A post-doctoral position is available in the Franz-Odendaal Bone Development Lab to study the developmental basis of the vertebrate ocular skeleton in a comparative context. Highly motivated and independent individuals with excellent interpersonal skills are encouraged to apply. The successful applicant will take a key role in our research program which spans evo-devo, developmental genetics and phenotypic variation. Opportunities to teach undergraduates, supervise students, and outreach are available to those interested.
We are seeking a recent doctoral student in Biological Sciences or related fields with experience in Molecular Biology, Cell and Developmental Biology. Experience with zebrafish and/or chick embryos is desirable but not required. Opportunities to supervise students will be available.
Please send curriculum vitae and summary of research interests via email.
Dr Tamara Franz-Odendaal
Franz-Odendaal Bone Development Lab
Mount Saint Vincent University,
Halifax, Nova Scotia, Canada
I am looking for talented and driven candidates to apply for a position in a 4yr-PhD programme and join my laboratory at the European Cancer Stem Cell Research Institute (Cardiff University). The studentship is funded by the South-West Doctoral Training Partnership of the BBSRC and starts September 2017.
The project aims at understanding how adult stem cells respond to the local needs for cell replacement through lineage tracing, genetic manipulation, confocal microscopy, mathematical modelling and the development of a new method for the temporal control of transgene expression in Drosophila. This will be done in collaboration with Prof Helen White-Cooper’s lab in Cardiff and with Dr Edward Morrissey in Oxford.
Earlier this week, I took part in a workshop on preprints – organised by Alfonso Martinez-Arias and held in Cambridge, UK. Inspired by the ASAPbio movement in the States, Alfonso felt it would be useful to bring discussion of the potential value of preprints more to the forefront in the UK. Happily, he was able to get John Inglis, co-founder of bioRxiv (the primary preprint server for the life sciences), to speak at this event, and also invited several other speakers – including myself – to talk about their experiences with preprint servers.
If you’re interested, the whole event was live-streamed, and you can watch the video here. But for those (understandably!) not wanting to spend 3 hours listening to people talk about preprints, here’s a (not-so-brief) summary. Or check out #pptsCamOA for the live tweeting.
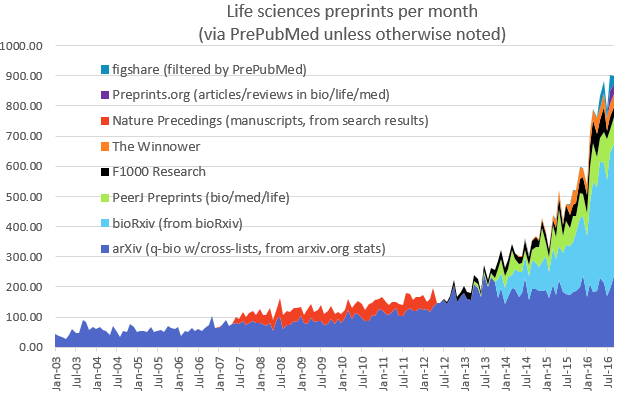
If you’re not familiar with bioRxiv or preprints more generally, I’d definitely encourage you to watch John’s talk at the beginning of the video (starting around 7 minutes) – he gave a great overview of the history of bioRxiv and its potential future. One thing I find interesting is why it appears that bioRxiv is succeeding while previous efforts – most notably Nature Precedings – failed (see graph below). Presumably this is primarily because bioRxiv got its timing right: we’re in a period of changing culture with respect to science publishing, and bioRxiv is riding a wave of discontent with traditional publishing systems. Perhaps Nature Precedings just came too early, but perhaps also people have been more willing to rally behind a not-for-profit enterprise not directly linked with a publishing house (bioRxiv is supported by Cold Spring Harbor Labs not by the Press).
Image from http://asapbio.org/preprint-info/biology-preprints-over-time Credit: Jessica Polka
As you can see from the graph, bioRxiv is expanding rapidly (now with around 450 deposits and over 1 million article accesses per month), and it’s clear to me that preprints are here to stay – though there are still a number of important challenges to address (check out the video from around 32 minutes to see where bioRxiv is going).
That preprints are becoming increasingly mainstream is also reflected in the fact that most journals are now happy to consider papers that have been posted on preprint servers – including Cell Press, who were initially and somewhat notoriously reluctant to make a clear statement on that front. Two representatives from the publishing industry talked about their attitudes to preprints – me (representing Development and The Company of Biologists) and Mark Patterson from eLife. Mark identified three reasons why eLife supports preprints: that ‘journals shouldn’t get in the way’ of what scientists want to do, that ‘journals and preprints complement each other’ and, somewhat enigmatically, that ‘new possibilities arise’. Here, Mark introduced the audience to a concept that most of the audience (myself included!) weren’t familiar with – a preprint-overlay journal. Exemplified by the maths journal Discrete Analysis, the idea here is that the journal is built on top of the preprint server, with the server (in this case arXiv) hosting the content, and the journal providing the peer review element. Later on in the workshop, Aylwyn Scally returned to this idea as his vision for how academic publishing might work in future, and it’s certainly an interesting idea…
Unlike eLife and its somewhat revolutionary ethos, The Company of Biologists is a much older and more traditional publishing enterprise, and it has taken us some time to embrace preprint servers, with each journal changing policy at different times according to feedback from its community – as shown in the excerpt from one of my slides below (I make an appearance at 56 minutes on the video). While it’s only recently that all our journals have supported preprint deposition, we’re now fully on board, and will shortly be fully integrated with bioRxiv for simple bidirectional transfer between journal and preprint server.
