We are seeking outstanding candidates to lead a project studying Notch, ephrinB2, and TGF-b signaling pathways in arterial venous programming/reprogramming during development and disease processes. We take a conditional mouse genetic approach to manipulating gene expression in endothelial cell-specific and temporally controlled fashion. We also use cutting edge in vivo real time imaging technology, including an in-lab constructed two-photon microscope, which provides exceptional access to gene function in vivo at the cellular resolution along with blood flow measurement overtime in live animals. This basic approach is complemented by preclinical studies with our elegant mouse models of diseases, offering outstanding opportunities for translational research. The laboratory is well equipped with state-of-the-art capabilities at the molecular, cellular, and organismic levels. In addition to funding from the PI, we also have an excellent track record in sponsoring postdoc fellowships. We are interested in a well-trained, highly productive recent Ph.D. to continue our innovative breakthroughs in a rewarding training program. This postdoctoral research is an excellent platform for a highly productive Ph.D. with a strong motivation to become a future group leader. Experience with mouse techniques is a plus. UCSF offers outstanding postdoctoral career development opportunities. Please submit your CV, research interests, and the names of three references by email with a subject title “postdoc application” to:
Rong Wang, Ph. D.
Professor
UCSF
rong.wang@ucsf.edu
Multiple postdoctoral positions are available in the field of quantitative stem cell biology at the Warmflash lab at Rice University. Our lab uses human embryonic stem cells to model early embryonic development with a particular focus on understanding the mechanisms of spatial patterning and morphogen signaling dynamics. Our work combines quantitative experimentation with data analysis and mathematical modeling. For more details, see our lab webpage here and examples of our work can be found in our recent publications: Warmflash et al. Nature Methods 2014, Sorre et al Dev Cell 2014, Nemashkalo et al. Development 2017.
Positions are available for:
Theoretically trained scientists interested in working closely with experimentalists. Experience with either mathematical modeling of biological systems or analysis of biological data is preferred but not required.
Experimental biologists interested in quantitative approaches and working closely with theorists. Experience with cell culture, microscopy, or molecular biology is preferred but not required.
Interested candidates should email a CV and a brief statement of past research accomplishments and future research interests to aryeh.warmflash@rice.edu.
A Research Technician position is available in the research group of Mina Gouti at the Max Delbrück Center for Molecular Medicine in Berlin. The group is using human and mouse pluripotent stem cells to study the development and disease of neuromuscular system. Further information about research in the lab can be found at: https://www.goutilab.com
The successful candidate will have a B.Sc. or M.Sc. degree in Biology or other related discipline, extensive experience in molecular biology and cell culture. Excellent communication, organization and prioritization skills are required, as well as flexibility to the work schedule as stem cell maintenance involves weekend attention (on a rotating basis). Proficiency in writing and speaking English is essential. Knowledge of German language will be considered as an additional advantage.
Key responsibilities will include:
Performing research projects involving molecular biology techniques, pluripotent stem cell culture and differentiation as well as mouse and chick embryological techniques.
Maintenance of mouse and human pluripotent stem cell lines for the lab stock.
Generation of new pluripotent stem cell lines using the Crispr/Cas9 system.
Oversee the maintenance of mouse colony, monitoring and submission of animal protocols.
Provide technical support to other lab members when required.
Establish, maintain and improve research protocols and maintain lab records.
Oversee the effective running of the lab by monitoring stock levels, ordering consumables and reagents, maintaining equipment and updating lab databases.
Attending safety courses in order to support and maintain good laboratory practice and safety procedures.
Minimum Experience:
Two years of lab experience working with molecular biology techniques. Previous experience with maintenance of a mouse colony and/or pluripotent stem cell culture will be an additional advantage.
The salary will be according to the TV-L9a scale and the contract will be initially for two years with the possibility of renewal. Applicants should send their CVs along with names and emails of at least two referees to Dr Mina Gouti (mina.gouti@mdc-berlin.de).
Blog post written by Isabelle Vea – 2017 Embryology Student
All 24 of the 2017 Embryology students came to Woods Hole to learn from the best scientists in the developmental biology field. We were immersed in a unique setting to interact with established and promising investigators. In general, each invited lecturer came and spent from a few days to a couple of weeks at the Marine Biological Laboratories and was dedicated to interacting with us. Through our interactions with the faculty we quickly learned that it was more than just learning knowledge and techniques. For instance, the course directors carefully planned lunches and dinners with one faculty and two of the students allowing us to discuss informally about all sorts of topics.
In this post, I would like to share some of the interactions I had with the course faculty that are not related to embryos, yet were extremely meaningful.
Science and art
I always thought that science required creativity and so for me, art/crafts and science is a natural combination. But is it possible to combine both in our daily lives? Isn’t science already a lot? One of my most memorable discussions was with Bob Goldstein from UNC Chapel Hill, who managed to integrate his artistic views into the academic world.
Bob creates posters for scientific seminars in his department using screen printing. It was refreshing to be able to discuss screen printing techniques, inks etc. with Bob. We also discussed using arts and craft as a medium for talking about science at outreach events.
I do think it is possible to combine research with other hobbies or non-scientific activities. The bonus (and actually what Bob taught me through meeting him) is to be able to find other researchers with the same artistic interests.
If you are an artistic scientist, give a shout out in the comments! And if you are interested in discovering scientists with an artistic mind, check out #sciart on social media platforms.
Leadership
As graduate students and postdocs, our primary concern is to find the next research position. As a postdoc myself, I am still struggling to figure out what type of researcher I want to become in the long term. I have always wanted to know how one scientist decides on becoming a leader in his department or institution. Unfortunately, I have never had the opportunity to ask such questions in my home institutions. Here at the course, asking these questions seemed natural and many discussions took place informally in a pub or the hallway.
I was particularly interested to hear from Claudio Stern about his leadership experiences. Claudio Stern from UCL is involved in service to the scientific community, he is part of the scientific council at the Institut Pasteur in Paris and the previous president of the International Society of Developmental Biology. We discussed how you decide to become a leader in academia, and what opportunities may lie beyond your own lab. I learned that at some point in our science careers, we may ask ourselves whether we would like to help improve our colleagues’ work environment and to do so, we need to be able inspire them.
Enthusiasm in research
Despite my undivided love for invertebrates and the excellence of every module, my favorite week was the zebrafish/frog one. Into the second week of the course, I had been overwhelmed by C. elegans powerful tools to examine cellular mechanisms and going into vertebrate species was quite intimidating. I initially thought that my lack of knowledge in vertebrate anatomy would be detrimental to learn the techniques suggested during the module but something special happened.
Left to Right – Joaquin Navajas Acedo (graduate student in Piotrowski lab and course TA), Elke Ober (University of Copenhagen), Isabelle Vea, Sara Caviglia (postdoc in Ober lab and course TA), Tatjana Piotrowski (Stowers Institute)
The zebrafish lecturers (Elke Ober from University of Copenhagen and Tatjana Piotrowski from Stowers Institute) were very present during our lab time. They were not only physically here but kept checking on our experimental progress throughout the week and transferred a lot of their enthusiasm to us.
