This year, is the BSDB’s 70th anniversary! Expect many extras on our Spring Meeting (15-18 April 2018 in Warwick)! For example, the history rap, a contemporary oral rendering of BSDB history, featuring Jerry aka Gerald H Thomsen PhD and produced and mixed by Philip Larsen – BSDB member and DJ!
Here at Development towers the excitement is mounting for the BSDB’s Spring meeting, which starts in Warwick on Sunday. The meetings are always great fun but this year promises to be particularly special – the society is celebrating its 70th birthday and has assembled an all star cast of speakers.
The epic conference poster designed by Sally Lowell!
If you’re coming, be sure to check out The Company of Biologists’ stand and look out for Katherine Brown (Development Executive Editor), Seema Grewal (Development Reviews Editors) and Aidan Maartens (the Node Community Manager & Development Online Editor). We’ll also be accessible at the bar to talk about everything related to publishing, including preLights, the new preprints service.
As we looked at the line up we realised we’d interviewed many of the speakers and chairs for Development over the last few years, either in print or on screen, so have decided to share them with you here.
This year we are pleased to announce Professor Wolf Reik as our keynote speaker. His research group, based at the Babraham Institute, investigates the roles of epigenetic gene regulation in mammalian development.
We are also honoured to have Dr Susan Cox and Dr Germano Cecere as our outstanding invited speakers. Dr Susan Cox, from King’s College London, has a strong background in biophysics and has done quantitative analysis using super-resolution microscopy. Dr Germano Cecere, from the Institute Pasteur, focuses on the characterization of short-RNA-based mechanisms of epigenetic inheritanceduring animal development and upon environmental changes.
As it is traditional, we will end up the meeting with a panel discussion, this time dedicated to Publishing, Editing and Journals. There we will be able to hear the views and experiences of Jennifer McLennan, Head of External Relations at eLife, Robert Kiley, Head of Open Research at the Wellcome Trust, and Katherine Brown, Executive Editor of Development.
Finally, we are looking for additional talks and posters from PhD students and postdoctoral researches who work on the fields of Evo-Devo, Stem Cell Biology, Embryology or more general Developmental Biology. Experimental and theoretical approaches are equally acceptable.
Whether you want to submit an abstract for a talk or poster, or just attend the meeting, you can register here. Registration is free, and there will be prizes for the best talks and posters.
Deadline for abstract submission is midnight on 14th April 2018.
Last day tomorrow to apply for the EMBO practical course on 3D Developmental Imaging (April 11th!).
Are you a developmental biologist struggling with 3D imaging (confocal, 2p, light-shet, OPT)? Don’t miss this opportunity: registration fee INCLUDES meals, course materials, hotel and local transport, all made possible by generous sponsorship from EMBO, the Gulbenkian Institute and our commercial partners. EMBO provides in addition a limited number of travel grants for candidates from Croatia, Czech Republic, Estonia, Greece, Hungary, Italy, Malta, Lithuania, Poland, Portugal, Slovakia, Slovenia, Spain and Turkey and Chile, India, Singapore and Taiwan.
There is a vacancy for a postdoctoral research fellow position at the Sars International Centre for Marine Molecular Biology (www.sars.no) in the research group headed by Dr. Pawel Burkhardt. The position is for a period of 4 years and is funded on the Sars Centre core budget. The Sars Centre belongs to the University of Bergen and is partner of the European Molecular Biology Laboratory (EMBL) (www.embl.de). The place of work will be at the Sars Centre. The starting date is negotiable but preferably no later than 01 September 2018.
About the project/work tasks: The Burkhardt group combines comparative biological systems in the laboratory to understand when and how the first synapses and neurons evolved. The group is particularly interested in studying the origin and evolution of synaptic proteins (Burkhardt et al, 2011 PNAS; Burkhardt et al, 2014 MBE; Bhattacharyya et al, 2016 eLife). We are looking for a highly self-motivated and enthusiastic Postdoctoral Research Fellow with interests in evolutionary biology, neurobiology and cell biology. The project will focus on the characterization of synaptic protein homologs in sponges and ctenophores to better understand the evolution of first neuron-like cell types in animals. The successful candidate will undertake research with the possibility to use a variety of techniques, ranging from super resolution immunofluorescence and electron microscopy, biochemical methods (protein purification, co-IPs, and analytical ultracentrifugation), mass spectrometry to X-ray crystallography to study synaptic protein homologs in sponges and ctenophores. The successful candidate will work in close association with the group leader and other lab members with the aim to eventually contribute to the further development of the project in line with her/his interests.
Qualifications and personal qualities:
The applicant must hold a Norwegian PhD or an equivalent degree or must have submitted his/her doctoral thesis for assessment prior to the application deadline. It is a condition of employment that the PhD has been awarded
Strong motivation/enthusiasm to perform research at an internationally competitive level
Practical experience in biochemical techniques (protein purification, Co-IPs) and with different fluorescence imaging techniques is highly desirable
Specific experience with sponges or ctenophores is beneficial, but not essential
The ability to work both independently and to cooperate with others in a structured manner is essential
Proficiency in both written and oral English
About the position of postdoctoral research fellow:
The position of postdoctoral research fellow is a fixed-term appointment with the primary objective of qualifying the appointee for work in top academic positions. The fixed-term period for this position is 4 years. Individuals may not be hired for more than one fixed-term period as a postdoctoral research fellow at the same institution. Upon appointment, applicants must submit a project proposal for the qualifying work including a work schedule. It is a requirement that the project is completed in the course of the period of employment.
