6 members). Deletion of the mir-35 family phenocopies the pash-1 allele, whereas deletion of the mir-51 family resulted in animals that arrested slightly later during embryogenesis. Rescue of a miRNA gene family knockout strain can be achieved by any one of the family members.
By sequencing miRNA from embryos, Luisa revealed that 75% of all miRNA molecules belonged to one of the mir-35 or mir-51 families. Using a micro-processor by-pass strategy, which uses splicing machinery rather than Dicer to process synthetic ‘mirtrons’, the lab was able to rescue pash-1 mutant embryos with only two mirtrons, representing each of the mir-35 or mir-51 families. However, these animals still arrested during larval development, suggesting a role for the remaining 25% (by expression) of miRNAs. Despite some of these 25% of miRNAs being expressed at very low levels, Luisa hypothesised that they may be essential for very specific developmental events, and could be expressed in as little as one cell during development. She tested the expression patterns of these lowly-expressed miRNAs and found that they were, indeed, expressed in very restricted domains. One such miRNA, mir-791, was found to be expressed in a subset of neurones: BAG, AFD and ASE. These are all CO2-sensing neurones. In behavioural assays, loss of mir-791 resulted in a reduced avoidance response, and could be rescued by reintroduction of mir-791. This indicated that mir-791 has a specific role in conferring fate of these neurones, and is not merely involved in the robustness of the expression of target genes. Luisa found that the target genes for mir-791 were two ubiquitously expressed genes, akap-1 and cah-3. The only cells where the expression of these genes is repressed is in the CO2-sensing neurones. In this way, she has found that miRNAs may act in parallel to transcription factor control of gene expression to confer specific cell fates.
Keynote talk
Paul Sternberg from CalTech in California delivered the keynote talk at the meeting. The worm community owes Paul a huge debt for the many research resources that he has cofounded in his time working on C. elegans – including Wormbook, Textpresso and Wormbase. He began his career with C. elegans with the Horwitz lab at MIT at about the same time as John Suston’s lineage was published, at the start of what would become a hat tick for C. elegans research: the lineage, the connectome and the genome. Using this information, he set out to ‘solve the worm’. His lab maintain a very broad spectrum of research interests, and here he highlighted their recent work on the dauer life-form of the worm. C. elegans larvae enter an alternative lifecycle during conditions of starvation or overcrowding, in which they may remain as a dauer animal for up to three months. Indeed, in the wild this is the most commonly-found life stage. Upon finding food, the dauer animals re-enter the life cycle as L4 larvae, and progress to adulthood. Paul suggests that the dauer stage might also serve other functions in the worm, however, than just a means to survive starvation. He notes that animals of the genus Steinernema are able to jump significant distances as dauers, due to the energy stored in their cuticle. Many genes are shared between Steinernema and Caenorhabditis, and his lab are now investigating the differences in gene expression between dauers and other larvae in C. elegans.
During his talk Paul highlighted the many resources available for C. elegans researchers, including the CRISPR-based knock-out by knock-in (using the hygromycin resistance cassette) library being created to complement the knockout consortium database of null alleles. He also mentioned the cGAL bipartite expression system that his lab has recently adapted for the worm, and finally his initiatives in micropublication. Micropublication is available through Wormbase and allows concise (one figure) reports to be made and referenced within the scientific community. As he reminds us at the end, the 1953 DNA double helix paper also had just one figure…
Keynote address: Worm Tales by John White
One of the meeting highlights was the keynote address delivered by John White, FRS. John was a graduate student in the lab of Sydney Brenner in the 1970s, and is now Professor Emeritus at the University of Wisconsin-Madison. During his time at the LMB, John co-developed confocal microscopy and published ‘The Mind of a Worm’, and in so doing founded the field of connectomics. His address focused on the historical perspective of modern Caenorhabditis research, from its origin as a brain child of Sydney Brenner to its establishment as a major genetic model organism. (Wormbook has a wonderful memoir describing John White’s experience as part of the first C. elegans research lab that is definitely worth a read.)