I had decided to perform a simple experiment that would back up my very risky one. As expected my risky experiment failed but I had managed to obtain time lapse images of the simple one. The night before show and tell, I asked one of the lecturers if there were still time to set up a new experiment: transferring cells of an embryo that I would have injected with fluorescent dye, into another embryo and see where the cells develop. I had never transferred any cells from one organism to another and the task seemed impossible to do with less than 24 hours left. But Elke just looked at me with excited eyes: “Yes! You should try! I will help you.” I could not not try.
Next thing I knew, I was injecting embryos with fluorescent dyes and transferring cells at 1 am in the very last hours of the module and both Elke and Tatjana were there as moral supports! The next day we checked the embryos, some of them survived and it worked! It was the most rewarding result of the course, not only because it worked but because I was inspired by their enthusiasm.
For sure, on a daily basis, experiments are not always successful (a big proportion of mine actually failed during the course), but just having one experiment work and a supportive and passionate community makes your day.
Overall, each faculty member came to Woods Hole with something to share with us. They taught us what they know about embryos but to me, they also conveyed their passion, shared their life experience and how this can be relatable to us, as budding independent investigators. And for this, the course was invaluable.
Isabelle Vea
Isabelle Vea is a Marie Skłodowska-Curie Fellow at the University of Edinburgh interested in the evolution of scale insects.
A Wellcome Trust/Royal Society funded Research Assistant position is available in Dr Kyra Campbell’s research group. This is a fantastic opportunity to join the Campbell group, who are focused on identifying the molecular mechanisms underlying epithelial cell plasticity during development and disease. We study how this fundamental property is orchestrated during morphogenesis of the Drosophila midgut, and also in exciting Drosophila cancer models that we have recently generated.
We are looking for a motivated and enthusiastic candidate who will play a central role in the lab. You will be involved in all aspects of an ambitious research programme combining molecular biology and Drosophila genetics with high-resolution microscopy on our own dedicated multiphoton confocal, working closely with Dr. Campbell. In addition to carrying out research, you will also be expected to support the research of the laboratory, for instance by providing technical assistance, maintaining and keeping records of reagents and Drosophila stocks and as well as general laboratory organization.
Applicants should have a good honours degree or equivalent in a discipline relevant to studying animal development and cellular behaviours, and must have previous experience in the use of model genetic organisms, and molecular biology techniques. Previous experience in generating CRISPR mutants or Drosophila transgenics will be an advantage. This is a unique opportunity for you to carry out cutting-edge microscopy and develop your skills in an exciting multidisciplinary environment.
Candidates must be highly enthusiastic and committed to the project in addition to displaying outstanding motivation and commitment to research in a fast-moving and competitive field. Excellent organisational and interpersonal skills along with the ability to work effectively with other members of the research team are also essential.
To interview Jim Smith I took a train to London and visited the Francis Crick Institute for the first time. The building had opened in 2016 and, by the time I visited, most if not all of the labs had settled in. Architecturally it was quite stunning, especially looking down from one of the higher floors and particularly so on a bright spring day. We sat down in one of the many open areas and chatted for an hour about Jim’s life and work both inside the lab and at the helm of large research institutions and funding agencies. I left thinking how positive it was for the field to have a developmental biologist with such energy and enthusiasm in such high places.
Jim Smith is Director of Science at the Wellcome Trust and a group leader at the Francis Crick Institute, where he was formerly Director of Research. A Fellow of both the Royal Society and the Academy of Medical Sciences, he was knighted for his services to medical research and science education in 2016. His lab works on mesoderm induction in the early vertebrate embryo. We met Jim in the Crick to hear about his life in science, his visions for the Crick and the Wellcome Trust, and his advice for early career scientists.
Image courtesy of Kirsty Watt
Let’s start with your first paper, which describes an investigation into the patterning of the chick limb from your PhD with Lewis Wolpert. How did this work come about?
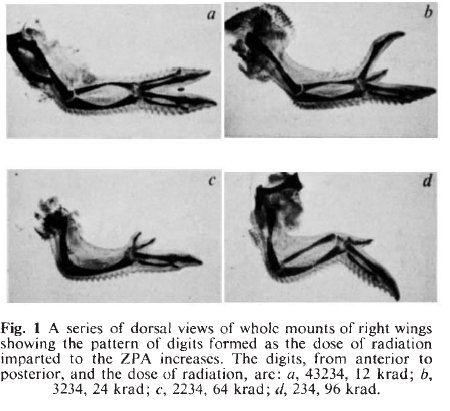
When I went to university I was interested in maths, physics and chemistry, and at Cambridge you could do those three subjects without having to choose between them. But I had to do one more subject, and my director of studies convinced me to try biology, which I’d never done before because I’d done my O-levels a year early and there wasn’t time in the curriculum. So I gave it a go, and loved it. I was sent down the developmental biology pathway through lectures by John Gurdon and Peter Lawrence, and was introduced to Lewis Wolpert’s work by Peter. I joined Lewis’s lab to do a PhD at a very exciting time – they had just grafted mouse polarising regions into chicken limb buds and seen that you got duplicated limbs, which was a really dramatic illustration that there are universal molecular signals in limb development.
Now, that first paper – at the time I shared an office and a lab with Cheryll Tickle, and Lewis set me to work on the zone of polarising activity (ZPA), this special instructive region of the posterior part of the limb bud. Following on from wonderful work by Cheryll and Dennis Summerbell, the rationale for my experiments was to ask whether you needed cell division in order for the ZPA to influence development. So I irradiated these limb buds, first with X- and then γ-rays, and found that even without dividing the cells were still able to signal, but also that the more radiation you gave them, the weaker the signal was in terms of which extra digits it produced. At the time, I thought it was rather obvious and not was worth publishing or mentioning, but when Lewis heard about it he was much more interested and said we should write it up for Nature right away. Six weeks later, we had a thin envelope back saying it had been accepted – it was the easiest paper of my life, and it’s been downhill ever since.
And did you keep in touch with the limb field in your later career?
I did and I do, though of course there is so much literature to keep up with these days. I remember being at a meeting in Switzerland and bumping into Cliff Tabin, who took me somewhere – maybe his hotel room – and showed me the first in situ hybridisation data for Sonic Hedgehog. My mind was completely blown! In Lewis’s lab we had mapped out where the ZPA was by grafts and so on, we knew where it was at different stages, and these in situ data just overlapped perfectly. It was probably the most exciting thing I’ve ever seen – it was extraordinary.
After your PhD you went to Harvard for a short postdoc with Chuck Stiles – was this your first brush with growth factors?
Although I had always wanted to go to America, ending up with Chuck was a bit of an accident, but a happy one because I wanted to learn cell biology and biochemistry. Lewis’s lab was great to teach you to think, but (with all due respect) much of what you did was graft bits of tissue around, and I knew that to work out what the polarising signal was I’d need to know a little bit about cell biology and biochemistry. So in Chuck’s lab we did some stuff on the regulation of the cell cycle, and I learned cell culture and cell fusion. We discovered – in another paper that got published surprisingly easily – that growth factors remain active when stuck to substrates. But perhaps the most important thing was the recognition that growth factors might be important not only to make cells grow, but also for embryos and patterning as well. I published a paper in a supplement of the Journal of Embryology and Experimental Morphology (now Development) in 1981 called ‘Growth factors and pattern formation’, which was quite prescient I guess, though it remains relatively uncited. All this work got me thinking about how to assay for the activities of these factors in embryos.
