Two Postdoctoral positions are immediately available at the University of California, San Francisco (UCSF), in the laboratory of Sarah Knox.
Position 1: The applicant will be part of an NIH-funded investigation into the development, regeneration and aging of exocrine organs including salivary glands, pancreas and ocular organs. The project specifically focuses on nerve-stem cell communication and how these interactions govern organ morphology, homeostasis and regeneration throughout the lifetime of the organism. In addition to defining key mechanisms regulating organogenesis, we pay particular attention to the impact of aging on organs and their potential for rejuvenation by employing parabiosis based methods in combination with 3D imaging, deep sequencing (scRNA and bulk RNA), and mouse genetics (including CRISPR generation of mouse models).
Position 2: The successful applicant will be part of an NIH-funded, pre-clinical investigation to develop a novel therapy that restores ocular and exocrine gland function through modulation of immune-nerve-epithelial cell interactions. The candidate will use genetic mouse models of autoimmune disease in conjunction with ex vivo and in vitro systems to study the effects of inflammation on epithelial cell-nerve interactions in the cornea and other ocular organs.
For both positions, applicants with a strong background in developmental biology, stem cell biology, neuroscience, mouse genetics and/or immunology are preferred. Technical expertise in the areas of mouse handling and dissection, immunoassays, immunohistochemistry, adoptive transfer, flow cytometry, qPCR, RNAseq, bioinformatics, and confocal microscopy are a plus. Applicants must possess an M.D., Ph.D., or equivalent degree. The candidate should have the proven ability to independently design and execute experiments, as well as interpret and publish results. They should also possess excellent communication skills and the ability to work as part of a team.
To apply, submit a C.V., a cover letter with a brief description of research interests and experience, and the names of at least 3 references via email: sarah.knox@ucsf.edu
The Landsman Lab invites highly motivated students to join its effort to decipher the role of the microenvironment in beta-cell development and function. We combine transgenic mouse systems with physiological, morphometric and molecular tools to define cell-cell interactions within the pancreas, with the aim of facilitating cell replacement therapy for diabetes.
The lab welcomes outstanding international students with a Masters degree in life sciences and a passion for research. The lab provides full fellowships for qualified students.
The Landsman lab is located at Tel-Aviv University, which is the leading interdisciplinary research and teaching university in Israel. Tel-Aviv is the cultural and commercial heart of Israel. Situated on a beautiful coast of the Mediterranean Sea, it is a vibrant, young city, with great food and weather.
Lectures and courses are in English, and Ph.D. studies usually last four years. Housing on campus is an option.
Please apply by sending CV (including transcripts) and a cover letter detailing your research experience and interest through our website.
Today we are delighted to announce the winner: Zubair Nizamudeen, a PhD student in Virginie Sottile’s lab at the Wolfson STEM Centre, School of Medicine, University of Nottingham, UK. Zubair’s prize was free registration for the meeting, which he will report from for us!
Here Zubair gives us the background to his research and tells us what he’s particularly looking forward to in the meeting.
Our lab is based in the Wolfson Centre for Stem Cells, Tissue Engineering and Modelling (STEM), and are part of the Division of Cancer & Stem Cells in the School of Medicine of the University of Nottingham, UK. We focus on the genetic aspects of stem cell biology, and on the molecular mechanism regulating the differentiation ability of adult stem cells compared to embryonic stem cells. We isolate and grow adult stem cells from different origins, in order to identify the key factors involved in the acquisition of a specific lineage identity, around two central themes:
1) the control of mesenchymal stem cell differentiation, and its use for translational tissue repair,
2) neural stem cell distribution in vivo, as well as differentiation using in vitro models.
We are interested in investigating the relationship between stem cells, differentiation control, tissue repair and neoplasia.
As a final year PhD student focused on understanding the potential of adult neural stem cells in the mammalian brain, I am very excited to attend the esteemed ‘Adult Neurogenesis 2018’ conference in Dresden, Germany. In particular, I am looking forward to attending the talks related to regulation and dynamics of adult neurogenesis by F. Gage (US) and S. Jessberger (Switzerland). I am eager and passionate to learn about new research ideas and explore networking and career opportunities that are to be presented in this conference.
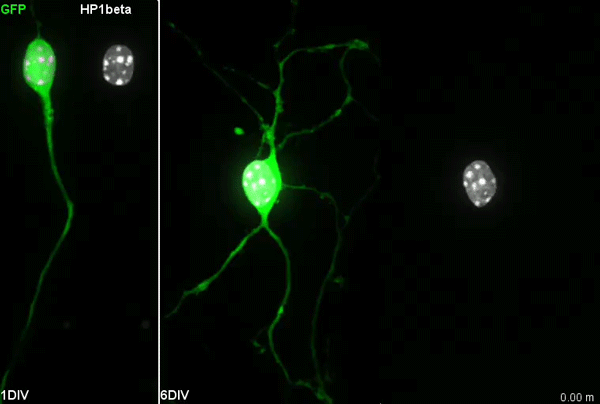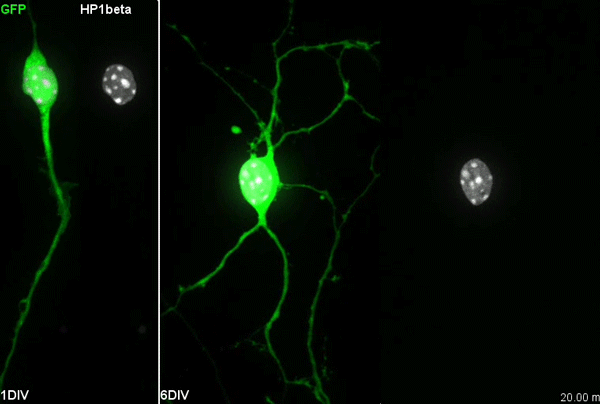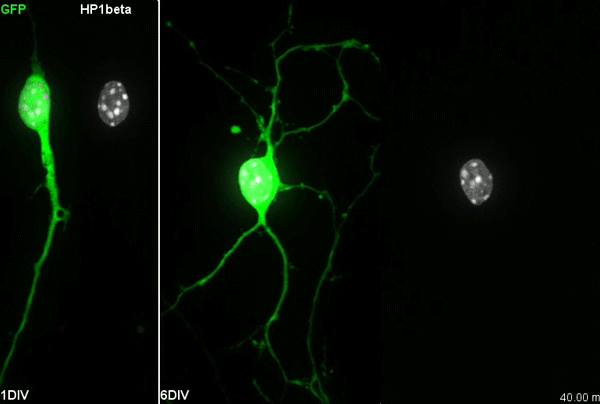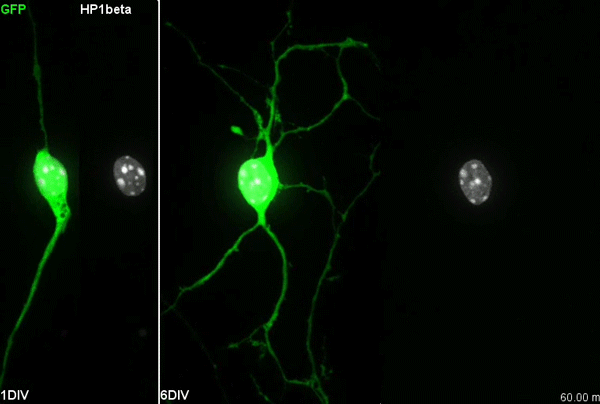
The story behind our paper:Cell-Intrinsic Control of Interneuron Migration Drives Cortical Morphogenesis. Carla G. Silva, Elise Peyre, Mohit H. Adhikari, Sylvia Tielens, Sebastian Tanco, Petra Van Damme, Lorenza Magno, Nathalie Krusy, Gulistan Agirman, Maria M. Magiera, Nicoletta Kessaris, Brigitte Malgrange, Annie Andrieux, Carsten Janke, Laurent Nguyen
The research behind this article is a good example of how looking beyond expected results can lead to unexpected discoveries. This story is not what we originally thought it was going to be about. But simplistic hypotheses failed to explain standard measurements and this lead us to a deeper question.
