PhD position opened in Switzerland for a developmental or cell biologist
Development & Evolution of structural colours in vertebrates
In the context of a multidisciplinary study combining cell biology, evolutionary developmental biology and physics, we offer one PhD student position for an outstanding, highly motivated, and creative experimental wet-lab biologist with strong skills in developmental biology or cell biology.
A major research project in Michel Milinkovitch’s group (Laboratory of Artificial & Natural Evolution; www.lanevol.org) at the University of Geneva (UNIGE) is to better understand the molecular developmental mechanisms generating a diversity of skin colours and patterns in reptiles.
One aim is to understand the development & evolution of surface gratings and intracellular nano-structures generating colours through the physical phenomenon of light interference. The successful candidates will use state-of-the-art microscopy and biochemical/genetic manipulations in both the zebrafish and chameleons to understand the development and evolution of surface gratings and intracellular photonic crystals.
Candidates must have a Master in biology or biochemistry. Skills and experience with developmental biology and/or cell biology are mandatory. The successful candidate will have a genuine interest for organismal biology and will appreciate interactions with developmental biologists, evolutionary biologists, physicists and computer scientists. The PhD student will be localised in the Milinkovitch lab (http://www.lanevol.org) and will have very regular interactions/collaborations with members of Marcos Gonzalez-Gaitan’ lab (https://cms.unige.ch/sciences/biochimie/-Marcos-Gonzalez-Gaitan-Lab-.html).
The University of Geneva is world-renowned for its research in Biology and Physics and is among the top 1% best universities in the world.
PhD students are remunerated according to the standards of UNIGE, which are very generous when compared to other international programs.
Geneva is an international city occupying a privileged geographical situation with its beautiful lake and the close-by Alps.
Refs: Saenko et al.Precise colocalization of interacting structural and pigmentary elements generates extensive color pattern variation in Phelsuma lizards.BMC Biology 2013, 11: 105; Teyssier et al.Photonic Crystals Cause Active Colour Change in Chameleons.Nature Communications 6: 6368 (2015); Tzika et al. Reptilian Transcriptomes v2.0: An Extensive Resource for Sauropsida Genomics and Transcriptomics. Genome Biol. Evol. 7: 1827-1841 (2015); Ullate-Agote et al.The genome sequence of the corn snake (Pantherophis guttatus), a valuable resource for EvoDevo studies in squamates.Int. J. Dev. Biol. 58: 881-888 (2014); Saenko et al.Amelanism in the corn snake is associated with the insertion of an LTR-retrotransposon in the OCA2 gene.Scientific Reports 5, 17118 (2015); Di-Poï & Milinkovitch. The Anatomical Placode in Reptile Scale Morphogenesis Indicates Shared Ancestry Among Skin Appendages in Amniotes. Science Advances 2, e1600708 (2016).
Candidates must send their application — in the form of a single PDF file including a brief letter of interest, a CV, as well as contact information (not support letters) of two persons of reference — to: Prof. Michel Milinkovitch (michel.Milinkovitch@unige.ch).
We are looking for a postdoc interested in studying neural development in tunicates, the sister group to the vertebrates. The larval nervous system of the tunicate Ciona has only 177 neurons and its entire “connectome” has been recently mapped (Ryan et al., eLife 2016). The Ciona genome is also highly compact and easily manipulated using CRISPR/Cas9 (Stolfi et al., Development 2014). This tractability offers an unprecedented opportunity to understand gene networks underlying the development of every single neuron in a chordate nervous system.
We have several potential projects centered on understanding the link between transcriptional regulation and cell behavior during the development of the Ciona nervous system. Specific cells of interest include homologs of vertebrate spinal cord neurons (Stolfi et al., Development 2011) and neural crest-derived sensory neurons (Stolfi et al., Nature 2015). Projects will involve isolating neural progenitors from transgenic Ciona embryos, profiling their transcriptome and chromatin states using next-generation sequencing, and targeting candidate regulators and effectors using CRISPR/Cas9.
Our lab is located at the Georgia Institute of Technology in Atlanta, USA. Georgia Tech offers first-rate research facilities and a highly collaborative research environment with traditional strengths in engineering, computing, and math. We are dedicated to promoting equity of under-represented minorities in academia.
Position highlights: The Grant Strategist will support the productivity of Zuckerman Institute scientists by editing and consulting on grants, particularly sections on research strategy, aims, and narratives; acquire and maintain sound knowledge of the Institute and the research priorities; use that knowledge and understanding to seek and recommend grants for individual as well as multi-investigator grants within and between disciplines.
The Grant Strategist will serve as a project manager to pursue, identify and promote funding opportunities for multi-PI grants. The Grant Strategist will also consult with and keep abreast of the current granting environment to provide suggestions to individual researchers on specific grant opportunities.
Position requirements: Ph.D. in the biological sciences or related field, including extensive writing experience; or advanced degree in journalism with deep expertise in science. Minimum of four (4) years of successful science grant writing experience or related experience in science communication field is required. Extensive knowledge of scientific granting processes, particularly in the biological sciences; successful grant writing or consulting record is vital.
When blood goes bad, a replacement is often needed. Each year, thousands of patients in the US receive bone marrow transplants to treat life-threatening diseases like blood cancer.
But in some cases, the transplant itself can become deadly.
The problem is not necessarily the one most people think of: rejection of the transplanted material. Instead, the donor hematopoietic stem cells (HSCs), delivered to repopulate the whole blood system, sometimes produce immune cells that attack anything foreign. In these cases, the transplant rejects the patient. This condition, called graft-versus-host disease (GVDH), is common in transplant patients and often fatal.
Developing a reliable method to make transplantable stem cells that would avoid GVHD is of considerable interest. That’s why we and others have been interested in learning how to direct the differentiation of HSCs in vitro from pluripotent precursor cells.
