I joined Dr. Craig Albertson’s lab as a graduate student in 2009, where I quickly became fascinated by these cute cichlid fishes. They’re colorful, they breed their young in the mouth, and some of them have funny looking faces like this blue mbuna (Labeotropheus fuelleborni):
My research started on the genetic control of bone development in these cichlids, trying to understand the development and evolution of their facial shape. We soon discovered that the ptch1 gene, a member of the Hedgehog signaling pathway, plays a major role in shaping multiple bones in the cichlid head. In brief, some cichlid species, such as the blue mbuna above, have one version of the ptch1 gene that leads to a facial skeleton suitable for scraping algae off the surface of rocks; some other cichlid species have a different version of the ptch1 gene which leads to a slightly different skeletal shape that make them good at capturing evasive prey via suction feeding. These findings add up to a fast-growing body of knowledge about the genotype-phenotype map, which has been of great interest to biologists for decades.
While I’m proud of my contribution as a young scientist, one thought often haunts me: although I call it a “major role”, the genetic difference in ptch1 only accounts for 10-20% of the variation in bone shape. The vast majority of the variation remains to be explained, what’s going on there?
From a geneticist’s point of view, this is exactly what you would expect to find in a trait as complex as the shape of facial skeleton: there’re probably many genes working together to produce the final phenotype. Some genes, like ptch1, can have a relatively large effect and will pop out in our analysis. But most of genes will have such a small contribution that could be easily swamped by all kinds of background noise and therefore, would be very difficult to detect. Considering the size and life history of cichlids, it’s logistically impossible for me to trace down those genes.
How about environmental factors? Genetics is certainly not the only player, I should take a detour.
I was really intrigued by a plasticity experiment in the lab, where we fed the cichlids with different food and induced skeletal changes in the face. And I started wondering, is there any other behavior that can also change their bones?
Then, the gaping behavior of cichlid larvae quickly caught my eyes.
6 day old cichlid larvae gaping
The cichlids we study are mouth brooders. Mom usually keeps all her eggs in the mouth for several weeks. But in order to study their development, we will extract the embryos from the female’s mouth and raise them in a flask. This is one of the first skills I learned when I joined the lab, and I noticed this gaping behavior right away – well, dozens of cute little baby fish gaping their mouths is kind of hard to miss. I didn’t pay much attention to this behavior at that time. And I think probably most people would have thought the same thing when they first see the video above – these larvae are just doing their normal fishy thing, they’re just breathing. It looks cute, but that’s pretty much it.
However, when I looked at this behavior with the new perspective, I soon realized its bizarreness: they were gaping really fast. Some of them would gape more than 250 times in 1 minute, so fast that I could barely count it in real time. And I had to count in Chinese, English numbers were just too long for me. That felt like a lot of gaping, were they really just breathing? A quick literature search said no: fish larvae at such a young age rely mostly on their skin for breathing. I was excited to find this out. Initially, I thought maybe this larval breathing behavior could influence bone development as a side effect. Now I got a new idea: perhaps it’s not about breathing at all. They might actually be working out their mouths to stimulate bone development.
As a graduate student, the first thing you need to do when you have an idea like this is to convince your adviser that you’re not crazy, that it’s a legit scientific hypothesis. So, I started to collect some preliminary data, which turned out to be encouraging: I found that the larvae of the blue mbuna, who has a longer jaw bone called the retroarticular process (RA), gaped at a higher frequency than another species with short RA. This is exactly what I expected to see, more gapes more bones.
At this point, both Craig and I became very interested in this hypothesis, and started to brainstorm ways to test it. We thought about all kinds of ways to manipulate the gaping behavior, from varying environmental factors like oxygen and temperature, to chemical treatment like caffeine and tricaine (commonly used for anesthesia in fish), to more goofy ideas like botox and gag. Most of them were rejected either because they would cause an overall impact on development, or simply not feasible in our fish. In the end, I decided to take a rather adventurous route, that was to surgically cut the ligament attached to the RA. We liked this approach because it’s a local and targeted experiment: the surgery would only attenuate the mechanical stimuli being applied directly to the RA without affecting the other bones and muscles that participates in the gaping behavior. The surgery worked beautifully: when I cut the ligament in the larvae of the fast gaping species, they developed a shorter RA.
The surgery was sort of a loss-of-function experiment, and we also did a gain-of-function experiment that originated from what I call a happy accident in the lab. One day I ran out of standard containers for the larvae when I was monitoring their gaping behavior, so I had to substitute with small beakers. Then I found that when restricted to a smaller environment, the larvae will gape at a higher frequency. Although it ruined my gape counts for that brood, I took advantage of this phenomenon and tried to induce gaping behavior in the larvae of slow-gaping species with smaller container. Just as I expected, they developed longer RAs.
These two experiments suggested that the seemingly trivial gaping behavior of cichlid larvae affected the development of their RA development, and the effect size was, interestingly, on par with what we found with genetic difference in the ptch1 gene. But what’s more interesting, is that we found larvae that gaped more is expressing more ptch1.
Taken together with our previous studies, we think the Hedgehog signaling pathway plays a dual role in shaping the facial skeleton. On one hand, genetic differences in the ptch1 gene leads to differential bone growth in the facial skeleton; on the other hand, mechanical stimuli also induce bone growth via Hedgehog. At this point, we’re curious about how these mechanisms evolved. It will be super cool if the epigenetic (or plastic, whichever way you prefer to call it) route came in place first, and genetic mutations came later to fix the difference – genes as followers.
And for me personally, I really want to learn the larval gaping behavior in nature. My observation of the gaping is done in beakers and Petri dishes after all. And I just can’t stop thinking what the larvae are doing in their mother’s mouth, which is a much smaller environment than my restriction experiment. Do larvae gape even faster? Does the mom modulate their gaping? Or perhaps they don’t gape at all? These questions won’t help us cure cancer. They’re not major challenges with great impact in academia or industry. I’m just curious, simple as that, which is exactly why I love doing science.
Curator position at Xenbase, the Xenopus Model Organism database
Xenbase (www.xenbase.org) is the Xenopus bioinformatics and genomics resource. Xenopus is a major model for fundamental cell and developmental biology and a model for human disease. Xenbase is a totally free, and globally accessible database, used by Xenopus researchers worldwide, and is funded by the National Institute of Child Health and Human Development. Xenbase has two performance sites: the curation team is based in Cincinnati, OH (PI: Dr. Aaron Zorn) and the developer/database management team is based at the University of Calgary in Canada (PI Dr. Peter Vize).