We can offer:
A professional, challenging and international working environment.
Well-equipped, modern laboratories and facilities
Salary at pay grade 57 (code 1352 / pay range 24, alternative 1) according to the state salary scale upon appointment. This constitutes a gross annual salary of NOK 490.900. Further promotions are made according to length of service. For particularly highly qualified applicants, a higher salary may be considered
Enrolment in the Norwegian Public Service Pension Fund (SPK)
A position in an inclusive workplace (IA enterprise)
Good welfare benefits
Your application in English must include:
A cover letter of the applicant’s research interests and motivation for applying for the position.
The names and contact information for at least two reference persons. One of these must be the the main advisor from the PhD programme.
CV including most relevant (i.e. first author) published papers and pre-prints
Transcripts and diplomas and official confirmation that the doctoral thesis has been submitted
Relevant certificates/references
List of any works of a scientific nature (publication list)
Detailed information about the position can be obtained by contacting: Group Leader Pawel Burkhardt, tlf.: +47 55 58 43 57, email: Pawel.Burkhardt@uib.no.
The state labour force shall reflect the diversity of Norwegian society to the greatest extent possible. Age and gender balance among employees is therefore a goal. People with immigrant backgrounds and people with disabilities are encouraged to apply for the position.
We encourage women to apply. If multiple applicants have approximately equivalent qualifications, the rules pertaining to moderate gender quotas shall apply.
The University of Bergen applies the principle of public access to information when recruiting staff for academic positions.
Information about applicants may be made public even if the applicant has asked not to be named on the list of persons who have applied. The applicant must be notified if the request to be omitted is not met.
Further information about our employment process can be found here.
There is a vacancy for a PhD position at the Sars International Centre for Marine Molecular Biology (www.sars.no) in the research group headed by Dr. Pawel Burkhardt. The position is for a fixed-term period of 4 years and is subject to funding on the Sars Centre core budget. The Sars Centre belongs to the University of Bergen and is partner of the European Molecular Biology Laboratory (EMBL) (www.embl.de). The place of work will be at the Sars Centre. The starting date is negotiable but preferably no later than 01 September 2018.
About the project/work tasks: The goal of the Burkhardt group is to reconstruct the evolutionary origin of synapses and neurons. The group is particularly interested in studying synaptic protein homologs in choanoflagellates, sponges and ctenophores. We are looking for a highly self-motivated and enthusiastic PhD student with interests in evolutionary biology, neurobiology and cell biology. The project will focus on when the protein signalling complexes required for synaptic activity first evolved and how they functioned at a molecular level (Burkhardt et al, 2011 PNAS; Burkhardt et al, 2014 MBE; Bhattacharyya et al, 2016 eLife). The successful candidate will undertake research with the possibility to use a variety of techniques, ranging from super resolution immunofluorescence and electron microscopy, various biochemical methods to X-ray crystallography to study synaptic protein homologs in choanoflagellates. The successful candidate will work in close association with the group leader and other lab members with the aim to eventually contribute to the further development of the project in line with her/his interests.
Qualifications and personal qualities:
The applicant must hold a master’s degree or the equivalent or must have submitted his/her master’s thesis for assessment prior to the application deadline. It is a condition of employment that the master’s degree has been awarded
Routine experience in standard molecular and cellular biology techniques is required
Practical experience in biochemical techniques (protein purification, Co-IPs) and with different fluorescence imaging techniques is highly desirable
Specific experience with choanoflagellates is beneficial, but not essential
The ability to work both independently and in a structured manner, cooperate with others and a possess high motivation and enthusiasm is essential
Proficiency in both written and oral English
About the PhD:
The duration of the PhD position is 4 years, of which 25 per cent of the time each year comprises required duties associated with research, teaching and dissemination of results. The employment period may be reduced if you have previously been employed in a recruitment position.
About the research training:
As a PhD Candidate, you must participate in an approved educational programme for a PhD degree within a period of 4 years. A final plan for the implementation of the research training must be approved by the faculty within three months after you have commenced in the position. It is a condition that you satisfy the enrolment requirements for the PhD programme at the University of Bergen.
We can offer:
A professional, challenging and international working environment
Well-equipped, modern laboratories and facilities
Salary at pay grade 50 (Code 1017/Pay range 20, alternative 8) in the state salary scale. Currently equal to NOK 436.900. Further promotions are made according to qualifications and length of service in the position
Enrolment in the Norwegian Public Service Pension Fund (SPK)
A position in an inclusive workplace (IA enterprise)
Good welfare benefits
Your application in English must include:
A brief account of the applicant’s research interests and motivation for applying for the position
The names and contact information for two reference persons. One of these must be the main advisor for the master’s thesis or equivalent thesis
CV
Transcripts and diplomas showing completion of the bachelor’s and master’s degrees, or official confirmation that the master’s thesis has been submitted
Relevant certificates/references
A list of any works of a scientific nature (publication list)
Detailed information about the position can be obtained by contacting: Group Leader Pawel Burkhardt, tlf.: +47 55 58 43 57, email: Pawel.Burkhardt@uib.no
The state labour force shall reflect the diversity of Norwegian society to the greatest extent possible. Age and gender balance among employees is therefore a goal. It is also a goal to recruit people with immigrant backgrounds. People with immigrant backgrounds and people with disabilities are encouraged to apply for the position.
