Development and homeostasis depend crucially on the maintenance of cell identity, and in gamete-producing tissues the somatic/germline distinction is paramount. A recent paper in Developmentexplores how cell identity is secured in the Drosophila ovary by studying the function of the conserved tumour suppressor L(3)mbt. To find out more about the story, we caught up with first author Rémi-Xavier Coux and his supervisor Ruth Lehmannof the Skirball Institute at New York University School of Medicine.
Remi and Ruth at graduation
Ruth, can you give us your scientific biography and the questions your lab is trying to answer?
RL I grew up in Germany and received my PhD in the lab of Christiane Nuesslein-Volhard. I was in her lab at the very beginning when the genes required for establishing embryonic polarity and pattern were identified. My project involved the genetic analysis of gap genes and a group of maternal effect genes, we termed the posterior group of genes, as mutation in these genes affect the development of the embryonic abdomen and my cytoplasmic transplantation experiments suggested a gradient of patterning activity emanating from the posterior pole. I became intrigued by this posterior pole plasm as it is also the site of germ cell formation in Drosophila. So, I made the more unusual and often discouraged decision to stay with my graduate project, yet decided to delve into molecular analysis of these genes. By the time I arrived for my first independent position at the Whitehead Institute at MIT in 1988, I had molecular entry points for the identification of the nanos, oskar and pumilio genes in my suitcase.
Analysis of these genes was really exciting as it showed that the mRNAs encoding Oskar and Nanos were localized to the posterior pole and that the translation of these RNAs was specifically regulated such that only the localized RNA (and now we know that is only a very small fraction of the total RNA) is translated while unlocalized RNA is translationally repressed. From the beginning, I have been amazingly lucky to have incredibly talented graduate students and postdocs in my lab, not only did they make all these discoveries but they also taught me a lot.
In 1996, I was recruited to the newly founded Skirball Institute at NYU School of Medicine. Here my lab focused completely on germ cell biology studying the germ line life cycle. We are particularly interested in three areas:
1. How the germ line-soma dichotomy is initially established in the early embryo. Critical for the initial distinction between soma and germ line are the properties of membraneless germ granules that co-ordinate the posttranscriptional regulation of RNAs specifically needed for germ cell formation, specification, transcriptional silencing and germ cell migration to the somatic gonad. Crucial for progress here has been the development of ever so powerful imaging modalities (from light sheet microscopy and super resolution microscopy to electron microscopy) and the development of many different ways to mark molecules and observe them in vivo.
2. How interactions between cells from different origins coordinate growth and differentiation of the gonad so that primordial germ cells mature into germ line stem cells and deposit eggs during adult life. Initially, we relied on forward genetic screens to address this big and fascinating problem of organ development but now we are increasingly take advantage of whole genome genetic analysis by RNAi and Crispr/Cas9 as well as high-throughput molecular analysis such as single cell sequencing at different developmental stages.
3. Finally, we are interested in how the unique role of germ cells as the only cells of the body with the potential to give rise to a complete new organism manifests specialized adaptions. So, we have become intrigued by the broadest sense of mutual ‘host-pathogen’ interactions as they relate to the germ line. Here, we are interested in how germ line regulatory mechanisms manage to control transposable elements activity, the interplay between genome and mitochondria during the germ line life cycle and emerging optional relationships like the ability of the intracellular bacterium Wolbachia to grow in the Drosophila host without direct harm. We reason, that such germline specific control and defence mechanisms protect the germ line, but may also provide opportunity for species to evolve via changes to the germline.
And Rémi-Xavier, how did you come to be involved with this project?
RXCl(3)mbt was identified in the laboratory of Elisabeth Gateff, who was the first to use Drosophila to identify tumor-suppressor genes (Gateff, 1978, Gateff et al., 1993). As the name says, ‘l(3) malignant brain tumor’ mutations cause brain tumors in Drosophila larvae. Many years later, Chris Yohn, a postdoc in our lab, identified several alleles of l(3)mbt in a clonal screen for maternally expressed genes that affected germ cell formation in the embryo. While Chris determined that the PGC formation defect was a secondary consequence of L(3)mbt’s role in early embryonic nuclear divisions, he also observed that l(3)mbt females were viable at the permissive temperature (brain tumors form only at high temperature) but they were completely sterile and produced no eggs. This suggested an additional role for L(3)mbt in gonadogenesis (Yohn et al., 2003). The project rested for a while, until Cayetano Gonzalez and his group at the IRB in Barcelona published an intriguing observation. The Gonzalez lab profiled l(3)mbt larval brain tumors and showed that in these tumors many germline genes were apparently derepressed (Janic et al., 2010). We were intrigued by this possible soma-to-germline transformation, so when I joined the lab as a graduate student I started two projects on l(3)mbt. First, I asked whether l(3)mbt brain tumor cells indeed behaved like bona fide germ cells and could be used to identify novel regulators of the germline fate. Second, I wanted to test whether the sterility phenotype was also due to a soma-to-germline transformation. Our paper describes the results of the later study that proved much more successful than the former.
Drosophila ovaries and the l(3)mbt phenotype, from Fig. 1 in the paper
Can you give us the key results of the paper in a paragraph?
RL&RXC In addition to its role in suppressing brain tumors in the larvae, L(3)mbt functions in two tissues of the ovary to safeguard oogenesis: the somatic support cells and the germline. We found that in both tissues, L(3)mbt prevents expression of genes incompatible with normal development: in the somatic ovarian cells, L(3)mbt represses genes normally expressed in the germline while in the germ cells, it silences testis and neuronal genes. This, and the fact that l(3)mbt mutant tissues still express genes characteristic of the tissue of origin revealed a function broader than previously thought. Our study therefore suggests that L(3)mbt functions as a tissue specific transcription repressor rather than simply silencing germline genes in somatic tissues. L(3)mbt binding sites overlap with insulator elements (Richter et al., 2012) so it is also possible that L(3)mbt functions in insulator complexes.
Do you think L(3)mbt work with different partners in different tissues? Any ideas how it plays tissue-specific roles?
RL&RXC Indeed, L(3)mbt has been found to function with the dREaM/MMB and LINT complexes in S2 and Kc167 somatic embryonic cells (Georlette et al., 2007; Meier et al., 2012). However, our genetic studies suggest that L(3)mbt functions independently of dREaM in the ovary. Several components of the dREaM complex are required for endo-replication (polyploidization) of both somatic ovarian cells and nurse cells in the germline, but these processes seem not to require L(3)mbt function. It would be interesting to test if L(3)mbt functions in only one of these two chromatin complexes in other tissues besides the ovary and how the potential switch between complexes is regulated.
Larval ovaries from Fig. 3 in the paper
Your results suggest that rather than guarding against transdifferentiation, L(3)mbt guards against adoption of mixed identity. Where does this leave the concept/importance of transdifferentiation in development?
RL&RXC This is a conceptual question we really struggled with: the “orthodox” definition of transdifferentiation is complete acquisition of another cell identity. However, functionally testing fate switching in vivo is very challenging. Jarrault and colleagues beautifully showed that in C. elegans the Y epithelial cell transdifferentiates into a fully functional neuron (PDA) in wild-type larvae (Jarrault et al., 2008). It is one of the only examples of complete transdifferentiation to our knowledge. There are many more examples, especially with mutations in chromatin factors, where cell-specific gene signatures are misexpressed. At the end, it comes down to the assay that is used to define a ‘cell fate switch’. For example, can we call a cell that aberrantly expresses most of another cell type’s transcriptome trans-differentiated without a functional assay?
The l(3)mbt, nos double mutant ovaries, from Fig. 5 in the paper.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
RXC Yes, I was very glad to observe that nanos mutations suppress the l(3)mbt mutant ovarian phenotypes. It was late at the confocal and I immediately emailed Ruth. The morning after, we checked that embryos laid by l(3)mbt, nos double mutant females had the typical patterning defects caused by nos mutations. When this control was done, we were really excited!!!
And what about the flipside: any moments of frustration or despair?
RXC I tried transplanting l(3)mbt tumorous brain cells into embryos devoid of germline to test if they could, at least partially, behave as germ cells. This experiment was quite challenging, and we could not detect the cells a few hours after transplantation, which was very frustrating! We now know that this experiment likely did not work because these neural-origin cells may not be completely transformed.
E2f2 mutant clones from Fig. 7 in the paper
What next for you Rémi-Xavier?
RXC I just started a postdoc in the Cohen-Tannoudji and Navarro-Gil labs in the Stem Cell Biology and Development Department, Pasteur Institute in Paris. I will study and characterize Transcription Factor bookmarking in the early mouse embryo.
Where will this work take the Lehmann lab?
RL Further study of L(3)mbt may provide us with clues about the transcriptional mechanisms underlying germline-soma dichotomy. We found in our study that mutating a key regulator of germline fate, the translational repressor Nanos, suppresses the somatic gonadal defects of l(3)mbt mutant ovaries almost completely. Thus, Nanos targets may be key regulators that distinguish between the germline and soma program and the l(3)mbt mutants may guide us towards their identification.
Finally, let’s move outside the lab – what do you like to do in your spare time?
RXC In my free time, I enjoy music, modern art, playing rugby and outdoor activities such as sailing and mountain activities.
One of the most talked about preprints this month was a corrigendum to that Nature Methods paper reporting widespread off-target mutations following CRISPR-Cas9 editing in mice. The paper was challenged quickly on bioRxiv, and four days after the corrigendum went up, it was retracted, as reported by Retraction Watch! Blink and you’ll miss it.