Xenbase is seeking to fill 1 full time Curation position to join the curation team at the Division of Developmental Biology, Cincinnati Children’s Hospital, Cincinnati, OH, USA. Curation positions offer a challenging job away from the wet-lab and research bench, where interpreting, annotating and displaying complex data is our main task. Curators also develop strategies to improve data curation; work to improve data display/querying on the website; interact with our user community at research conferences; develop programming skills; and contribute to Xenbase publications.
Job Description:
• Curation and annotation of published Xenopus scientific literature, focusing on gene expression and the extraction of other research data: genes, transgenic constructs, antibodies, morpholinos, phenotypes, genetic interactions, gene product functions and models of human disease.
• Import and annotate data from large-scale screens (e.g., loss-of-function morpholino screens, gain-of-function mRNA screens).
• Help develop new features: curation and processing of public and directly submitted RNA-seq and ChIP-seq NGS data from Xenopus experiments, curation of mutant phenotypes and transgenics; implementing GO annotations.
• Co-author reports and publications, and give presentations at national and international meetings/workshops.
Qualifications:
• MSc or PhD degree in bioinformatics and/or developmental biology, genomics, genetics, molecular biology, zoology, anatomy or related field.
• Demonstrated ability to produce scientific papers, reports and presentations
• Demonstrated ability to work in a team as well as independently, efficiently (i.e., both quickly and accurately) and be self-motivated
• Strong interpersonal and communication skills, including excellent written and spoken English.
Preference will be given to an applicant with:
• Experience with a bioinformatics, genomics or model organism database
• Experience in data annotation/biocuration, knowledge of relational databases, and familiarity with ontologies.
• Experience in Xenopus or other vertebrate (mouse, zebrafish or chick) developmental biology.
• Experience in analyzing genomics data, using GRN software, genome browsers and common bioinformatics tools.
How to Apply:
Please submit your application to: aaron.zorn@cchmc.org
with the following information:
• A cover letter, including a statement of interest/purpose
• CV/Resume.
• Copy of your degree(s).
• List 3 references/referees whom we may contact (please include their postal address, email and phone number).
Salary and Start Date:
Salary will be commensurate with qualifications and experience. Start date is negotiable, but expected to be in late 2017.
The successful applicants will be employees of Cincinnati Children’s Hospital and will undergo background checks, orientation and a 6-month probationary period. Employees are required to receive an annual flu vaccination.
More information about working at Cincinnati Children’s Hospital, and living in Cincinnati, can be found here: http://www.cincinnatichildrens.org/careers/working/default/
This is an exciting opportunity for a skilled research technician/assistant to join the Ear Repair Group at the National Institute for Health Research (NIHR) Biomedical Research Centre (BRC) at Guy’s and St Thomas’ and King’s College London. The Ear Repair group is led by Professors Andrea Streit, Karen Steel and Abigail Tucker, based in the Centre for Craniofacial & Regenerative Biology and the Wolfson Centre for Age-Related Diseases at King’s College London. The successful applicant will be employed by Guy’s and St Thomas’ NHS Trust.
The successful applicant will have advanced knowledge of cell and molecular biology techniques and have significant previous experience in a research laboratory environment. They will provide technical support for the research programme relating to ear repair and regeneration. This will include studies on drug design and delivery to different compartments of the ear, testing their uptake and efficiency as well as their influence on cell restoration and repair. Please note that previous experience in a clinical setting is not required for this role.
A position as postdoctoral research associate is available in the Ear Repair group within the NIHR Biomedical Research Centre at Guy’s and St. Thomas’ NHS Foundation Trust and King’s College London. The Ear Repair group is led by Professors Andrea Streit, Karen Steel and Abigail Tucker, and is part of the BRC Oral Health Theme based in the Centre for Craniofacial and Regenerative Biology at King’s College London. The Centre is one of the leading centres for Craniofacial Biology and offers a vibrant research environment in the heart of London.
We are looking for an enthusiastic and highly motivated candidate holding a PhD in biomedical science or a related field with a background in molecular biology, mouse genetics, neuroscience and/or developmental neurobiology. Understanding of and expertise in ear development and physiology will be an advantage, as will experience in drug delivery strategies. The project investigates the mechanisms underlying ear homeostasis and hearing loss, looking at new ways of restoring function, with the long term aim to design new strategies to treat hearing loss. The successful candidate will have training and expertise in one or more of the above areas, an excellent academic track record, be committed to a scientific career and will be keen to work in an interdisciplinary team. The project itself offers many training opportunities in cutting-edge biomedical science, as well as in different transferrable skills.
To apply for this post go to https://www.hirewire.co.uk/HE/1061247/MS_JobDetails.aspx?JobID=77419
Just over two years ago, while I was a Masters of Neuroscience student at University College London, I became interested in the emerging concepts of brain lymphatics and sleep dependent macromolecule clearance within the brain. Until quite recently, the brain was considered to be without lymphatic vasculature or dedicated mechanisms of clearance of its own. Then in 2013, Lulu Xie of the Nedergaard lab published a blockbuster paper in Science that suggested that sleep facilitates fluid and metabolite clearance from the brain (Xie et al., 2013). Following in 2015, a double header in Nature and The Journal of Experimental Medicine (Louveau et. al. 2015, Aspenlund et al. 2015) demonstrated that lymphatic vessels exist within the dura mater of mouse and human meninges. Together these papers opened an entirely new outlook for how our energy intensive brain might maintain homeostasis, with potential implications for neuropathology and degenerative disease. I was attracted to the interdisciplinary aspects of this budding field, and the topic became a fundamental obsession for me after several involved discussions with Dr. Jeff Illif who had written the original brain paravascular clearance paper (Illif et al., 2012) and moved from the Nedergaard lab to head his own at Oregon Health and Science University in 2013. I was deeply encouraged by the time he took to act as a sounding board and to offer mentorship and direction as I decided how to pursue this field for my PhD.
I had a potential idea for how to tackle this field from a new angle after Dr. Jason Rihel from UCL’s Department of Cell and Developmental Biology gave a lecture for the Master’s program, during which I discovered that his lab uses molecular and genetic techniques to study sleep using zebrafish. I found his approach and style as a creative, big-picture scientist appealing. Zebrafish also made sense to me as a model through which to investigate this complex clearance mechanism, as they are small and optically transparent throughout larval stages, allowing for accessible imaging and high throughput studies. However, when I approached Dr. Rihel (Rihel et al. 2010) with a PhD proposal to investigate these processes in zebrafish, he warned me that not only would this be a new area for his lab, requiring new methods, but we also might find that zebrafish simply lack brain lymphatics/clearance. To finally convince Dr. Rihel that it was worth investigating, I poured over these new papers along with zebrafish papers describing lymphatic development, many of which came from a well-known lymph and vascular lab in Muenster, Germany led by the seasoned scientist, Professor Stefan Schulte-Merker.