We encourage women to apply. If multiple applicants have approximately equivalent qualifications, the rules pertaining to moderate gender quotas shall apply.
The University of Bergen applies the principle of public access to information when recruiting staff for academic positions.
Information about applicants may be made public even if the applicant has asked not to be named on the list of persons who have applied. The applicant must be notified if the request to be omitted is not met.
The successful applicant must comply with the guidelines that apply to the position at all times.
Cell fate commitment relies on both activation of appropriate genes and suppression of inappropriate ones. Polycomb group proteins are known to be crucial epigenetic silencers of developmental genes, but the manner by which they control fate in vivo, and the relative roles of different Polycomb proteins in silencing, have remained unclear. A new paper in Development tackles this problem using the Drosophila eye a developmental model – we caught up with authors Jinjin Zhu and Justin Kumar, Professor of Biology at Indiana University in Bloomington, to find out more.
Jinjin and Justin
Justin, can you give us your scientific biography and the questions your lab is trying to answer?
JK I started my career in Drosophila eye development while I was an undergraduate in the laboratory of Karl Fryxell at the University of California, Riverside. He was a wonderful mentor and it was my time in his lab that convinced me that I wanted to be a professor one day. While I was in Karl’s lab, I read Don Ready’s seminal paper on the morphogenetic furrow (Ready et al., 1976) and was totally amazed by the cellular mechanism of pattern formation. From the images in the paper, I could see a field of undifferentiated cells being transformed into the periodic units of photoreceptor clusters right before my eyes. I knew then that I wanted to join his lab for my PhD studies. Being in Don’s lab at Purdue University as a graduate student was a privilege. He taught me how to love the fly eye for its own sake and to appreciate its intrinsic beauty.
I then went on to do my post-doctoral fellowship with Kevin Moses initially at the University of Southern California and then at Emory University. It was in Kevin’s lab that I finally settled on the research questions that still drive me today. I stumbled on what I thought to be an astonishing phenotype. Quite by accident I discovered that manipulations of the Notch and EGF Receptor signalling pathways led to the homeotic transformation of the eye into an antenna. While Hox mutants change entire body segments, I was able to observe fate transformations occurring within a single imaginal disc. When I joined the faculty of Indiana University I set out to understand how the fly eye-antennal disc is first set apart from the other discs (i.e. leg, wing, haltere, genital) and then how it is later subdivided into distinct territories such as the compound eye, ocelli, antenna, maxillary palps, and head epidermis. Over the years my research group has discovered that while gene regulatory networks promote fate specification, growth, and patterning, they also influence development by repressing alternate and inappropriate tissue fates. My current interests are to understand how the retinal determination gene regulatory network cooperates with signalling pathways and epigenetic complexes to prevent the eye from adopting distant fates such as the wing and more local fates such as the head epidermis.
And Jinjin how did you come to be involved with this project?
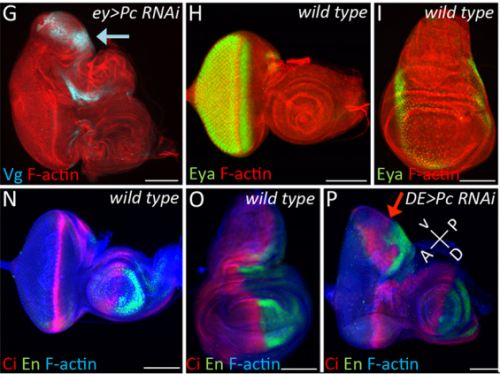
JZ I was really interested in the fate transformation caused by mis-expression of Hox genes when I was in college. When I joined the Justin’s lab, I did a genetic screen to find upstream regulators of eyeless in the developing eye disc. I knocked down eyeless and Sfmbt together and found this amazing eye-to-wing transformation phenotype. Meanwhile, Ali Ordway (second author) joined the lab and decided to screen other PcG proteins. She knocked down Pc and saw similar phenotypes. We were both fascinated by the images of these chimera tissue, in which the dorsal part of the eye disc turned into a wing disc while the ventral part remained as an eye disc, so we decided to figure out what happened to these mutant discs.
A selection of eye and wing discs from Fig. 1 in the paper.
How did you come to be interested in the phenomenon of transdetermination?
JK I got interested in transdetermination purely by accident. When I joined Don’s lab in 1991, I made sure that I read every paper that he had published. In one of his papers, he and Ricky Lebovitz had transplanted eye-antennal disc fragments into host larvae and then recovered the tissue after the host had emerged as an adult (Lebovitz and Ready, 1986). These experiments were aimed at understanding if the morphogenetic furrow was pushed or pulled across the eye field. I was really intrigued by the disc transplantation method used in his paper so I started reading about the history of this method, which I learned was pioneered by Beadle and Ephrussi in the 1930s. In the course of these readings I came across the work of Ernst Hadorn. It was from his papers that I learned about the concepts of determination and transdetermination. When Jinjin and I saw the eye to wing transformation, it reminded me of the fact that Hadorn remarked that the eye could only transdetermine into a wing. And it was at that point that I realized that the loss of PcG and Pax6 that we noticed gave the eye to wing switch might be the molecular explanation for the eye to wing transdetermination event that Hadorn observed many decades ago.
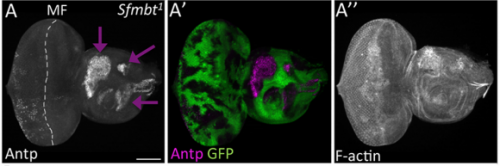
Third instar larval eye-antennal disc following Pc RNAi, from Fig. 2 in the paper.