Intuitively, a way to make new HSCs would be to replicate the same instructions used to make them in the developing embryo. But first we have to identify the signals that make up those instructions. During embryonic development, HSCs are born from the descending aorta, the primitive vessel that carries blood to the lower half of the body. Decades of research have uncovered numerous signaling inputs that direct formation of the descending aorta, and its subsequent conversion to the first HSCs. However, we are still defining the complete set of signals, and how they are regulated and integrated.
We set out to examine this coordination and identify novel factors that might play critical roles in HSC development. We used zebrafish because their blood development is nearly identical to that of humans. We noticed a common theme: signaling pathways required for hematopoietic stem cell formation also frequently regulate vessel patterning. Specifically, well-established signaling pathways, like the Vegf, Notch, and Wnt pathways, play a role in both processes.
In the trunk of the embryo, smaller blood vessels called “intersegmental vessels” sprout from the developing descending aorta at about the same time that HSCs are born. We became intrigued by published results identifying a new signaling pathway necessary for the sprouting of intersegmental vessels from the aorta (Gore et al., 2011). This pathway involves R-spondin-1 (Rspo1), a secreted factor that augments Wnt signaling. Intriguingly, Gore et al. also showed rspo1 was expressed in the dorsal aorta during the time of HSC specification, the point when blood stem cells start to assume their future identity. We wondered if this pathway might also be necessary for HSC specification.
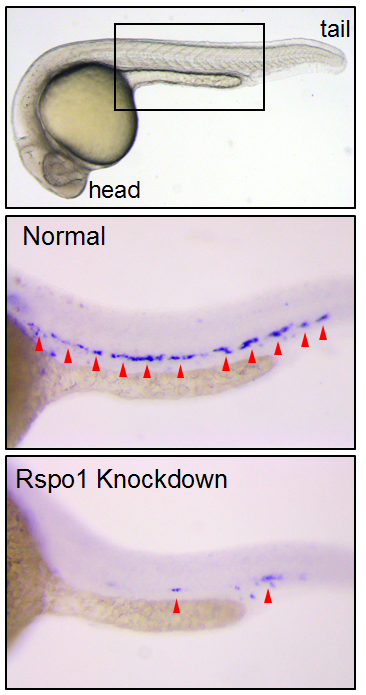
Figure 1. Transgenic zebrafish with GFP-labelled blood vessels. Rspo1 knockdown causes loss of blood vessel growth as seen by decreased intersegmental vessel (ISV) growth, providing a visual means of seeing that Rspo1 is inactivated.
Our first goal was to determine if Rspo1 is required in HSC development. We knocked down Rspo1 in zebrafish embryos using antisense oligonucleotides and asked what would happen to HSCs. Strikingly, markers of HSC development were visibly decreased at all time points we examined. We subsequently confirmed the rspo1 requirement in an established zebrafish hypomorphic mutant.
Figure 2. HSCs marked by runx1 in 24 hours post fertilization zebrafish embryos. Rspo1 knockdown causes loss of HSCs. Red arrowheads indicate HSCs.
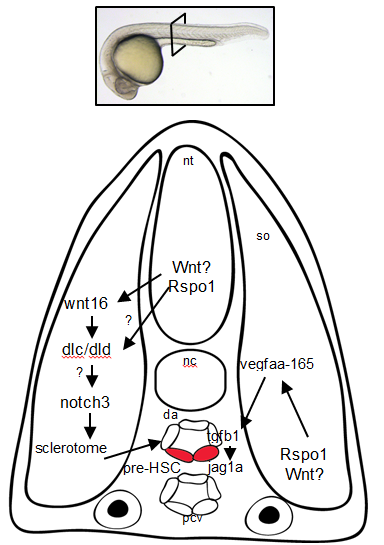
So Rspo1 is required for HSCs to form, but how does it work? To answer this question, we looked at the activity of other pathways already known to play a role in HSC specification to see if we could detect any changes in their activity. Our results identified two of these – the Wnt16 and Vegfa pathways – that showed alterations in expression of their downstream components.
Our findings had some surprises. Most notably, although vegfa expression was lower when Rspo1 was knocked down, a key target of Vegf signaling, notch1b, was unaffected. Our finding suggested that there might be a Notch1-independent role for Vegf in specifying hematopoietic stem cells.
As we were trying to define how this might work, the group of Roger Patient (Monteiro et al., 2016) identified precisely such a pathway. We determined that Rspo1 acted via this Notch-independent pathway, and discovered that a particular Vegfa splice variant was involved.
Figure 3. Rspo1 signaling pathways needed for the development of HSCs. Cross-section of a 24hr old zebrafish embryo.
Overall, these findings point to Rspo1 as a new master regulator of blood stem cells that controls two pathways both needed for the birth of these stem cells. In the future, we hope that understanding the full set of signals and their integration will provide the key to unlocking our ability to make fully functional hematopoietic stem cells in the lab. Eventually we will have the tools to create designer treatments for leukemia and other blood disorders while making GVHD a thing of the past.
Here are the highlights from the new issue of Development:
Adding a new layer of complexity to pre-eclampsia
Pre-eclampsia (PE) is a pregnancy complication associated with abnormal formation of the placenta. To date, most studies of PE have focussed on cytotrophoblasts (CTBs) within the villous placenta (the chorion frondosum); the deficient invasion of these cells into the uterine wall is thought to lead to abnormal placentation and hence PE. Here, on p. 767, Susan Fisher and colleagues reveal that CTBs within a different region of the human placenta – the smooth chorion – are implicated in severe PE. They first report that the CTB layer in the smooth chorion is expanded in severe cases of PE and is less organised. These morphological changes are accompanied by changes in the expression profiles of smooth chorion CTBs. In particular, smooth chorion CTBs exhibit enhanced expression of HLA-G, integrin α4 and E-cadherin, all of which are factors that extravillous CTBs normally modulate as they invade the uterine wall. The researchers further show that severe PE is associated with global gene expression changes in smooth chorion CTBs that are distinct from the transcriptional responses of villous and extravillous CTBs to severe PE, which they previously described. Overall, these findings suggest that smooth chorion CTBs play a greater role in placentation and pregnancy outcome than previously appreciated.