About the same time, in Professor Schulte-Merker’s lab, a postdoctoral researcher by the name of Max van Lessen had also caught the brain lymphatics ‘bug’. Digging through the same literature, Max had his own “eureka” moment when he realized that the transgenic vascular and lymphatic lines, including a few created by his colleague Andreas van Impel, might have labelled the brain. Such transgenic tools would allow for dynamic investigation of the origins and molecular properties of brain lymphatics, if they were present and conserved in bony fish.
Serendipitous timing brought Professor Schulte-Merker to our doorstep last March for an unrelated meeting with our colleague and neighbour, Professor Steve Wilson. When I heard from my colleague Dr. Eirinn Mackay that he was down the hall, I jumped up and caught him as he was heading out the door. Taking advantage of the brief moment, I thrust my hand at him for a hearty shake and I began excitedly outlining that I believed brain lymphatics found in mice and humans would be conserved in zebrafish. I told him that in some of the images from papers, I had seen evidence of lymphatic markers present in the head of the zebrafish and wanted to know his opinion. Professor Schulte-Merker smiled and nodded. “Indeed, we have noted this and are looking into it.“ I asked out of curiosity why had everyone in the zebrafish vascular community focused exclusively on the trunk of the fish? Most images of zebrafish lymphatics I found in the literature had frustratingly cropped the top of the head from full view. Professor Schulte-Merker smiled wryly and stated that “everyone was so engrossed with the peripheral system in the past that no one ever bothered to look up.” It struck me that so much of discovery is about simply “looking up”. Not being afraid to challenge long standing norms and established or popular avenues of thought to follow instincts that bring us to look with new eyes upon something we have seen before a billion times.
I also discovered that science is, more often than not, luck, happenstance, and the willingness to cooperate to get more done. We can work alone and hoard, or we can branch out and be, as my friend Seanne in California always likes to put it, “better together”. My PhD supervisor Dr. Rihel cultivates an atmosphere of collaborative research, and his fearlessness in sharing ideas and taking chances paved the way. The Schulte-Merker lab didn’t hesitate to reciprocate in the same spirit, and immediately a cooperation was forged. To begin parallel investigations into brain lymphatics, the Schulte-Merker lab sent us a combined transgenic line with kdr-l expression labelling all vasculature in mCherry (Hogan et al., 2009) and flt4 (Vegfr3), an established venous-lymphatic marker in mCitrine (van Impel et al., 2014).
We at the Rihel Lab have an interest in the conservation of these systems and mechanisms to ultimately elucidate the impact on potential clearance through sleep, and the Schulte-Merker lab thinks about this problem through the angle of vascular and lymphatic origin and development. We have complementary skill sets that allowed the work to begin at an incredible pace, building a whirlwind storyline. My colleagues often remark that this project made them feel like they had been strapped to a rocket ship.
The Mystery Cells
To help with the anatomical search for brain lymphatics, I recruited my remarkably talented second supervisor as a co-investigator (Dr. Tom Hawkins), as he is one of the most highly inquisitive people I have met, with a wide breadth and depth of technical expertise which would clearly be needed. We began by looking at adult transgenic brains, carefully dissecting and removing them using confocal microscopy to examine both the surface of the brain as well as the interior portions of the skull. What we saw initially was somewhat astonishing. We had imagined we might find lumenized vessels; however, along much of the surface of the brain were what looked to be flt4 positive cells of some sort, glowing brightly in mCitrine, clearly differentiated from the kdr-l blood vascular marker in brilliant mcherry. While kdr-l expression labels both arteries and veins, flt4 expression is an established marker of venous and and lympahtic endothelium. Importantly, our cells were entirely negative of kdr-l expression, which in the trunk can only be found in lymphatic vessels.
They possessed large inclusions and spindly processes which appeared to be connecting each to the other in close proximity to surface vasculature. A Sir Henry Welcome Fellow in our lab, Sabine Reichert, took one look at the images and without blinking stated, “they look like some sort of endothelial cell type.” It quickly became clear that she was right.
Brightly coloured Brain Lymphatic Endothelial Cells along blood vasculature (blue) on the surface of the zebrafish brain’s Optic Tectum (Photo: Shannon Shibata-Germanos and Tom Hawkins).
We dove in with immunohistochemistry using anti GFP and RFP antibodies followed by cryosections in adult brains to investigate carefully where these cells were positioned. Our cryosections revealed that these cells populated the meninges surrounding the entire brain exclusively and were associated with particular vessels diving into the parenchyma from the meningeal layer. This was an exciting moment for us as we recognized that the work done in mice and in humans had placed brain lymphatics in the meninges and our cells were positioned in a similar location. Although much work remained, we in London felt we were on track and may have identified a brain lymphatic system in the fish, suggesting evolutionary conservation within the vertebrate lineage.
Origins
Around the same time Professor Schulte-Merker and I had met, Max and the Schulte-Merker lab in Germany had found beautiful evidence of head lymphatics in the embryonic brain. Max had been capturing live images from the earliest stages of development onwards, patiently waiting to see when, how, and from where expression of these flt4 positive structures would emerge to get a hint of their developmental origin. They reasoned that if peripheral lymphatics sprout from venous structures early in development, perhaps the same would occur in the brain. While training the confocal microscope on the choroidal vascular plexus (CVP) and mesencephalic vein (MsV) which loops around the optic tectum of the zebrafish brain, they saw a clear sprout comprised of flt4 positive cells emerging around 56 hours post fertilization (hpf) from the CVP that then grew in an arc along the MsV at the edges of the tectum in parallel to the vasculature. Residual kdr-l expression in these flt4 positive cells suggested differentiation from venous endothelium.
Over the course of three days, this flt4 positive ring from what we began calling the stereotypical tectal “loop”, a cohort/string of loosely connected single cells which aligns closely to the MsV. In the peripheral nervous system of zebrafish, angiogenesis begins with the sprouting and development of arteries, followed by the concurrent development of veins and lymphatic vessels with both requiring three key genes: ccbe1, vegfc and flt4. Both lymphatic sprouts and venous sprouts initially originate from the posterior cardinal vein and ultimately form their own independent structures as they grow towards the myoseptum (van Impel et al., 2014). Since these novel brain cells sprouted from a vein and expressed flt4, Professor Schulte-Merker remarked that they were most likely lymphatic in origin.