In his talk, John explained how Sydney would holiday at the laboratory of marine biology in Woods Hole, contemplating his next scientific adventure (he had already been involved with solving the genetic code, in the process discovering a STOP codon). He was particularly interested in a molecular solution to behaviour, and had the foresight to understand that this would involve a precise understanding of the research organism’s anatomy. However, current model organisms were either too complex (Drosophila has approximately 135,000 neurones) or genetically intractable for this purpose. Sydney came across Bovari’s lineages of the nematode Ascaris, which suggested that these worms were likely to have stereotyped lineages, and also Goldschmidt’s work, which showed Ascaris to have a very simple nervous system. A 1948 paper published in Nature by Dougherty and Calhoun had also suggested the possible significance of nematodes as model organisms. Thus, after some experimentation which showed the nematode nervous system could be visualised with electron microscopy, Sydney settled on the worm as his model. The first strain of Caenorhabditis elegans was isolated from mushrooms in Bristol in England, becoming the ‘canonical’ research strain, and 55 years ago Sydney isolated the first mutant worm.
According to John White, Sydney undertook a very successful PR campaign in the early ’70s to garner support for his new research model. He travelled around America, speaking at many events in order to attract the best and most adventurous postdocs to his laboratory – who also came with funding. People who worked at the LMB with Sydney included many current lab heads and notable researchers: Bob Horwitz, Marty Chalfie, Judith Kimble, Cynthia Kenyon, Barbara Meyer, Jim Preiss, Iva Greenwald, Andy Fire, Julie Ahringer, Benjamin Podbilewitz, John Sulston, Bob Edgar and Ed Hedgecock, amongst others. Graduate students included John White, Jonathan Hodgkin, Mario de Bono, Richard Durbin and Tony Hyman. John explained how Sydney’s approach to supervision could be described as ‘benign neglect’, centring around 10am coffee break discussions that may or may not be related to the lab’s current research. John attributes much of the success of early worm research to this strategy, which forged strong communication and support links between researchers.