I enjoyed living in the States – I wish I had been able to stay there longer. I’d never had more disposable income in my life before, or indeed since – I remember going into record shops and just being able to buy whatever I wanted! Lab-wise, the differences were extraordinary. Lewis’s lab was on the top floor of the Windeyer building of the Middlesex Hospital Medical School, and it was really just an extended hut – if the weather was hot, you didn’t need to put the chicken embryos in an incubator because it went up to 38°C anyway. But this did illustrate to me that you can do great science, fun science, in an environment that might look unpromising. And Lewis was a fantastic supervisor – he kept out of the way when he had to, gave advice when it was necessary, and supported me and continues to support me today.
Back in England for a postdoc with Jonathan Slack, you started your career-long affiliation with Xenopus. What do you think of the state of Xenopus as a model, and its future?
Xenopus is a very powerful model organism that you can still do great work with, and I claim – and will fight anyone who disagrees with me – that we have learned more about the generalisable aspects of early vertebrate development from Xenopus than from any other species. But I do think that people worry too much about model organisms, and Xenopus people are no exception in being defensive over their model. In an ideal world, you would work on a model that best allowed you to answer the question you are interested in. The trouble is that it takes a certain amount of investment and infrastructure to work on any model, which raises the activation energy for shifting to another. This means it’s very helpful to work in an environment that has as many organisms around as possible, which was what we had at Mill Hill in the nineties. If we had a problem that would be better approached in another animal, we’d just go and collaborate. The Crick will I hope foster the same sorts of collaborations, but also beyond developmental biology.
First with Jonathan Slack and then in your own lab at Mill Hill, you helped to identify the molecules responsible for induction of the mesoderm, and later the transcriptional networks these molecules regulated. What are the key open questions in how mesoderm induction works today?
The way I like to think about it is reflected in the title of a talk I gave recently – ‘The first ten hours in the life of a frog’. Mesoderm induction happens during those first ten hours or so, and it’s becoming clear that everything in this time is coordinated and interdependent, so if you want to understand mesoderm induction you cannot look at it in isolation. You have to look at it in terms of fertilisation, chromatin structure and dynamics, the early activation of transcription and transcription factor cascades, and the cell cycle, which we are working on quite a lot at the moment and which is proving to be very interesting. And intercellular signalling – we still don’t quite know how signals travel between cells, the details of how the signal transduction pathways activate the genes, or how the different pathways interact with each other. We don’t really understand the cell movements, why it is that particular cells move earlier than others, and why they move where they move. There’s still so much to find out, and it will certainly see me out as a problem.
If you want to understand mesoderm induction you cannot look at it in isolation
And where do you think developmental biology as a field is going?
I think we’re getting down to a deeper understanding of the spatial and temporal aspects of development. What I’d like to do is to develop simple real-time in situ hybridisation, to watch genes turning on and off in real time in a living embryo – not snapshots of fixed embryos. If you just watch things happen, you’ll get a really good idea of the dynamics of the processes. I think what it comes down to is technology – a lot of what we do will be driven by new advances. As new technologies come along, we’ll be able to ask new questions, some of which we can’t conceive of at the moment. Technology is highly under-rated: as I think Mike Levine said in a previous Development interview, the low-hanging fruit in developmental biology has been hoovered up mercilessly by the old farts – my generation – and we were very lucky. The future will lie in the ability to ask new questions with new technologies.
As well as a productive research career, you have held top administrative jobs throughout your career. As head of the National Institute for Medical Research (NIMR), you were involved in the development of The Francis Crick Institute, where you were Director of Research and where your lab is now based. What was the extent of your involvement, and, a few months in, is it too early to say whether the initial aims are being met?
I think it might be helpful to go back and ask why I do these leadership jobs in the first place. One of the things about early scientific success, which I was fortunate enough to have, is that people think you will be good at other things besides being good at science. So people ask you to do stuff, and if you’re not strong enough, as I wasn’t, you just say yes, and end up wrapped up into these leadership positions. Not that I’m complaining, you understand!
So I went to the Gurdon Institute to be Director from 2000 to 2008, and that was a very valuable experience. It was a small-ish place with terrific, collegial people working there, who made it very easy to do my job. And then I moved back to the NIMR as director, having worked there previously for 16 years from 1984. Coming back, it was a torrid time for the institute, because its members did not know what was going to happen to it and were very anxious as a result. We were aware that there would be this thing into which NIMR would move, and as the idea took shape, it was my job to shepherd NIMR into this new world. It was an interesting and fun task to work together to get the momentum and enthusiasm going for the move. By this time, Paul Nurse (then at The Rockefeller University), Richard Treisman (then Director of the CRUK London Research Institute) and I were working together quite closely on this, and we made sure, I hope, that everyone in the two founding institutes played as much of a part as they could in the design of the building and the way it would work. In designing an institute, the building itself is important, but just as important, or more so, are the people you bring in and the mindset that they bring in with them.
Richard and I spent a lot of time on building design, with a great deal of help from Steve Gamblin and John Diffley, and with the architects of course, and together designed the layout you can see today. The building’s lines of sight, its break-out areas, its central staircase, all reflect our aim for a design that would encourage people who work on different things to meet, interact, and talk with each other. And is it working? It’s been about six months, and the answer I think is yes – people seem to be happy and to like the building. I hope the mix of people we’ve brought together will mean we’ll have more collaborations than we had at the two founding institutes.
Well, of course I was sorry to leave – I had known the place for a third of a century, and I’d done my best work there, for sure. It was a quirky building in a quirky place, miles from anywhere – but it worked, and people loved it. It was a real wrench for many people to leave, but as this all coincided with its centenary, we had some really fantastic send-off parties! One hopes that the spirit and the collaborations will happen here as they happened there.
Your recently took up your job as Director of Science at the Wellcome Trust. What do you hope to achieve there?
Well, it’s quite a long list. First, I want to make sure that we fund the best science, and keep science at the centre of everything Wellcome does. We need to make sure the people we fund are supported properly, regardless of where in the country they work, and irrespective of their gender, social background, sexual preference, disability or religion. We have a drive at Wellcome focussing on inclusion and diversity, led by Lauren Couch, and that’s really important to me. We know for instance that 50% of graduate students in biomedical sciences are women, for example, but once you get to my exalted heights it’s something like 16% – we are losing some of our best scientists for no other reason than that they are women, and we have to stop that.
Second, I want to make sure that science is an attractive career with an appropriate career structure. We need to make sure that people are trained – for example in data science and statistics – at the right stages of their careers so that they have the best chance to advance. It’s also important that molecular biologists are aware of clinical medicine, and that people understand how pharmaceutical companies work, and how things operate in government. This will be crucial – not least because we need more people with a scientific background moving in to areas like law or journalism or government.
Third, while of course I want to emphasise and support basic science, it’s also important that when a basic scientist does something that can be translated, it is translated, and Wellcome’s Innovations division led by Steve Caddick can help with this. Everything depends on basic science but you can’t just do basic science and ignore the translation. It behoves people like me particularly to make sure that translation happens.
Fourth, I think it’s important to make sure that the UK is a good place to do science. At Wellcome, Simon Chaplin leads the Culture and Society division, which helps ensure that science is recognised and celebrated as being the best way to understand the way the world works, and to integrate science into our everyday lives.