Macromolecule Uptake
If these novel cells were in fact functionally similar to mammalian lymphatic structures, Max hypothesised that these cells may have function in the uptake and clearance of macromolecules, similar to what has been reported for murine brain lympahtic vessels. Incredibly, tracers injected either into the tectal neuropil or into the hindbrain ventricle accumulated at flt4 positive cells forming the tectal loop within minutes of injection. This uptake is so rapid and specific that when I first showed Dr. Rihel a similar dye injection experiments that my colleague Dr. Chintan Trivedi and I had performed on the two-photon microscope in a wild type fish, he thought I was showing him an image of the lymph transgenic line, as the dye took on the precise location and shape of the flt4 positive cells! With so much activity going on in both of our labs, we realized that it was time to have an in-person meeting to facilitate discussion and avoid excessive experimental overlap.
I made a pre-Christmas journey to Muenster to mutually share our findings along with new ideas and techniques. During my time there, I was also introduced to Andreas van Impel, another postdoctoral researcher in the Schulte-Merker lab, with a humble and steadfast demeanour who had created a number of the stunning complex lymphatic lines we were currently using. He brought a great deal of experience and insight to our discussions and grounded us with his thoughtful interjections. When Max presented his work, we both were so excited that we were wildly gesticulating and scribbling notes. At one point, one of us climbed atop of the table to point out a key piece of research from an old paper from the 1930’s, with Andreas calmly leaning in with an amused smile on his face. It was one of the most vibrant scientific visits I have had, and it solidified a strong sense of teamwork. Face to face contact I found is the cornerstone of collaboration. Trust must be earned and built, while looking one another in the eye, so colleagues can both sense and see our intentions, and know we are all labouring for the same goal.
Shortly after my visit, Max shared he had mused that the cells had macrophage or dendritic-like characteristics, even though their lymphatic origin suggested a novel cell type. This had Max hunting after shared receptors or other common characteristics, which may offer further clues about our cells. In this reading, Max discovered a universal pattern recognition receptor called the mannose receptor found in both macrophages and dendritic cells and was a major way in which cargo was endocytosed into these cell types. Thus, he decided to apply both Pyrimidyn-7, a quintessential blocker of dynamin-dependent endocytosis, and mannan, a bacterial polysaccharide that binds to and saturates the mannose receptor in independent experiments to see if he could stop tracer uptake. He shared on a call that he was left deeply puzzled as it looked like the tracers had still localized to the flt4 positive cells anyway, even in the presence of the endocytic blocker.
Dorsal view of the zebrafish brain at 5dpf. The two yellow rings are the early lymphatic endothelial cells in the brain surrounding the Mesencephalic Vein around the zebrafish brain’s Optic Tectum (Photo: Max van Lessen).
Fortunately, Max had also injected the pH sensitive dye (pHrodo-advin) which would only fluoresce when it encountered an acidic environment inside of a cell. In previous experiments, he had seen both the tracers and the pHrodo fluoresce at the flt4 positive cells together, demonstrating for the first time lymphatic uptake of macromolecules at the single cell level. Being in the habit of injecting pHrodo along with his usual tracer, he also used this dye cocktail in his search for an effective concentration of mannan or Pyrimidyn-7, which revealed that the usual fluorescent tracer still accumulated at the cells. However the injection of either blocker effectively stopped the internalization of pHrodo. It didn’t make sense! How could the fluorescent tracer be taken up, but the pHrodo not? Suspicious, he took a closer look, and realized that the usual fluorescent tracer had merely localized to the membrane of the cell and was not actually being internalized. 3D reconstructions revealed that the fluorescent tracers were lodged into the membrane. If pHrodo was injected before the blocker it still happily fluoresced in the acidic lysosomal compartments within the cell but never co-localized with the fluorescent tracers lodged in the membrane after endocytic blockage.
Lateral view of the zebrafish. In red are the newly discovered cells on the top of the Optic Tectum of the zebrafish brain (Photo: Max van Lessen).
Thus, it seemed that one process “docks” the tracer to the cell membrane while subsequent cargo internalization into acidic compartment is likely mannose receptor dependent. If it hadn’t been for his habit of using the pHrodo he may just have given up thinking the blockers hadn’t worked. I learned a few valuable lessons from Max’s results. First, I learned that experiments don’t have to be complicated and showy. They can often be incredibly straightforward. The other lesson was to always look closer and deeper before walking away from an experiment in frustration. So much of science emerges from happy accidents. We hear it again and again, and that is because it’s just so true.
Max and his colleagues expanded the analysis to additional lymph markers as well as markers for macrophages, etc. For example, they examined transgenic lines labelling classic lymphatic markers such as Lyve1 (Okuda et al., 2012) and Prox1 (van Impel et al., 2014) to further confirm these cell’s lymphatic identity. Much to our combined excitement, each lymphatic line labelled the quintessential tectal loop. Finally, to rule out that these flt4, prox1, lyve1 positive cells were macrophages, Professor Schulte-Merker suggested breeding the lyve1 line in DsRed (Okuda et al., 2012) with the mpeg1 macrophage line in GFP (Ellett et al., 2011). This showed clear differences between the cell types based on location, morphology, and expression. To completely eliminate the possibility, they also examined whether the morpholino knockdown of the macrophage transcription factor pu.1 had any effect on the lymphatic tectal loop. Andreas injected morpholinos, and after a tense few days of waiting for the results, the experiment revealed that our flt4 positive cells remained undisturbed, even as all macrophages were eliminated. We were elated, and we all finally allowed ourselves to believe that we had a novel cell-type on our hands.
Back in London, Tom and I began our hunt for the identity of these cells from a very different angle, using electron microscopy to obtain a definitive view of the location and ultrastructural features of these cells in relation to blood vasculature. Tom has had decades of experience with transmission EM, so he warned me not to get too excited, as disappointment was likely. Using the enormous TEM microscope with the assistance of Mark Turmaine: the quiet EM master, we happily navigated our way over the ultrathin slices in what, to me, was reminiscent of the broadcasts from the first moon landing. To our delight, and despite the forewarning, our first EM run was crystal clear with excellent structural resolution. The cellular landscape was wild and extraordinary, full of intricate interdigitations of membranes and strangeness all around. Guided by venerable EM studies on goldfish meninges, we could clearly identify a distinct meningeal tri-layer, inside which the cells that contained these large inclusions were exclusively found. We were able to immediately identify structures with multiple large inclusions in close proximity to surface brain vasculature with the size and distribution shown in our confocal images. They were clearly separable from perycites, vascular endothelial cells, and their basement membranes.
This was the day we captured the TEM (Transmission Electron Microscope) images at UCL.
Tom Hawkins hard at work cutting glass knives for TEM block cutting.