Sydney’s first task was to get the genetics of the worm up to the standard of Drosophila, generating mutants and tools such as balancers, and also mapping the C. elegans genome. This work culminated in the publication of a single author paper in 1974. At the same time, the neuroanatomy of the worm was being dissected using electron microscopy. However, Sydney soon moved onto other scientific projects and left C. elegans in the hands of his coworkers. Jonathan Hodgkin took over the construction of the genetic map, and John Sulston decided to help with the effort of White and colleagues to reconstruct the neural network by completing the post-embryonic lineage of the worm. After finding out that those in charge of constructing the embryonic cell lineage were ‘doing it wrong’, Sulston apparently locked himself away for 18 months and produced the accurate map we have today. This work paved the way for the identification of lineage mutants, and the work on cell-cell communication and programmed cell death that was pioneered in the worm. An obituary for John Sulston written by John White can be found here.
John Sulston holding his cell lineage of the nematode C. elegans.
The whole meeting represented a wonderful collection of the continuation of the work initiated by John Sulston, during which he was remembered and missed by the C. elegans community. The historical perspective of John White was a good reminder of how creativity is fostered by the combination of curiosity, tenacity and communication between researchers, values that I believe are central components of the C. elegans research community and make it such a pleasure to be part of.
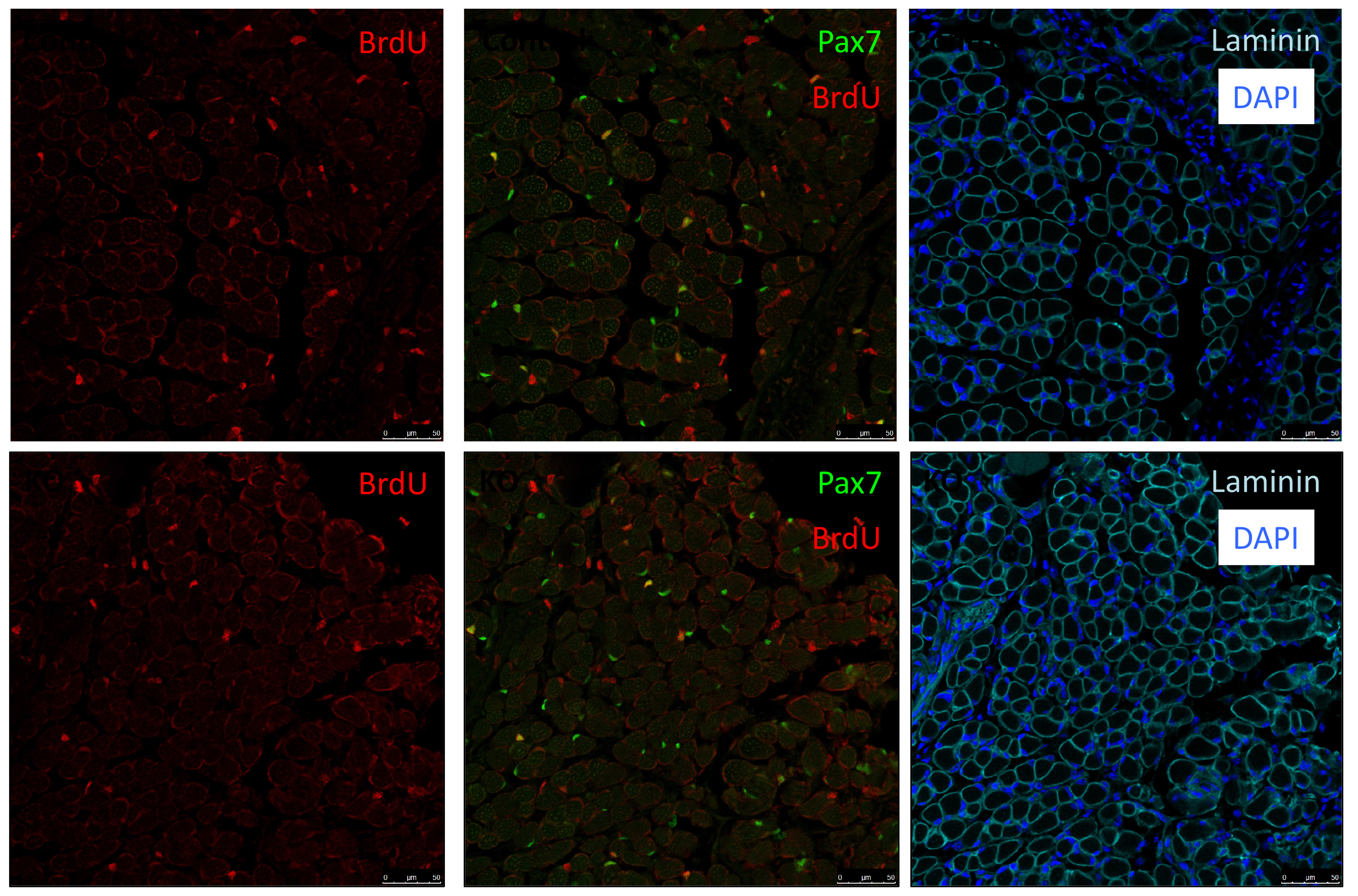
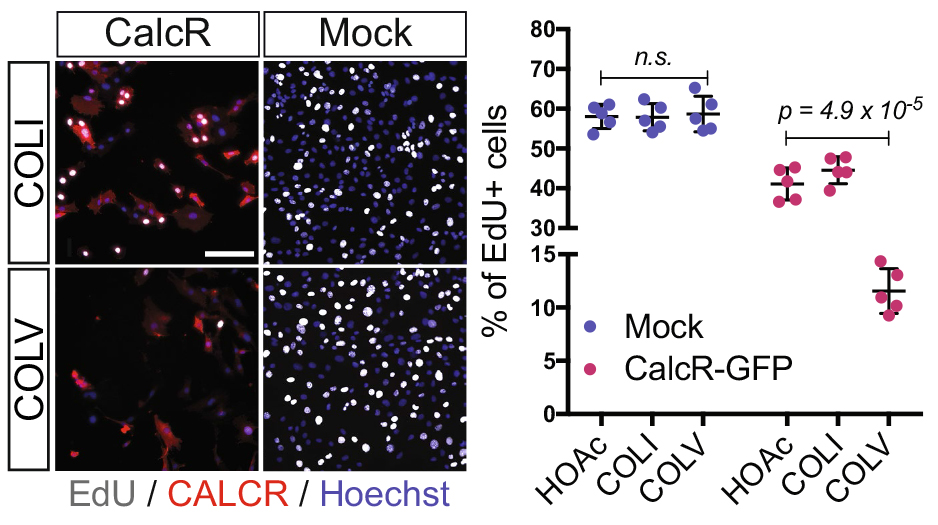
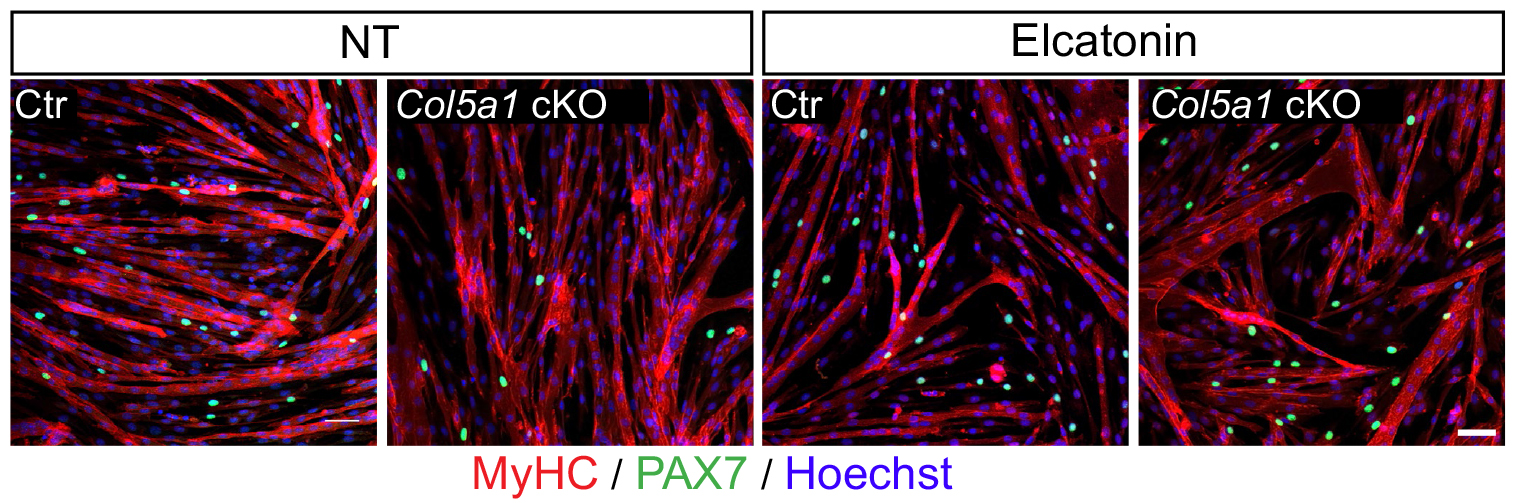
In our recent paper published in Nature, we unravel a new mechanism of an extracellular matrix protein secreted by muscle satellite (stem) cells, thereby playing the unusual role of acting as a signaling molecule to maintain the stem cell population. Here, I share the story behind this discovery and discuss the questions related to niche regulation.