It’s a lot, what I’ve just described – basic science, the right facilities, equality, infrastructure, careers, translation, research culture – but I think all these aims tie together.
Everything depends on basic science but you can’t just do basic science and ignore the translation
How do you see the prospects for funding of basic versus applied research in this country?
I’ve been Deputy Chief Executive at the Medical Research Council, and am now Director of Science at the Wellcome Trust, and when I speak to people in high office there is a clear understanding, belief and acceptance that basic science is the only way to go. But, as I touched on earlier, there is also the expectation that the basic science will be translated. Scientists cannot simply work in their basic science bubble – it’s important, as well as interesting and fun, to be around clinicians, chemists, pharmaceutical companies. As an example, the Crick has a fantastic interaction with GlaxoSmithKline that is not only advancing our understanding of particular problems, but also advancing our understanding of the environment in which both kinds of science work. This breaks down the illusion that they are different kinds of science – we are all in the end trying to work out how life works and how to make the world better, and the more you put people in bubbles, the worse off they will be. I do feel quite strongly about this.
How will institutions like the Wellcome Trust and the Crick adapt once Britain has left the European Union?
Both organisations are in a privileged position because people listen to us; for instance, Paul Nurse has been very visible in the debate about the effect of Brexit on science. In the immediate term, we care about our staff: in the Crick, about 30% of total staff and 56% of postdocs are from non-UK EU countries – we need to establish their right to remain in the UK. The postdoc stage is a very mobile time in your career: all of a sudden, young researchers go all over the world, and very frequently return to their home countries. That experience is incredibly valuable both to the country that sends them out and the country that receives them. So we are working hard to ensure that any migration system after Brexit should recognise that the UK in general, like the Crick, will benefit hugely from international researchers. I think that we should use Brexit to introduce a more streamlined system to cover all highly skilled research staff, whether EU or non-EU, and in particular we need to help people early in their careers to come to the UK – this has been difficult because the immigration system uses salary as a measure of seniority and skills, but postdoc salaries aren’t always enough to qualify them. And then of course there is the funding – access to EU funding and the European Research Council framework has been very valuable, if occasionally bureaucratic – and conversations are currently under way as to how we’ll maintain access to this.
You have a long history with Development, first publishing with us in 1979, and taking the reins from Chris Wylie to be our Editor-in-Chief from 2003 to 2009. How did the journal change during your tenure, and where do you see the journal’s present and future?
I looked it up – I think I’ve published 68 papers in Development, so it’s probably my main journal. Chris made some huge changes to the journal, turning it upside down, and Development is still his journal really: he gave us the structure, the format, the look. When I came in I saw my main job as not screwing up what Chris had done! With Jane Alfred, who joined as Executive Editor during my tenure and with whom I worked very well, we did make a few changes. It was stem cell time, or as Doug Melton and I both like to call it, ‘applied developmental biology’, so we got Ken Chien, Ken Zaret and Austin Smith in as editors. We introduced the ‘Research reports’ section, and I was keen to make sure that the review process was quick and decisive. I also wanted to make sure the papers were interesting – for a while there were too many papers with a generic title such as ‘The role of gene X in organ Y in species Z’, and I didn’t like that sort of title; we tried to make them more question-driven. I think we also had the initial discussions for Development’s community blog, the Node, and I’m quite proud of that even though I didn’t take it forward personally.
As for the future – I am quite taken by preprints, and I think Development’s two-way integration with bioRxiv is terrific. I am also keen on the Wellcome Open Research journal, and the planned Gates Open Research, both using the F1000 publishing platform – they’re interesting models to keep an eye on. We’re at quite an interesting time where we’re circling around trying to decide what the best way forward is for publishing.
And this year you were knighted – congratulations! What does the recognition mean to you?
Well it was fantastic for the field – with Ottoline [Leyser], Mandy [Amanda Fisher] and me all being recognised in one go. But yes of course I was pleased, and proud I suppose – I wished my parents were still alive to see it. I guess it sounds kind of hokey but it makes you think about all the people that work and have worked with you, the postdocs and students without whom I couldn’t have achieved much. I’ve been tremendously lucky in having great people in my lab, many of whom have become firm friends – part of the fun of doing science is that you meet people and make friends. The knighthood’s a hoot – my kids love it, I love it, but you can still call me Jim!
Do you have any advice for young scientists today?
I’ll repeat what I said earlier: if you can become expert in a technology it is not going to do you any harm at all. Enjoy yourself, remember how lucky you are to be doing what you’re doing, be generous with reagents and data, and magnanimous about authorship where necessary. The truth will always out, and the minuscule risk of sharing stuff and being scooped is far outweighed by the opportunities that will come along and the friends you will make – as I’ve said, one of the best things about science is the friendships you make along the way. But most importantly, I’d say to take your career into your own hands. Far too many people are passive in their careers, and wait for stuff to happen to them, but if you just sit back and wait, it won’t happen. Other people won’t do it for you.
Is there anything that Development readers would be surprised to find out about you?
Well I was thinking about this – I don’t think so, actually. Thanks to Twitter, I’m probably an open book – so people can find out anything about me, whether it’s the music I like or my love for running, by following me @ProfJimSmith.
Collated below are responses from the field (or at least, those of them on Twitter!) and links to opinion pieces. If we missed any links, let us know and we’ll update.
We’d also love to hear your thoughts on the work – use the comment section below.
Ion channels are famous for their roles in neurons and muscles, but the spectrum of phenotypes seen in ion channel mutants indicate a diversity of roles in development; the underlying mechanisms, however, have remained opaque. This week we feature a paper published in the latest issue of Development that reveals a link between potassium channels and morphogen signalling during epithelial morphogenesis. Co-first-authors Giri Dahal and Sarala Pradhan, and their PI Emily Bates of the University of Colorado Denver, told us more.
Giri, Sarala and Emily
Emily, can you give us your scientific biography and the main questions your lab is trying to answer?
EB My long term goal as a high school student was to understand the mechanism behind migraine headaches and I thought that genetics would be the best tool to do that. The ACCESS program for women in science at the University of Utah placed me in Dr. Anthea Letsou’s lab and that was a great head start for my career. Dr. Letsou introduced the powerful tools available in Drosophila genetics and BMP/Dpp signalling. I stayed in her lab for four years working on defining the role of Punt, a type II Dpp receptor. I then went on to Harvard Medical School for my PhD in Dr. Anne Hart’s laboratory to focus on genetic neurological disease. I took classes about ion channels at Harvard, getting me closer to my goal. Then I joined Dr. Louis Ptacek’s lab at UCSF to study a genetic link to migraine. Dr. Ptacek’s lab focuses on episodic disorders including migraine and Anderson Tawil Syndome, caused by mutations in Kir2.1, a potassium channel. When I saw that in addition to periodic paralysis, mutations in Kir2.1 cause birth defects in people and that the phenotypes in model organisms look like the BMP signalling mutants, I became intrigued. That brought me to one of the central questions my lab is trying to answer: How do ion channels and electrical activity influence developmental signalling? We also have project that are trying to figure out how properties of the cytoskeleton influence neurodevelopment and neurodegeneration.
Giri and Sarala – how did you come to join Emily’s lab?