We combed over mammalian and teleost EM images of the meninges to try to identify these cells. One candidate was the Fluorescent Granular Perithelial Cells (FGP), which were discovered and characterized in rodents by Maso Mato in the 1980’s. Mato cells were known to reside in the meninges and contained numerous, heterogeneous inclusions (Mato et al., 1986, 2002), and we had several hours of heated debates in Dr. Rihel’s office as we stared at side-by-side comparisons of classic EM images and our new data. Ultimately, we decided that our cells were most probably distinct from Mato cells, or at least they represented a specific subtype, as our new cells’ inclusions were less heterogeneous than expected for Mato cells, and they had morphologically distinct shapes.
The ”Competition”
As the first part of our story of these cells was beginning to wrap up, Max and I were made aware that two other labs had come to similar conclusions to us. Suddenly, we were plunged into a race in which we weren’t precisely clear who had what, and when, in entirety.
Moments like these, when you find out other labs are working on similar avenues, can be stressful at best, and to be frank, Max and I had moments of panic together. The prospect of potentially being “scooped” looms heavy above all of our heads at any given moment. Each one of us is deeply connected to our research and have spent countless hours sweating and toiling over our projects. They become a part of us. And hearing that others have found something similar can be particularly disorienting as discovery, though made for the common good, can feel so deeply personal. Professor Schulte-Merker and Dr. Rihel kept insisting that each lab would be contributing to the scientific enterprise and bring novel insights into the nature of these cells.
This also meant that the community would be excited about this discovery. In retrospect, with the three respective papers now published, (Galanternik et al., 2017, Bower et al., 2017, van Lessen et al., 2017), it is clear that each lab approached these cells from unique perspectives. Galanternik et al. discovered these cells using a transgenic mannose receptor line and indicated that they believed these to be fluorescent granular peritheliaI cells (FGP or Mato Cells) (Galanternik et al., 2017). Bower et al. utilized the transgenic lyve1 lymphatic line to reach their discovery, and amongst other interesting findings they ablated these cells to find that the ablations impacted the integrity and development/regeneration of meningeal vasculature (Bower et al., 2017). Both those papers also performed valuable RNAseq analysis using differential gene expression analysis to rule out the cell’s having blood endothelial, smooth muscle, macrophage, or pericyte identity. This exciting combined evidence opens up an entirely new avenue of research into brain lymphatics in zebrafish.
Arachnocyte, Kumocyte, BLEC
Finally, as we debated what to name these cells, the other labs disclosed the names with which they were identifying the cells in their papers, with Galanternik et. al. calling them FGPs and Bower et al. settling on perivascular Lymphatic Endothelial Cells (pLECs). We ultimately decided on Brain Lymphatic Endothelial Cells (BLECs) to be precisely descriptive of their properties; however, internally we all enjoyed the name we had additionally selected which was Kumocyte. Kumo means spider in Japanese, describing their spidery appearance, they had previously been colloquialised as arachnocytes for the same reason. I think they will always remain Kumocytes to us somehow, as it speaks to their unique morphology and “personality” and we have taken to fondly calling them that within the Rihel lab.
We are grateful for this opportunity to share our experience as we are so often confined by formality, word limits, and jargon that we miss the opportunity to tell “the story”. The story which discusses the more personal aspects of the craft of discovery, especially the journey behind the scenes that keeps all of us all coming back again and again despite the tough days, which are many. After all, in biology and science, we are all just witnesses to nature, merely reporting what we see with awe and wonder. It’s always been there, but there’s that kid in all of us that gets our mind-blown every time we get to be the ones to see something with human eyes for the “first time.“ I also never want to forget on this journey, not just in science, but in life, that we truly are only as good as those we surround ourselves with. Each and every colleague on my floor has contributed, and given their time and expertise without hesitation. They say it takes a village to raise a child…..and so it also goes with raising a PhD student.
We can’t wait to see what this field of brain lymphatics and clearance holds. Certainly, dogma that the brain is without dedicated lymph structures of its own is being rewritten, even in fishes. As we start at the drawing board and re-examine all of our old assumptions, what else will we discover around the corner?
Shannon Shibata-Germanos and Max van Lessen
Professor Stefan Schulte-Merker presenting the finished paper on the enormous stage at the 10th European ZebraFish Meeting. (Photo: Renato Martinho)
Rihel lab out for pizza! Our colleagues are what make us what we are. They each contribute to the puzzle, as each has an expertise. Science is a team sport! (Photo: Sumi Lim)
Schulte-Merker lab after a 5k run! These guys don’t mess around! Our labs become larger and more capable as we partner and open ourselves to cross-pollinating science.
Cluster Hire in Quantitative and Computational Developmental Biology, North Carolina State University
As part of the Chancellor’s Faculty Excellence Program, NC State University seeks three outstanding faculty at any rank to expand the new interdisciplinary faculty cluster on Quantitative and Computational Developmental Biology. Together with NC State’s existing strength in quantitative/computational sciences and engineering, this cluster will propel NC State to the forefront of efforts to define new principles in developing biological systems.
Successful candidates will engage in transformative research which integrates expertise in biological, biophysical, computational, and/or mathematical methods to investigate dynamic events in human, animal or plant development on any level, from the nanoscale to the whole organism. The cluster seeks individuals with expertise in using state-of-the-art quantitative methodologies and/or computational modeling to address fundamental aspects of Developmental Biology such as:
the mechanisms that regulate individual cell behavior, proliferation, differentiation, potency, and reprogramming,
how material properties and dynamic physical / mechanical forces influence collective embryonic cell behavior and tissue shape,
the formation, maintenance and appropriate scaling of the geometric configurations of cells and tissues that comprise organs and organisms, and
the mechanisms that underlie the evolution of new biological forms and facilitate morphological plasticity in response to stress or environmental change.
Although we are interested in individuals who can bridge experiment and theory and have the ability to translate research outcomes to address biomedical or agricultural challenges, applicants whose expertise is mainly in the computational realm are also strongly encouraged to apply. Demonstrated collaboration as a member of a multi-disciplinary team is essential.
Hiring may occur at the level of Assistant, Associate, or Full Professor. The home department is anticipated to be in one of the four participating colleges (Engineering, Sciences, Veterinary Medicine and Agriculture and Life Sciences) and will be determined based on credentials and research fit. Hires will be expected to provide key leadership in quantitative and/or computational biology across the university, teach in existing courses and develop specialized coursework in their area of expertise.