In the beginning:
In the 1970’s a Purkinje cell degeneration mutant mouse had been studied1 and the deficient gene responsible for the phenotype was later shown to be Ccp12. This gene codes for an enzyme, called carboxypeptidase 1, capable of digesting chained glutamates on tubulin (the building block of microtubules) or on a wide range of additional proteins3.
When we first started working in the lab of Laurent Nguyen, we were given access to the CCP1 conditional knock out mouse3. At the time, the lab was focusing on understanding the mechanisms underlying interneuron migration during cortical development. This migration is a fascinating process. The cortical interneurons are born in regions located far away from the residence of their future, mature selves. As the embryonic brain is forming, a steady flow of interneurons moves away from their birth place to reach the cortical plate. The developing brain environment gives cues to steer them in the right direction, both repulsing them from the zones they should not invade and attracting them towards correct paths. Each individual interneuron senses its environmental cues and adapts its movements accordingly. This information is translated to directed movements thanks to the dynamic remodelling of the cytoskeleton, which gives scaffolding structure to the cell but also generates contraction and pulling forces4. Microtubules and acto-myosin fibres are the main components of the interneuron cytoskeleton and their contractions and dynamics are tightly regulated. Two general levels of cytoskeletal regulation can be found: 1) gene-encoded signalling cascades regulate contraction or polymerisation/depolymerisation; 2) posttranslational modifications of cytoskeleton components can fine-tune the movement. One of these posttranslational modifications is the addition/removal of glutamate.
We reasoned that by removing Ccp1 and hyperglutamylating cytoskeletal proteins, we would subtly modify interneuron migration without stalling cells.
The phenotype observed:
Our primary observation was that too many interneurons were invading the cortex upon loss of Ccp1 expression during development. It would then take us about 2 years to understand why.
We first characterised cortical invasion by cortical interneuron in more detail. We showed that between E12.5 and birth, more Ccp1 cKO interneurons were invading the cortex of mouse embryos. This was an unusual phenotype since so far, cytoskeletal modifications lead to delayed migration and reduced number of interneurons in the cortex.
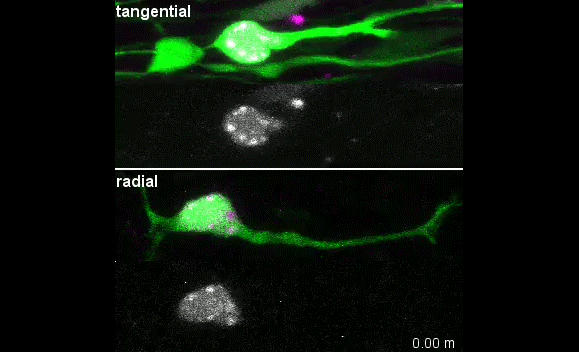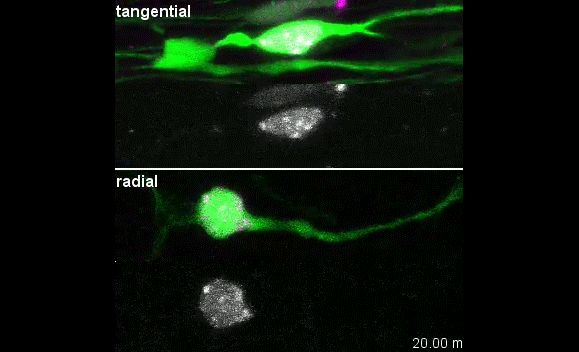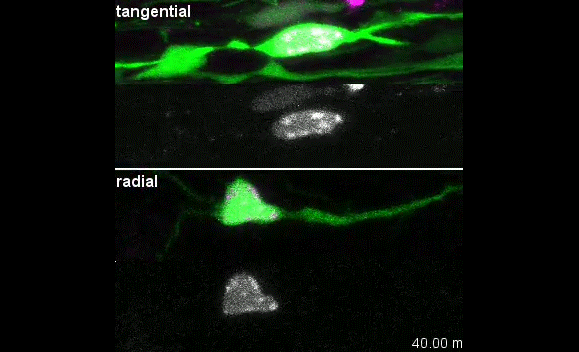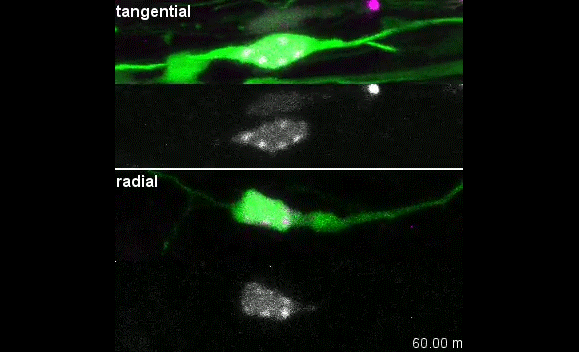
In parallel we characterized the cellular pattern of migration of these interneurons, using time lapse video microscopy. We were able to follow their behaviour as they moved toward their destinations.
GFP-expressing cortical interneurons migrating away from MGE explants prepared from E13.5 CCP1 WT embryos. Duration of recording is 5h.
Not only could we measure the average speed of displacement but we were also able to study the way interneurons cortical interneurons were moving. We observed that indeed, the behaviour of interneurons lacking Ccp1 was slightly changed despite having a preserved average speed of migration. Instead of alternating phases of nuclear pauses and large amplitude jumps called nucleokinesis5, mutant interneurons were pausing for a shorter amount of time and the amplitude of their nucleokinesis was reduced. Smaller jumps compensating shorter pauses, amounting to similar average speed.
The next question was to understand what could explain our observation of larger number of interneurons in the cortical plate?
The simplistic hypotheses:
To answer this question we tested a large number of hypotheses that yielded negative results and frustrating statistically non-significant histograms.
We first tested if more cortical interneurons were generated in upon loss of Ccp1. Our genetic recombination tool (Dlx5.6 CRE mouse line) was expressed in a small portion of dividing cells. We counted the number of mitotic cells, the number of cells replicating their DNA and the general cell cycle phases distribution of our CCP1 mutant cells and nothing was changed in our mutant.