GD I came to the USA in the fall of 2008 to pursue graduate study at Brigham Young University. Emily was an Assistant Professor there. She had a positive outlook towards life and her views in science were inspiring. She wanted to pursue two projects, both were interesting but something about this project caught my attention. I was intrigued to find that we do not know why Kir channels cause dysmorphic features in Andersen-Tawil Syndrome even though scientists have been studying the channels for a long time. I liked both her personality and the project so decided to join her lab. I went with her to Colorado when she started her position at the University of Colorado.
SP Two ‘D’s attracted me, Drosophila and Denver, but certainly the research that was happening in the lab was key in my decision.
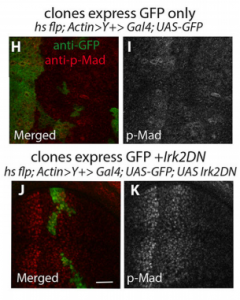
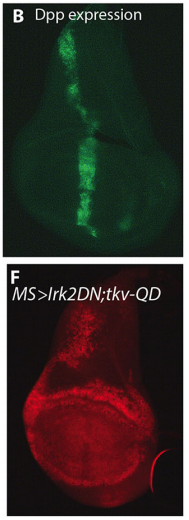
Wing phenotypes following Irk2DN expression, from Figure 1, Dahal, et al. 2017
Can you give us key results of the paper in a paragraph?
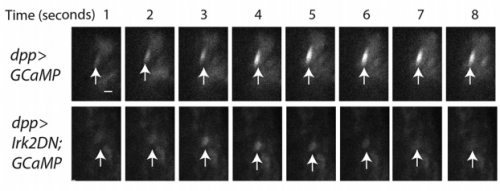
EB/GD/SP We knew that Irk channels played a role in Dpp signalling from our first paper that was also published in Development: Dahal et al 2012. The goal of our work since then has been to determine the molecular mechanism. We found that human Kir2.1 could substitute for fly Irk channels, showing that the developmental role of Irk/Kir channels is conserved between flies and humans. We found that Irk channels are important in the cells that produce Dpp and not in the cells that receive the signal. Inhibition of Irk channels in the Dpp-producing cells does not decrease the amount of Dpp that is made, but it changes the temporal dynamics of Dpp release. Irk channels modulate intracellular calcium levels to influence releases of insulin from pancreatic beta cells and neurotransmitter from neurons, so we hypothesized that Irk channels could regulate Dpp release via the same mechanism. Therefore, we looked for calcium activity in the wing disc. We found native transient increases in intracellular calcium that were altered by inhibition of Irk channels. Lastly, we found that depolarizing cells by applying a potassium solution to the extracellular bath caused transient increases in intracellular calcium and Dpp release.
Why might the timing, and not just the bulk amount, of Dpp release be important to development?
EB/GD/SP The timing of exposure to Transforming Growth Factor-beta (TGF- β) impacts the transcriptional response in tissue culture. For example pulsed exposure to TGF-B had a much greater transcriptional response than constant exposure to the same concentration of the ligand (See Sorre and Warmflash 2014). Dpp belongs to the TGF- β superfamily. Perhaps exposure to these ligands must be pulsed to signal efficiently. There are other examples of this in biology such as insulin signalling affecting metabolism and neurotransmitter affecting neurons. Similarly, the timing of notch signalling in somitogenesis is essential for its effect on the development of somites.
Clones in the wing disc, from Figure 2, Dahal, et al. 2017
How similar is Dpp release from wing cells to neurotransmitter release from synapses?
EB/GD/SP Irk channels help to maintain resting membrane potential in neurons and therefore help to determine when a neuron will “fire” or undergo an action potential, increase intracellular calcium, and release neurotransmitter. In the context of the wing disc, we have shown that Irk channels influence calcium activity and Dpp release. We do not yet know if the release machinery is the same for neurotransmitter release and Dpp release.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
GD I would rather like to say that I have experienced surprising moments instead of a eureka because when it happens my first inclination is to think it is a fluke, which is true most of the time. By the time you confirm the result with enough samples and rule out other possible explanations the prospective eureka moment disappeared long ago because there is no element of surprise. I had small eureka moments with each new channel tested during the rescue experiment.
SP Yes, there were several. The most memorable were being able to see the regulation of calcium transients in the wing disc cells by Irk and the evoked Dpp release in wing disc cells by depolarizing with high extracellular potassium. These kinds of experiments that can be visually examined leaves one with a long lasting excitement.
Changes in GCaMP6s fluorescence, from Figure 5, Dahal, et al. 2017
And what about the flipside: any moments of frustration or despair?
GD Initially, we tried electrophysical approach to study mechanism of the Irk channels I took a course on the topic, and spent months to learn the technique. However, when I tried to express the fly Irk channels in Xenopus egg to measure current these channels did not express well so we abandoned this approach
SP There were many moments of frustration, despair, bruises and cuts. Thankfully these are like labor pains, once the child is born; they seem to fade away and you are ready to start again.
What are your career plans following this work?
GD For my postdoctoral training, I joined Dr. Howard Rockman lab at Duke University, where I worked to develop exosome-mediated drug delivery system to modulate cardiac function. Gradually, my interest shifted towards the downstream process of drug development. I decided to move out from academia. At present, I work in a company as a part of clinical operation team to conduct oncology trials.
SP I have always been ambitious to explore career challenges. As a child in Nepal I envisioned myself in white lab coat being a scientist somewhere. I first set foot in science with a degree in microbiology and taught in a brand new medical school in Nepal. It was a good job but I did not find myself challenged enough, so over-ambitiously I set my foot in USA to pursue my PhD. The day I graduated I felt overwhelmingly happy. Deep in my heart I was bothered with a question….that now I have achieved my life’s biggest goal….. what next? It may not be easy to live rest of my life without a goal. So I thought about it and set myself another goal and this time I did not want to be over ambitious. I just wanted to carry the responsibility and legacy of this degree with dignity for rest of my life. I feel that I have achieved this to some extent with this post doc paper in ‘Development’, and with being a biology faculty in college. So with this, I will continue to pursue a career in science. This may take different directions depending on the opportunities that open up and the different paths of life. I am open to new experiences and challenges that will build on my foundation in science and research.
And what next for the Bates lab?
EB We are now identifying other ion channels and ion channel associated proteins that are important for wing morphology and trying to determine if the SNARE complex is important for release of Dpp. In addition, we have evidence that Kir/Irk channels influence BMP signalling in mammals for craniofacial and limb development. To understand how Kir functions in mammals, we are extending our studies into mice.
Wing discs from Figure 2, Dahal, et al. 2017
Finally, what do you like to do when you are not in the lab?
EB My husband and I have a new baby girl, so currently my favourite thing to do outside the lab is play with her and watch her play with her Dad. I also enjoy family hikes and anything outdoors.
GD I live with my wife and two children. I spend most of our time outside of the lab taking care of our children.
SP Physically and mentally travel to Nepal and elsewhere if I can afford it in terms of time and money. But if you are talking about day-to-day activities, I enjoy playing with my son, being in nature, specially around water. I find the sound of water peaceful. And cooking delightful and delicious food at home.