Minimum requirements include a PhD in a relevant field from an accredited institution. Interested candidates should submit: a CV, a 2-3 page research plan, a cover letter describing prior multi-disciplinary research efforts and how their research prospectus addresses the goals of the cluster (see https://facultyclusters.ncsu.edu/clusters/modeling-the-living-embryo/), and
contact information for 3 references. Materials for consideration will be accepted electronically via http://jobs.ncsu.edu/postings/88538. Review of applications will begin immediately and continue until the position is filled. Questions about the position may be directed to Dr. Nanette Nascone-Yoder (nmnascon@ncsu.edu).
NC State University provides a vibrant environment for research, teaching and mentoring across disciplines; ample opportunities will be available for collaborations with existing faculty and other newly hired colleagues in the cluster. Our location in the Research Triangle also facilitates interaction with faculty at Duke University and the University of North Carolina at Chapel Hill, as well as with industry and government agencies.
The Chancellor’s Faculty Excellence Program is bringing some of the best and brightest minds to join NC State University’s interdisciplinary efforts to solve some of the globe’s most significant problems. Guided by a strong strategic plan and an aggressive vision, the cluster hiring program is adding new faculty members in select fields to add more breadth and depth to NC State’s already strong efforts. The Chancellor’s Faculty Excellence Program marks a major initiative of the university’s strategic plan, “The Pathway to the Future” We invite you to explore more information about the Chancellor’s Faculty Excellence Program and this cluster at http://workthatmatters.ncsu.edu/.
NC State University is an equal opportunity and affirmative action employer.All qualified applicants will receive consideration for employment without regard to race, color, national origin, religion, sex, gender identity, age, sexual orientation, genetic information, status as an individual with a disability, or status as a protected veteran. Persons with disabilities requiring accommodations in the application process please call (919) 515-3148.
The Mokalled laboratory is seeking applicants for a Research Technician II position in the Department of Developmental Biology at Washington University School of Medicine. Our lab uses the zebrafish model system to study neural regeneration after spinal cord injury or disease. We welcome ambitious applicants with enthusiasm for neuroscience, regenerative biology, and zebrafish research. Training will be provided. This position offers opportunities for co-authorships on published manuscripts.
Duties:
Assists with research studies, experiments and assays including collection of data, preparation of solutions and set-up and maintenance of equipment. Main experimental duties include molecular biology, zebrafish husbandry and microinjection.
Performs data entry and maintains data files on research.
Complies with established safety procedures and maintains required documentation on laboratory and specimen conditions.
Ensures lab conditions and equipment are properly cleaned and maintained in accordance with established procedures.
Assists with general lab maintenance and cleaning.
Required qualifications:
Bachelor’s degree and up to 1 year of experience in lab setting or equivalent combination of education and experience equaling 4 years required. Please send a cover letter, CV, and list of 3 or more references to mmokalled@wustl.edu.
Each of our cells has the same genetic information and thus the same potential to become a part of a heart, brain, or a finger. Somehow though, during development our cells manage to figure out exactly which type of cell they should be and which body parts they should help compose. The key to making this work is precise control over gene expression, such that as a single cell divides to make trillions, the correct genes are turned on and off at precisely the right times and places. When gene regulation fails, developmental disorders and diseases like cancer can occur.
Today, transcriptional regulation in animals is understood to involve a complex integration of cis-regulatory elements (enhancers), cell signaling pathways, nucleosome occupancy, and higher order chromatin architecture that all work in concert to direct gene expression. In the McKay Lab we are interested in how spatial and temporal regulatory information is integrated during development, and how that integration produces the distinct cell types and body parts of animals.
Central to the process of differential gene regulation are relatively small genomic regions called enhancers. Enhancers, as their name implies, have long been known to increase gene expression, often over long distances. In extreme cases an enhancer can act on a gene that is over 1 million bases away, such as in the case of the enhancer that regulates the Sonic hedgehog gene1.
Enhancers mediate gene activation by serving as landing pads for proteins (transcription factors) that recruit the machinery required for gene transcription. Enhancers are remarkable not only for their ability to regulate gene expression across long genomic distances, but also because they exhibit highly flexible sequence characteristics, they work independently of their orientation, and they can be transplanted to different parts of the genome and still retain their ability to activate nearby genes2. The role and importance of enhancers has become increasingly apparent, especially as the links between human disease and mutations within enhancers have become much clearer3-5.
One of the major current question in the field is how networks of enhancers direct the differential gene expression programs during development.
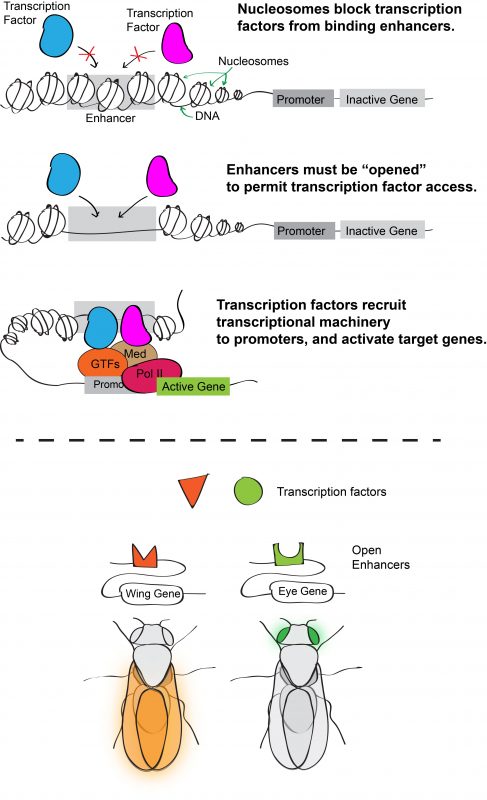
Inside eukaryotic cells, DNA is packaged into chromatin, the basic unit of which is the nucleosome. Wrapping DNA around nucleosomes succeeds in packaging two meters of DNA into a ten-micron nucleus. It also serves as a potential means of controlling access to the DNA sequence because nucleosomes typically block transcription factors from binding their target enhancers. For a transcription factor to bind DNA, its target site must be sufficiently depleted of nucleosomes or “opened.” Consequently, one hypothesis for how transcription factor binding is controlled (and hence for determining which enhancers are active) is through regulation of chromatin accessibility. According to this model, an enhancer with cell-type specific activity would be open in cells in which it is active, but closed in cells in which it is inactive (Figure 1).
Figure 1. (Top) DNA wraps around an octamer of histone proteins forming the nucleosome complex. This allows for tight packing of DNA in the nucleus, and blocks transcription factors from binding target enhancer sites. Enhancers must be “opened” to allow for transcription factor binding and subsequent gene activation. (Bottom) One model of how enhancers direct differential gene expression is by having tissue-specific accessibility, such that broadly expressed transcription factors only activate genes in cells where target enhancers are open.