Otherwise this month we found plenty of plant patterning, lovely larval lampreys and ovulating roundworms, and the developing diaphragm finding its voice.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
Megan B O’Hare, Alamin Mohammed, Kyle J Connolly, Katelyn C Aitchison, Niki C Anthoney, Amy L Roberts, Matthew J Taylor, Bryan A Stewart, Richard I Tuxworth, Guy Tear
Analysis of novel domain-specific mutations in the zebrafish ndr2/cyclops gene generated using CRISPR-Cas9 RNPs
Ashley N Turner, Reagan S Andersen, Ivy E Bookout, Lauren N Brashear, James C Davis, David M Gahan, John P Gotham, Baraa A Hijaz, Ashish S Kaushik, Jordan B McGill, Victoria L Miller, Zachariah P Moseley, Cerissa L Nowell, Riddhi K Patel, Mia C Rodgers, Yazen A Shihab, Austin P Walker, Sarah R Glover, Samantha D Foster, Anil Kumar Challa
Deep proteomic analysis of chicken erythropoiesis
Marjorie Leduc, Emilie-Fleur Gautier, Anissa Guillemin, Cédric Broussard, Virginie Salnot, Catherine Lacombe, Olivier Gandrillon, François Guillonneau, Patrick Mayeux
A critical role for miR-142 in alveolar epithelial lineage formation
Amit Shrestha, Carraro Gianni, Nicolas Nottet, Ana Ivonne Vasquez-Armendariz, Susanne Herold, Julio Cordero, Indra Bahadur Singh, Jochen Wilhelm, Guillermo Barreto, Cho-Ming Chao, Elie El Agha, Bernard Mari, Jin-San Zhang, Saverio Bellusci
Chromatin accessibility dynamics across C. elegans development and ageing
Jurgen Janes, Yan Dong, Michael Schoof, Jacques Serizay, Alex Appert, Chiara Cerrato, Carson Woodbury, Ron Chen, Carolina Gemma, Ni Huang, Djem Kissiov, Przemyslaw Stempor, Annette Steward, Eva Zeiser, Sascha Sauer, Julie Ahringer
Measuring cell area in iPSC lines, from Vigilante, et al.’s preprint
Identifying the genetic basis of variation in cell behaviour in human iPS cell lines from healthy donors
Alessandra Vigilante, Anna Laddach, Nathalie Moens, Ruta Meleckyte, Andreas Leha, Arsham Ghahramani, Oliver J. Culley, Annie Kathuria, Chloe Hurling, Alice Vickers, Mukul Tewary, Peter Zandstra, HipSci Consortium, Richard Durbin, Franca Fraternali, Oliver Stegle, Ewan Birney, Nicholas M Luscombe, Davide Danovi, Fiona M Watt
Placozoa and Cnidaria are sister taxa
Christopher E. Laumer, Harald Gruber-Vodicka, Michael G. Hadfield, Vicki B. Pearse, Ana Riesgo, John C. Marioni, Gonzalo Giribet
Frederic Delsuc, Herve Philippe, Georgia Tsagkogeorga, Paul Simion, Marie-Ka Tilak, Xavier Turon, Susanna Lopez-Legentil, Jacques Piette, Patrick Lemaire, Emmanuel J. P. Douzery
Quantifying the impact of public omics data
Yasset Perez-Riverol, Andrey Zorin, Gaurhari Dass, Mihai Glonț, Juan Antonio Vizcaino, Andrew Jarnuczak, Robert Petryszak, Peipei Ping, Henning Hermjakob
Helena Cousijn, Amye Kenall, Emma Ganley, Melissa Harrison, David Kernohan, Thomas Lemberger, Fiona Murphy, Patrick Polischuk, Simone Taylor, Maryann Martone, Timothy Clark
Gerald G Singh, Vinicius Farjalla, Bing Chen, Andrew Pelling, Elvan Ceyhan, Martin Dominik, Eva Alisic, Jeremy Kerr, Noelle Selin, Ghada Bassioni, Elena Bennett, Andrew Kemp, Kai MA Chan
Single-cell approaches are revolutionizing developmental biology. We can now trace in time the behavior of each cell in a live developing organism (1). In parallel, single-cell transcriptomics and genomics gives access to the transcriptional state of each cell (2). Combination of these two approaches promises to unravel how genomic information translates into individual cell behaviours.
We are harnessing these approaches to the embryos of ascidians (Tunicates, 3,4), a group of marine invertebrate chordates. These embryos develop with such stereotyped cell lineages that each cell of an embryo of one species has an exact homolog in all embryos of different, even distantly related, species. Contrasting with this extraordinary morphological conservation, the genomes of ascidians are widely divergent.
We are looking for a PhD student, who will study how the transcriptional program of homologous cells has evolved between distantly related tunicate species. She/He will focus on two classes of genes, those building the gene regulatory networks (GRN) driving development and the effector genes controlling cell behaviours through their action of cytoskeletal architecture.
The selected student will first establish single-cell RNA-seq for a range of developmental stages in 4 ascidians and 2 thaliaceans (a different class of tunicates, 5). From this dataset, She/He will analyse the level of conservation of the transcriptional programme across species, which could lead to a classification of genes/subnetworks according to their level of expression conservation. In a second step, she/he will select a few genes based on their pattern of expression conservation and their functional annotation and study their function in cell fate specification or embryonic morphogenesis using CRISPR/Cas9 technology.
The project is mostly experimental. Bioinformatics knowledge or a desire to acquire it will be a plus. More about the host group and institute can be found on the CRBM website: http://www.crbm.cnrs.fr/.
Collaborations with the Christiaen (NYU, New-York, USA), Zinzen (MDCC, Berlin, Germany) and Aerts (KU, Leuven, belgium) labs are foreseen.
References:
1) L. Guignard*, U.-M. Fiuza*, B. Leggio, E. Faure, J. Laussu, L. Hufnagel, G. Malandain, C. Godin#, P. Lemaire# (2017) Contact-dependent cell communications drive morphological invariance during ascidian embryogenesis. bioRxiv 238741 https://www.biorxiv.org/content/early/2017/12/24/238741 2) Karaiskos N, Wahle P, Alles J, Boltengagen A, Ayoub S, Kipar C, Kocks C, Rajewsky N, Zinzen RP (2017) The Drosophila embryo at single-cell transcriptome resolution. Science. 358:194-199 3) Lemaire P. (2011) Evolutionary crossroads in developmental biology: the tunicates, Development, 138(11):2143-52. 4) Lemaire P. (2009) Unfolding a chordate developmental program, one cell at a time: Invariant cell lineages, short-range inductions and evolutionary plasticity in ascidians. Developmental Biology ;332(1):48-60. 5) Piette, J. and Lemaire, P. (2015) Thaliaceans, the neglected pelagic relatives of ascidians: a developmental and evolutionary enigma. The Quarterly Review of Biology. 90(2):117-145.
How to apply?
Interested candidates are encouraged to contact Patrick Lemaire (patrick.lemaire@crbm.cnrs.fr) directly prior any formal application to the Life Science doctoral school in Montpellier (Deadline for application May 18th 14:00, French time).
International students are encouraged to apply, provided their master English. Understanding or speaking French is not necessary.
►Criteria to apply to the call
The contest for a doctoral contract is open to all candidates regardless of their nationality and institutions of graduation.
Twenty-four 3-year doctoral positions will be funded by the doctoral school
To be eligible, candidates must hold a Master’s degree (or diploma recognized as equivalent to a Master’s degree).
►Timeline for the 2018 campaign (All times are in Paris time zone.)
18th May 2018 at 14:00. Deadline for submission of applications
4th June: short listing of candidates
28th and 29th June 2018. Interviews of shortlisted applicants (video-conference for distant applicants should be possible)
Applications (single pdf file) should include the following documents:
A cover letter and CV, including academic background and professional experience, in particular internships accomplished as part of the study program.
Copies of diplomas. It is mandatory to hold a Masters degree by the beginning of the 2018/2019 academic year).
Academic transcripts of all courses taken as part of university degrees *.
At least two letters of recommendation. Referees are expected to explain how they have known the candidate, their frank evaluation of his/her capability and motivation to undertake a Ph.D., as well as of his/her creative and analytical skills. It is recommended that referee rank the student among all students they mentored (Top 5%, 10%, 20%).
The Company of Biologists is looking to recruit a Scientific Copy Editor for the journal Development – one of the leading international journals in the field of developmental biology. This is a permanent full-time position.
The role entails copyediting articles to a high standard, compiling author corrections, overseeing the journal production process, and liaising with authors, academic editors, external production suppliers and in-house staff to ensure that articles are published in a timely and professional manner.
Candidates should have a degree (ideally a PhD) in a relevant scientific area, and previous copyediting experience is strongly preferred. Additional requirements include excellent literacy skills, high attention to detail, a diplomatic communication style, good interpersonal and IT skills, a flexible approach and the ability to work to tight deadlines.
The position represents a unique opportunity to gain experience on our highly successful life-science journals and offers an attractive salary and benefits. The position will be based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV to recruitment@biologists.com, along with a covering letter that summarises their relevant experience, why they are enthusiastic about the role, and their current salary.
Applications should be received by 16 April and we welcome early applications or expressions of interest. Late applications may still be considered.
As many of you will be aware, The Company of Biologists initiated a search last year for a new Editor-in-Chief for Development, after Olivier Pourquié announced his intention to step down in September 2018. Following community consultation and a shortlisting and interview process, we are delighted to announce that James Briscoe will be the journal’s new Editor-in-Chief.
Many of you will already be familiar with James and his research. As a developmental neurobiologist working on the vertebrate spinal cord, he has a particular interest in using quantitative approaches to understanding how signalling pathways (particularly sonic hedgehog) regulate gene expression networks to control patterning, cell fate specification and growth of this tissue. His lab uses a range of in vivo, in vitro and in silico models to gain insight into this question at the molecular, cellular and tissue scales. For those interested in finding out more about James, we invite you to read our Spotlight interview with him elsewhere in this issue.
The position of Editor-in-Chief of Development is an important one in the developmental biology field – given Development’s position as a key community journal. The Editor-in-Chief is responsible for (among other things) appointing the editorial team, overseeing the handling of papers submitted to us, and setting new priorities for the journal. It was therefore important to us to gather input from the community as we set about appointing a successor to Olivier. We are hugely grateful to those of you – editorial board members, referees, authors and readers – who took the time to respond to our community consultation and provide feedback, not only on who they would like to see running Development, but also more broadly on how we are doing as a journal. Many of you are aware that, as a member of The Company of Biologists’ Board of Directors, James was part of the advisory group who initiated the consultation. He stepped away from any involvement in the process as soon as his name began to appear in nominations. The responses were collated by the three of us, and here we would like to share with you some of that feedback.
Your responses were, in general, consistent with our own assessment of the journal’s standing. We heard that Development is seen as a high-quality, rigorous venue for publication, with excellent academic editors, a ‘tough but fair’ review process and good production values. We were delighted that many respondents picked up on some of our more recent innovations – cross-referee commenting during peer review, openness to preprints, and our strong online presence, particularly through our community blog the Node. Given that the Node was launched at least in part in response to the consultation we conducted when looking for a new Editor-in-Chief to replace Jim Smith, it was fantastic to see how much traction the Node has now gained in the community. A recurring theme in the feedback on the journal’s strengths was that Development is the ‘journal of reference’ for the field, and that Development papers ‘stand the test of time’. If this is how we are seen in the community, we are clearly doing something right!
You also told us that Development can be seen as ‘hard to get into considering its impact factor’ and that competition from newer journals means that Development is sometimes seen as a less attractive choice, especially for early career researchers. These are of course issues of which we are all too aware. As signatories of the San Francisco Declaration on Research Assessment (DORA), we would argue that impact factor is a poor proxy for journal quality, and an even worse one for individual papers. That this measure still holds so much sway, particularly in certain geographic regions, is disappointing, and is something that The Company of Biologists – as a supporter of the newly revamped DORA project – is trying to change. Nevertheless, Development recognises that you as authors have a wide choice of journals to which you can submit, and the team will continue to work to make the journal an attractive option.
Looking to the future, we were delighted to hear that many respondents feel it is important for Development to maintain and strengthen the focus on stem cells, regeneration and human development. However, this is clearly not the only area in which developmental biology is growing: there was strong support for increased visibility in the genomics, biophysics, quantitative and systems biology, and evo-devo fields. These are all fields that Development has highlighted as future priorities for the journal, and the team looks forward to working with many of you to realise the potential of these areas.
Away from consideration of specific research areas, we heard that Development should do more to support and advocate for the field (this is actually something the Development team is actively working on – look out for news in an Editorial in the near future!) and that we should continue to support our communities through our charitable activities – and, if possible, grow these. Development and The Company of Biologists see these activities – meeting grants, travelling fellowships, workshops and so on – as a key part of our ‘raison d’être’, and you can rest assured that The Company will continue to give back to the community as much as we can.
On the specifics of the choice of new Editor-in-Chief, your feedback helped us to draw up a ‘wish-list’ for the kind of person we wanted to lead the journal. We were looking for an individual whose research is at the cutting edge of developmental biology, with interdisciplinary skills, broad interests in the field and a strong vision for the future of the journal – to build on the developments that Olivier initiated, bring new ideas to further strengthen the journal, and be an active advocate for Development. James fulfils all these criteria and more, as evidenced by the fact that his name came up over and over again in the feedback we received; in fact, he was suggested by almost half of those who provided specific names – around four times as often as any other single individual. The unanimous view was that James is an outstanding candidate to succeed Olivier, and we are delighted that he has agreed to take on this role.