In a ChIP-sequencing screen in myogenic cells performed by the Tajbakhsh and Stunnenberg labs (Castel et al., 2013) to identify direct targets of nuclear NOTCH (NICD) and its downstream effector RBPJ, we found an enrichment of extracellular matrix proteins, among which were specific collagens genes including Collagen 5 (Col5a1 and Col5a3) and Collagen 6 (Col6a1 and Col6a2). I was in the first year of my PhD when Dr. Philippos Mourikis, an ex post-doc in the Tajbakhsh lab, asked me to help with some collagen immunostainings on embryonic muscles over-expressing NICD (E17.5 Myf5Cre; R26NICD) (Mourikis et al., 2012). We nicely showed that both COLV and COLVI secretion were upregulated upon Notch induction.
As Col5a3 gene was the most responsive gene to Notch activity, both in vitro and in vivo, we thought that Col5a3 KO mice might present a muscle phenotype (Huang et al., 2011). In collaboration with Dr Guorui Huang from the Greenspan lab, we analyzed the number of satellite cells (PAX7+) in adult and postnatal P8 pups muscle sections. Unfortunately, we could not observe any alterations in stem cell number nor proliferation capacity (BrdU pulse).
PAX7 (satellite cells), BrdU (proliferation) and Laminin immunostaining on transverse sections of Tibialis Anterior muscle of Control (WT) and Col5a3 KO P8 (8 days after birth) juvenile mice. No difference in satellite cells number and proliferation capacity between control and mutant pups.
As my initial PhD project was on hold due to mice breeding issues, we decided that I could perform a simple culture experiment to close up the collagen story. We performed two sets of experiments: in the first, we added different collagens in soluble form on cells plated on gelatin-coated dishes; in the second, cells were cultured directly on collagen-coated dishes. Surprisingly, only COLV in its soluble form retained cells in a more stem-like state delaying their proliferation and differentiation. With these unexpected results, we realized that COLV might be acting as a signaling molecule rather than a biomechanical cue. As collagen molecules are known to bind to specific receptors such as Integrins or Discoidin Receptors (Leitinger, 2011), we tested whether their inhibition could abrogate COLV anti-myogenic effect. To our disappointment, COLV was still able to delay myoblast differentiation even when these receptors were inhibited.