Our latest monthly trawl for developmental biology (and other cool) preprints. See last year’s introductory post for background, and let us know if we missed anything
This month we found lots of plant preprints and plenty of evo-devo work, as well as a series of preprints characterising features of the genome at distinct stages or places during development. The preprints were hosted on bioRxiv, PeerJ and arXiv. Use these links to get to the section you want:
In vivo validation of predicted E12.5 enhancers, from Gorkin, et al.’s preprint
Systematic mapping of chromatin state landscapes during mouse development. David Gorkin, Iros Barozzi, Yanxiao Zhang, Ah Young Lee, Bin Lee, Yuan Zhao, Andre Wildberg, Bo Ding, Bo Zhang, Mengchi Wang, J. Seth Strattan, Jean M Davidson, Yunjiang Qiu, Veena Afzal, Jennifer A Akiyama, Ingrid Plajzer-Frick, Catherine S Pickle, Momoe Kato, Tyler H Garvin, Quan T Pham, Anne N Harrington, Brandon J Mannion, Elizabeth A Lee, Yoko Fukuda-Yuzawa, Yupeng He, Sebastian Preissl,Sora Chee, Brian A Williams, Diane Trout, Henry Amrhein, Hongbo Yang, J. Michael Cherry, Yin Shen, Joseph R Ecker, Wei Wang, Diane E Dickel, Axel Visel, Len A Pennacchio, Bing Ren
Spatiotemporal DNA Methylome Dynamics of the Developing Mammalian Fetus. Yupeng He, Manoj Hariharan, David U Gorkin, Diane E Dickel, Chongyuan Luo, Rosa G Castanon, Joseph R Nery, Ah Young Lee, Brian A Williams, Diane Trout, Henry Amrhein,Rongxin Fang, Huaming Chen, Bin Li, Axel Visel, Len A Pennacchio, Bing Ren, Joseph R Ecker
Genetic variation and gene expression across multiple tissues and developmental stages in a non-human primate. Anna J. Jasinska, Ivette Zelaya, Susan K. Service, Christine Peterson, Rita M. Cantor, Oi-Wa Choi, Joseph DeYoung, Eleazar Eskin, Lynn A. Fairbanks, Scott Fears, Allison Furterer, Yu S. Huang, Vasily Ramensky, Christopher A. Schmitt, Hannes Svardal, Matthew J. Jorgensen, Jay R. Kaplan, Diego Villar, Bronwen L. Aken, Paul Flicek, Rishi Nag, Emily S. Wong, John Blangero, Thomas D. Dyer, Marina Bogomolov, Yoav Benjamini, George M. Weinstock, Ken Dewar, Chiara Sabatti, Richard K. Wilson, J. David Jentsch, Wesley Warren, Giovanni Coppola, Roger P. Woods, Nelson B. Freimer
An Algorithm for Cellular Reprogramming. Scott Ronquist, Geoff Patterson, Markus Brown, Stephen Lindsly, Haiming Chen, Lindsey Muir, Max Wicha, Anthony Bloch, Roger Brockett, Indika Rajapakse
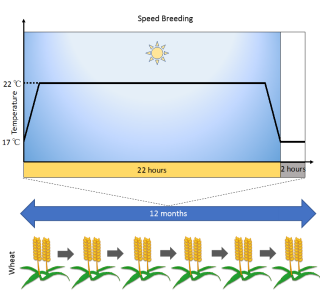
Speed breeding: a powerful tool to accelerate crop research and breeding. Amy Watson, Sreya Ghosh, Matthew Williams, William S. Cuddy, James Simmonds, Maria-Dolores Rey, M. Asyraf Md Hatta, Alison Hinchliffe, Andrew Steed, Daniel Reynolds, Nikolai Adamski, Andy Breakspear, Andrey Korolev, Tracey Rayner, Laura E. Dixon, Adnan Riaz, William Martin, Merrill Ryan, David Edwards, Jacqueline Batley, Harsh Raman, Christian Rogers, Claire Domoney, Graham Moore, Wendy Harwood, Paul Nicholson, Mark J. Dieters, Ian H. DeLacy, Ji Zhou, Cristobal Uauy, Scott A. Boden, Robert F. Park, Brande B. H. Wulff, Lee T. Hickey
Repeat associated mechanisms of genome evolution and function revealed by the Mus caroli and Mus pahari genomes. David Thybert, Maša Roller, Fábio C. P. Navarro, Ian Fiddes, Ian Streeter, Christine Feig, David Martin-Galvez, Mikhail Kolmogorov, Václav Janoušek, Wasiu Akanni, Bronwen Aken, Sarah Aldridge, Varshith Chakrapani, William Chow, Laura Clarke, Carla Cummins, Anthony Doran, Matthew Dunn, Leo Goodstadt, Kerstin Howe, Matthew Howell, Ambre-Aurore Josselin, Robert C. Karn, Christina M. Laukaitis, Lilue Jingtao, Fergal Martin, Matthieu Muffato, Michael A. Quail, Cristina Sisu, Mario Stanke, Klara Stefflova, Cock Van Oosterhout, Frederic Veyrunes, Ben Ward, Fengtang Yang, Golbahar Yazdanifar, Amonida Zadissa, David Adams, Alvis Brazma, Mark Gerstein, Benedict Paten, Son Pham, Thomas Keane, Duncan T. Odom, Paul Flicek
Signatures of the evolution of parthenogenesis and cryptobiosis in the genomes of panagrolaimid nematodes. Philipp H. Schiffer, Etienne Danchin, Ann M. Burnell, Anne-Marike Schiffer, Chris Creevey, Simon Wong, Ilona Dix, Georgina O’Mahony, Bridget A. Culleton, Corinne Rancurel, Gary Stier, Elizabeth A. Martinez-Salazar, Aleksandra Marconi, Urmi Trivedi, Michael Kroiher, Michael A. S. Thorne, Einhard Schierenberg, Thomas Wiehe, Mark Blaxter
Genome expansion and lineage-specific genetic innovations in the world’s largest organisms (Armillaria). Gyorgy Sipos, Arun N Prasanna, Mathias C Walther, Eoin O’Connor, Balazs Balint, Krisztina Krizsan, Brigitta Kiss, Jaqueline Hess, Jason Slot, Robert Riley, Bettina Boka, Daniel Rigling, Kerrie Barry, Juna Lee, Sirma Mihaltseva, Kurt Labutti, Anna Lipzen, Rose Waldron, Nicola Moloney, Christoph Sperisen, Laszlo Kredics, Csaba Vagvolgyi, Andrea Patrigniani, David Fitzpatrick, Istvan Nagy, Sean Doyle, James B Anderson, Igor V Grigoriev, Ulrich Guldener, Martin Munsterkotter, Torda Varga, Laszlo G Nagy
Loss Of PTEN Promotes Formation Of Signaling-Capable Clathrin-Coated Pits. Luciana K. Rosselli-Murai, Joel A. Yates, Sei Yoshida, Julia T. Bourg, Kenneth K. Y. Ho, Megan White, Julia Prisby, Xinyu Tan, Megan Altemus, Liwei Bao, Zhi-Fen Wu, Sarah L. Veatch, Joel A. Swanson, Sofia D. Merajver, Allen P. Liu
Versatile open software to quantify cardiomyocyte and cardiac muscle contraction in vitro and in vivo. Luca Sala, Berend J. van Meer, Leon G. J. Tertoolen, Jeroen Bakkers, Milena Bellin, Richard Davis, Chris Denning, Michel A. E. Dieben, Thomas Eschenhagen, Elisa Giacomelli, Catarina Grandela, Arne Hansen, Eduard R. Holman, Monique R. M. Jongbloed, Sarah M. Kamel, Charlotte D. Koopman, Quentin Lachaud, Ingra Mannhardt, Mervyn P. H. Mol, Valeria V. Orlova, Robert Passier, Marcelo C. Ribeiro, Umber Saleem, Godfrey L. Smith, Christine L. Mummery, Francis L. Burton