Over the next few months, James will be working alongside Olivier to ensure a smooth handover when Olivier steps down in September. You’ll be hearing more from him later in the year when he officially takes over the editorship. As James gets ready to take the reins, he (and the journal more broadly) welcomes any feedback or suggestions that you might have for how you would like to see the journal develop. In the meantime, we hope you will join us in congratulating James on his appointment, and in wishing him luck as he steps up to this challenging and important position!
An interview with James Briscoe
Katherine Brown
James Briscoe is a group leader at The Francis Crick Institute in London. His lab’s research focusses on the developing vertebrate spinal cord, with a particular interest in how sonic hedgehog gradients, and the downstream signal transduction and transcriptional networks, regulate the development of this tissue. In September 2018, James will become the new Editor-in-Chief of Development. We met with James to discuss his career and research interests, the importance of interdisciplinary thinking in developmental biology, and his views on the current state and future opportunities in scientific publishing.
Let’s start at the beginning: how did you first become interested in biology?
There was never a moment of epiphany and I didn’t have a well thought out plan. At school I enjoyed science and did well at it, but I was not aware that a career in academic research could be a possibility. Around the time I was thinking about going to university, I watched the film Life Story, based on Jim Watson’s book The Double Helix, and read Richard Dawkins’ The Blind Watchmaker. I think both of these tipped me towards biology. I also remember reading Microbes and Man by John Postgate, which is less well known but a really superb book, and it’s probably at least partly a consequence of this that I ended up at Warwick University studying microbiology and virology.
During your PhD, you worked on interferon signalling in cell culture. What attracted you to that field, and what then prompted you to move into developmental neurobiology for your postdoc?
During my undergraduate degree, I became fascinated by how the study of microbiology and virology had taught us fundamental aspects of eukaryote molecular biology. This led to an interest in interferons – the secreted cytokines that are the major cellular response to viral infection – and I joined Ian Kerr’s lab at the Imperial Cancer Research Fund (ICRF; which later became Cancer Research UK and then merged to form part of The Francis Crick Institute) to study the mechanism of interferon signalling. By chance my timing was perfect, as we were identifying what became known as the Jak-STAT signalling pathway. Our neighbouring labs were also investigating signal transduction and elucidating pathways such as MAP kinase, RAS, PKA and more. I became very interested in understanding how cells perceive and respond to extracellular signals. Most of the groups at ICRF used cell lines but there were a few developmental biology labs, including those of David Ish-Horowitz, Julian Lewis and Phil Ingham. I went to their seminars and this introduced me to the beauty of embryos and to hedgehog signalling. I decided I wanted to continue studying signal transduction but to learn developmental biology so that I could investigate signalling in its natural environment rather than just in cell culture.
You joined Tom Jessell’s lab at what must have been an exciting time: sonic hedgehog (Shh) had only recently been cloned and it was known to be involved in spinal cord patterning, but little was understood about how the system worked. What were the questions you set out to answer during your time there?
Again, I was lucky with my timing. As you say, it was a really exciting time to be in the Jessell lab and at Columbia University. I wanted to understand Shh signalling and how it could function in a graded manner to control cellular responses. Very little was known about the molecular mechanism of signal transduction – I think even today we still have an embarrassingly poor understanding of this pathway – and I thought we could figure this out using the neural tube. Being in the Jessell lab, however, I soon became fascinated with and distracted by broader aspects of developmental neurobiology and how the spinal cord forms and functions. It became apparent that understanding the role of Shh in the spinal cord required studying the gene regulatory network that it controls and this has become the passion I’ve pursued ever since.
So what are the problems you’re working on now?
Broadly, they’re still the same problems that I was interested in when I was in Tom’s lab. For me, the fundamental question in developmental biology is how the right cells are produced in the right place, at the right time and in the right amounts in a developing tissue. The spinal cord turns out to be a fantastic system to address this and it’s revealing general principles that are applicable to many, if not most, developing tissues. Addressing these issues covers some of the most basic questions in biology. How is gene activity controlled? How is cell function determined? How are tissues shaped and organised from cells? Over the last couple of decades, developmental genetics has identified at least some of the key players in these processes. Now, new approaches and technology, from imaging to genomics to genome engineering, are providing unprecedented insight and resolution. I’ve been keen to move beyond purely qualitative explanations to a more dynamic and quantitative understanding of how the neural tube is formed and patterned. We’re trying to bridge scales from molecules to cells to tissues, to explain how the cells make the key decisions and how this guides the assembly of a functional, well-organised neural tube. I think this is of fundamental interest, but it also has practical implications for understanding disease and for progress in regenerative medicine and tissue engineering.
You’ve embraced mathematical modelling of developmental systems: why do you think this is important, and what are the challenges involved in trying to model something as complex as the developing spinal cord?
This was a decision I made about 10-12 years ago and, in part, it is because of the complexity of the problems we were investigating that I thought mathematical modelling was important. We were analysing increasingly complicated gene regulatory networks and we were trying to get away from simplistic reductionist descriptions to find explanations of how the system functioned as a whole. It’s often difficult to form an intuitive understanding of even relatively small systems if there are multiple interactions and feedback – it’s very easy to fool or confuse yourself. Mathematical modelling provides a rigorous way to describe and investigate a system. It tests whether your assumptions and interpretations are compatible. And different kinds of modelling can be useful at different levels of organisation, from the molecular to the tissue level, allowing us to look at the problem at different scales. I’ve been fortunate to have had great collaborators over the years that have taught me a lot of maths and physics. One thing I’ve learnt from this is that developing productive interdisciplinary collaborations takes a long time and requires a lot of trust and patience, but the investment can be very rewarding. We now think of mathematical modelling as just one of the available techniques that we use to tackle a problem, alongside more conventional molecular and cellular experiments.
More broadly, how important do you think it is for today’s young scientists to think interdisciplinarily?
Perhaps because it operates across multiple scales (both spatially and temporally), I think developmental biology has always been interdisciplinary. The molecular genetics revolution that transformed our field over the last 30-40 years is just one example of this. The importance of genetics is now taken for granted but it was once seen as pioneering and innovative. Today, it’s increasingly recognised that biologists need to be more quantitative and computationally literate and many of the applicants we see at The Francis Crick Institute have these skills. Nevertheless, it is something that needs to be strengthened through training at undergraduate and graduate levels. Having said that, one of the pleasures of academic research is that you’re continually learning and challenging yourself, so there are always opportunities to fill any gaps in your knowledge and learn something new. I often participate in advanced study courses run by organisations such as the Marine Biological Laboratory at Woods Hole and the Kavli Institute for Theoretical Physics at UC Santa Barbara. Even though I’m nominally ‘faculty’ on these courses, I usually end up learning at least as much as the students.
Developing productive interdisciplinary collaborations takes a long time and requires a lot of trust and patience, but the investment can be very rewarding
You recently moved into The Francis Crick Institute, which formed through the merger of NIMR at Mill Hill (where your lab was based), Cancer Research UK Lincoln’s Inn Fields and a number of other partners. How has the move been?
The planning took a long time, but the move itself went much more smoothly than I had anticipated. It’s been less than 18 months since we moved but it feels much longer; I guess that means we’ve settled in well. While I miss the charm and familiarity of our old institute, the new building more than makes up for it. The merger of the two institutes and our new partners mean I have new colleagues, which is stimulating and invigorating. I also love being in the centre of London – both because of the proximity to other academic institutions, and because we’re in a very vibrant part of town just minutes from the West End.
You’re active on Twitter: where do you see the value in social media for science and scientists?
I firmly believe that communication is a central part of science and academia: knowledge that isn’t passed on is wasted. Although not everything that appears on Twitter necessarily contributes to human progress, I enjoy being involved and I find it very useful. I get a lot of scientific information from Twitter. Particularly in fast-moving fields; for example, when CRISPR/Cas9 was coming on to the scene, I found Twitter was a great way to stay up to date and hear the latest developments. Also, like many scientists, I have friends and colleagues all over the world and Twitter is an easy way to stay in touch as well as to share and discuss (albeit briefly) papers and ideas.
You’ll be taking over as Development’s Editor-in-Chief in September, but you’ve been associated with our publisher, The Company of Biologists, for many years. Can you tell us a bit about your role as Director on The Company’s board and why you chose to get involved?
Yes, I’ve been involved a long time – since 2004. The Company of Biologists is a not-for-profit scientific publisher that publishes five journals: Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open. But unlike commercial publishers such as Elsevier and Springer Nature, we put the money that we make back into the scientific community. We give away about £1 million per year across our range of charitable activities: funding scientific societies such as the British Society for Developmental Biology (BSDB), sponsoring conferences, running workshops, and promoting research collaboration through travelling fellowships. We also support non-profitable, community-focussed activities such as the Node and preLights. There are 17 Directors of The Company and we’re almost all active scientists. We aren’t directly involved in the day-to-day business but we are responsible for the major strategic decisions, such as deciding to launch a new journal or start a big new project. In my role as Director, I was very involved in setting up the Workshops programme, and I’ve chaired the Grants Committee for the last few years, which decides how we spend our charitable funds. I’ve really enjoyed being on the board: it’s been a great opportunity to support the scientific community and I hope we’ve done at least a little bit of good.
Remember, by submitting your paper to Development or reviewing a paper for us, you’re supporting not only the journal but also all our other charitable activities
Scientific publishing is going through some significant changes at the moment, with the open access movement, developments in online publishing technology, the rise of preprints and so on. What do you think the future holds for small not-for-profit publishers like The Company of Biologists?
It’s definitely an exciting time in the publishing world. While all the changes create a lot of uncertainty both for people working in publishing and for scientists, I think there are lots of opportunities for small publishers such as The Company of Biologists. One of our advantages is that we’re run by scientists and we can respond in ways that help scientists and science and we’re not driven by profit margins. We’re also small enough and bold enough to experiment occasionally. Eight or nine years ago, we saw the increasing use of social media in our labs and heard calls from the community for an online discussion forum, so we launched the Node as a way to encourage informal discussion and communication about science within the developmental biology community. This has continued to grow in popularity. Just last month, we launched another experiment – preLights, the preprint highlighting and commenting service. This was in response to the growing numbers of preprints and it will be interesting to see how this develops in the coming months and years. More broadly, The Company’s goal is to support and inspire our fields and the people working in them and these aims will continue to drive our thinking and any future innovations.
Fundamentally, though, the journals and the research papers we publish remain at the heart of The Company of Biologists and it’s crucial that we all continue to read, write and referee for Development, if we are to continue to benefit from all the other initiatives and charitable support that The Company offers. Remember, by submitting your paper to Development or reviewing a paper for us, you’re supporting not only the journal but also all our other charitable activities.
Given the huge volume of published research, it’s increasingly difficult to keep up with the scientific literature. How much do you read and how do you choose what you read?
I agree it can be daunting keeping up with literature. I have a routine where I read a paper first thing in the morning when I get in to work (often I’ll choose it the night before). So I read at least one paper every day, although usually I’ll also read or review an article during the day. I try to read things outside my own research area, as this can provide ideas for my own work. I often pick up paper or preprint suggestions from people I follow on Twitter and I expect I will increasingly use preLights to get recommendations. I also think this is where community journals such as Development play a crucial role. Every issue of Development has papers handled by academic editors who are leaders in the field, so it offers a curated collection of the latest developmental biology research selected by experts. I always browse through the list of newly published papers and I often find myself picking one or two to read – it might be on neural development but as often as not it’s a Drosophila paper, an evo-devo study or even sometimes a plant paper.