After looking at the results from several angles, we came to the conclusion that we lacked a mechanism and we should probably wrap up the story with what we had, so that I could go back to my initial PhD project. With a sense of unfinished business, we started gathering the data into figure formats. We were then struck by an old RT-qPCR assay on satellite cells that were treated with the different collagens, where Calcitonin receptor (CalcR), a known satellite cell quiescence marker, was upregulated more than 20-fold. We remembered that CALCR is a G-protein couple receptor (GPCR) and found in vitro evidence that some collagens could indeed bind to GPCRs (Luo et al., 2011; Paavola et al., 2014). This was a major turning point in the project, and the path was wide open for detailed mechanistic studies.
In parallel on the other side of the world, our collaborator Dr. So-Ichiro Fukada in Osaka had just a few weeks earlier published a very nice study showing that specific ablation of Calcr in muscle stem cells led to their spontaneous exit from the quiescent niche (Yamaguchi et al., 2015). Using their retroviral tools to overexpress CALCR and Calcr null muscle stem cells, we were able to demonstrate that CALCR was necessary to respond to COLV. This was too exciting, my other project had to wait!
EdU (2h pulse) and CALCR staining of GFP+ C2C12 cells isolated by FACS and transduced with Calcr-GFP or mock-GFP retrovirus and cultured for 24 h with COLI (top) or COLV (bottom). Quantification of EdU+ Calcr-transduced C2C12 cells or mock-GFP cells treated for 24h with COLV or with the controls, COLI and HOAc. There was no significant difference between HOAc and COLI treated samples.
On the other hand, these results were puzzling…CalcR already has its ligand: Calcitonin hormone produced by the parathyroid gland, suggesting that satellite cell quiescence is under systemic control. Why would the regulation of muscle stem cells that are dispersed throughout muscle masses in the body – and that would be solicited with different kinetics – be under systemic control? Can COLV act as the local ligand of CalcR for muscle stem cells?
Meanwhile, our first priority was to assess whether the loss of function of COLV specifically in satellite cells using Tamoxifen-inducible Tg:Pax7-CreERT2; Col5a1flox mice could affect their behavior. Another unexpected surprise was the observation that only 18 days following recombination, satellite cells escaped the quiescent niche, differentiated and fused to the pre-existing fibres.
Another consideration is the source of collagen in skeletal muscle – being predominantly from interstitial myofibroblasts, which deposit collagen between myofibres (Zou et al., 2008). We also found expression of Col5a-1, -2, and -3 in other cell types present in the muscle; namely Fibro-Adipogenic Progenitor (FAPs) and resident macrophages.
So why does this source of COLV not compensate for the loss of satellite cells-produced COLV?
Possible explanations include the basal membrane (BM) forming a physical diffusion barrier between the satellite cells and the interstitial space – rendering them blind this source of COLV. Although the myofibre that is in intimate contact with satellite cells under the BM could be a provider, we showed that myofibres do not express detectable COLV to sustain satellite cell quiescence.
Another possibility is that COLV produced by satellite cells has a specific configuration. In our siRNAs experiments on isolated myofibers, we observed that targeting of the Col5a3 isoform phenocopied siRNA against Col5a1, strongly suggesting that the active triple helix is the a3-COLV isoform composed of both a1 and a3 chains as an [α1(V)α2(V)α3(V)] heterotrimer. Although the presence of soluble COLV is difficult to demonstrate in vivo, we favoured the possibility that COLV produced by satellite cells rapidly binds to CALCR upon secretion, rather than being integrated in the collagen network. This would then explain why loss of COLV function exhibited a phenotypic loss of satellite cells within such a short period.
We then needed to demonstrate that the newly discovered Notch/COLV/CALCR axis is functional for the maintenance of quiescent cells in vivo. To do so, we injected Col5a1 cKO (Tg:Pax7-CT2; Col5a1flox) mice subcutaneously with a derivative of the known CALCR ligand calcitonin (Elcatonin). Interestingly, exogenous Elcatonin administration compensated for the lack of COLV and prevented their exit from quiescence, thereby validating our results linking COLV and CALCR. It also told us that endogenous calcitonin was not bioavailable in the muscle to rescue the loss of COLV. Interestingly, thyroidectomy in adult individuals does not show any striking overall pathological consequences (Russell et al., 2014); it would be interesting to study satellite cell dynamics in mice suffering from hyper/hypothyroidism.