Integrins worm their way into brain regeneration
Tissue regeneration involves a number of cellular processes, including proliferation, differentiation, migration and patterning, but it is not clear how all of these processes are coordinated to allow the correct generation and assembly of cells during regeneration. Using the planarian flatworm Schmidtea mediterranea, which displays remarkable regenerative capacity and can regenerate any missing body part, two new studies reveal a key role for integrins in regulating tissue organisation during brain regeneration.
In the first study (p. 784), Nicolle Bonar and Christian Petersen use RNAi to show that β1-integrin regulates cell number and tissue organisation during regeneration following decapitation; in the absence of β1-integrin, brain tissue is disorganised and ectopic cell aggregates form in the head region. By contrast, global patterning is largely unaffected. The authors further show that β1-integrinRNAi animals exhibit an initial delay in regeneration but that this is followed by tissue overproduction. Finally, they report that integrin signalling, likely via a β1/α-2 complex, is required for the proper localisation of neoblasts and progenitor cells during regeneration. Together, these findings suggest that integrin signalling acts to recruit and localise progenitor cells following injury, thereby promoting the correct organisation of regenerating planarian tissue.
In the second paper (p. 795), Florian Seebeck, Kerstin Bartscherer and colleagues demonstrate that β1-integrin RNAi animals, as well as α-integrin-2 RNAi animals, exhibit impaired regeneration following amputation. They show that, in β1-integrin RNAi animals, the newly formed tissues – including the muscle and gut – display structural defects. The researchers also reveal that β1-integrin is required for neoblast migration towards the wound site. Finally, they report that β1-integrin RNAi causes the formation of ectopic neural spheres within the regenerating brain region that are composed of various neuronal cell types and that undergo continuous growth. Overall, these results suggest that integrins are required for the formation of organised tissues and for restricting neurogenesis during planarian regeneration.
Apoptosis: a delayed gut reaction to bacteria
The intestine is a tissue that is known to undergo regeneration, both continuously as part of tissue homeostasis and in response to damage – for example, that induced by bacterial aggression. While many studies have examined how the gut responds to large amounts of pathogenic or opportunistic bacteria, it is unclear how low levels of bacteria might influence gut homeostasis. Here, on p. 808, Armel Gallet and co-workers tackle this issue. They report that small amounts of the opportunistic Gram-positive bacterium Bacillus thuringiensis var. kurstaki induce a mild early stress response mediated by JNK signalling in the Drosophila midgut. This, in turn, induces the proliferation of intestinal stem cells and leads to the accumulation and overcrowding of differentiated intestinal cells (enterocytes). The authors further report that low amounts of ingested bacteria do not trigger apoptosis, whereas larger amounts do. However, they find that a wave of apoptosis is observed days after infection and acts to eliminate the excess enterocytes. Finally, they demonstrate that the Hippo pathway functions cell-autonomously to trigger the removal of supernumerary enterocytes. These findings lead the authors to propose that the mechanisms involved in the response to the ingestion of low amounts of opportunistic bacteria are different to those mediating the ʻregenerative cell deathʼ that occurs following a stronger aggression.
Nucleogenesis gets active
Neurons within the central nervous system can assemble into clusters, termed nuclei, that house neurons with similar synaptic inputs, outputs and function. This process of nucleogenesis, which is crucial for correct circuit formation, is poorly understood. Now, on p. 830, Sarah Guthrie and colleagues show that the correct assembly of developing motor neurons into nuclei in the chick brainstem requires interplay between spontaneous activity, type II cadherins and gap junctions. Using the genetically encoded calcium indicator GCaMP6, they first show that facial motor neurons exhibit activity patterns that change over the course of nucleogenesis. These patterns can be disrupted by perturbing the expression of the type II cadherin Cad20 or the gap junction protein Cx43. The authors further demonstrate that the inhibition of spontaneous activity (using calcium channel inhibitors) results in neuronal disaggregation, and also causes a reduction in the levels of Cad13, another type II cadherin, suggesting the presence of a feedback loop. In summary, these observations suggest that a network of interactions between cadherins, gap junctions and spontaneous activity governs nucleogenesis.
PLUS:
Creating to understand – developmental biology meets engineering in Paris
In November 2016, developmental biologists, synthetic biologists and engineers gathered in Paris for a meeting called ‘Engineering the embryo’. The participants shared an interest in exploring how synthetic systems can reveal new principles of embryonic development, and how the in vitro manipulation and modeling of development using stem cells can be used to integrate ideas and expertise from physics, developmental biology and tissue engineering. In their Meeting Review, Anna Kicheva andNicolas Rivron provide a summary of this meeting and highlight the challenges arising at the intersection of these fields.
Krüppel-like factors in mammalian stem cells and development
Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their Primer, Agnieszka Bialkowska, Vincent Yang andSandeep Mallipattu review key functions for KLFs in mammalian embryogenesis, stem cells and regeneration.
Plasticity in the lung: making and breaking cell identity
In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their Review, Purushothama Tata andJayaraj Rajagopal summarize our current understanding of the different cell lineages of the adult mammalian lung and discuss how these populations respond to injury.