Can you give us the key results of the paper in a paragraph?
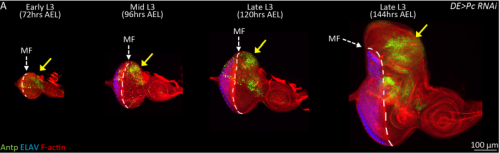
JK In this paper we demonstrate that the Pax6 transcription factor and the Polycomb group (PcG) of epigenetic silencers prevent the eye from adopting a wing fate. This decision is made early in development during the last stages of embryogenesis and during the first larval instar. The eye transforms into a wing because the chromatin around the Antp locus fails to be compacted thereby allowing for its activation by the zinc finger transcription factor Teashirt (Tsh), which is normally expressed in and required for the specification of the eye. Our findings suggest that in addition to promoting the primary fate of a tissue or organ, gene regulatory networks must play defense and suppress the activation of inappropriate selector genes and/or entire gene regulatory networks. This repressive activity appears to require cooperation from epigenetic silencing complexes such as PcG.
Ectopic wings in Pax6 / Sfmbt double knockdown flies, from Fig. 5 in the paper
Why do you think the dorsal region of the eye might be more susceptible to wing transformation than the ventral region?
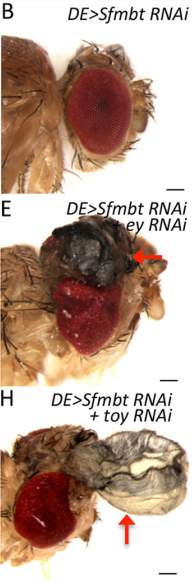
JZ This is a very interesting question and I think the answer lies down to the gene regulatory network controlling dorsal eye disc. 1) Wingless (wg), which is normaly required for wing development, is expressed in a higher level in the dorsal eye disc than in its ventral compartment. 2) engrailed (en) and cubitus interruptus (ci), which regulates A/P compartment of the wing disc also meets in the dorsal portion of the eye disc. Thus, the expression pattern of the endogenous genes in the dorsal eye disc contributes a lot to the eye-to-wing fate transformation. In addition, the dorsal eye disc is more susceptible to adopt wing fate because the level of Pax6 is significantly lower in the dorsal eye disc, especially in the region which will become the future dorsal head capsule. We have demonstrated that eye disc is more resistant to loss of Sfmbt function than the antennal disc. This is very likely due to the presence of Pax6 in the eye progenitor cells, because simultaneously knocking down Pax6 and Sfmbt induces the fate transformation.
It must have been particularly satisfying to discover the molecular underpinnings of phenomena first described in the pre-molecular biology age?
JK Yes, it was indeed satisfying to provide a modern perspective to an old problem/observation. In general I really enjoy reading the literature from the pre-molecular age – that is how I got interested in transdetermination in the first place. I also like scour the older literature for problems in eye development that were once studied but later abandoned due to the lack of the right genetic or molecular tools. If you look carefully enough, there is a wealth of such studies out there that are ripe for a modern perspective. For example, one of the exciting new areas of research in my laboratory today is the role that the peripodial epithelium, a tissue that overlies the eye-antennal disc, plays in development. There were several papers in the 1970s that suggested that its role was limited to the fusion of the two eye-antennal discs during pupal development. However, between 2000-2002 several laboratories provided evidence that signalling existed between these two tissues. But very little has been done since on this topic. Now several researchers in my lab are looking at the role that transcriptional networks in the peripodial epithelium play in promoting the fates of the eye-antennal disc.
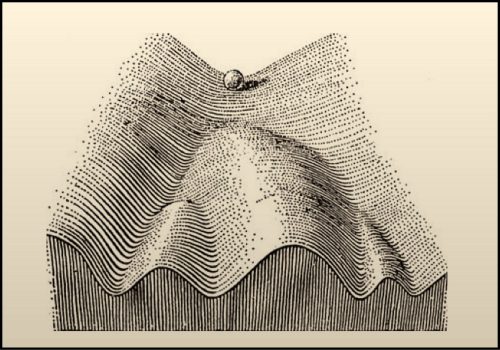
I also like how researchers of times past thought about development. The Epigenetic Landscape Model by C.H. Waddington is one of my favourite examples. To represent the process by which individual cells make fate decisions, he proposed that a cell can take different paths during development with each path representing a journey towards a unique fate. He drew a diagram to represent this idea – in his drawing a ball rolling down a mountainside presented a cell on its way to adopting a final fate. This drawing and the concepts that it represents is very inspiring to me. One can think about how to push cells developmentally down different trails or how to push the cells uphill (de-differentiation) and then down another trail (fate reassignment). For my own work, I try to think of the ball not as a single cell. Instead, to me it represents the entire eye-antennal disc. My lifetime goal is to figure out how the eye-antennal disc is guided down the mountain and how it ultimately gives rise to five distinct tissues and organs.
Sfmbt loss-of-function clones in eye-antennal discs, from Fig. 5 in the paper
When doing the research, did you have any particular result or eureka moment that has stuck with you?