An interesting outcome of the rescue experiments was that Elcatonin administration in control mice also showed an improvement in self-renewal capacity, suggesting that this could be a good therapeutical candidate for myopathies.
Rescue of loss of COLV by Elcatonin in an ex vivo self-renewal reserve-cell model, where PAX7+ non-proliferative cells return to quiescence. MyHC and PAX7 staining of Control (Ctr: Tg:Pax7-CT2; Col5a1+/+; R26mTmG) and Col5a1 null (Tg:Pax7-CT2; Col5a1flox/flox; R26mTmG) non-treated (NT) or treated with Elcatonin (Elcat). No GFP+/EdU+ cell (12h pulse) could be detected in any of the conditions indicating GFP+ cells are quiescent (data not shown).
In addition, it has been shown that the satellite cell population is heterogenous according to the muscle anatomical location and even within a single muscle (Rocheteau et al., 2012; Sambasivan and Tajbakhsh, 2015). Therefore, we cannot exclude the possibility that some satellites cells do indeed respond to calcitonin stimulation. Our results showing that Elcatonin administration induces a deeper quiescent state could support this notion. In addition, CALCR is expressed in about 80% of satellite cells, suggesting that 20% of them are most likely not under direct Notch/COLV/CALCR control.
While exploring the quiescence in muscle stem cells, we learned that when studying the role of collagens one should consider not only the traditional mechanotransduction role of these molecules, but also