Reviews Editor: biomedical and life sciences journals
Based in Cambridge, UK
Applications are sought for the role of Reviews Editor for The Company of Biologists’ journals: Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open. The role will be offered for an initial term of three years.
Joining a long-established and successful publishing company, this is an exciting opportunity to make a significant contribution to our highly respected biomedical and life science journals. The role will initially focus on Disease Models & Mechanisms, a growing Open Access journal in the fast-moving field of translational research.
The front sections of the journals include commissioned reviews and poster articles, thought-provoking editorials and interviews with leaders in the field.
Applicants will have a PhD or MD in a relevant area. Post-doctoral and/or previous editorial experience is desirable, although we will provide full training. The successful candidate will work alongside an established publishing team in our Cambridge office.
Core responsibilities include:
• Commissioning, handling peer review and developmental editing of material for the front section of the journals.
• Travelling to international scientific conferences and research institutes, representing the journals, keeping abreast of the latest research and making contacts in the community.
• Spotting newsworthy articles, writing informative press releases and handling any media enquiries.
• Interviewing high-profile scientists.
• Contributing to our social media output.
• Creative involvement in the journals’ development and marketing activities.
Additional responsibilities may be provided for the right candidate.
Essential requirements for the job are enthusiasm, commitment, judgement and integrity. Candidates should have excellent interpersonal skills and confidence, and excellent oral and written communication skills. The successful candidate will have a broad interest in science, the scientific community and publishing.
The position is full time and will be based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK. It offers an attractive salary and benefits.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals –Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV by email to recruitment@biologists.com along with a covering letter that states their current salary and any limitations regards possible start date, and summarises their relevant experience and why they are enthusiastic about this opportunity.
Applicants should be eligible to work in the UK and should be able to travel internationally. Applications should be received by 20th March 2017.
Alana I. Mendelsohn, Departments of Neuroscience and Biochemistry and Molecular Biophysics, Columbia University
Jeremy S. Dasen, Department of Neuroscience, NYU
Thomas M. Jessell, Departments of Neuroscience and Biochemistry and Molecular Biophysics, Columbia University
The ability of the nervous system to perform complex tasks depends on a diverse set of circuits that underlie specific behaviors. The nervous system generates distinct neural circuits in several ways. One strategy is to make many different types of neurons, varying individual properties such as cellular morphology, settling position, electrical conductance and pharmacological signaling. The other method of forming distinct circuits is to have neurons form selective connections with only a subset of other neurons. Given the scale of the human nervous system, where 100 billion neurons each forms an average of 7000 synaptic connections, the establishment of defined neural circuits is one of the most complex facets of organismal development. The scientific challenge of identifying mechanisms underlying neural specification and circuit formation is in large part what drew me to Tom Jessell’s lab as a graduate student. Tom’s lab works on the mammalian motor system, which carries a number of advantages for studying fundamental principles of nervous system development. Namely, the spinal cord is an excellent model system for studying development because it has well characterized and relatively stereotyped circuit anatomy, with defined sensory inputs and topographically segregated motor outputs.
The first project I worked on in graduate school looked at the formation of monosynaptic reflex circuits between sensory neurons and motor neurons, a process long-thought to be under total genetic control. Although our work showed that the development of these circuits is in fact shaped to some extent by neural activity, it also made clear that one fundamental aspect of circuit formation, the choice of synaptic partner, was made in this context solely on the basis of molecular recognition (Mendelsohn et al., 2015). With this finding in mind, I decided follow up this project by designing a transcriptional screen of different motor pools in mouse, with the intention of identifying molecular surface recognition molecules that might underlie selective circuit formation. Instead, the results of this screen led me down a very different scientific path, towards addressing how motor neurons acquire distinct subtype identities.
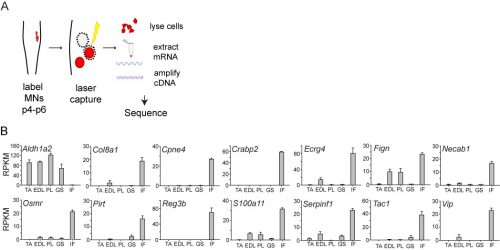
In initially following up the results of the screen, I was unable to identify new genes that could plausibly account for differences in sensory-motor connectivity, or even genes that could distinguish motor neurons projecting to different muscles in the leg. But to my surprise, the screen also revealed something completely unexpected: a large number of genes expressed selectively in the motor neurons that project to the intrinsic muscles of the hand and foot. Moreover, I also found that one particularly important gene, Aldh1a2, which encodes the retinoic acid synthesis enzyme Raldh2, was absent in these motor neurons. Given the historical difficulty of defining selective markers of motor pools, this was an exciting and shocking finding.
(A) Schema for motor pool genetic screen experiment. (B) Pool-specific levels of gene expression in reads per kilobase per million. Pools profiled were tibialis anterior (TA), extensor digitorum longus (EDL), peroneus longus (PL), gastrocnemius (GS) and intrinsic foot (IF).