And how would you like to see the journal (Development) evolve under your editorship?
I’m not taking over until late September 2018, so although I have lots of ideas I’m still refining my plans and taking suggestions. Olivier has done a great job during his time as Editor-in-Chief. He’s strengthened the journal, incorporating and encouraging new areas of research, and I certainly want to keep building on his accomplishments as developmental biology continues to change and grow over the next few years. I’m also very keen that Development continues to innovate and support our community. I’d welcome suggestions from readers, authors and referees about what we should do. What areas of science should we be encouraging in Development? What more can Development do to help our field? I’d urge anyone with thoughts or ideas to contact me.
Finally, is there anything that Development readers would be surprised to find out about you?
Some people find it surprising that I’m really a country boy at heart. Despite having lived most of my adult life in big global cities – London and New York – I was brought up in a 300-year-old thatched cottage on the Sussex Downs. I spent most of my teens working in stables and riding horses. We always kept lots of animals. As well as cats, dogs and rabbits we also had hens, ducks and goats; as a consequence, I can milk a goat.
The Department of Pediatrics at the University of Colorado School of Medicine invites applications for a faculty position in the tenure track. Appointment is expected at the Associate Professor level, but more senior individuals may apply. Applicants must have a Ph.D. and/or M.D. degree and demonstrated excellence in research.
We are particularly interested in individuals studying cardiac cell diversity, cardiovascular cell lineages and embryonic origins, gene regulatory networks and systems biology, cardiac cell regeneration and reprogramming, cardiovascular matrix biology and developmental models of cardiac biology. Applicants should have a record of creative and cutting-edge research, consistent and impactful publication and external funding. Applicants also should have a strong commitment to graduate and medical education and a successful record of trainee mentorship. Individuals who can enhance the diversity and accomplishment of our campus academic community are especially encouraged to apply.
Dr. Bruce Appel, Head of the Section of Developmental Biology, is Chair of the search committee. Questions can be sent to bruce.appel@ucdenver.edu.
The Neoproterozoic Earth System and the rise of biological complexity Thematic Project (FAPESP 2016/06114-6) directed by Prof. Ricardo Trindade at the Instituto de Astronomia, Geofisica e Ciências Atmosféricas (IAG-USP) and in collaboration with A. Morandini, M. Custodio, and F. Brown at the Instituto de Biociências (IB-USP), and D. Galante at the Laboratório Nacional de Luz Síncroton (LNLS) is recruiting a postdoctoral researcher with experience in physiology, cell biology, and/or developmental biology. We know little about the relative roles of environmental and biological factors involved in the late Neoproterozoic events leading to the ultimate oxygenation of the ocean-atmosphere system and dawn of biological complexity. We intend to fill these gaps by integrating information about the chemistry of oceans, evolution of complex life, paleogeography and tectonics between the Cryogenian and the early Cambrian. The selected postdoc will examine the physiology, ecology and developmental behavior of several groups of metazoans to test hypothesized connections between changes in ocean redox, nutrients and the evolution of life.
The main objectives of the postdoctoral project include:
(1) To study the tolerance of modern metazoans to Neoproterozoic ocean chemistry. The postdoc will examine tolerance to extreme oxygen level, as well as other Neoproterozoic environmental conditions, in several species (e.g. sponges, cnidarians, acoels, platyhelminthes, nematodes, and tunicates) to evaluate conserved adaptive physiological or phenotypic responses.
(2) To search for conserved metabolic pathways in species adapted to distinct oxygen levels. Using a comparative genomics approach, the postdoc will search for ancestral vs. derived gene pathways involved in oxygen metabolism. He/she will evaluate selection on oxygen metabolic pathway genes.
(3) To carry artificial selection experiments in C. elegans. To what extent can extreme oxygen variations alone drive the evolution of novel phenotypes that originated early Metazoans? We will evaluate the evolution of morphological and phenotypic complexity, and also evaluate alterations or changes in tolerance of phenotypically plastic physiological, developmental, or behavioral responses during the life cycle of C. elegans, an model animal with extensive understanding of the genetic, cellular, and developmental processes that generate phenotypes.
The candidate will be mainly based at the IB-USP in São Paulo to work with cultures of live animals, and at the LNLS in Campinas to use the space environment simulation chamber that will be used to recreate Neoproterozoic Earth conditions. The research team maintains an international working environment, speaking Portuguese is not required but it would be advantageous.
It is ESSENTIAL that:
-the candidate has a doctoral degree in Biological Sciences or related fields
-the candidate has experience in any of the following fields (the more the better): physiology, cell biology, developmental biology, evolutionary biology, astrobiology or bioinformatics.
-the candidate can coordinate a highly collaborative and integrative research project
-the candidate is willing to co-supervise students together with the PIs involved in the project
-the candidate has excellent communication skills in spoken/written English.
The applicant should contact directly to Dr. Federico Brown (fdbrown@usp.br), Dr. André Morandini (acmorand@usp.br) or Dr. Marcio R. Custódio (mcust@usp.br), and provide a letter of interest, a CV, and contact information of three potential referees until May 15th. Start date is any time after August 2018.
Often, the most enjoyable research projects are the ones that were never planned; and so it was with the current study.
In developmental biology, genetic lineage tracing methods have become a standard tool to interrogate the lineage potential and fate behaviour of cells [1]. By activating fluorescent reporter genes in individual cells, qualitative and quantitative methods have been defined to translate the size distributions of marked cells and their progeny – clones – into information on the statistical fate behaviours of constituent cells [2]. These approaches have led to important insights into the dynamics and functional behaviour of stem cells and progenitors during the development, maintenance and regeneration of tissues, as well as the dynamics of tumour growth during cancer initiation and progression.
Unlike continuous in vivo live-imaging approaches [3], the interpretation lineage tracing data, based on the analysis of fixed samples, can be confounded by uncertainties in the history of clonal evolution: First, there is a question of whether the tracing assay targets a single (functionally equivalent) cell population, or whether there is heterogeneity within the induced cell fraction. Second, there is an intrinsic ambiguity in the lineage reconstruction since multicellular clones can typically be generated from a multitude of potential lineage trajectories: Different combinations and permutations of cell divisions leading to symmetric and asymmetric fate outcomes can generate clones of the same size and cell composition. Fortunately, to a degree, both problems can be mitigated post hoc by the nature of the clonal dynamics itself!
Figure 1. Surface of a mouse heart at embryonic day (E)12.5 showing clusters of cells derived from cell precursors marked by a multicolour fluorescent (confetti) reporter system under the control of a Mesp1-promoter at E6.5. For details, see the primary paper. Looking at the image, can you tell which clusters belong to the same clone…?
Previously, research by our group and others have shown that, while the short-term dynamics of labelled progenitors may be highly complex, in the long-term, the size distribution of clones anchored in equipotent cell populations typically converge to rigid “scaling” behaviours in which the chance of finding a clone larger than some multiple of the ensemble average becomes unchanging over time [4]. Indeed, in self-renewing adult tissues, the constraint of homeostasis limits strongly the variety of scaling distributions, so that information on cell fate behaviour can be recovered “rigorously” from the functional form of the scaling distribution.
However, the utility of these methods is heavily reliant on the integrity of clonal assignments; and therein lies the rub! During development, and in the course of adult tissue turnover, cell rearrangements due to active cell migration, or serial rounds of cell loss and replacement, can lead to the fragmentation or merger of clones, corrupting the data and, potentially, misleading efforts to interpret the underlying clonal dynamics. Yet, in a lineage tracing approach based on the analysis of fixed samples, we can’t know what were the individual histories of clones and their fragments. Developments of multicolour lineage tracing strategies, based on the “brainbow” or confetti reporter constructs, can mitigate the potential for such effects. However, in the context of tissue morphogenesis or tumour development, large-scale cell rearrangements caused by tissue remodelling can lead to the long-distance segregation of clonal fragments rendering the task of clonal reconstruction potentially infeasible. Worse, confronted with such challenges, many researchers may feel inclined to abandon attempts to reconstruct putative clones dispersed across the three-dimensions of a solid tissue and instead to characterize the size of a clone by its intersection with a thin section of tissue – “surely a rough indication of the complete clone size…?”
Unfortunately, such hopes are dashed by the inconvenience of a mathematical riddle – the corpuscle problem – which states that, without making detailed assumptions, the distribution of three-dimensional objects cannot be recovered (even in principle) from the statistics of their random intersections [5]. Yet, for those brave enough to try to reconstruct the entirety of the clone, the situation is still treacherous since, at first sight, it is difficult to know whether the clonal assignment is faithful. With the increasing prevalence of genetic lineage tracing assays, this is the situation that prevails – with legions of researchers (and reviewers…!) questioning whether assays are clonal. Of course, statistical analyses can help. At low enough clonal induction frequency, fragmented clusters of cells can be associated with defined statistical confidence. Yet, in a developmental context, this may require organs with as few one or two clones per animal, a daunting, expensive and arguably unethical endeavour! So, can anything be done to alleviate this problem?
With the benefit of hindsight, we should have asked ourselves whether, in common with “true” clonal dynamics, the statistical distribution of “putative” clones could, in of itself, provide evidence that the source data was corrupted by clonal fragmentation and/or merger events. But we didn’t! Instead, we stumbled into this idea through an intriguing empirical observation. Our collaborators in the Blanpain lab at the Interdisciplinary Research Institute at the Université Libre de Bruxelles were interested in resolving the potency and proliferative potential of early Mesp1-expressing precursors in the developing mouse heart. To trace the fate of these cells, confetti labelling was induced in a fraction of Mesp1+ cells early in embryonic development, and the size and regionalization of clonal footprints were scored on the developing mouse heart (Fig. 1) [6]. Notwithstanding the corpuscle problem, we (recklessly) constructed the size distribution of clonal “fragments” on the embryonic heart and found a size dependence that was strikingly broad yet was almost void of structure (i.e. information on the underlying clonal dynamics): The average sizes of labelled fragments differed vastly between different heart compartments and time points in development – consistent with the complexity of developmental programmes during heart morphogenesis. However, to our surprise, once we divided fragment sizes by their ensemble average, the resulting rescaled probability distributions became indistinguishable between different heart compartments (Fig. 2). What could it mean…? Did this reflect a rigid growth characteristic of precursors in the maturing mouse heart – with some cells destined to proliferate and expand prodigiously while others are set to exit cycle early? Or was there a less “glamorous” explanation, at least from the perspective of our experimental collaborators…?