We then reasoned that if more cortical interneurons were reaching the cortex, it could be explained by an increased survival, leading to fewer cells dying on the way. However, we did not detect any reduction of apoptosis in Ccp1 cKO cortical interneurons as compare to their controls.
Another plausible explanation was that mutant cortical interneurons would take “short-cut” pathways to the cortex and thus would be more efficiently reaching their destination. We counted the number of cortical interneurons crossing the striatum, a no-go zone for cortical interneurons and again, we did not measure any differences.
Finally, we tested whether loss of Ccp1 could affect cell fate and favour generation of cortical interneurons at the expense of oligodendrocyte that are born in overlapping regions of the forebrain from a common pool of progenitors. We showed that the mutant brain did not lack oligodendrocytes. To be sure that the conditional knockout of Ccp1 knock-down in cortical interneurons was not resulting in higher interneurons generation through a non-cell autonomous activity, we counted the number of recombinant cells in the whole brain using FACS. At E13.5 we measured the same total number of cells in Ccp1 cortical interneurons brains as compared to WT controls. This observation suggested that more cortical interneurons were displaced in the cortical compartment in Ccp1 cKO embryos.
None of the above hypotheses proved to be true. So why were there more cells invading the cortex?
The unexpected:
By this point, the project was 3 years old and although several hypotheses had been eliminated we were not closer to an explanation for our phenotype. We sat down and looked at the data. It was the moment to decide to either put the project aside or take a fresh look at the whole thing in a different light. What if the slight change of individual behaviour could influence the way the entire population moved? To help us with this new hypothesis we called upon our colleague Mohit Adhikari, a talented physicist, to help modelling the behaviour of our cells in silico. Using the parameters of speed and pause duration of migrating cortical interneurons measured by time lapse experiments, we generated trials of surrogate cells displacements. In this in silico experiment, the interneurons are challenged along the simplest path, a straight line on a 2D plane to isolate and test the effect of kinetic behaviour on the population displacement.
Displacement of cell surrogates with WT or Ccp1 cortical interneuron migration parameters. Displacement of all cell surrogates in both groups (Group A gray and Group B red); lighter marker shade indicates higher total displacement of a surrogate. Horizontal bars mark the highest displacement thresholds crossed by 75% of surrogates in each group. Duration of simulation is 600min.
The simulation allowed us to conclude that a difference in the kinetic properties of migration, such as shorter pauses and smaller jumps, increases recruitment at short distances from the starting point of CCP1 mutant interneurons in the developing cortex. We finally had a possible explanation for our phenotype, and the reason why had been staring at us from the start.
The molecular regulation:
In parallel to running in silico simulations of cortical interneurons displacement, we set out to understand the molecular regulation of migration linked with the Ccp1 mutation. We first found that acto-myosin fibres, a component of the cytoskeleton, were not contracting normally in upon loss of Ccp1 in cortical interneurons. This observation was done in time lapse video microscopy experiments when cortical interneurons were electroporated with a acto-myosin contraction fluorescent probe.
Migration of pLifeAct-Ruby electroporated E13.5 WT or cKO Ccp1 cortical interneuron homochronic mixed cortical feeder
Mutant cortical interneurons were generating almost permanent actomyosin-derived forces instead of pausing. These forces were, however, either not strong or not focused enough at the rear of the nucleus to grant correct nuclear jump. We looked at the phosphorylation state of the myosin light chain (MLC), the effector of acto-myosin contraction, and we observed hyper phosphorylation, confirming MLC over-activation. We then turned our attention to the kinase that phosphorylates MLC, MLCK3,6, and showed that it was not only a substrate of our enzyme Ccp1 in cortical interneurons but also that it was hyperactive in the mutant cortical interneurons. This showed a new molecular regulation of acto-myosin contraction and helped us understanding what lead to the abnormal movement behaviour of our mutant interneurons.
The cherry on the cake:
Finally our last question was: why is cortical invasion by cortical interneurons regulated during cerebral cortical development? We raised this question because we had noticed that the mutant brains were able to correct their number of cortical interneurons after birth by killing off the supplementary cells by apoptosis. If the problem can be corrected later why does cortical interneurons cortical invasion need to be regulated? To answer this question we looked at the cortex, the compartment receiving migrating interneurons. During our birth dating experiments, we injected BrdU to birthdate and track cohorts of cortical interneurons. We analysed BrdU injected brains at E13.5 and noticed an abnormal proliferation of the projection neuron progenitors, the intermediate progenitors in the cortex of Ccp1 mutant brains. This lead to an overproduction of projection neurons persisting in postnatal brains. This result was unexpected as our genetic recombination tool is specific to subpallial regions and does not target the cortical progenitor cell populations. Interestingly this increased proliferation was detected in regions with higher numbers of interneurons. We postulated that during development, the number of interneurons entering the cortex regulates the production of the projection neurons with which they will form connections later on. To test this hypothesis we analysed another mouse model (Nkx2.1 mutant). In these mutants brains no cortical interneurons entered the cortex and this resulted in a decreased proliferation of intermediate progenitors, suggesting again a regulatory link between the number of interneurons in the cortex and the proliferation of intermediate progenitors.
These observations are original and they unravel a regulatory crosstalk between cortical progenitors and interneurons regulating corticogenesis progression. This could be relevant for human diseases as increased cortical thickness are described in patients suffering from autism spectrum disorder 7,8.
Conclusion
Overall this work took 5 years to complete and in the end, it is far from what we expected it to be when we first started. The challenge of getting to an explanation beyond the obvious lead us to unexpected and novel results.