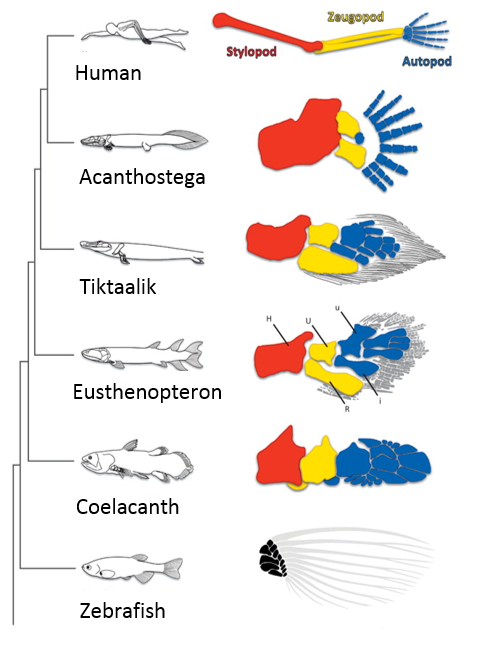
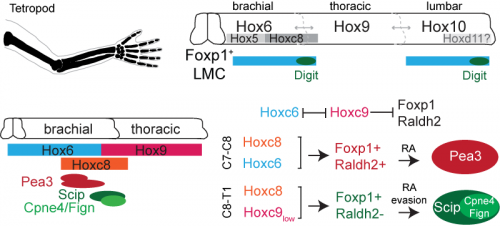
When I first showed these results to Tom, he immediately grasped that they carried profound consequences beyond the scope of developmental neurobiology. The formation of the motor system does not occur in a vacuum, as the development of motor circuits has to correspond with the formation of muscles in the limb that receive motor innervation. In fact, the formation of the limb and motor system use many similar patterning mechanisms, including Shh, FGFs, BMPs, retinoic acid and Hox genes (Cooper et al., 2011; Dasen and Jessell, 2009; Mercader et al., 2000; Roselló-Díez et al., 2011; Zakany and Duboule, 2007). Consequently, Tom realized that the divergent transcriptional profile of digit-innervating motor neurons could have something to do with the natural history of digits themselves. Thinking now in evolutionary terms, Tom quickly sent me off to read Neil Shubin’s best-selling book “Your Inner Fish,” along with a stack of academic papers on, among other things, the embryology of shark and paddlefish fins. I soon learned that evolutionary biologists have long been concerned with the question of how digits formed during the transformation of fish fins into tetropod limbs (Shubin et al., 1997). It appears that the emergence of distal limb elements is a specialized step in limb patterning, requiring evolutionarily conserved molecular determinants (Nakamura et al., 2016; Schneider and Shubin, 2013; Woltering and Duboule, 2010). As a result, we started to wonder whether the specialized evolutionary emergence of digits might explain why we found such divergent patterns of gene expression in digit-innervating motor neurons.
The fossil lineage shows the progressive elaboration of the distal skeleton and appearance of digits. Adapted from (Schneider and Shubin, 2013).
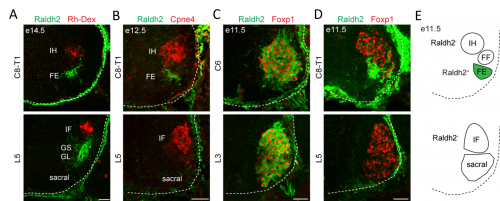
In order to clarify the mechanisms underlying digit-innervating motor neuron specification, we first sought to characterize the patterns of gene expression that emerged from the screen using a combination of in situ hybridization, immunohistochemistry and retrograde anatomical tracing. This work revealed several markers, including Cpne4 and Fign, which selectively labeled digit-innervating motor neurons throughout embryonic development. It also confirmed that digit-innervating motor neurons fail to make retinoic acid from the moment of their specification. In looking at patterns of gene expression under the microscope day after day, I stumbled on another interesting way in which digit-innervating motor neurons are unique. Unlike other limb-innervating motor neurons, digit-innervating motor neurons express the signaling molecule pSMAD in a manner that is dependent on retrograde cues from the limb. Though intrigued by this finding, we weren’t sure how to proceed given that not much is known about the role of the SMAD family in motor neuron development. On the other hand, it was already well known that retinoic acid is involved in the initial generation of motor neurons, as well as their acquisition of broader divisional identities (Sockanathan et al., 2003). As a result, we decided to focus on the role of retinoic acid signaling in digit-motor neuron development.
(A) Raldh2+ motor neurons are not labeled after Rh-Dex injection into the intrinsic hand and intrinsic foot muscles at e14.5. (B) Cpne4+ intrinsic hand and intrinsic foot motor neurons do not express Raldh2 at e12.5. (C) Rostral LMC motor neurons express Raldh2 at e11.5. (D) Caudal LMC motor neurons do not express Raldh2 at e11.5. (E) Summary of Raldh2 expression by spinal motor neurons.
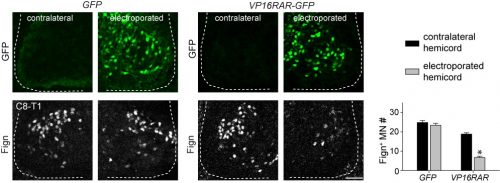
Since digit-innervating motor neurons fail to make retinoids, we wondered whether the absence of retinoic acid was necessary for their development. To address this, we decided to force developing motor neurons to engage in retinoic acid signaling, reasoning that the non-digit innervating motor neurons that already made retinoids would be unaffected. The quickest and easiest way to test this question was to go into chick, where embryologists have long used in ovo electroporation as a technique to manipulate gene expression in developing embryos. We electroporated chicken embryos with a genetic construct in which the retinoic acid receptor RAR was fused to a motif that would render the receptor constitutively active, and found that while other limb-innervating motor neurons were unaffected, the development of digit-innervating motor neurons was selectively disrupted.
Electroporation of VP16RAR-GFP, but not control GFP, reduces the number Fign+ motor neurons at brachial levels.
The finding that retinoid evasion was important for digit-innervating motor neuron development was satisfying for several reasons. One is that retinoic acid is responsible for the downstream regulation of hundreds of other genes, providing a potential explanation for why digit-innervating motor neurons express so many different genes from other limb-innervating motor neurons. A second reason is that the absence of retinoic acid signaling also helped to explain why intrinsic hand and intrinsic foot innervating motor neurons expressed so many of the same genes, even though each pool is located in a different part of the spinal cord and innervates separate limbs. However, we were still left with the nagging question of why digit-innervating motor neurons were failing to make retinoic acid. To address this, we turned to Hox genes.