Of course, during development, clones are subject to numerous intrinsic and extrinsic influences that adjust cluster size: Labelled cell clusters can expand through the division of constituent cells; they can also expand through chance merger events that bring together clusters of the same hue. However, clusters can also diminish in size through cell loss – either through cell death, or by cells leaving the field of view in a sectional characterisation of cluster size; they can also diminish through fragmentation events caused by collective cell rearrangements created by larger-scale morphogenic changes or simply from stochastic forces exerted by the surrounding cells. Finally, new clusters can appear as labelled cells that are “out of frame” move into a sectional view. In short, in a model that mirrors both qualitatively and quantitatively the non-equilibrium dynamics of aerosol suspensions, the time-evolution of the distribution of labelled cell clusters conforms to a fiendishly complex set of possible processes that includes all the factors above, and potentially many more – a seemingly unpromising starting point for any analysis…
However, the rigour of statistical physics (and a little bit of hindsight) came to the rescue!! Indeed, during tissue development or turnover, constituent labelled cell clusters will stochastically expand and contract in response to the factors above. Under these conditions, the different possible influences impact on the resulting cluster size distribution to widely varying degree. Therefore, to quantify the relative contributions of these disparate processes, we employed a concept that has shaped the field of statistical physics: By progressively moving to coarser and coarser length scales, we obtained mathematical equations describing the varying contributions of different processes on these scales. We reasoned that, on the largest scales, i.e. as development proceeds, these equations then give an accurate, but reduced, description of the dynamics shaping the cluster size distributions. Notably, using an analytical approach inspired by the “renormalization group” methods of statistical physics, we could show that, over time, only the processes of clonal merger and fragmentation impact on the shape of the resulting cluster size distribution; in the parlance of statistical physics, these were the relevant operators. Crucially, if perhaps worryingly for the biologists(!), information encoded in developmental fate programmes becomes progressively erased from the cluster size record, while the emergent dynamics signals only the existence of fragmentation and/or merger events. The observed cluster size dependence was, indeed, not a reflection of some underlying fate programme, but an inevitable outcome of the collective dynamics of heart development. As a corollary, the erasure of biological information suggested that cluster size distributions obtained from lineage tracing experiments should take the same form across different biological tissues and even species. To drive home the message, we confirmed that the cluster size distributions of the developing zebrafish heart, as well as liver and pancreas, also conform to common – physicists would say “universal” – cluster size distributions with the same hallmark (in this case, log-normal) size dependence.
So far, so good, we thought; and we set off to inform the stem cell community of our findings. Not surprisingly, we found that not all experimentalists were equally enthusiastic to learn that the outcome of often challenging lineage tracing experiments might be, by design, void of biologically significant information… However, perhaps more surprisingly, we found that the very concept of universality – a rigorously defined mathematical concept at the heart of critical phenomena in statistical physics – caused additional irritation among our biological peers. More than once we were informed that “nothing in biology is universal…!” Indeed, this warning betrays something of the cultural differences that continue to divide the physical and biological science communities. While the former are trained to look for commonalities between different systems, the inherent complexity of biological systems makes it more natural to focus on differences and details.
Indeed, we believe that these observations have value both to the biology and physics community. For the former, these findings have a practical value: Although one cannot use statistical distributions to confirm positively the integrity of clonal assignments, one can at least use hallmark scaling dependences of cluster size to quantitatively and, therefore, rigorously signal when seemingly clonal data has been corrupted by fragmentation and/or merger events. At the same time, understanding the origins of universal scaling behaviour allows biologists to develop experimental strategies that can unveil the developmental programmes of labelled cells. For the physicists, the situation is more encouraging with clonal dynamics presenting yet another example of where biological systems can provide an arena in which powerful concepts from statistical physics and mathematics can find novel and practical application.
Figure 2. Cumulative distributions of fragment sizes from different tissues and species (dots) collapse onto the same universal scaling form (dashed line) when rescaled by the average size. For details, see primary paper.
In these lines I share with you some details of our recently published Nature paper. I will comment how this project was started and details which are not included in the manuscript. Finally, I will briefly comment on some questions we are working on today and others we believe are worth addressing in the future.
Historical context of the project.
One of the questions that has enthralled biologists for many years is how a single cell can give rise to highly organised multicellular organisms with extremely complex forms. Although finding an answer to this question represents a big challenge, the advances made by the cell and developmental biology community over the last 50 years have been truly astounding. The advent of molecular biology in the last century, allowed embryologists to generate a strong body of evidence whereby ‘old’ ideas based upon cellular observations have found their molecular explanation. One of the most astonishing examples of this type of molecular re-description of a cellular process is the molecular characterisation of the Spemann–Mangold organiser. This tissue was initially described in 1924 as an embryonic region exerting inductive effects on cells (1). More than 70 years later, Edward De Robertis’ group generated cDNA libraries from manually dissected organisers and found genes that were specifically expressed in this territory (i.e., goosecoid (gsc)) (2). This was the first demonstration that this structure was a molecular identity defined by specific gene expression patterns. Currently, most of the embryonic tissues and about 4 organiser regions have been molecularly defined across embryos of different species (3,4). Indeed, this field has progressed to such a level that well-defined networks of signalling pathways are now widely accepted as coordinators of morphogenesis, and that their most common modus operandi is the delivery of positional information allowing interaction among neighbour cells (3,4). However, despite such vast advancements in our understanding of molecular players during morphogenesis, the role of other types of cues in this process is comparatively less understood.
Morphogenesis is a highly regulated biological process which involves constant rearrangement of tissues and deep mechanical changes. However, the role of these mechanical changes is only just re-starting to be considered when looking for a more comprehensive explanation of the morphogenetic process. I use the term ‘re-starting’ because in the early 1900’s, and prior to the molecular ’boom’, scientists studied embryonic development by using theoretical, morphological, chemical, and mechanical approaches. More recently, and ‘swimming against the tide’, several groups have focussed their research on developmental mechanics as a means of understanding those aspects of morphogenesis that molecular patterning cannot explain by itself. Work from these contemporary groups has been nicely reviewed in Lecuit, 2008 and Gilmour et al., 2016 (5,6). Considering the strong molecular frame-work and new advances in tissue mechanics, the field is starting to look at embryos as finely regulated machines where tissue rearrangements lead to mechanical changes that, in turn, feed back into the cells to modify their gene expression and behaviour, ultimately coordinating morphogenesis.
In this context, and taking advantage of powerful mechanical and molecular tools, we revealed that tissue stiffening owing to cellular rearrangements occuring in the mesoderm during Xenopus gastrulation triggers the epithelial-mesenchymal transition (EMT) and collective migration of the neural crest (NC) at neurula stages. More generally, we showed that tissue mechanics act as long-range cues coordinating the timing of two developmental processes that, until now, were considered as unconnected events (7).
Early steps of the project
The initial question addressed in this project was: what triggers the migration of the neural crest? The neural crest forms by inductive signals at the border of the neural plate (induction) where it remains as a non-migratory population until, at some point and for some reason, they undergo EMT, delaminate, and collectively migrate toward their target tissues (7,8).
The conception of this project occurred towards the end of my PhD in 2014. I was working on the transcriptional regulation of Twist, an EMT regulator in the neural crest (9). A persistent question at the time was why Twist and most of the neural crest EMT-related transcription factors are expressed at induction stages, 12 hours before the EMT and migration take place. At the same time, I was due to start a postdoc, requiring me to design and write a project. By 2012–14 there was a wave of in vitro studies showing that stiffer substrates favour migration and EMT in several cell types (10) but whether this was the case in vivo, was lacking a demonstration. That is how, with Roberto, we began to speculate that mechanics of the tissues surrounding the neural crest may change during development and that, consequently, the neural crest could feel this change and migrate. I started to read about the topic and I was impressed by the vast morphological changes that Xenopus embryos undergo in these 12 hours prior to neural crest migration, during which embryos pass from a spherical to a rather more elongated shape due to convergent extension (11,12). I even found an article showing that paraxial mesoderm stiffens towards the onset of NC migration (13). Many questions emerged at the time i.e.: Does the environment play a role in NC migration? Does NC actually reply to mechanical cues from its substrate? What are the mechanical properties of the NC microenvironment? How are these mechanical properties regulated? So, we put these questions down in a project, got the funding, and start working on this story. Not as simple as it sounds; I had to apply twice for funding, but I was finally awarded an EMBO and a Marie Curie postdoc fellowship.
Main findings
We can divide our story in two parts. First, we studied the mechanical interaction of the NC with its surroundings and the role of the temporal environment in controlling NC migration. Second, we sought to understand the mechanisms underlying mesoderm stiffening.
Before discussing the results, I would like to clarify the nomenclature we use for the different stages in which we analysed the NC. Non-migratory NC are defined as, even if it sounds obvious, cells that are not migrating and that instead resemble a more epithelial phenotype. These are normally analysed between stages 12–14 in our work. Pre-migratory NC are motile but a migratory stream is not yet observable, and we analysed them at stage 17–20. Finally, migratory NC, which are defined when migratory streams are evident, are typically analysed between stages 22–25 in our article. Stages as in Nieuwkoop and Faber (14).
I was forgetting this! When I talk about stiffness throughout the text, I am referring to the apparent elasticity of the tissue or the apparent elastic moduli contained in it, if you prefer.
Mesoderm stiffening is necessary and sufficient to trigger NC collective migration in vivo
As most of the molecules required for NC migration are already expressed at non-migratory stages, we asked whether non-migratory and migratory NC had the same migratory potential. To address this, we explanted non-migratory and migratory neural crests and evaluated their chemotactic response towards Sdf-1, a well-known NC chemoattractant (15). To our surprise, ‘non-migratory’, pre-migratory, and migratory neural crests equally migrated to SDF-1 indicating that, regardless of their stage, NC cells have the same migratory potential. Based on this in vitro observation we hypothesised that, in vivo, the temporal environment may be contributing to govern the migratory potential of the NC. Therefore, we performed heterochronic grafts, where non-migratory cells where grafted into migratory hosts and vice versa. It was nice to observe that non-migratory or migratory NC cells grafted into migrating hosts almost immediately start migrating. However non-migratory or migratory NC cells grafted into non-migratory hosts took more than 12 hours to start migrating, the same time taken by host embryos to reach migratory stages. These results strongly suggested that there is ‘something’ in the environment that changes from non-migratory to migratory stages and that this ‘something’ is very likely to be involved in the initiation of NC migration.
Personal note 1: experiments like these are highly motivating (at least for me). They do have this beautiful mixture of simplicity, creativity, and a touch of classic embryology… but with a high reward in terms of information.
As in vitro studies show that substrate stiffness triggers collective migration (10) we next characterised the mechanical environment of the NC (and yes, we did rule out other environmental factors as potential triggers of NC migration). Because the NC migrates sandwiched between the mesoderm and epidermis, we measured stiffness of these tissues in the region just in front of the NC at non-migratory, pre-migratory, and migratory stages. Our weapon of choice to measure tissue stiffness in vivo was a very novel and versatile setup of in vivo atomic force microscopy (iAFM). This method was developed by one of our co-authors, Kristian Franze (University of Cambridge) (16). After adapting this setup to our experimental requirements, we discovered that the mesoderm, used by the neural crest as a substrate, stiffens by 3 times from non-migratory to migratory stages (Figure 1). Our controls revealed that the epidermis has either similar or lower stiffness than the mesoderm. Until here, our results showed that there is a strong correlation between mesoderm stiffening and neural crest migration.
Figure 1. Mesoderm stiffens towards the onset of neural crest migration.A Schematic showing the region where the measurements were taken. NP, neural plate; NC, neural crest; ep, epidermis; NT, notochord; msd, mesoderm. B Chart summarising the spread of data of our iAFM results in two of the analysed stages. Each dot is an AFM indentation, data was analysed with Mann-Whitney test.
Based on this, our next question was whether this correlation plays a functional role in NC migration. We addressed this question by developing an ex vivo system consisting of 2D hydrogels containing stiffness values equivalent to those obtained in embryos at migratory (stiff) and non-migratory (soft) stages. NC plated on stiff substrates chemotax towards Sdf-1, disperse, and undergo a switch of E- to N-cadherin (EMT). However, these parameters were drastically reduced in cells plated on soft substrates. This was a nice control that showed us that cranial neural crest replies to changes in substrate mechanics.