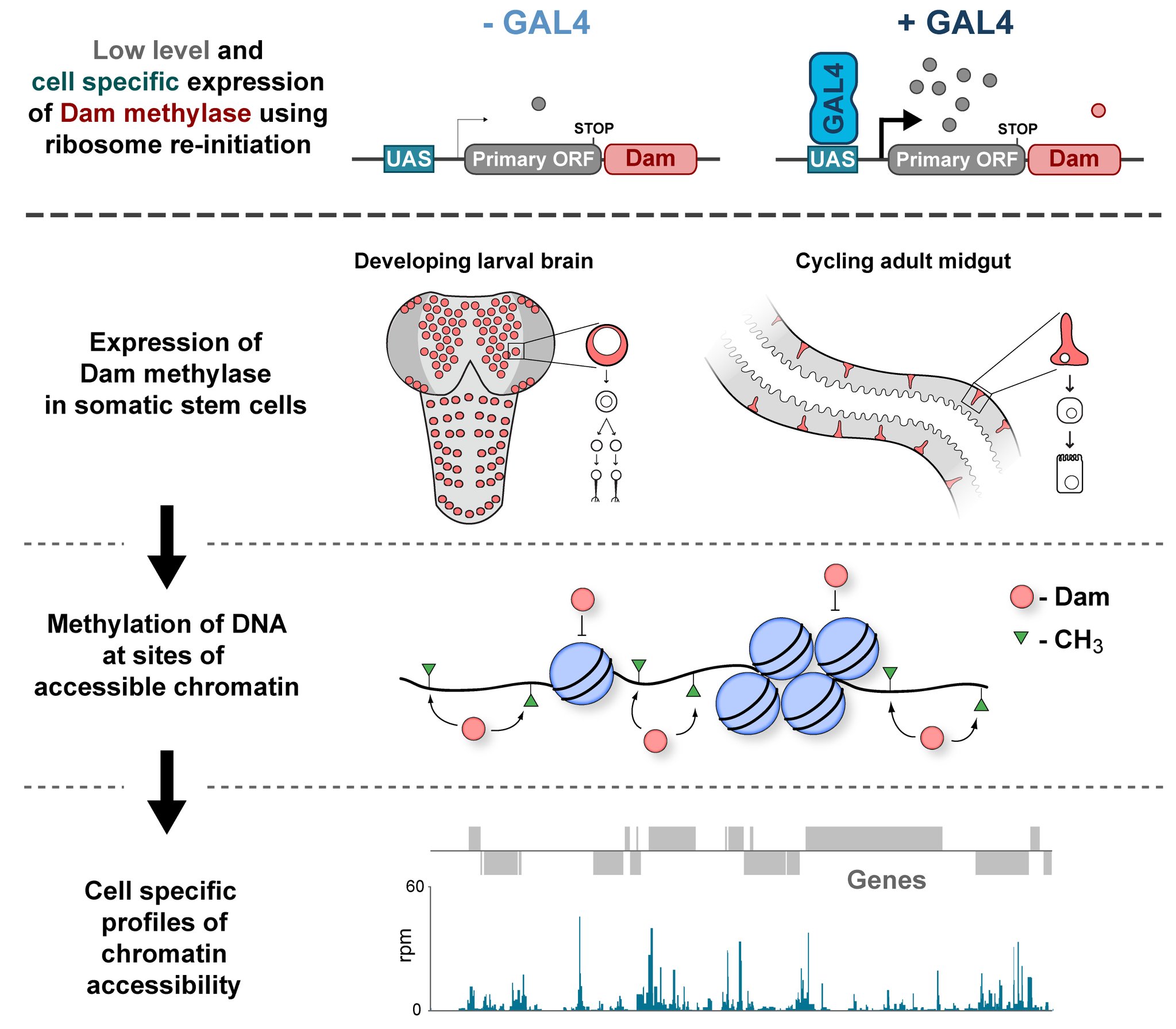
Attempting to understand the biology of a complex organ, like the brain, comes with an array of technical challenges. Those of us who would like to understand development in the context of a living organism are faced with an enormously complex arrangement of cells, which may be superficially indistinguishable from one another, and furthermore, are inconveniently hidden away inside the body of an animal. Typically, if we want to examine the molecular characteristics of one of the constituent cell types of an organ, we need to physically isolate these cells from their biological context before applying the relevant assay. Cell separation can be very technically challenging and has been recently reported to cause artefacts in gene expression [1]. Therefore, methods in which cell types within a complex tissue can be examined without the need to remove them from their in vivo environment are of great interest to biologists working on in vivo models. Measuring the accessibility of chromatin to external factors is commonly used as a way of detecting functional regions of the genome. Techniques such as ATAC-seq or DNAse-seq are widely used for this purpose [2]. However, before now, we had no way of performing these chromatin accessibility assays without cell separation. In our recent paper we attempted to address this problem [3].
TaDa! – Chromatin interactions the easy way:
In the Southall lab we make extensive use of the Targeted DamID system (TaDa), which circumvents some of these issues when assaying protein-DNA interactions. This is a development of the well-established DamID system in which a chromatin-binding protein of interest is tethered to the E. coli adenine methylase, Dam [4]. Dam activity is relatively promiscuous, and so methylates any DNA that it comes into proximity with. Once this methylation is detected, we can infer the binding sites of a protein of interest. Targeted DamID expands on this approach by expressing the Dam-fusion protein as a bicistronic transcript, downstream of a primary open reading frame (ORF). This allows for very low level expression of the Dam-fusion so that the animal does not experience toxicity related to high Dam expression, and the methylation levels are not saturating – allowing for a semi-quantitative readout of protein binding [5]. Crucially, this construct can be combined with tissue specific binary expression systems (e.g. Gal4/UAS), allowing us to profile chromatin-protein interactions in a cell type of interest. (A full review of DamID and its applications can be found here: [6]).
Chromatin accessibility profiling with CATaDa
For TaDa experiments to work it is necessary to perform a control experiment in which untethered Dam protein is expressed in the cell-type of interest. The data produced from this control is then used to normalise the Dam-fusion binding data so that a reliable DNA-interaction profile can be generated. (This can be considered to have an analogous function to the input chromatin in a ChIP-seq experiment). Having generated much of this control data in the course of performing TaDa experiments, we wondered whether it might have applications beyond just normalisation. As Dam is highly active, we reasoned that the signal obtained after expression of untethered Dam would represent interactions between Dam and any regions of chromatin that it can access. Therefore, we inferred that Dam-only data may be a good proxy for chromatin accessibility.
Given that we had gigabytes of Dam-only sequencing reads stored on our hard drives going unused, we were excited by the possibility that we could leverage these data into yielding further biological insights. To determine whether these data were comparable to established chromatin accessibility techniques, we compared some of our Dam data to previously published ATAC-seq or FAIRE-seq results [7]. We were excited to see that the signal from Dam methylation displayed many of the hallmarks of chromatin accessibility data that are observed with alternative techniques. For example, enriched signal at the transcription start site of genes. Overall, our data compared favourably to both ATAC and FAIRE-seq data. We found highly significant agreement between these datasets, and encouragingly, we saw that our data was comparable to ATAC-seq and out-performed FAIRE-seq in identifying previously validated enhancer regions. Therefore, we concluded that these data are appropriate for inferring meaningful chromatin accessibility data. We coined the term CATaDa (Chromatin Accessibility profiling using Targeted DamID) to describe this use of Dam methylation data for cell-type-specific profiling of chromatin accessibility.
Chromatin accessibility during neurogenesis
Our group is interested in understanding how the development of the nervous system is coordinated by changes to chromatin states. Therefore, we decided to apply CATaDa to the cells of the developing nervous system, including neural stem cells, their intermediate progeny (ganglion mother cells), and mature neurons. These data showed us that chromatin accessibility is incredibly dynamic between cell-types, with many regions becoming open or closed as development processes. Furthermore, we were able to examine regions of increased accessibility to detect enriched sequenced motifs which point towards factors that are involved specifically in various stages of nervous system development. We were also able to use our data to identify enhancers promoting cell-type specific gene expression.
It is often said that chromatin is more open in stem cells, then becomes less accessible when differentiated. This has been shown to be the case when comparing embryonic stem cells to (e.g.) neurons [8]. This feature is often stated to be a general feature of stem cells, and is often applied to somatic stem cells (such as neural stem cells), despite the fact that we could find no evidence for the existence of any data attempting to examine this question in vivo. We realised that with our CATaDa data, we could answer this question by looking at the distribution of sequencing reads across the genome in each cell type. We were able to confirm that chromatin was globally more open in neural stem cells, which then becomes restricted in fully differentiated neurons. Interestingly, we saw that intermediate cell types showed intermediate chromatin accessibility which was closer to that of their stem cell progenitors. At individual loci this was also observed – regions of high accessibility associated with stem cell genes were also seen to be accessible (albeit at lower levels) in their progeny, despite the fact that expression of some of these genes is tightly restricted. To see if these trends were consistent in other tissues, we also assayed cells of the adult midgut. We chose this tissue because in contrast to neurogenesis, intestinal stem cells continue to divide throughout the lifetime of the animal for maintenance and repair. The previously described trends were also seen in these cells. Together these data suggest that chromatin accessibility decreases during the development of somatic lineages and intermediate cell types may have unexpected plasticity despite being fully committed to their cell fate.