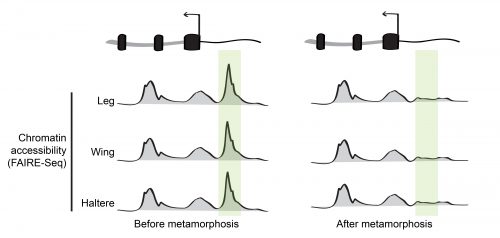
During his postdoctoral work, our PI Dan McKay tested this idea by profiling the open chromatin regions during appendage development in the fruit fly Drosophila melanogaster6. Using a high-throughput sequencing method that identifies genomic regions of low nucleosome occupancy, called FAIRE-Seq, Dan found that, despite having different morphologies, transcriptional programs, and transcription factors that specify the distinct identity of each appendage (so called master transcription factors), the patterns of open chromatin were surprisingly similar between the cells of wings, legs, and halteres. Even more surprising was the finding that although these patterns were dynamic over time, they remained highly similar between the appendages. In other words, the same regions of the genome appeared to be opening and closing at similar times, even though the cells were in completely different parts of the animal. This indicated that the temporal control of chromatin accessibility might be a more significant driver of differential gene expression during appendage development than spatial control (Figure 2).
Figure 2. In many cases, chromatin accessibility remains highly similar between the leg, wing, and haltere during development. Many of the temporal changes that occur are coordinated between these spatially separated appendages.
Dan’s 2013 paper raised a big question: If the transcription factors that specify the distinct identity of each appendage weren’t primarily responsible for directing chromatin accessibility, what exactly was? And on top of that, how were these changes in open chromatin being coordinated across spatially-separated tissues?
One of the phenomenal things about insects is that the timing of their development is precisely controlled by a steroid hormone called ecdysone. In Drosophila, levels of ecdysone increase at stereotypical developmental timepoints, particularly during major transitions like molting7. The work of Michael Ashburner in the 1970s revealed that these ecdysone pulses were responsible for activating a large set of genes, many of which were eventually found to be DNA-binding transcription factors8. Because ecdysone acts systemically, and it has a known role in controlling gene expression, we wondered if ecdysone, and in turn ecdysone-induced proteins, contributed to the coordinated changes in chromatin accessibility that Dan had observed.
Focusing our attention on wing development, our first step was to get a better sense of how chromatin accessibility was changing in the wing9. Our original FAIRE data had only looked at chromatin accessibility at two time points: a stage right before metamorphosis and a much later stage at the end of metamorphosis. Consequently, it was essential to get FAIRE-Seq data in the pupal wing at finer timepoints. With collaboration from the Buttitta Lab at the University of Michigan, we obtained FAIRE-Seq data that confirmed Dan’s observations; chromatin accessibility was changing significantly, with thousands of regions opening and closing over a relatively brief two-day period. Based on these results we knew that the pupal wing, which undergoes striking morphological changes as it develops, was going to be an excellent model system for us to examine how chromatin access is regulated.
Since ecdysone is a steroid hormone and is essential for even the earliest stages of fly development, we couldn’t remove it completely. Instead, we focused our attention on one of the transcription factors directly induced by ecdysone, Eip93F (E93). To ask whether E93 was involved in changing chromatin accessibility, we repeated our FAIRE-seq experiments in an E93 mutant fly. We found that many regions (~50%) that originally showed dynamic changes in accessibility, failed to change their state in the mutant. So, it appeared that we were on the right track and that E93, and by proxy ecdysone signaling, was required for directing many genome-wide changes in chromatin accessibility. Importantly, our data showed that E93 was required for both opening and closing of chromatin.
Although these data showed us that E93 was required for changes in chromatin accessibility, it didn’t tell us whether it was directly causing these changes. To help answer this question, we wanted to determine where E93 was physically bound in the genome. These days assaying protein-DNA binding by ChIP-Seq is a standard practice, but the method requires a significant amount of input to get good results. When working with cell culture this isn’t much of a problem, but it presents a serious challenge when working with tissues in tiny animals like Drosophila, because we needed over a thousand wings to have sufficient input. Over the course of a week, the lab worked together to dissect all the wings necessary for the experiment, and in the end the effort paid off. We found that not only was E93 required for many of the changes in chromatin accessibility, it actually bound to about half of these dynamic regions, arguing that in many cases E93 had a direct role in coordinating both opening and closing chromatin.
Next, we had to tackle whether the temporally dynamic chromatin regions we observed in our FAIRE data actually corresponded to functional enhancers. The best method for testing enhancer functionality is still to clone them upstream of a fluorescent reporter gene. This can work great, but makes it much harder to test many candidates with a high degree of throughput, as it can take months to establish and test the transgenic flies. This was right around the time I joined the lab as a first-year graduate student, and I got the chance to work on characterizing the function of several of the putative enhancers we had cloned. We were thrilled to find that the E93 dependent dynamic chromatin regions we had cloned successfully drove GFP expression, and that the timing of their activity correlated with the timing of the accessibility of the native enhancer.
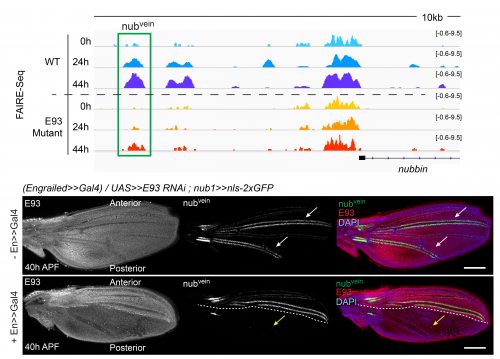
The crux of this project was to finally test if the failures that we saw in enhancers opening or closing in E93 mutants correlated with a functional defect in our reporters. We found that in the absence of E93 there were dramatic changes in enhancer activity, and that these changes followed what we would expect based on the changes we saw in chromatin accessibility. For example, the nubvein enhancer, which normally opens during pupal wing development, produces striking patterns of fluorescence along the wing veins when it’s cloned upstream of a GFP gene. However, in an E93 mutant fly, this region fails to open to the same degree. When we imaged the nubvein enhancer reporter in the presence of E93 RNAi we saw complete loss of the normal vein pattern of expression (Figure 3).
Figure 3. The nubvein enhancer becomes more accessible over the course of wing development (top panels), driving expression in cells that flank two of the major wing veins (white arrows). But in an E93 mutant, the enhancer fails to open to the same degree, and when E93 RNAi is produced in the posterior half of the wing the normal nubvein expression pattern is lost (yellow arrows). Genome tracks are shown in the top panel, with immunofluorescence images in the lower panel.