Hox genes are involved in both limb and nervous system planning, and have been a longstanding subject of interest in Tom’s lab for their role in specifying motor neurons. Much of this work had been carried out by Jeremy Dasen when he was a post-doc in the lab, in a series of classic studies that I first became acquainted and enamored with as an undergraduate student (Dasen et al., 2003, 2005, 2008). In re-reading these studies, as well as more recent work from Jeremy’s lab at NYU, I started to hone in on the role of two Hox genes, Hoxc8 and Hoxc9. Hoxc8 is expressed at brachial levels of the spinal cord and contributes to limb-innervating motor neuron specification, whereas Hoxc9 is expressed at thoracic levels and is thought to block limb-innervating motor neuron identity. What I found interesting is that even though most limb-innervating motor neurons were unaffected by the loss of Hoxc9, there was a surprising reduction in the number of motor neurons in these mutants projecting to the distal limb (Jung et al., 2010). After initially confirming that digit-innervating motor neurons at brachial levels expressed Hoxc8 as well as low levels of Hoxc9, I started to wonder whether they might both be required for their development. This possibility was especially tantalizing given previous evidence that Hoxc9 is capable of blocking the expression of Raldh2 (Dasen et al., 2003, 2008; Jung et al., 2010), possibly serving as an explanation for why digit-innervating motor neurons fail to make retinoic acid. At this point in the project, it was clear that we would benefit from directly enlisting Jeremy’s help.
We first asked whether Hoxc8 and Hoxc9 were necessary for digit-innervating motor neurons specification, examining the expression of digit motor neuron selective markers in Hoxc8 and Hoxc9 mutants, which Jeremy’s lab provided for us (Catela et al., 2016; Jung et al., 2010). We found that whereas all caudal motor neurons, including those projecting to the hand, were lost in the Hoxc8 mutants, only digit-innervating motor neurons were lost in the Hoxc9 mutants. Moreover, when we ectopically expressed Hoxc8 and Hoxc9 together in chick embryos through electroporation, we could induce the generation of digit-innervating motor neurons, suggesting that this combination of gene expression was also sufficient to specify digit-innervating motor neurons.
Digit-innervating motor neurons can be distinguished from motor pools that innervate more proximal muscles in the forearm by their low levels of Hoxc9 expression. The specification of digit-innervating motor neurons at brachial levels requires both Hoxc8 and Hoxc9. At high levels, Hoxc9 blocks expression of Foxp1 and Raldh2. Low levels of Hoxc9 in digit-innervating motor neurons may thus be sufficient to block expression of Raldh2 while maintaining Foxp1. The absence of retinoid production in digit-innervating motor neurons consequently contributes to their proliferation and maintenance.
Together, our Hox and retinoid manipulation studies suggested a model for the specification of digit-innervating motor neurons, implicating low levels of Hoxc9 in regulating the absence of Raldh2 in these neurons. While this work provides an important initial step to clarifying how digit-innervating motor neurons acquire their divergent identity, it raises many more questions than it answers. We still don’t understand the role of pSMAD signaling in these motor neurons or how Hox genes operate at lumbar levels of the spinal cord. We also don’t know how retinoid evasion coordinates the expression of downstream genes or the molecular mechanisms by which digit-innervating motor neurons form appropriate connections with muscle in the periphery. But what made this project so exciting and satisfying to work on was that it emerged spontaneously. I had no idea when I started my PhD that I would end up working with chick embryos or reading about the latest advances in paleontology. The freedom to pursue unforeseen and interesting questions and the joy of unexpected discovery has been the most meaningful part of my graduate work, and one that I will hopefully continue to experience throughout my scientific career.
Jung, H., Lacombe, J., Mazzoni, E.O., Liem, K.F., Grinstein, J., Mahony, S., Mukhopadhyay, D., Gifford, D.K., Young, R.A., Anderson, K. V, et al. (2010). Global control of motor neuron topography mediated by the repressive actions of a single hox gene. Neuron 67, 781–796.
Positions are available in the Gray Laboratory within the Dell Pediatrics Research Institute at University of Texas at Austin Dell Medical School. The primary research focus of these positions will be to utilize novel zebrafish and cartilage cell culture models to study the genetic susceptibilities of spine and cartilage diseases. One of the major focuses of the Gray Lab is to leverage zebrafish and mouse models coupled with cell culture, transcriptomic, and proteomic approaches to understand spine and cartilage development and disease. Trainees should have proven skills working with animal models with zebrafish experience is a plus. Postdoctoral and research associate candidates should provide a cover letter, CV, and contact information for three professional references. Applications will be reviewed immediately and accepted until filled
Send information and applications to:
RYAN SCOTT GRAY, PhD
Assistant Professor
Department of Pediatrics
Dell Medical School | The University of Texas at Austin
The BSDB invites nominations for three new committee members, who will commence their 5 year term in October 2017 replacing three members who will retire this year. Committee members are required to (make every attempt to) attend two committee meetings per year, one of which coincides with the annual BSDB Spring Meeting.
All nominations must be proposed by a member of the BSDB, and seconded by another member, and must also be accompanied by a statement from the nominee that they are willing to serve if elected. Please, send your nominations to secretary@bsdb.org as soon as possible, latest by Friday 7th of March 2017.If a vote is required, the election will take place at this year’s BSDB AGM, which will be held during this year’s Joint Spring Meeting (Warwick, 2-5 April 2017). In this case, a short verbal statement of support for the nominee will be required, preferably presented by the proposer or seconder.