Conclusion
We hope that we have shown that CATaDa is a useful approach for assaying chromatin accessibility that will appeal to researchers who are interested in in vivo approaches. Although we have only demonstrated CATaDA in flies, Targeted DamiD has recently been shown to be effective in mammalian cells, therefore our method can reasonably be expected to be effective in any model organism in which transgenic animals can be obtained.
Dam-ed if you do and Damned if you don’t – When to use CATaDa:
Yes:
You want to profile chromatin accessibility of a subset of cells within a complex tissue / whole animal. (We typically start with dissected brains, but it is possible to use whole animals if the drivers used are specific enough)
You have a limited amount of starting material (approx. 1000 cells minimum).
You would like to profile the in vivo chromatin accessibility of a tissue, avoiding fixation or tissue dissociation artefacts.
You have done a DamID/TaDa experiment with a conventional dam-only control and want to get the most out of your data!
No:
You want to find the exact positioning of nucleosomes / enhancer sequences (CATaDa resolution is limited by availability of dam target motif – GATC).
You need to detect changes in chromatin accessibility over very short timescales. Dam must be expressed over a relatively long time period for sufficient methylation to be detected. Consequently, the output of the experiment in slightly different to that of e.g. ATAC-seq (i.e. ATAC-seq represents a snapshot of a population of cells frozen in time, whereas CATaDa will include signal representing all accessible regions over the period assayed).
We are currently seeking applications from post-doctoral candidates (LabEx-funded positions).
Project: Christophe Heinrich “Cellular Reprogramming and Brain Repair” was recently awarded a LabEx Attraction Package at SBRI. Our group focuses on direct lineage reprogramming of somatic cells into induced neurons as a novel cell-based therapy for brain repair. We previously showed that glial cells can be converted into functional induced neurons (Heinrich et al., PLoS Biology 2010, Nature Protoc 2011, Stem Cell Reports 2014, Nature Cell Biology 2015; Karow et al., Cell Stem Cell, 2012). We aim at reprogramming glial cells in the injured brain into neurons that integrate into endogenous networks and modulate the activity of these networks with beneficial effects.
Environment: The University of Lyon (IdEx Initiative of Excellence) and LabEx CORTEX represent an outstanding scientific environment. Research at SBRI is at the interface of development biology with a strong emphasis on stem cell biology, neurogenesis and functional neurobiology of the brain. SBRI benefits from state-of-the-art research platforms for stem cell bioengineering and provides a high-quality scientific, technological and medical environment.
Candidates: We are looking for enthusiastic and highly motivated scientists holding a PhD and willing to join a challenging research area at the edge of cellular biology and neurosciences. Applicants are expected to have a strong background and proven track record in neurosciences and/or cell biology. Experience in working with mice, cellular and molecular biology (cloning), and/or electrophysiology would be helpful. English proficiency is mandatory and strong communication skills and team spirit are expected.
Last Friday March 9, a research symposium was held at Duke University in Durham, North Carolina to honor the career and retirement of Professor Brigid Hogan, Chair of the Department of Cell Biology (Hogan Lab Webpage: http://bit.ly/2ImZxn5). Current and former Hogan Lab members, colleagues, and friends came from across America, Japan, and the United Kingdom to join in the celebration of a truly remarkable scientist. There were 14 invited speakers, including former students, postdocs, colleagues from Brigid’s days at Vanderbilt University in Nashville, Tennessee, current members of the Department of Cell Biology at Duke, and friends in the mouse development and genetics field. More than 150 participants that included local students and postdoctoral fellows came to hear outstanding research talks. Among the participants, luminaries in the mouse developmental biology field were there to honor Brigid, including Gail Martin, Liz Robertson, Liz Lacy, Frank Costantini, Phil Soriano, Terry Magnuson, Blanche Capel, Kat Hadjantonakis, and Mary Dickinson. The symposium started with surprise videos from friends Fiona Watt (King’s College London) and Jim Smith (Francis Crick Institute), sending their congratulations to Brigid and one from Brigid’s third graduate student, Peter Holland (University of Oxford), praising her skills at inspiring his confidence as a young scientist during his thesis research. The research talks discussed current research, including gene regulatory networks, cutting-edge microscopic imaging, organogenesis, the genetic basis of human disease, novel gene manipulation approaches, embryos on a chip, organ-specific stem cells, high-throughput mouse mutant phenotyping, and tissue regeneration. The talks highlighted the advances in the field of cell and developmental biology and why this area of research is so important for basic knowledge and human health. To learn more about Brigid’s background and motivation to study mouse embryos and organs see her 2015 interview with The Node (http://bit.ly/2DqfZiM).
In addition, to the wonderful science that was presented that day, all of the speakers had a “Brigid story” that they shared with the audience. Many spoke of her drive, curiosity, generosity, patience, and for those who were trained in her lab, the lessons they learned from her. These included ‘don’t talk yourself out of an experiment, sometimes you just have to do it’, ‘be brave’, ‘finish what you start’, ‘speak up and speak out’. My favorite was ‘don’t apologize for being a tall, confident woman’. Everyone praised Brigid’s skills as a mentor. You can read about Brigid’s thoughts on mentoring in a recent interview in Cell Stem Cell – Mentoring the Next Generation (http://bit.ly/2Dk1PiM). Yes, Brigid is “retiring” but she will still be very active. At the end of the symposium, Brigid thanked everyone for attending and participating, especially those who traveled such long distances. She said it brought “a joy to my heart” and was a “day I’ll always remember”.
Symposium Poster
Brigid (peacock-colored highlights) with Chris Wright (left) and Ray Dunn (middle). Chris is a close colleague from Vanderbilt University and Ray was one of Brigid’s graduate students also at Vanderbilt.
Symposium organizers Emma Rawlins (middle) and Jason Rock (right) both former postdocs with Brigid and Gail Martin.
Brigid’s former students and postdocs gather together at the Symposium Dinner.
Neuronal migration is critical for mammalian brain development. In many migrating neurons, the nucleus translocates from the trailing to the leading edge of the cell in a manner dependent on the actin and microtubule cytoskeletons, but how these cytoskeletons interact and their relative contribution to the forces that move the nucleus has remained unclear. This week we feature a paper published in the latest issue of Development that uses live imaging to understand this process, revealing some fascinating cellular behaviours. We caught up with first author You Wu and his supervisor Mineko Kengaku, Professor at the Institute for Integrated Cell-Material Sciences at Kyoto University, to find out more about the story.
From left to right: Mineko Kengaku, You Wu, Hiroki Umeshima and Junko Kurisu.
Mineko, can you give us your scientific biography and the questions your lab is trying to answer?