What we found most exciting about this project is that it demonstrated that an extrinsic signal, ecdysone, which pulses at specific times in development, alters the accessibility of enhancers to binding by transcription factors. In other words, for a genome that contains far greater regulatory capacity than is used at a given point in time, hormone signaling can help to determine which subsets of the genome are accessible for use at given time in development. Thus, ecdysone-regulated chromatin accessibility provides a temporal-specific input, which is combined with spatial-specific input in the form of tissue-specific transcription factors. Amazingly, both forms of input are integrated by enhancers.
This has left us with a whole new set of questions to go after. What are the mechanisms that drive ecdysone-dependent changes in chromatin accessibility? Does E93 act alone? How is it that E93 is required for both opening and closing chromatin? How does chromatin accessibility fit into other aspects of gene regulation such as long-distance enhancer-promoter interactions? There’s a lot more to figure out, and these questions are actively pushing us to expand our research focus. Fortunately for me, that means there’s a lot of room to explore and opportunity to make some meaningful contributions to the field.
Deciphering the Mechanisms of Developmental Disorders (DMDD) is a large-scale imaging and phenotyping programme for genetically modified mouse embryos. For embryos at E14.5, the key imaging technique is High Resolution Episcopic Microscopy (HREM), and the resulting images are used to comprehensively phenotype the embryos using a systematic approach.
With a combination of lectures, demonstrations and hands-on sessions, this three-day workshop (20-22 October, The Medical University of Vienna) will introduce HREM technology and discuss the value of the resulting images when used to score morphological phenotypes. The HREM procedure will be described, while sample preparation and data generation will be demonstrated.
As an introduction to phenotyping, the workshop will cover the normal anatomy of E14.5 mouse embryos and the morphology, topology and tissue architecture of their organs as presented in HREM data. A special focus will be given to developmental peculiarities and norm variations in anatomy. A protocol for scoring abnormalities will be demonstrated, after which hands-on sessions will allow participants to practice scoring both wild-type and mutant embryos whilst receiving feedback.
I recently saw drawings by Maria Sybilla Merian at Kupferstichkabinett Berlin and the University Library Dresden. Merian, who lived from 1647 to 1717, is renowned for her exceptional illustrations of biological specimens and gained recognition as a scientist for her nature observations, for example, of insect metamorphosis.
Merian evidently was genius in choosing frame and magnification in her drawings, but her pictures lack indications of scale*, which are essential in today’s science images. Scales give the reader the key for aligning the image content with reality. To my knowledge, neither Merian nor her predecessors from Antiquity, Byzantium, or Renaissance included scales in their medical and natural science images*. Even in the beginning of the 20th century, images were often considered a waste of space and scales unnecessary as scientists were familiar with each other’s apparatuses and objects. Today however we study invisible processes and structures that are unfamiliar to most of our colleagues and therefore have to include scales in our images.
Comment from Benjamin Moore in nature (1910) when reviewing a biochemistry handbook.
We often include in images a familiar object of a standard size for scale: a penny placed on a rock, a person standing beside a large animal or in a landscape, a measuring tape next to a fossil (or an Earth worm!).
Bar = 1cm (Earth worm lovingly raised by Jeff Woodruff).
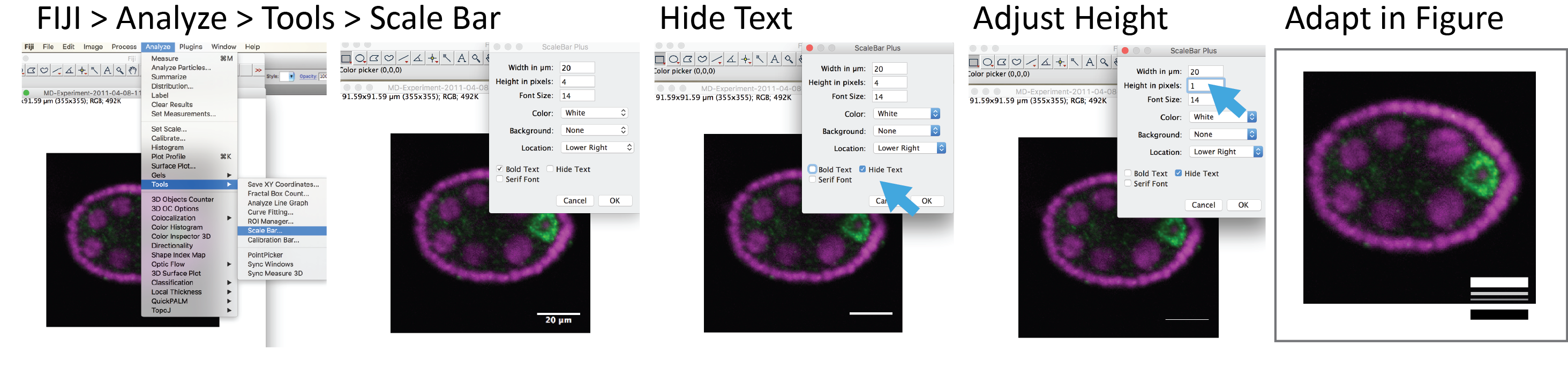
Using familiar objects for scale isn’t possible for tiny things. We don’t have a clear mental image of the size of a salt grain or sesames seed to reliably use them to scale for instance cells**. We therefore include scale bars in microscopy images. With ImageJ/FIJI files from any microscope system can be read in along with their scaling information (shout-out to Curtis and Melissa and the Bio-Formats project!). By using Analyze > Tools > Scale Bar we can add the scale bar with a user-defined length, width, color, position, and label. Now the audience can calculate the actual size of objects and relate image with reality.
Four tips for superb scale bars
Length: Be kind to your audience and use simple units, such as 100um, 50um, 10 or 2um.
Color: Scale bars should have a high contrast with the background. Avoid red, green, or blue bars, as these colors might be considered part of the image.
Position: Lower left corner is a safe place. The upper space should be kept for important information like species, cell type, or gene name.
Add scale bar last: In the process of writing your manuscript you may re-think the figure size. Also images are re-sized for posters and slides. It is therefore easierst to add only a very fine scale bar with FIJI and then re-draw it in Adobe Illustrator (or PowerPoint, as I I know that about half of you out there use PowerPoint for making figures and posters!).
And finally, do not miss this article by Monica Zoppe with an interesting idea on how to communicate subcellular sclales better!
* I’d be delighted to stand corrected, and if you find old scientific images with scale bars, or interesting scales, send them my way for my collection!

 (3 votes)
(3 votes) (No Ratings Yet)
(No Ratings Yet)





 (8 votes)
(8 votes)