As we learned that mesoderm stiffens and that NC behave differentially on substrates of varying stiffnesses, we tested the relevance of mesoderm stiffening for NC migration in vivo. We used a combination of ablation experiments and targeted injections of myosin contractility inhibitors to release tension and reduce stiffness in the migratory substrate of the neural crest. We controlled that our treatments effectively reduced stiffness by using iAFM. In all our conditions, NC migration was analysed by in situ hybridisation. Our results were very clear and we observed that migration of the NC was inhibited when the stiffness of the substrate was reduced by tissue ablation or targeted injection experiments (Figure 2 A–C). These results indicated that mesoderm stiffening is necessary to promote migration of the neural crest in a non-autonomous manner.
Figure 2. Mesoderm stiffening is necessary and sufficient to promote neural crest migration in vivo.A Representative images showing lateral views of embryos hybridised with a probe against a NC marker. Treatments as indicated. B Chart summarising the spread of data of our iAFM results. Each dot is an AFM indentation. C normalised NC migration. D Representative images showing lateral views of embryos hybridised with a probe against a NC marker. Treatments as indicated. E Tissue elasticity plotted as a function of the AFM indentation. F chart showing normalised NC migration.
We next asked whether substrate stiffening was also sufficient to trigger migration of the NC in vivo. For this, we used a novel ‘strain stiffening’ assay to promote early stiffening of the tissue in front of the NC. This is an AFM-based method in which we can exert a controlled force for a determined amount of time in a precise location (16). As a result, we observed that applying extrinsic stress was sufficient to stiffen the mesoderm and promote premature migration of the NC (Figure 2 D–F). This was a very surprising result and it represented one of the first demonstrations of premature migration of the NC. Whilst former studies showed delamination and ectopic migration of delaminated single NC cells upon cadherin inhibition, the premature migration of the NC by following its stereotypical migratory paths had not been previously shown.
Personal note 2: designing and performing these experiments was a great experience for me. It was something not related at all to my biological background and it is one of the major sources of new knowledge from my postdoc. I have no doubt that around this time I changed the way I address biological questions.
Taking together this set of results we concluded that stiffening of the mesoderm is necessary and sufficient to promote NC migration in vivo, very likely by triggering its EMT. We believe this is not only relevant for the NC. Numerous groups of cells and single cells are challenged by the mechanics of their surroundings when migrating in vivo. There are several types of mechanical challenges that cells need to sort in order to reach their target tissues. Hence it would be of great value to start exploring the mechanical interaction of other cellular systems with their surrounding tissues in diverse biological contexts.
PCP-mediated convergent extension is the driving force of mesodermal stiffening in vivo
Our next goal was to unveil the driving force of mesodermal stiffening. The first idea we explored was an accumulation of extra cellular matrix (ECM). In many systems, accumulation of ECM is one of the main components of tissue stiffening (17). It was disappointing to discover that in our system this could not be the case. We found out that i) there is no difference in ECM deposition among non-migraotry and migratory stages; ii) removing ECM under the NC did not affect mesoderm stiffness; and iii) collagen is not expressed at these stages in Xenopus.
Thus, we next turned our attention to actomyosin contractility, as it has been previously shown that this activity could play a role in paraxial mesoderm stiffening (13). We analysed expression of phosphorylated myosin and f-actin in the head lateral mesoderm (mesoderm used by the NC as a substrate). We observed no differences in expression levels or sub-cellular localisation when comparing non-migratory vs migratory stages. We even inhibited contractility once the stiffness was already increased in the head mesoderm. As measured by iAFM, only a very small and non-significant difference was observed when compared to control embryos, indicating that although it may play a role, contractility was not the main component of head mesoderm stiffness.
Personal note 3: These were very frustrating results at the time, I was running out of options and the explanation about what was the driving force of mesodermal stiffening seemed more complicated than what we thought. But if you ask me today I think that this complication is from where one of the most interesting parts of our discovery was originated; mechanical coordination among temporally unrelated embryonic tissues.
We did not give up and kept looking for options. At the time, Kristian’s lab was just publishing their work about differential cell density as the source of a stiffness gradient that guided the growth of the optic tract (16). We went back and looked into the anatomy of the embryo and realised that from non-migratory to migratory stages, embryos elongate and cells accumulate dorsally. We did a very simple sectioning experiment on wild-type embryos with colour-labelled mesoderm and NC. We quantified cell density in the mesoderm under the NC at non-migratory, pre-migratory, and migratory stages and observed a strong accumulation of cells and increased cell density that nicely correlated with mesoderm stiffening and onset of NC migration (Figure 3 A–C).
Figure 3. PCP-dependent CE is the driving force of the cell accumulation required for mesoderm stiffening. A Representative images showing transverse sections with labelled nuclei of neural crest (cyan) and mesoderm (magenta) cells. Stages as indicated. Dorsal to the top. B Chart showing the total number of nuclei under the neural crest. C Chart showing the density of nuclei at the area immediately under the NC. D Representative images showing lateral views of embryos hybridised with a probe against a NC marker. Treatments as indicated. E Chart summarising the spread of data of our iAFM results. Each dot is an AFM indentation. F chart showing normalised NC migration. Treatments as indicated.
To functionally test this correlation, we quantified cell density upon application of distinct treatments into the mesoderm without inducing a direct effect on NC migration. We found that treatments resulting in blocked NC migration correlated with low mesodermal cell density and low stiffness, whereas treatments promoting NC migration perfectly corresponded with high mesodermal cell densities and high stiffness values. This strongly suggested that one of the main components of mesodermal stiffness is the accumulation of mesodermal cells that lead to increased cell density.
This matched seamlessly with what we observed before and at this point we re-interpreted our myosin knockdown results, where early inhibition of myosin contractility in the mesoderm led to decreased stiffness and inhibited NC migration. It is well-known from John Wallingford’s lab work that polarised activity of myosin is required for mesodermal cell migration (12). Consequently, it makes sense to think that what our treatments were doing was inhibiting mesoderm cell migration which in consequence decreased cell density and stiffness under the NC.
As planar cell polarity (PCP) is a master regulator of convergent extension (CE) (11,12) we inhibited PCP in the mesoderm. As expected, mesoderm stiffness was decreased and NC migration inhibited (Figure 3 D–F). As depleting PCP has many biological consequences, this treatment represented an excellent scenario to test whether introducing extrinsic stress to stiffen the mesoderm of soft embryos was enough to promote NC migration. So, as a final experiment we inhibited PCP, applied extrinsic stress with iAFM and observed that NC migration was mechanically rescued (Figure 4). These results led us to conclude that the driving force behind the increased cell density that promotes mesoderm stiffening and NC migration are PCP-driven cellular rearrangements during CE, with a minor contribution of myosin contractility.
Personal note 4: Something which I believe is really worth mentioning is the role of myosin and ECM in mesoderm stiffening. There is a large body of evidence from excellent groups that demonstrate the requirement of Fibronectin deposition and myosin activity to ensure correct convergent extension movements. Hence myosin and Fibronectin do contribute to build-up mesodermal stiffening by allowing mesoderm migration and accumulation of cells under the NC, but they represent a minor contribution to stiffness once cells have already accumulated.
These results were very exciting as they demonstrated for the very first time that two embryonic processes that have been always studied as separate events, CE during gastrulation and NC migration, are connected via tissue mechanics. Our data is an example of how applying more integrative approaches to study morphogenesis can lead us to actually understand how the process organises itself– such an approach has the potential to deeply impact our advances in the study of ‘self-organising’ biological systems.
Figure 4. Ectopic compression rescues NC migration in soft embryos. A Diagram describing the extrinsic stress experiments used to stiffen the mesoderm after PCP inhibition. B Representative images showing lateral views of embryos hybridised with a probe against a NC marker. Treatments as indicated.
Open questions
Although we demonstrate in our article that integrin, vinculin and talin, well-known mechanosensors (18), are required for NC migration we are now actively studying the mechanism by which NC cells sense and translate mechanical stimuli in a cellular and eventually molecular response. Another question that we are addressing now is related to a potential role for durotaxis in vivo. Durotaxis is the process by which cells migrate from compliant to stiffer substrates. In this front, we are studying how stiffness gradients may emerge as a result of mechanical heterogeneities of embryonic tissues and whether these gradients can direct cell migration in vivo. In the future, it may be interesting to address whether substrate mechanics are just permissive cues allowing cells to exert traction and migrate or instead are a real source of signalling that ‘permit’ migration but also modify the genetic program of the NC. Similarly, a question that may be linked to environmental mechanics and that is relevant for development, regeneration, and cancer is how cells know when to stop migrating and differentiate once they reach their target tissues.
Acknowledgements
I would like to thank the co-authors of this article Roberto Mayor, Guillaume Charras, and Kristian Franze for their input and the funding sources EMBO (LTF-971) and Marie Skłodowska Curie (IF-2014_ST VivoMechCollMigra). Dr Ailin Buzzi (KCL) and Emily Atalar (UCL) for their comments on this post.
References
1. Spemann H, & Mangold H. Induction of embryonic primordia by implantation of organizers from a different species (1924). Roux’s Arch. Entw. Mech. 100, 599–638.
2. Cho, K. W., et al., (1991). Molecular nature of Spemann’s organizer: The role of the Xenopus homeobox gene goosecoid. Cell, 67, 1111–1120.
3. Anderson, C. & Stern, C. D. (2016). Organizers in development. Curr. Top. Dev. Biol. 117, 435–454.
4. De Robertis EM (2006). Spemann’s organizer and self-regulation in amphibian embryos. Nat Rev Mol Cell Biol. 7, 296–302.
5. Thomas Lecuit. (2008). “Developmental mechanics”: cellular patterns controlled by adhesion, cortical tension and cell division. HFSP Journal 2, 72-78.
6. Gilmour, D., Rembold, M. & Leptin, M. From morphogen to morphogenesis and back. Nature 541, 311–320 (2017).
7. Barriga E.H., et al., (2018). ‘Tissue stiffening coordinates morphogenesis by triggering collective cell migration in vivo’. Nature 554, 523–527.
8. Betancur, P., Bronner-Fraser, M. & Sauka-Spengler, T (2010). Assembling neural crest regulatory circuits into a gene regulatory network. Annu. Rev. Cell Dev. Biol. 26, 581–603.
9. Barriga, E. H., et al., (2013). The hypoxia factor Hif-1α controls neural crest chemotaxis and epithelial to mesenchymal transition. J. Cell Biol. 201, 759–776.
10. Roca-Cusachs, P., Sunyer, R. & Trepat, X. Mechanical guidance of cell migration: lessons from chemotaxis. Curr. Opin. Cell Biol. 25, 543–549 (2013).
11. Wallingford J.B. & Harland R.M. (2001). Xenopus Dishevelled signaling regulates both neural and mesodermal convergent extension: parallel forces elongating the body axis. Development 128, 2581–92.
12. Shindo A. & Wallingford J.B. PCP and septins compartmentalize cortical actomyosin to direct collective cell movement. Science 343, 649–652 (2014).
13. Zhou J., Kim H. Y. & Davidson L. A. (2009). Actomyosin stiffens the vertebrate embryo during crucial stages of elongation and neural tube closure. Development 136, 677–688.
14. Nieuwkoop, P. D. & Faber, J. Normal Table of Xenopus laevis (Daudin): a Systematical and Chronological Survey of the Development from the Fertilized Egg till the End of Metamorphosis 2nd edn (North-Holland, 1967).