MK We study how neurons are built up into the beautiful architecture of the brain cortex. A major effort of my lab has been devoted to live-imaging of growing neurons for a mechanistic understanding of brain formation, using the clear and simple cerebellar cortex as a model. We especially focus on the mechanisms and dynamics of cytoskeletal organization during migration of granule cells and dendrite arborization of Purkinje neurons, the two major events during the formation of the three-layered cortex of the cerebellum. I did my PhD study on the molecular mechanism of the primary axis formation in the Xenopus CNS, under supervision of Dr Harumasa Okamoto at the University of Tokyo. I wanted to further study the molecular signals regulating morphogenesis, and joined the laboratory of Cliff Tabin at Harvard Medical School as a post-doc. His lab had just discovered Sonic hedgehog as a bona fide morphogen in the chick embryo. I was extremely lucky to be present in the lab at moments when long-standing questions were clarified, such as the mechanisms of left-right asymmetry, bone formation, fore- and hind limb differentiation, etc. As a consequence of this rapid progress, many fundamental questions in morphogenetic signals had been clarified by the time I started my own lab at the RIKEN Brain Science Institute. The major interests in developmental biology then shifted toward stem cells and disease mechanisms. But I was still fascinated with animal morphology, and therefore decided to look into the beautiful shapes of neurons in the well-organized cortex of the brain.
And You, how did you come to be involved with this project?
YW I was attracted by the beautiful morphology of neurons and the structure of the cortex, and joined the Kengaku Lab as a Master’s student. I first worked on hippocampal neurons to seek how they achieve their unique dendritic shape by live-imaging. The dynamics of development really excited me. Around the time I completed my M.S. degree, I saw a movie of nuclear rotation in a migrating neuron, and was simply interested in the phenomenon.
What was known about the role of the cytoskeleton in CGC nuclear migration before your work?
MK & YW Unlike mesenchymal cell migration, neurons typically form a long leading process and move the nucleus into the leading process in a saltatory manner independently of the constant leading process extension. Since the discovery of an evolutionary conserved cytoplasmic dynein complex protein LIS1 as a responsible gene for smooth brain syndrome (type 1 lissencephaly), accumulating evidence has supported the model that the nucleus is pulled forward by dynein motor activity along the microtubules extended from the leading process. Subsequent studies have also implicated the contractile force of actomyosin in nuclear movement, although competing hypotheses for the site of actomyosin force generation have been proposed using different neuron models.
CGC migration has provided an in vitro model for cell biology study of neuronal migration ever since the first time-lapse observations in 1980s. Recent studies from the groups of Xiao-bing Yuan and David Solecki have provided strong evidence that actomyosin localized in the proximal leading process exerts the driving force for nuclear migration in CGCs. However, the precise interplay and relative contribution of actomyosin and microtubules during nuclear migration have remained to be elucidated.
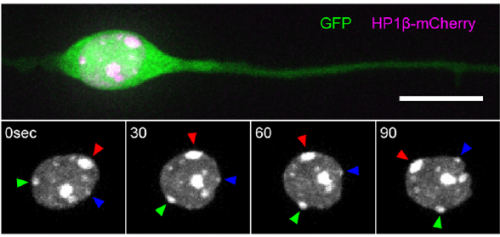
Heterochromatin spots reveal a migrating nucleus, from Figure 1, Wu, et al. 2018
Can you give us the key results of the paper in a paragraph?
MK & YW For better understanding of the complicated interplay between microtubule- and actin-dependent mechanisms driving nuclear migration in CGCs, we performed a high spatiotemporal resolution imaging using spinning-disk confocal microscopy. We found that the nucleus displayed unexpectedly dynamic motion during migration, including fast rotation in the soma. By quantitative 3D image analysis combined with molecular and pharmacological manipulations, we demonstrated that the nuclear rotation is driven by the same strain energy inducing nuclear translocation. Furthermore, among the two major cytoskeletal forces driving nuclear migration, rotation is solely dependent on microtubule systems but not actomyosin, while nuclear translocation requires both actomyosin and microtubules. We thus used nuclear rotation as a readout of microtubule-dependent tensile stress during nuclear migration, which led to the second major finding that both dynein and kinesin-1 are involved in the force transmission. Our results suggest that microtubules and associated bidirectional motors exert force to small points on the nuclear envelope via interaction with nesprins, and steer rotation or translocation depending on the position of the force points.
Nuclear dynamics of a migrating and a post-migratory CGC in vitro. Movie 1 from Wu, et al. 2018
I have to say that the videos in your paper are remarkably beautiful! Can you tell me what it was like to first see nuclear rotation in migrating CGCs?
MK Our co-author Hiroki Umeshima is highly skilled in live imaging, and he set up most of the imaging systems used in the present study. His first movie of nuclear rotation caught my eye, but I did not begin the present study for more than a year, as I was unsure if the rotation is a biologically significant event or an idling motion caused by excessive force to the nucleus in culture. But, the more movies I watched, the more confident I became that the rotation was driven by active forces on small points of the nucleus during nuclear migration. Around that time, You had developed his skill in image analysis, so I asked him if he was interested in analyzing this mysterious phenomenon, and he said ‘Yes’.
YW It was really interesting. I never imagined the nucleus would rotate so fast and frequently during migration. The movies also showed the dynamic deformation of the nucleus. I reckoned these motions could be indicating force transmission, and started to find a way to analyze the dynamics.
How do you think the opposing microtubule motors dynein and kinesin work together to drive unidirectional nuclear motion?
MK & YW We observed frequent switches of rotation direction both in vivo and in vitro (Movie 4 and 5), indicating that the nuclear surface readily moves in both anterograde and retrograde directions by dynamic interplay of the bidirectional motors on bipolar microtubules. Given that the polarity of perinuclear microtubules is mixed but strongly biased, dynein probably acts as the predominant motor and moves the nucleus along the retrograde microtubules (with minus-end toward the leading process) as previously indicated, while kinesin drives back-step movement or rotation. Involvement of bidirectional motors in nuclear migration along uniformly oriented microtubules has been found in long hypodermal precursor cells in nematode embryos (Bone et al., Development 2016; Fridolfsson and Starr, Dev. Biol. 2010). Both hypodermal cells and CGCs are elongated and move nuclei through narrow interstitial and tissue spaces, where dynamic bi-directional movement by opposing motors might adjust the precise positioning of the nucleus and help it squeeze through constrictions.
Nuclear dynamics of migrating CGCs in an organotypic slice. Movie 4 from Wu, et al. 2018.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
I was excited to observe the dynamics of the nucleus in the organotypic slice culture. Until then, I was focused on the in vitro culture system, where the rotation and deformation occurs sporadically. I found the nucleus in 3D tissue much more dynamic, and there was almost no moment when the nucleus stayed still. It was interesting to see that forces seemed to be always applied to the nucleus, even if it was not translocating
And what about the flipside: any moments of frustration or despair?
While it was an exciting moment to see the nuclear motion in tissue, it was very challenging to analyse it quantitatively. It would be interesting and informative to characterize its properties in tissue precisely, but it was difficult to acquire enough resolution in live-imaging.
What next for you, You?