Conjoined twins have fascinated biologists for centuries. In twins joined at the thorax, left-right patterning is disrupted, but only in one half of the right hand twins. Today’s paper, from this week’s issue of Current Biology, tackles this enigmatic phenomenon using Xenopus, and reveals that laterality in conjoined twins is determined by cilia-driven leftward flow. We caught up with lead author and soon-to-graduate PhD student Matthias Tisler, and his PI Martin Blum of Hohenheim University in Germany.
Martin (L) and Matthias (R)
So Martin, can tell us your scientific biography and the aims of the Blum lab?
MB Well, it took some time until I became an embryologist. I trained as a biologist and got my Ph.D. in Karlsruhe, Germany, working with Peter Herrlich on DNA repair and trying to clone the gene defective in patients with Xeroderma pigmentosum. A first postdoc led me to the Biocenter in Basel, where we studied genetic polymorphisms of drug metabolism in the lab of Urs Mayer and cloned the genes and variants responsible for slow acetylation. This is where I met Walter Gehring, whose work I had followed and admired for a very long time. He encouraged me to switch to developmental biology even at a relatively advanced stage of my career, and he introduced me to Eddy De Robertis, with whom I did a second postdoc in LA.
These were exciting years: the lab was at the forefront of unraveling the molecular secrets of Spemann’s organizer and cloned some of the first genes, such as Goosecoid and Chordin. After my return to Germany I set up my own lab and serendipitously cloned Pitx2, in an attempt to clone Gsc-related genes. From that time onwards, my lab has studied left-right asymmetry in various organisms, mouse and frog but also rabbit, pig and even sea urchins. We try to understand why and how left-right organ asymmetry evolved, at what stage cilia became instrumental, and how cilia-driven symmetry breakage works.
What is research like for developmental biology in Stuttgart?
MB Hohenheim is a small university and there is just one other embryology lab working on Drosophila. However, we are very close to Tübingen and Heidelberg, and we have established close ties with other Xenopus labs all over Germany. Therefore, we are not isolated at all, and we have quite a good standing with biology students at all levels that are interested in joining the lab for summer projects or their bachelor or master thesis. The lab usually consists of 5-6 Ph.D. students, 2-3 technicians, a staff scientist and numerous bachelor and master students. We also have two junior groups in the institute, with whom we have lab meetings and seminars together. I love this place, the beautiful campus and the small size which allows you to basically know all your colleagues and which makes for a very pleasant atmosphere.
And Matthias, how did you come to join Martin’s lab?
MT Back in 2006, when I started my studies in Hohenheim, Martin was in charge of introducing molecular cell biology and in the course of his lectures at one point passed a cartilage-stained E14.5 mouse embryo around. I think it was on one of my very first days at Hohenheim University. After this lecture, I was completely fascinated by the beauty and complexity of developmental biology and wanted to join Martin’s lab as soon as possible. After some time, I finally had the courage to ask for a job in the lab, just to get in contact with the people and to get my hands on embryos. Martin gave me a chance…and I think I did well over the years.
What can conjoined twins tell us about development?
MT Dating back to the experiments of Hans Spemann and colleagues, twins and their experimental induction led not only to the discovery of the famous Spemann organizer but also led him to think about the establishment of left-right asymmetry. So twinning has a long tradition in inspiring developmental biology.
MB I couldn’t agree more: conjoined twins have been a great assay ever since Hilde Mangold’s organizer transplantation in newt embryos in the 1920s in Hans Spemann’s laboratory that induced the formation of a conjoined twin on the ventral side. The molecular elucidation of the organizer used this assay time and again to demonstrate the potential of genes such as Wnt8 or cerberus to elicit the organizer phenomenon. For example, many Wnt pathway components have been investigated and their epistatic relationships unravelled in that assay, i.e. the dependence of twinning on the presence of such factors.
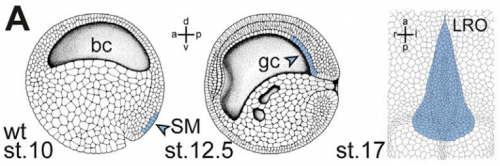
Development of the left-right organiser at the Xenopus gastrocoel roof, from Figure 1, Tisler, et al. 2017. Current Biology.
And why are Xenopus in particular a good model for twinning and laterality?
MT Xenopus is THE ideal model organism to induce a secondary body axis by the (even sided) injections of mRNAs during early stages of development. As frog embryos develop in a petri-dish, they are suitable for additional manipulations like morpholino oligomer-mediated gene knockdowns or – in experiments that we have used in the present study – injection of methylcellulose (wallpaper paste) into the archenteron to block cilia-driven leftward flow. And: all these treatments can be combined, that is only possible in Xenopus embryos.
MB By these sided injections, which Matthias mentioned, one can assess whether a gene works on the left or right side, and the contralateral (uninjected) side always serves as an internal control.
Can you give us the key results of the paper in a paragraph?