15. Theveneau, E. et al (2010). Collective chemotaxis requires contact-dependent cell polarity. Dev. Cell 19, 39–53.
16. Koser, D. E. et al (2016). Mechanosensing is critical for axon growth in the developing brain. Nat. Neurosci. 19, 1592–1598.
17. Moeendarbary, E. et al. (2017). The soft mechanical signature of glial scars in the central nervous system. Nat. Commun. 8, 14787.
18. Charras, G. & Sahai, E. (2014). Physical influences of the extracellular environment on cell migration. Nat. Rev. Mol. Cell Biol. 15, 813–824.

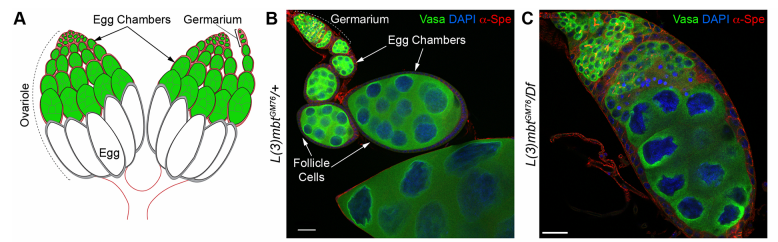
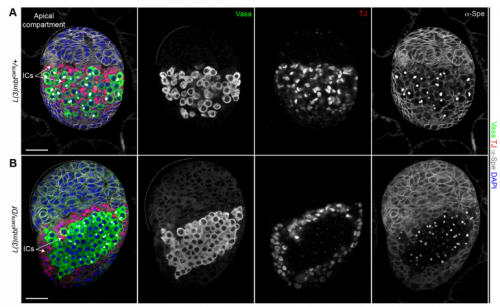
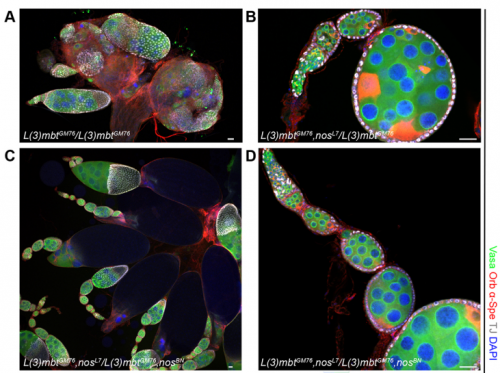