JZ Yes. For a long time, I couldn’t figure out why the eye disc adopts a wing fate but not any other tissue fates. I know it is likely due to the de-repression of Antp in the developing eye when PcG activity is impaired. However, why is Antp being activated when the epigenetic silencers are removed? This question was solved when a piece of data came back from another ongoing project, in which I found that over-expression of teashirt (tsh) was able to rescue the headless phenotype of Pax6 double mutant (Zhu et al, 2017). In those flies, little wings or thoracic bristles were found in the rescued head cuticle, so I wonder the endogenous expression of tsh might be the transcriptional activator of Antp when Pc is removed from the eye disc. I did an experiment to knockdown Pc and Tsh (Figure 3) at the same time and it turn out to be true. Tsh is required to activate Antp during the fate transformation. Although we do not know whether Tsh directly turns on Antp transcription or not, but at least we found some underlying mechanisms of the homology between the eye and the wing. I think the lesson here is to have multiple projects going on at the same time.
And what about the flipside: any moments of frustration or despair?
JZ For me, the frustration in this project is that isolating eye discs at early stages, such as in 1st and early 2nd larval instar, is almost impossible. Thus, we couldn’t verify our final model of PcG proteins being recruited to the genome at these specific stages using ChIP-Seq. I hope the techniques will advance and allow us to reveal how exactly these epigenetic regulators function in vivo.
What next for you Jinjin – I hear you’ve moved to Harvard?
JZ Yes, for my postdoctoral training, I will work with Dr. Robert Kingston, focusing on how PcG proteins control gene expression in mammalian system. The PcG proteins in mammals are much more complicated than in Drosophila. Different PcG complexes have multiple variants and each protein have multiple homologs. Thus, a diversity of possible mechanisms that might be used to generated a repressive state of gene expression, such as histone modification, chromatin compaction and higher-order genome organization. I think the eye-to-wing transformation project has brought me here, but I want to learn more about the underlying mechanisms of epigenetic regulators on the molecular level for my future research.
Where will this work take the Kumar lab?
JK My research group has started studying how the interplay between transcriptional networks and epigenetic complexes such as Polycomb, Trithorax, and SAGA controls fate specification within the eye-antennal. These studies build upon the findings of our paper described here in Development in which Pax6 and PcG proteins cooperate to repress wing fates from being adopted in the developing eye. Currently, we have evidence that Pax6 cooperates with Trithorax and SAGA complexes to control the number of antennae that are produced. We also have evidence that these same complexes work together to prevent the duplication of the entire eye-antennal disc. I am very excited about these preliminary findings and I think my lab, for the near future, will be focused on using the tissues within the eye-antennal disc to revisit several very basic questions – how does an organism control the fate, number, and placement of all of the organs that it needs for survival.
Finally, let’s move outside the lab – what do you like to do in your spare time?
JZ I travel with my husband during holidays and we have been to many national parks in US. We both like photographing wild animals. At home, I usually play piano and computer games if I don’t need to collect fly embryos.
JK When I am not in the lab or my office I enjoy playing tennis. It is a terrific sport and a good outlet to release stress. I try to mix tennis and work as well whenever possible. When I am at home working on a paper or grant, I have the Tennis Channel on in the background. If there is an important match, I will stream it on my work computer as well. I also take my camera wherever I go and enjoy photographing wild life and outdoor scenery.
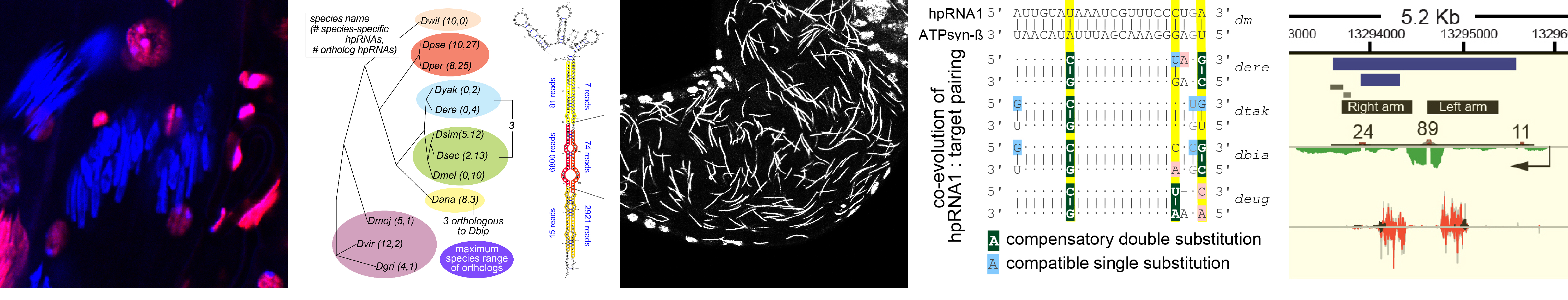
We have an opening for a postdoctoral position that will address fundamental questions in small RNA biology, genomic conflict, and speciation.
What is endogenous RNAi utilized for? We earlier described a mysterious class of endo-RNAi substrates termed hpRNAs (Okamura Nature 2008), and recently recognized that these mediate adaptive gene regulation in testis (Wen Molecular Cell 2015). These data open a window on a vital biology of RNAi, and now lead us to explore the evolution and function of RNAi systems across the Drosophilid phylogeny.
We discovered networks of rapidly evolving RNAi substrates we hypothesize resolve intragenomic conflicts, and successfully started to use CRISPR/Cas9 in non-model fly species to test some of these ideas (Lin 2018, in submission). Amazingly, while endogenous RNAi was not historically thought to have much phenotypic impact, we find that rapidly evolving genomic battles are being waged and critically depend upon the powerful weapon of RNAi silencing to propagate the species. Reciprocally, we are fascinated to understand by what novel molecular mechanisms de novo selfish meiotic factors can paradoxically drive population extinction.