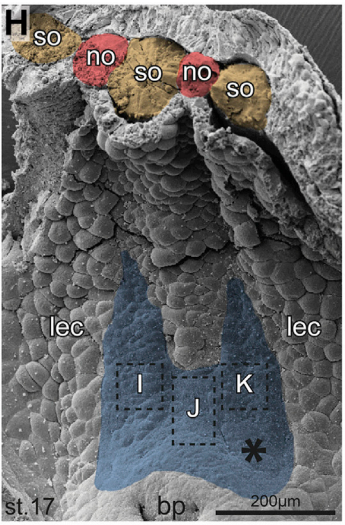
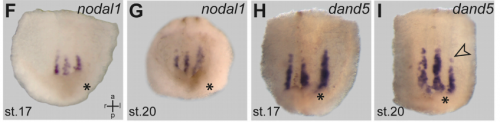
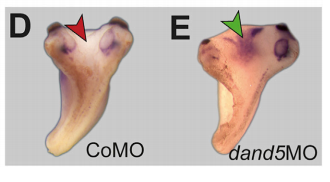
MB & MT The observation that the heart loops normally in left conjoined twins but is randomized in right twins (50% normal, 50% inverted) is old and has been reported in numerous human cases. Spemann reproduced this finding by performing partial ligatures of early stage newt and frog embryos. We injected ß-catenin to induce twinning at will on the left or right side of the endogenous twin and observed the same phenomenon. While two clearly and completely separated precursor tissues of the ciliated left-right organizer (LRO) formed in all cases, the LRO was partially fused, i.e. the right margin of the LRO in the left twin was joined to the left part of the LRO of the right one. Cilia were present on both sides, of normal length and polarity, and they were motile and produced a leftward fluid flow on both sides. Yet the asymmetric Nodal signaling cascade, which determines heart looping and the placement of the other asymmetric organs in the chest and abdomen, was only induced in the left but not in the right twin. This was consistent with the observed heart situs (normal in left and random in right twins), but odd, as flow was normal also in the right twin. A possible solution surfaced when we analyzed the Nodal inhibitor Dand5, which is coexpressed with Nodal at the LRO margins on both sides: in conjoined right twins, flow down-regulates Dand5 like in left ones, but only partially, likely because there is a vast excess available from the fused right side of the left twin. This observation predicted that a knockdown of Dand5 on the right side of the left twin should induce the asymmetric gene cascade also in the right twin, which is exactly what we observed in an experiment, that can only be performed in Xenopus. To round this off, we manipulated Dand5 and flow (using methylcellulose) in a sided manner and were able to induce the Nodal cascade in twins at will, which demonstrated that flow and its target Dand5 also determine laterality in twins, just as in singletons.
Scanning electron micrograph of representative twinned gastrocoel roof plate, from Figure 1, Tisler, et al. 2017. Current Biology.
And your results overcome any final objections to the role of cilia in breaking symmetry?
MB & MT A role for cilia-mediated symmetry breakage in fish, amphibian and mammalian embryos has been demonstrated in many genetic and embryological studies. However, it has been argued that cilia-driven leftward flow merely amplifies an earlier asymmetry which is present already during early cleavage stages (flow only sets in during neurulation). This argument was mostly based on organ situs determination in conjoined twins, which is triggered by activation of the Spemann organizer on the ventral side. Because the organizer acts during gastrulation, it was assumed that cilia could not directly impact on laterality determination. Our work now shows that the initially clearly separated axes fuse in a way that flow in the right twin is insufficient to completely repress the Nodal inhibitor Dand5. Because this riddle now is solved, there are no experiments left that argue against cilia and flow.
nodal1 and dand6 expression in twins, from Figure 2, Tisler, et al. 2017. Current Biology.
Is the same process is occurring in conjoined humans?
MB & MT We are pretty sure that this mechanism also works in humans: the twins that we can generate in Xenopus are fused at the thorax and have two heads, which is exactly the type of human twins that show this odd laterality defects (and which constitute some 70% of human conjoined twins. Also, the anterior-posterior position of the LRO corresponds to the thorax in the adult. Human twins fused at other sites show mostly normal organ situs, for example if they are just fused at the heads. In these cases, the LROs would be expected to be completely separated during neurulation.
When doing the research, was there a particularly exciting result or eureka moment that has stayed with you?
MT The initial observation of the duplication of the Left-Right Organizer in twins and that this could be the key to explain the century-old question of how the organ situs is determined in conjoined twins will stay with me.
pitx2c expression in control and dand5 MO injected animals, from Figure 3, Tisler, et al. 2017. Current Biology.
What about the flipside: any particular moments of frustration or despair?
MT Some moments of frustration that probably everybody working with frog embryos shares is when the manipulated specimens do not survive the night. Specifically regarding the twin project: the task of visualizing the flow in the archenteron of conjoined twins has had a lot of “potential” to causing frustration and despair.
And what next for you Matthias?
MT Now that my time in Hohenheim is ending (the defense of my thesis has been scheduled), it is time for me to try something new. As a consequence of this work I really got interested in human development and disease, so I decided to switch gears and to go to Med School. In the future I hope to be working as a clinical scientist from bench to bedside and backwards.
And where next for the Blum lab?
MB Off to the next frontier: linking the flow-dependent repression of Dand5 to upstream events. We really need to understand how this repression is brought about in a flow- and cilia-dependent manner. This task has the potential to keep us busy for some time. And most likely we will be using the twin assay again.
 (No Ratings Yet)
(No Ratings Yet)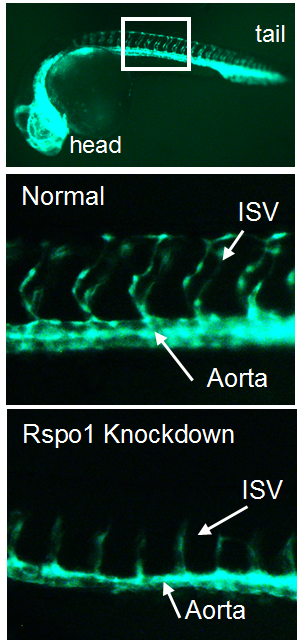
 (4 votes)
(4 votes)
 In the first study (p.
In the first study (p. 


 Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their
Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that regulate diverse processes such as cell proliferation, differentiation, development and regeneration. Several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. In their  In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their
In recent years, lineage tracing studies have identified distinct epithelial stem and progenitor cell populations in the lung. These cells, together with their differentiated progeny, maintain a stable identity during steady state conditions, but can display remarkable lineage plasticity following injury. In their