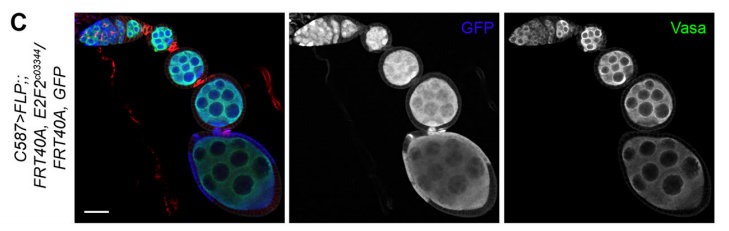
 (4 votes)
(4 votes)
 (No Ratings Yet)
(No Ratings Yet)

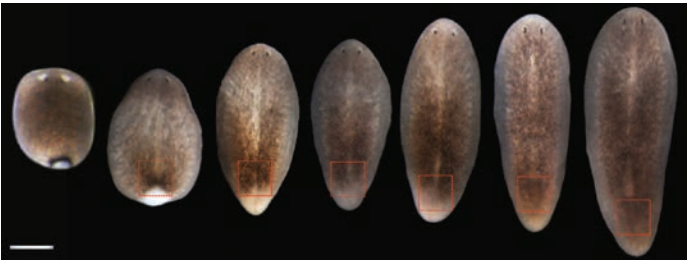
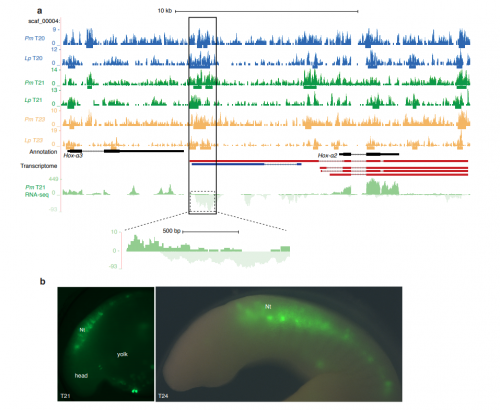
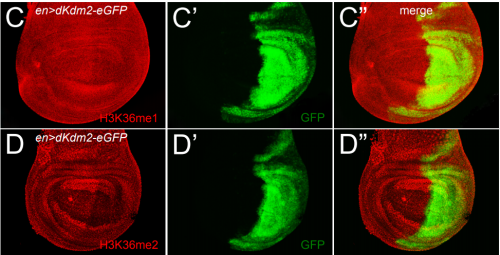
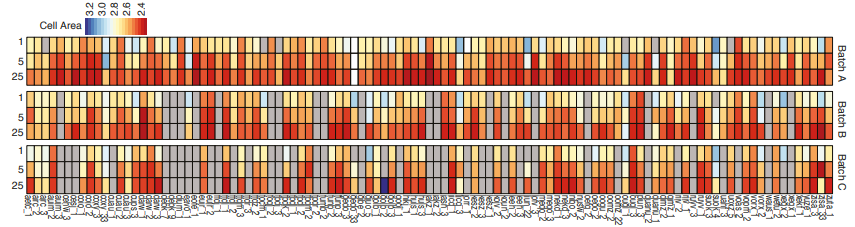

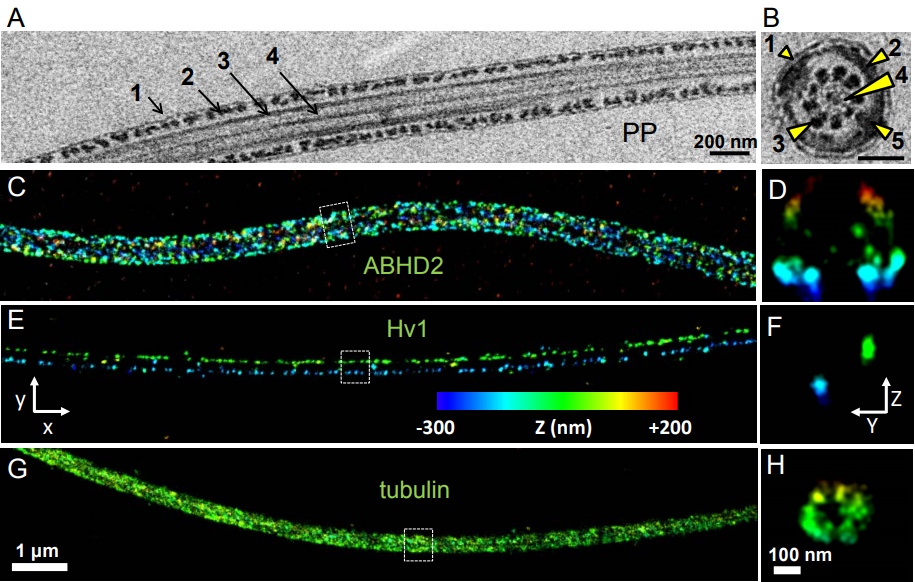


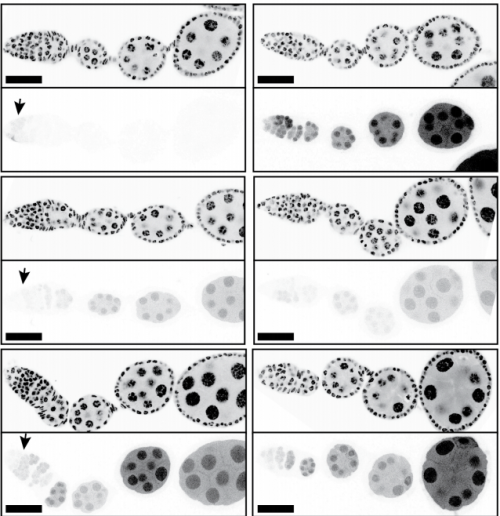
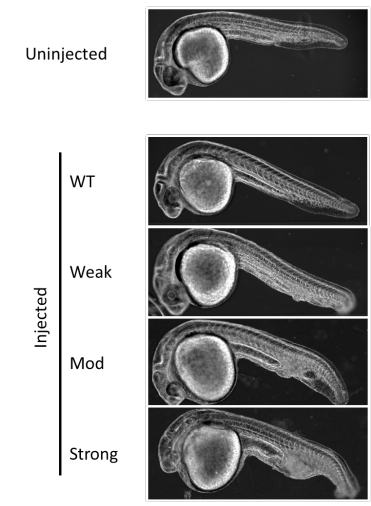

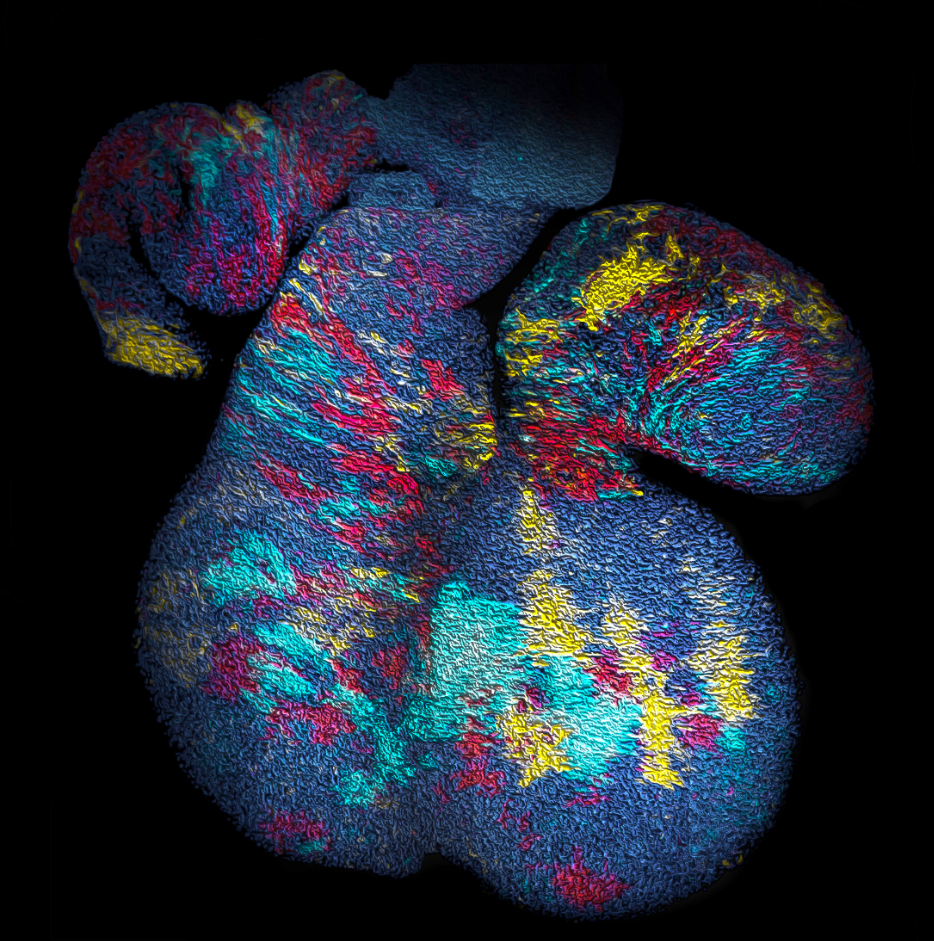
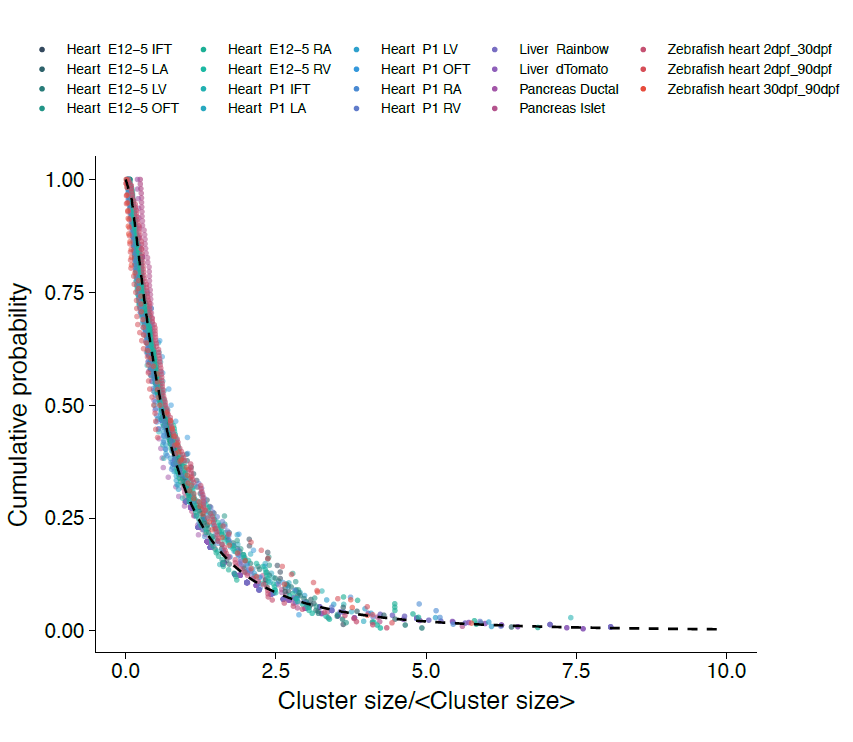
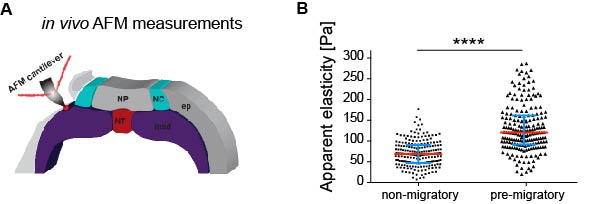
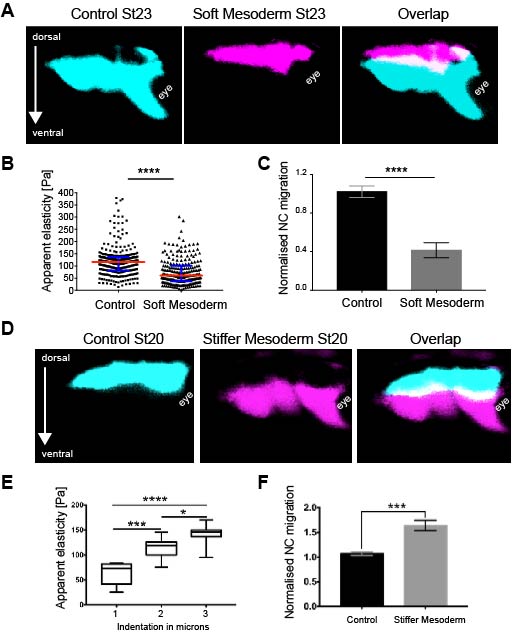
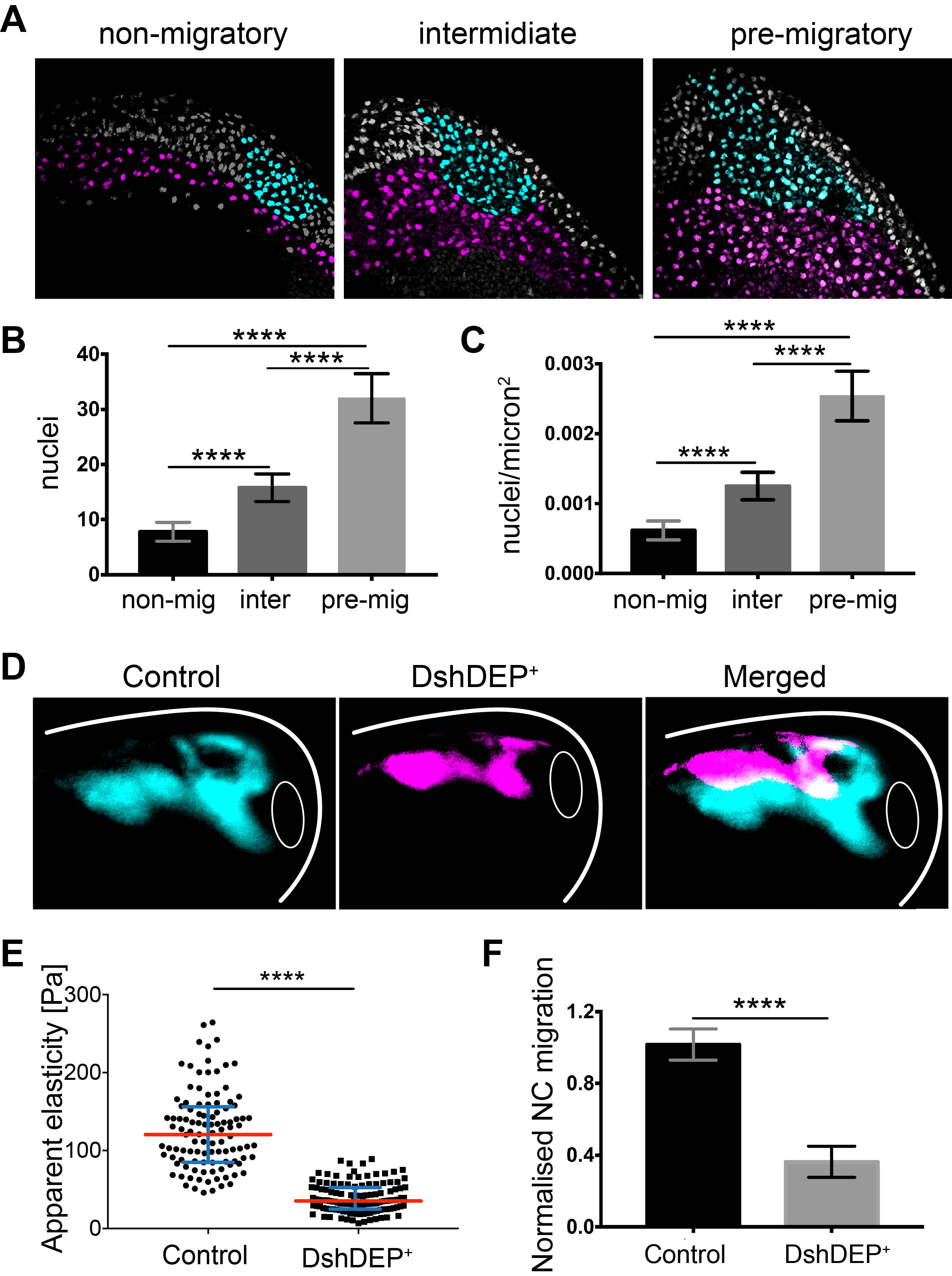
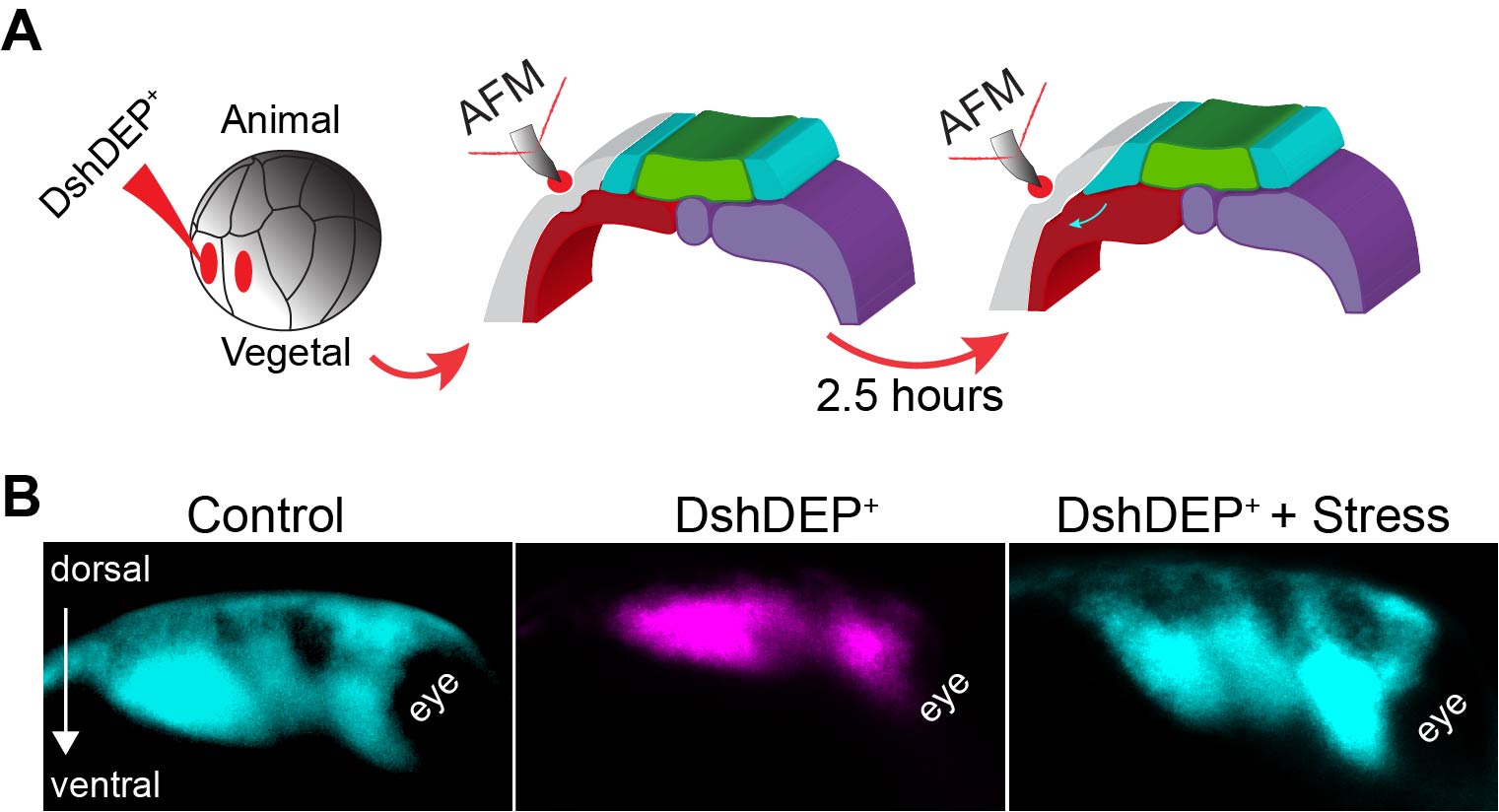
 (16 votes)
(16 votes)