We seek a motivated postdoctoral fellow with strong Drosophila molecular genetics experience and interest in integrating genome engineering, biochemistry, and bioinformatics to analyze the evolution and function of RNAi silencing systems in resolving deleterious intragenomic conflicts in testis. Although our entry point is RNAi biology, we anticipate that selfish factors yet to be discovered may mediate their effects through chromatin mechanisms.The successful candidate will integrate into a team that is actively engaged in diverse topics in gene regulation in Drosophila and mammalian models, and the Sloan-Kettering Institute provides a vibrant research community.
Funded position with housing and medical benefits are available immediately. Please provide CV, motivation letter and references to Eric Lai, laie@mskcc.org.
Relevant recent papers on hpRNAs and testis post-transcriptional regulation.
see also https://www.mskcc.org/research-areas/labs/eric-lai
Lin, C.-J., F. Hu, R. Dubruille, J. Wen, J. Vedanayagam, P. Smibert, B. Loppin and E. C. Lai (2018). The hpRNA/RNAi pathway is essential to resolve intragenomic conflict to preserve balanced sex ratio. Submitted.
Mohammed, J., A. Flynt, A. Panzarino, M. Mondal, M. DeCruz, A. Siepel and E. C. Lai (2018). Deep experimental profiling of miRNA diversity, deployment, and evolution across the Drosophila genus. Genome Research28: 52-65.
Kondo S., J. Vedanayagam, J. Mohammed, S. Eizadshenass, L. Kan, N. Pang, R. Aradhya, A. Siepel, J. Steinhauer and E. C. Lai (2017). New genes often acquire male-specific functions but rarely become essential in Drosophila. Genes and Development31: 1841–1846. (Highlighted in Genes and Dev 31: 1825-1826.)
Lin, C.-J., J. Wen, F. Bejarano, F. Hu, D. Bortolamiol-Becet, L. Kan, P. Sanfilippo, S. Kondo and E. C. Lai (2017). Characterization of a TUTase/RNase complex required for Drosophila gametogenesis. RNA23: 284-296.
Wen, J., H. Duan, F. Bejarano, K. Okamura, L. Fabian, J. A. Brill, D. Bortolamiol-Becet, R. Martin, J. G. Ruby and E. C. Lai (2015). Adaptive regulation of testis gene expression and control of male fertility by the Drosophila hairpin RNA pathway. Molecular Cell57: 165-78.
We are at the National Institute of Child Health and Human Development (NICHD) at NIH, Bethesda, Maryland USA. Our lab is interested in understanding cell lineage differentiation, gene regulation and how non-coding DNA elements and the 3D architecture of chromosomes contribute to these processes during early mouse development.
You share our enthusiasm for epigenetics, gene regulation, nuclear organization and mouse development.
You have PhD-experience in one or more of the following: mouse development, mouse genetics, epigenetics, massively-parallel sequencing techniques or computational biology.
What we offer:
Fully-funded postdoc positions.
Opportunity to start your own research program or lead ongoing projects.
2 paragraph cover letter explaining your scientific trajectory and why you would like to join us.
CV and email contacts for 3 references.
The NIH is dedicated to building a diverse community in its training and employment programs.
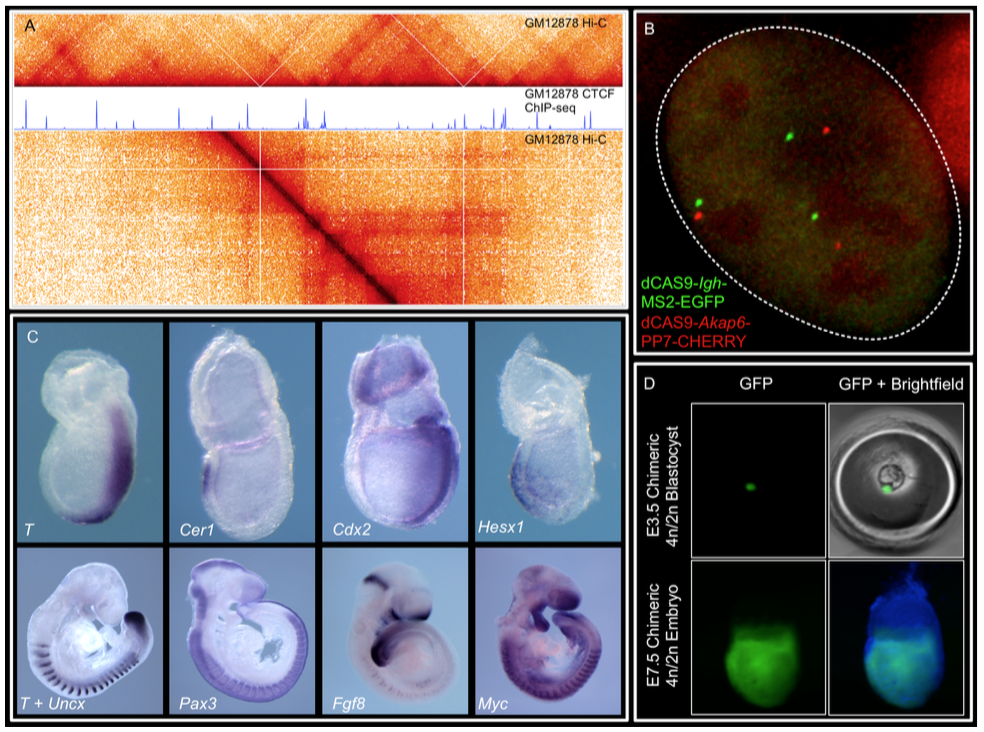
We combine imaging techniques in both fixed and living cells with sequencing- based genomic techniques that assess DNA-DNA interactions. (A) Hi-C and CTCF ChIP- seq of GM1278 cells (B) dCAS9 MCP-EGFP and PCP-CHERRY live imaging of the Igh and Akap6 loci. The mouse embryo is an unparalleled system in mammalian biology for understanding how tissue- specific gene expression is achieved. (C) Whole mount in-situ hybridization for patterning markers in mid and late gastrulating embryos. (D) Tetraploid aggregation with GFP ES cells allows generation of fully ES-cell derived embryos.