Traction force screening enabled by compliant PDMS elastomers. Haruka Yoshie, Newsha Koushki, Rosa Kaviani, Kavitha Rajendran, Quynh Dang, Amjad Husain, Sean Yao, Chuck Li, John K. Sullivan, Magali Saint-Geniez, Ramaswamy Krishnan, Allen Ehrlicher
One of Zimmerman, et al.’s antibodies and its target
P53 toxicity is a hurdle to CRISPR/CAS9 screening and engineering in human pluripotent stem cells. Robert J. Ihry, Kathleen A. Worringer, Max R. Salick, Elizabeth Frias, Dan Ho, Kraig Theriault, Sravya Kommineni, Julie Chen, Marie Sondey, Chaoyang Ye, Ranjit Randhawa, Tripti Kulkarni, Zinger Yang, Gregory McAllister, Carsten Russ, John Reece-Hoyes, William Forrester, Gregory R. Hoffman, Ricardo Dolmetsch, Ajamete Kaykas
Genome-wide genetic data on ~500,000 UK Biobank participants. Clare Bycroft, Colin Freeman, Desislava Petkova, Gavin Band, Lloyd T Elliott, Kevin Sharp, Allan Motyer, Damjan Vukcevic, Olivier Delaneau, Jared O’Connell, Adrian Cortes, Samantha Welsh, Gil McVean, Stephen Leslie, Peter Donnelly, Jonathan Marchini
Soon to be dosed fish from Monstad-Rios, Watson & Kwon’s preprint
Best Practice Data Life Cycle Approaches for the Life Sciences. Philippa C. Griffin, Jyoti Khadake, Kate S. LeMay, Suzanna E. Lewis, Sandra Orchard, Andrew Pask, Bernard Pope, Ute Roessner, Keith Russell, Torsten Seemann, Andrew Treloar, Sonika Tyagi, Jeffrey H. Christiansen, Saravanan Dayalan, Simon Gladman, Sandra B. Hangartner, Helen L. Hayden, William W. H. Ho, Gabriel Keeble-Gagnère, Pasi K. Korhonen, Peter Neish, Priscilla R. Prestes, Mark F. Richardson, Nathan S. Watson-Haigh,Kelly L. Wyres, Neil D. Young,Maria Victoria Schneider
Designing an intuitive web application for drug discovery scientists. Nikiforos Karamanis, Denise Carvalho-Silva, Jennifer A. Cham, Luca Fumis, Samiul Hasan, David Hulcoop, Gautier Koscielny, Michael Maguire, William Newell, ChuangKee Ong, Eliseo Papa, Andrea Pierleoni, Miguel Pignatelli, Sangya Pundir, Francis Rowland, Jessica Vamathevan, Xavier Watkins, Jeffrey C. Barrett, Ian Dunham
The York Gospels: a one thousand year biological palimpsest. Matthew D. Teasdale, Sarah Fiddyment, Jiří Vnouček, Valeria Mattiangeli, Camilla Speller, Annelise Binois, Martin Carver, Catherine Dand, Timothy P. Newfield, Christopher C. Webb, Daniel G. Bradley, Matthew J. Collins
Here are the highlights from the current issue of Development:
X-citing insights into dosage compensation
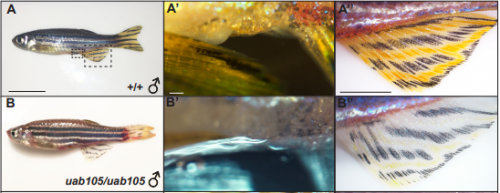
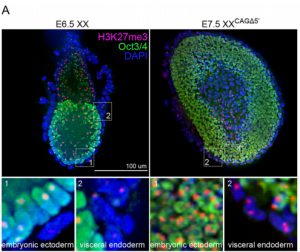
The non-coding RNA Xist plays a key role in the process of X chromosome inactivation (XCI) and is thus essential for dosage compensation of X-linked genes in females. The 5′ region of Xist RNA contains a conserved element termed the A-repeat that is required for the silencing function of Xist in embryonic stem cells, but how this region functions during development is unclear. Now, Takashi Sado and co-workers explore this by introducing into mice a mutated Xist allele that produces Xist RNA lacking the A-repeat region (p. 2784). They first report that imprinted XCI is compromised upon paternal transmission of this allele. The authors further show that the mutant form of Xist is able to coat the X chromosome but fails to silence it in embryonic and extraembryonic tissues. Surprisingly, however, mutant Xist RNA is still able to silence a subset of genes in the trophoblast. Finally, the authors reveal that the failure of imprinted XCI has a more significant impact on genome-wide gene expression than expected; changes in the expression of both X-linked and autosomal genes are observed. Together these findings provide new insights into Xist-mediated gene silencing but also raise the intriguing possibility that dosage compensation regulates X-linked genes as well as gene expression more globally.
Non-neural roles for acetylcholinesterase
Acetylcholinesterase (AChE) is a highly conserved protein that is known for its essential role in degrading the neurotransmitter acetylcholine at neural synapses. However, it is expressed more broadly, outside the nervous system, suggesting that it may carry out additional functions. Now, on p. 2764, Nanette Nascone-Yoder and colleagues reveal that AChE plays an essential non-classical role in Xenopus gut morphogenesis. By exposing tailbud stage Xenopus embryos to AChE inhibitors, or by injecting embryos with morpholinos to knock down AChE in the intestinal endoderm, they show that AChE is required for proper intestinal morphogenesis; in the absence of AChE function, intestines are short/malrotated and exhibit a disorganised epithelium. This function of AChE , they report, is independent of its cholinesterase activity. Further analyses demonstrate that AChE is required for endoderm cell rearrangement and polarisation – events that drive gut lengthening and morphogenesis – as well as endoderm cell differentiation. Finally, the researchers demonstrate that AChE regulates cell-substrate but not cell-cell adhesion. Overall, these results provide direct in vivo evidence for a morphogenetic function for AChE in non-neuronal tissues and suggest that AChE may function in other aspects of development and physiology, a find that has important implications given the widespread use of cholinesterase inhibitors in the treatment of human diseases.