It is always fun for me to observe dynamic things. I am still interested in neural development and also in brain function, which is achieved by such well-organized structure, and am thinking about doing postdoc in related fields.
Kinesin inhibition disrupts CGC migration in vivo, from Figure 7, Wu, et al. 2018.
Where will this work take the Kengaku lab?
We want to further elucidate the actual crosstalk of microtubule motors and nuclear envelope during nuclear motion. New imaging techniques with higher spatiotemporal resolution might be required to pursue the next problem. Also, the dynamic deformation and rotation shown in the present study suggest that the nucleus is very soft and flexible in newborn neurons. We are interested in how the mechanical properties of the nucleus is determined, and if they contribute to neuronal migration.
Finally, let’s move outside the lab – what do you like to do in your spare time?
MK I must confess that I do not have so much spare time for special activities after research and family. I am a food fan and enjoy cooking every day. I sometimes eat-out with my family and friends on weekends. Luckily Kyoto is a great place to explore nice restaurants of various styles, from traditional Japanese Kaiseki to casual French bistro.
YW I like to sing songs, sometimes in a bit loud voice if nobody is by.
Here we discuss the curious case of female-restricted epilepsy, an unusual disorder caused by mutations in the Protocadherin 19 (PCDH19) gene. How changes in this cell adhesion gene cause seizures and intellectual disability in girls (but not boys) has been a mystery since this condition was first described over 20 years ago. By pursuing several lines of enquiry including in vitro cell sorting assays, CRISPR/Cas9 mouse models and patient MRIs, we have finally “cracked the case”, although intriguing questions remain about the neuronal pathology that underpins this unique condition.
PCDH19-GCE-no other disorder is quite like it
The most striking characteristic of PCDH19-GCE is its unique X-linked inheritance pattern which was first described by Ryan et al in 1997. Typically, X-linked disease-causing mutations affect males as they do not have a second wild type (WT) copy of the gene to compensate. As the name suggests, Protocadherin 19 Girls Clustering Epilepsy (PCDH19-GCE) is caused by mutation in the X-linked gene PCDH19 but this disorder does not follow the “typical” X-linked recessive disease inheritance pattern. Instead, heterozygous females (who have one WT copy and one mutated copy of the PCDH19) are affected while hemizygous males are not. A mysterious condition indeed!
It was not until 2008 that PCDH19 mutations were identified as causing the disease, and multiple hypotheses were put forward to explain its unusual inheritance pattern. It was initially proposed that PCDH19 mutations may be dominant negative, but the identification of whole gene deletions and examples of nonsense mediated decay provided strong evidence opposing this hypothesis (Dibbens et al., 2008). It was also suggested that males may be able to compensate for the loss of PCDH19 through the Y-linked PCDH11Y gene. However, this has also been ruled out due to the discovery of several mosaic males with early somatic mutations in PCDH19 that phenocopy affected girls (Depienne et al., 2009; Terracciano et al., 2016). The third and final hypothesis was based on two key facts. The first is that PCDH19 is located on the X-chromosome and is subject to X-inactivation. This mechanism ensures that females randomly “silence” one copy of the X chromosome in every cell to match the expression levels in males (who are XY). Thus, females with PCDH19 mutations have a mixture of WT and mutant neurons. The second was PCDH19’s known role as a homotypic cell adhesion molecule. Together these pieces of information led Dibbens et al , 2008 and Depienne et al 2009 to suggest that mosaic expression of PCDH19 in female brains leads to abnormal neuronal connections between PCDH19 WT and PCDH19 mutant cells affecting neural network formation and ultimately giving rise to seizures and intellectual disability. However, it was unclear how mosaicism could lead to PCDH19-GCE at the molecular, cellular and network level. Furthermore, this model did not explain why the complete absence of PCDH19 (in males) does not cause epilepsy.
The smoking gun: cell sorting assays provide evidence for a PCDH adhesion code
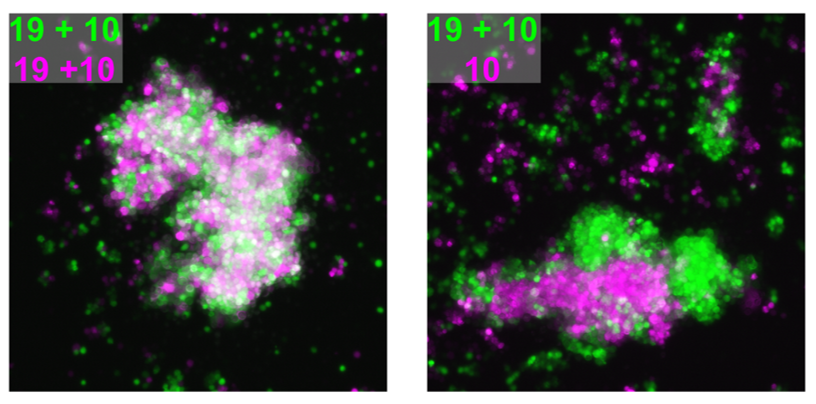
We attempted to unravel this mystery by combining complementary in vitro and in vivo experimental approaches. A valuable clue came from recent papers showing that clustered PCDHs (which are structurally similar to the non-clustered PCDH family to which PCDH19 belongs) act in combination to form complexes at the cell surface with highly specific homotypic binding affinities (Thu et al., 2014). This peaked our interest as it was unclear from the “cellular interference model” how the loss of a single cell adhesion molecule could disrupt normal interactions given that neurons typically express many PCDH family members. Using a co-immunoprecipitation and cell aggregation assays we demonstrated that PCDH19 can form promiscuous cis and highly specific trans interactions both of which are required to generate specific combinatorial binding affinities. We then mixed together two populations of cells expressing different combinations of PCDHs and observed segregation between the two populations when they differed by just a single PCDH (Figure 1). This confirmed that the “PCDH adhesion code” was acutely sensitive to the absence of PCDH19.
Figure 1 – Adhesion specificity is determined by single differences in PCDH expression. Two populations of cells expressing the same PCDHs show extensive mixing (left) compared to that of two populations with a single difference in PCDH expression, which display striking segregation (right).
The next step was to assess how disruption of PCDH19-dependent adhesion would impact the developing brain, where a complex array of cell adhesion molecules and binding partners are expressed. To investigate this, we turned to our previously characterised Pcdh19 Knockout (KO) mouse model (Pederick et al., 2016). Like humans, Pcdh19 is located on the X chromosome in mice and therefore heterozygous mice have mosaic expression of Pcdh19 due to X-inactivation. Interestingly, electrocorticogram analysis revealed significantly elevated activity in heterozygous mice when compared to WT controls. Importantly, this phenotype was not present in mice completely lacking Pcdh19, which matched the unique X-linked inheritance of PCDH19 epilepsy.
CRISPR/Cas9 joins the investigation
While the electrical phenotype of the mouse was consistent with humans, we couldn’t investigate the cellular impact of mosaic PCDH19 expression in the brain because we were lacking a specific PCDH19 antibody with which to identify PCDH19+ cells. We therefore turned to the CRISPR/Cas9 genome editing system and generated a mouse with an epitope-tagged (HA-FLAG) version of PCDH19, allowing us to identify PCDH19 WT cells with commercially available antibodies. To detect “PCDH19-expressing” Pcdh19 null cells we used the previously mentioned PCDH19 KO mouse which has a LacZ reporter. Now we were set to answer the key question- how does mosaic expression of Pcdh19 in heterozygous female brains affect the behaviour of the WT PCDH19 and null PCDH19 cell populations?