In a previous blog, I have disgraced parrots by associating them with P-values and discrediting them for their mechanic repetition. Nevertheless, I admire the vivid colours of these multifaceted birds. Here, I want to make it up by dedicating a pseudo-colour look-up table (LUT) to parrots.
The images produced by fluorescence microscopy are best displayed in grayscale for optimal contrast, using a linear relation between gray-level and pixel intensity (footnote 1). However, the number of gray levels that can be distinguished by humans is orders of magnitude smaller than the number of colours that can be discerned. Therefore, pseudo-colours (also often named false colours) can be used to achieve better contrast and convey a finer level of detail (Sheppard et al., 1968), at the cost of loosing the relation between intensities in the image and the actual pixel value. Thus, the choice between grayscale representation and pseudo-colour will depend on the information that is visualised.
A monochrome digital image can be converted into a pseudo-colour image by replacing each pixel value with a colour (Fink, 1976). The conversion is defined by a so-called look-up table (LUT). The LUT contains 256 RGB codes that define a colour for each of the pixel values between 0 and 255. The combinations are basically endless. A collection of LUTs for cartography, technical illustration and design is available here and the popular image processing software ImageJ/FIJI comes with a set of LUTs.
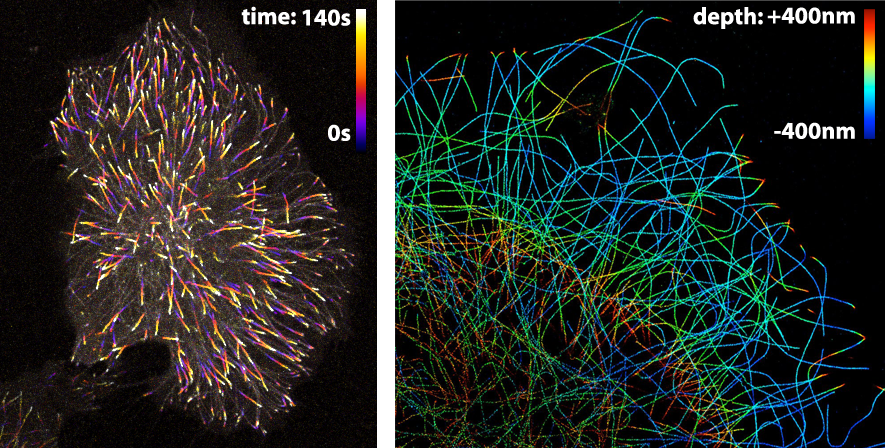
The LUT that is applied to an image largely depends on the information that one wants to visualise. Since the LUTs are critical as to the level of detail that can be discerned, careful selection is important. Here, I highlight different LUTs in the context of fluorescence images. Besides fluorescence intensity, several other parameters can be encoded by colour. Examples are (i) temporal encoding (figure 1 left), (ii) depth encoding (figure 1 right, and see several other examples here), and (iii) encoding of a spectroscopic parameter, e.g. fluorescence lifetime (Joosen et al., 2014).
Figure 1: Pseudo-colours are used (in the left panel) to encode time to show the dynamics of microtubule plus ends or (in the right panel) to encode depth in a 3D STORM image of microtubules. Image credits: Timelapse movie of EB3-mNeonGreen by Anna Chertkova, 3D STORM image by Christophe Leterrier.
Here, I will give examples of pseudo-coloured images that were obtained by FRET ratio-imaging. In FRET ratio-imaging the response of a biosensor is monitored by emission ratio-imaging. Both the spatial differences and the changes over time can be of interest. The spatial differences are usually presented with pseudo coloured maps of the FRET ratio. Movies of these FRET maps are generated to depict the ratio changes over time.