A chemical reset for pluripotency
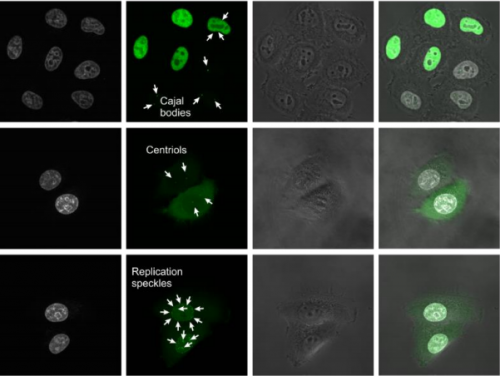
Pluripotency in mammalian stem cells is thought to pass through two phases – an initial naïve phase and a later primed phase, mimicked in vitro by mouse embryonic stem cells and human pluripotent stem cells (hPSCs), respectively. Much effort has gone into converting hPSCs to a more naïve state, but current methods are not always reliable or broadly applicable across cell lines. On p. 2748, Ge Guo, Austin Smith and colleagues provide a simple and efficient method for resetting human pluripotency based on transient inhibition of histone deacetylases (HDACs) with chemical inhibitors. HDAC inhibition leads to increased expression of naïve markers in a variety of different hPSC lines, and cells can be expanded in naïve culture conditions without requiring feeders. Chemically reset cells show a marked transcriptional difference to primed hPSCs and a similarity to epiblast cells of the preimplantation inner cell mass. Reset cells undergo global reduction in DNA methylation, have two active X chromosomes, and can differentiate into multiple lineages. This work provides a protocol for efficient resetting of hPSC pluripotency, and a transcription and methylation resource for further interrogation of the human naïve state.
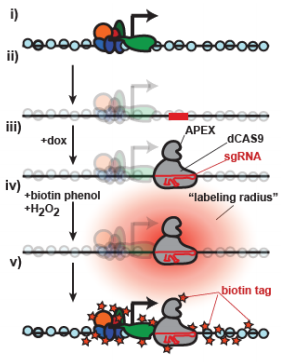
Morphogen signalling: Dpp gets Irked
Mutations in genes encoding ion channels cause severe defects in development across species but the underlying mechanisms, particularly in tissues other than neurons and muscle, have remained unclear. On p. 2771, Emily Anne Bates and colleagues describe a role for Irk2, an inwardly rectifying potassium channel in Drosophilaorthologous to Kir2.1 in vertebrates, in regulating the release of the BMP family morphogen Dpp during development of the wing epithelium. Building on their previous finding that a dominant-negative Irk2 reduces Dpp signalling and causes wing defects, they now show that human KIR2.1 can substitute for inhibited DrosophilaIrk2, and that Irk2’s role is restricted to the cells that produce, rather than just transduce, the Dpp signal. Surprisingly, inhibiting Irk2 broadens the distribution of Dpp in the wing, but also alters the dynamics of Dpp release from cells, suggesting that Irk2 controls the timing of Dpp secretion. Irk2 inhibition reduces the amplitude of calcium spikes in wing cells, and depolarising the membrane with extracellular potassium leads to an overall increased release of Dpp, implicating Irk2’s regulation of membrane potential in Dpp release. Together, these results suggest that ion channels influence tissue morphogenesis by regulating the release of morphogens from the membrane.
Development announces changes to the editorial team and invites community input in choosing a new Editor in Chief and in suggesting future areas for the journal to explore
Jim Smith, recently knighted for services to medical research and science education, talks about his research career and his hopes for the future of biomedical science.
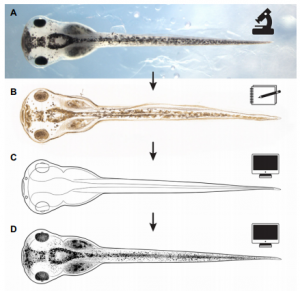
New, freely available illustrations of developing Xenopus laevis, drawn by Natalya Zahn, provide a resource for teaching and research, especially in the field of craniofacial biology.
This Meeting Review discusses recent progress in our understanding of neuronal programming, highlighting some of the common features of cell fate determination during development and directed reprogramming.
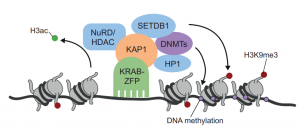
This Primer summarises our current understanding of the intriguing family of KRAB-ZFP transcriptional regulators and its contribution to the control, evolution and co-option of transposable elements.
 (No Ratings Yet)
(No Ratings Yet)

 (13 votes)
(13 votes)









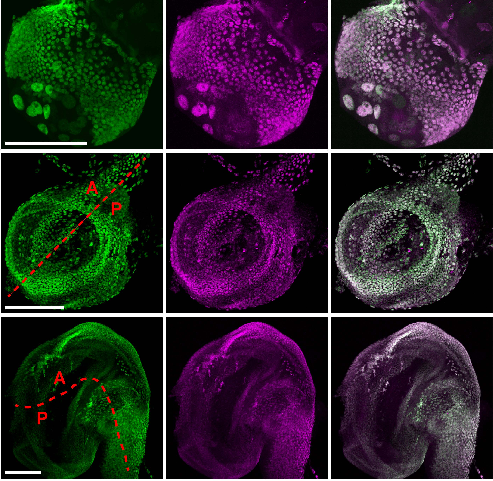
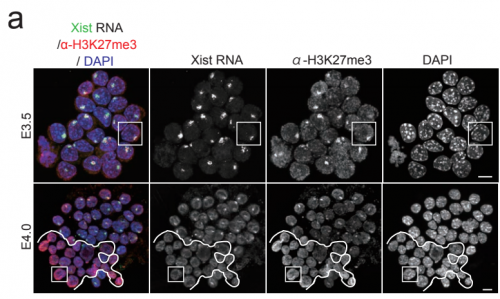
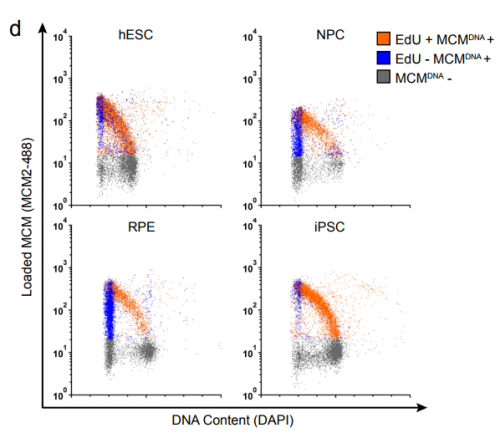
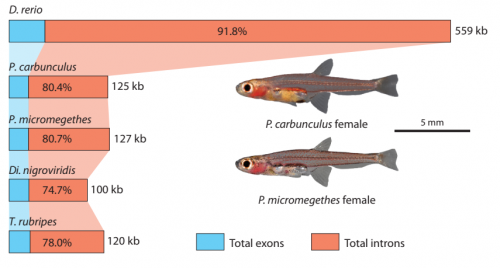




 The 5′ region of Xist RNA contains a conserved element termed the A-repeat that is required for the silencing function of Xist in embryonic stem cells, but how this region functions during development is unclear. Now, Takashi Sado and co-workers explore this by introducing into mice a mutated Xist allele that produces Xist RNA lacking the A-repeat region (p.
The 5′ region of Xist RNA contains a conserved element termed the A-repeat that is required for the silencing function of Xist in embryonic stem cells, but how this region functions during development is unclear. Now, Takashi Sado and co-workers explore this by introducing into mice a mutated Xist allele that produces Xist RNA lacking the A-repeat region (p. 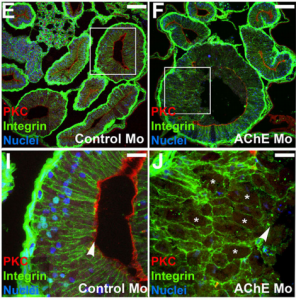 Now, on p.
Now, on p.  On p.
On p.  On p.
On p.