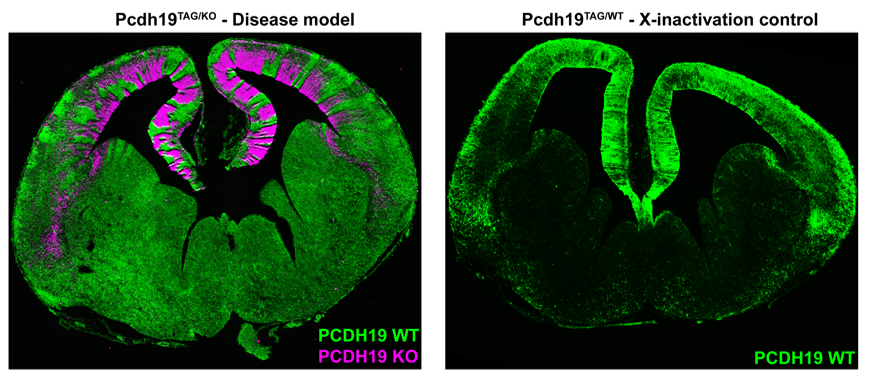
Simultaneous labelling of WT PCDH19 and null PCDH19 cells in PCDH19 heterozygous mice revealed a striking pattern of alternating PCDH19 +ve and PCDH19 -ve regions (Figure 2). This pattern was particularly obvious in the developing cortex where it resembled “tiger stripes”. Before getting too excited, we needed to address the possibility that the apparent segregation simply arose from X-inactivation and subsequent clonal expansion of PCDH19 +ve and PCDH19 -ve cells. We therefore generated “WT” mice with one tagged copy of PCDH19 and one (untagged) WT PCDH19 allele. This critical control was only possible because we had generated the tagged allele (i.e. even if we had a PCDH19 Ab we could not have done this experiment). Critically, there was no sign of the “tiger stripes” in this control- instead we detected small patches of tagged and untagged PCDH19 cells along the ventricle with subtle variations in the cortical plate (Figure 2). So, evidence for active cell segregation in heterozygous females was beginning to stack up. Interestingly, the abnormal segregation pattern of WT PCDH19 and null PCDH19 cells was different in each mouse, presumably caused by the random nature of X-inactivation. Since the symptoms of PCDH19 epilepsy are highly variable (even in identical twins (Higurashi et al., 2012)) it is possible that different segregation patterns contribute to the severity of the disorder. Although not performed in this study it would be interesting to correlate the X-inactivation patterns seen in individual mice with their ECoG recordings to identify any patterns that lead to higher electrical brain activity.
Figure 2 – Mosaic expression of PCDH19 leads to abnormal cell sorting in the developing cortex. Segregation of Pcdh19 +ve and Pcdh19 -ve cells results in distinct “tiger stripes” (left). Normal X-inactivation of Pcdh19 does not result in “tiger stripes” (right).
In our opinion the most interesting feature of PCDH19 epilepsy is not that mosaicism leads to disease but that individuals who completely lack PCDH19 do not have the disease. As mentioned above, homozygous null mice did not display increased electrical brain activity. We hypothesised that this phenotypic “rescue” was due to the uniform loss of Pchd19 and the consequent restoration of normal cell sorting during cortical development. To test this, we developed a strategy to rapidly assess in vivo cell sorting in Pcdh19 null embryos. We deleted the functional PCDH19TAG allele in heterozygous (PCDH19TAG/PCDH19LACZ) zygotes using CRISPR/Cas9, transferred the embryos to pseudopregnant females for further development, and then used X-Gal staining to track the location of the PCDH19 null cells. While negative (i.e. heterozygous) controls showed the expected segregation phenotype, no significant segregation occurred after deletion of the (WT) PCDH19TAG allele. These findings provided further evidence that abnormal cell sorting is caused by the differential adhesion affinities between WT PCDH19 and null PCDH19 cells. Furthermore, the absence of abnormal cell sorting in animals completely lacking PCDH19 provides a clear cellular phenotype that explains the unique inheritance pattern of PCDH19 epilepsy.
Finally, we postulated that abnormal cell sorting in humans could lead to brain malformations due to the prolonged expansion and increased cortical folding compared to mice. Abnormal cortical sulcation was observed in four girls with causative PCDH19 mutations. We identified variably positioned cortical defects that included bottom of the sulcus dysplasias, abnormal cortical folding, cortical thickening and blurring of the grey/white junction. It is not known how abnormal cell sorting may generate these cortical malformations, however, insight into the cellular mechanism could be gained by the generating a PCDH19 mutant ferret with CRISPR/Cas9 genome editing, a model organism that has extensive cortical folding.
For the future…
These findings provide some long-awaited answers to the mysterious inheritance of PCDH19 epilepsy. However, despite these insights, there is still much to discover about how mosaic expression of PCDH19 leads to epilepsy, intellectual disability and autism. It is possible that in addition to cell sorting, other processes such as neuronal wiring, synapse formation, maintenance and function are disrupted by differential adhesion affinities caused by mosaic expression of PCDH19. Perturbation of these fundamental neuronal processes is implicated in many neurodevelopmental disorders and it seems possible that they may also be altered in PCDH19 epilepsy. Much sleuthing lies ahead!
Higurashi, N., Shi, X., Yasumoto, S., Oguni, H., Sakauchi, M., Itomi, K., Miyamoto, A., Shiraishi, H., Kato, T., Makita, Y., et al. (2012). PCDH19 mutation in Japanese females with epilepsy. Epilepsy Res. 99, 28–37.
Applications are sought from established as well as career development researchers with interests in nervous system and/or human development, including stem cells and organoid approaches. We are also seeking cell biologists with interests in signalling, trafficking and cell adhesion.
Applicants will be expected to be competitive for personal fellowships and/or programme level grant support and have a strong publication record. For career development researchers the start-up package typically includes financial support for relocation and
essential equipment and a defined transition to tenure after 5-6 years. New researchers are supported by a structured mentoring programme. The University of Dundee operates policies enabling flexible working to ensure a good work life balance.
The Division of Cell & Developmental Biology consists of 8 research groups studying the regulation of differentiation in developing organisms, stem cells and adult tissues and how this is altered in disease states. Our work combines classical embryology, cell biological and genetic approaches as well as mathematical modelling to understand gene function and regulation in differentiating tissues. A particular shared interest is in epithelial cell biology, and this has led to our cooperative effort to maintain and develop state of the art imaging technologies. The Division also hosts the Dundee Human Pluripotent Cell Facility, which provides quality controlled human pluripotent cells and training in their use.
How to apply: please provide CV, details of 3 academic referees and a brief summary of future research plans. For informal enquiries please contact
 (No Ratings Yet)
(No Ratings Yet)


 (7 votes)
(7 votes) (2 votes)
(2 votes)