their potential role as signaling agents. Given that neural and intestinal, and perhaps other stem cells, are maintained by Notch signaling, it is possible that this model operates in stem cells located in other tissues and organs.
 (No Ratings Yet)
(No Ratings Yet)






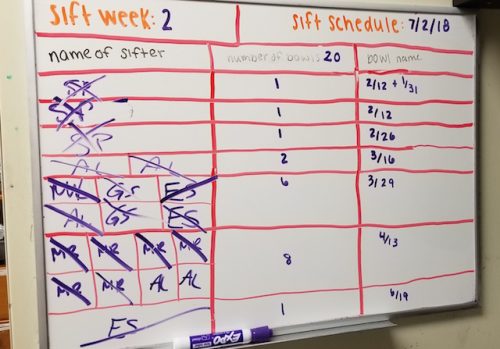


 (4 votes)
(4 votes)




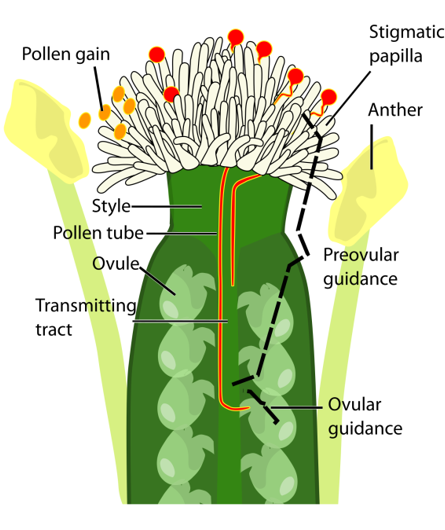

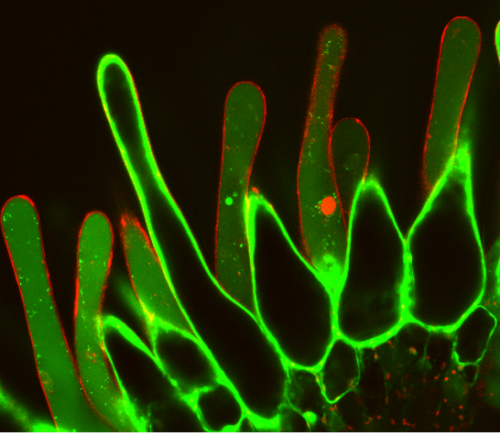
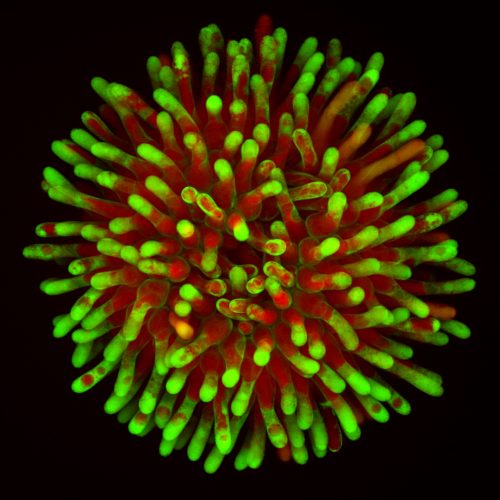


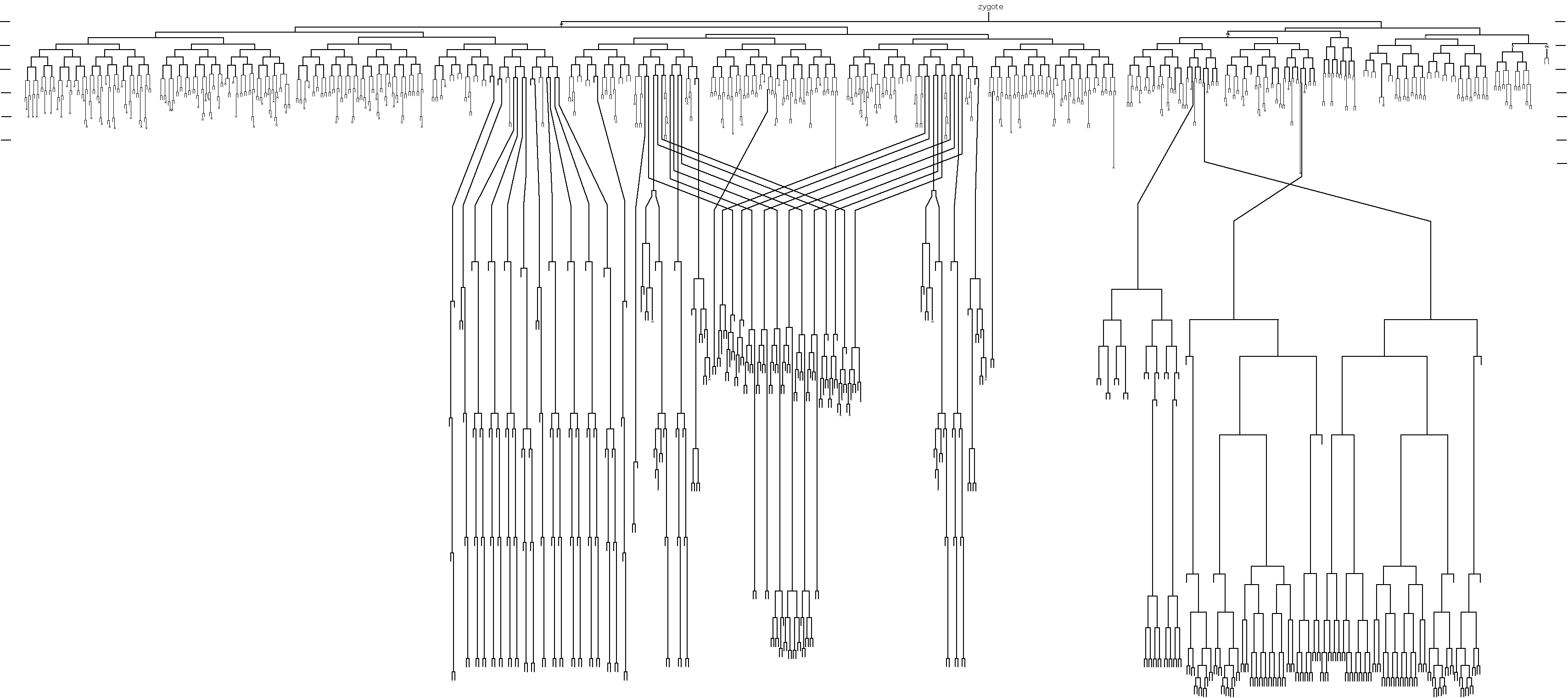


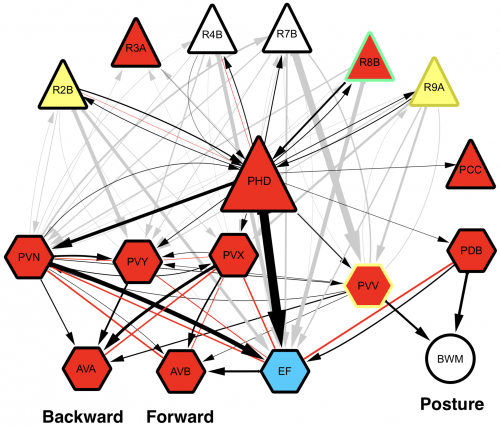
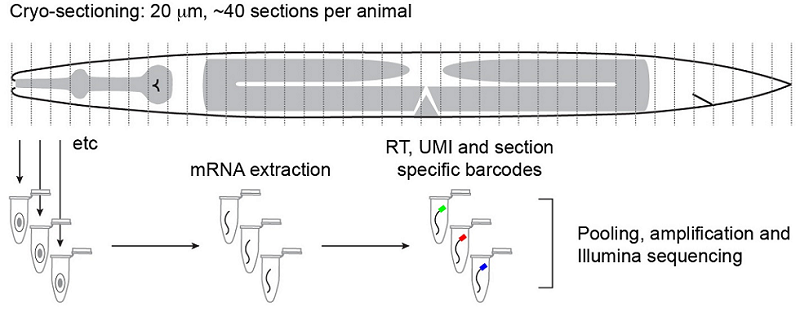
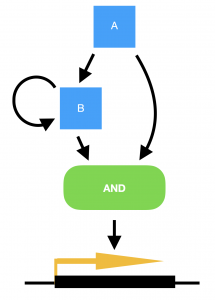
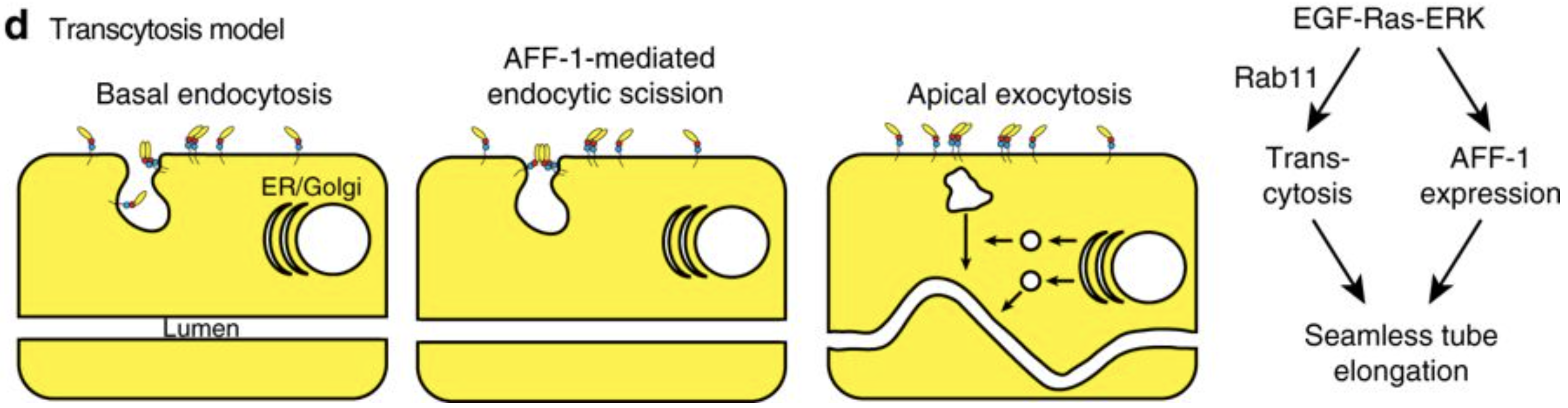
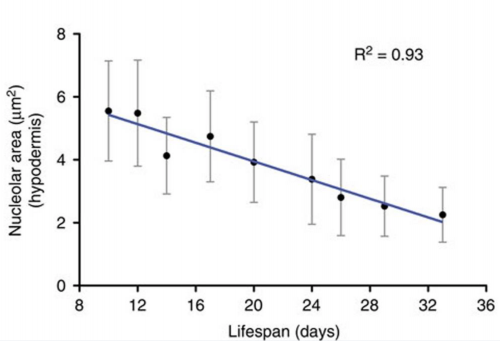


 (13 votes)
(13 votes)