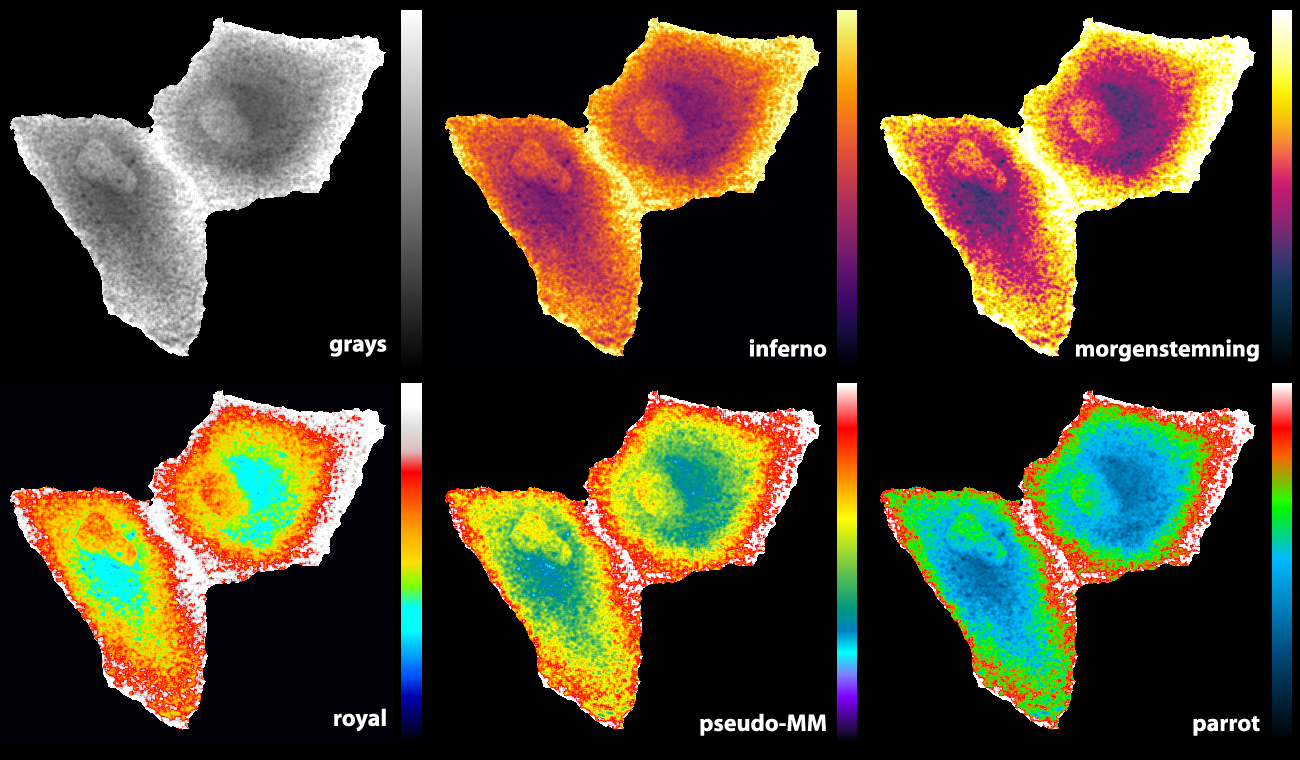
The ‘Fire’ LUT that comes with ImageJ works well for temporal colour coding (figure 1, left panel). A variant of the Fire LUT was generated and named ‘MorgenStemning’. This LUT is colourblind friendly and has a linear increase in luminance (Geissbuehler and Lasser, 2013). Similarly, several “perceptually uniform” colour palettes were generated that are also colourblind friendly. These LUTs, ‘Inferno’, ‘Viridis’, ‘Magma’ & ‘Plasma’, are described here. The Inferno LUT is a good choice, since it starts with black at zero intensity. This LUT does, however, not use white for maximal intensity, thereby not providing maximal contrast.
Some of the LUTs that we previously used are the ‘royal’ LUT of ImageJ (Unen et al., 2015) and a pseudocolor LUT that is part of MetaMorph software (Reinhard, 2016). Since I was not entirely satisfied with the existing LUTs, I designed a new LUT (footnote 2). The basic features are that it starts with black and ends with white to achieve maximal contrast between the minimal and maximal pixel values. The false colours that correspond with lower half of the pixel values does not have a red component to get a cyan colour which blends into bright green. For higher pixels values the false colours turn from green via orange and red to white. As the colours in the LUTs reminded me of parrots, I call this LUT ‘parrot’. Figure 2 shows how the parrot LUT compares to other LUTs. The parrot LUT is colour-blind-friendly (footnote 3), but it is not perceptually uniform and is suboptimal in this respect.
Figure 2: An overview of different LUTs applied to the same grayscale image that displays Rho GTPase activity in endothelial cells (Reinhard, 2017). The image and LUTs are available here.
Final words
The choice of a LUT is often based on a qualitative assessment of how it visualises the information. By trying a number of LUTS, the LUT that best conveys the information can be selected. Therefore, it useful to have a broad panel of LUTs to choose from and the parrot LUT is just another option. I encourage designing and sharing of LUTs (either as supplemental data to go with a paper or on data sharing platforms – footnote 4) to increase the number of options and the chances of finding the right LUT.
Acknowledgments: Thanks to Jakobus van Unen for tracing back the origin of the ‘pseudo-MM’ LUT to metamorph and to Marten Postma for making the LUT editor available.
Footnotes
Footnote 1: Achieving a true linear relationship between pixel value and displayed grayscale value may be impossible and irrelevant.
Footnote 2: The parrot LUT was created with a MatLab based LUT editor that was generated by Marten Postma
Footnote 3: I have a colour vision deficiency that prevents me from distinguishing blue from purple and bright green from yellow. This is the main reason to remove most of the red component and only have a limited band with a green/yellow component. Since I designed it, it is intrinsically colour-blind friendly. However, as there are several types of colour vision deficiency it may not work for everyone.
Footnote 4: The LUTs and related material described in this blog are available at Zenodo (Doi: 10.5281/zenodo.1211690).

 (2 votes)
(2 votes)
















 (No Ratings Yet)
(No Ratings Yet)