J2B portal is the system allowing simple transfer from the journal submission site to bioRxiv for immediate posting; B2J is the reverse portal whereby authors uploading a file to bioRxiv can easily submit to an integrated journal.
The final speaker for the first session was Hannah Hope from the Wellcome Trust. Given that preprints allow you to get your research out to the community earlier, but that that research is not yet peer-reviewed, funding agencies have to decide whether or not to ‘count’ preprints when assessing someone’s productivity for grant or fellowship applications. The Wellcome Trust has taken a forward-looking approach to this and supports the use of preprints in applications and end-of-grant review. They’re also working with other funders to develop a common set of principles – which should be made public soon. This is great news for researchers who need to apply for a grant before their paper is formally published but who want to demonstrate that they have a good track record in their field, and hopefully will become common practice among funders!
In addition to hearing the voices of publishers and funders, Alfonso had invited several preprint users to talk about their experiences. Raymond Goldstein, an interdisciplinary PI whose work spans the physics-biology divide (and includes IgNobel prize winning work on the shape of a ponytail!), gave us the viewpoint of someone for whom posting on arXiv has been common practice for decades and for whom the surrounding angst that many biologists feel is alien. He pointed out that, with LaTeX-based formatting tools, one can generate a beautifully formatted version of the manuscript to submit to a preprint server – common practice in the physical sciences, but with most biologists still married to Microsoft Word, isn’t something we’re yet comfortable with. Raymond also raised an important question about discoverability of preprints and argued that it might make more sense to have a central server to hold everything in one place, rather than multiple outlets for preprints.
Steve Royle told us ‘how and why I became a preprinter’ – explaining that his frustration with the amount of time it takes to get a paper published (see his blog post for some fascinating stats on this – it can be quicker to produce a baby than a paper!) induced him to start depositing preprints. As paraphrased by Richard Sever on twitter, Steve said that ‘preprints bring the joy back during the soul-destroying wait for journal publication’. Aylwyn Scally highlighted his concerns about how the peer review system works (what credit do referees get for the work they put in?) and how he feels preprints can help change the system. Sudhakaran Prabakaran‘s unusual career path – spending time as an editor at AAAS before returning to academia to set up his own lab – has given him a unique viewpoint on academic publishing. His main point was that review takes too long (mainly because reviewers take too long!), and that preprints therefore provide a possibility to get your work out more quickly, and to take advantage of altmetrics to get your work known in the community. Finally, Brian Hendrich took to the stage to explain why he felt compelled to post a preprint to alert readers to potential problems with a published paper (something that Steve Royle also mentioned), and why he then didn’t feel the need to get that preprint formally published elsewhere.
In addition to the talks, there was a lot of discussion surrounding why preprints are useful and where we are going with them. It was great to hear some of the early career scientists express their opinions – mainly but not exclusively positive – about preprints. One expressed concern about putting her work out on a preprint server without the ‘sanity check’ of peer review, while another explained how he felt his preprint helped him get his Henry Dale Fellowship. Discussion also centred around the degree to which preprint posting can claim precedence for an idea or a set of data (though without firm conclusion!), and to what extent peer review can claim to ‘validate’ a scientific paper. In general, I found it a hugely constructive discussion, and I came away feeling positive about how preprints and journals do and can integrate to allow researchers to better disseminate their research.
At the end of my talk, I posed a bunch of questions that I hoped would provoke discussion among the audience, and I’d also be really interested to hear people’s feedback here, so please feel free to comment on any of these below!
When should authors ideally submit to preprint server?
Should editors be searching through preprint servers and soliciting potential submissions?
What do preprints mean for precedent and scooping? Will researchers claim precedence through deposition of very preliminary data?
How might this impact on e.g. data/materials sharing or patent applications?
Is there a danger of making un-reviewed data publicly available – especially medically relevant/translational research?
Might commenting on preprints complement or even replace traditional peer review and what would it take to make this happen?
Is it good that we have one dominant preprint server for the life sciences or should there be several?
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute, University of Cambridge
Salary: £28,982-£37,768
Reference: PS10005
Closing date: 03 November 2016
Post-doctoral fellow: healthy ageing of human haematopoietic stem cells.
Fixed-term: The funds for this post are available until 31 October 2019 in the first instance.
The Wellcome Trust-Medical Research Council Cambridge Stem Cell Institute is an international centre of excellence for stem cell research and regenerative medicine. Scientists in the Institute collaborate to advance our knowledge of various stem cell types and to perform pioneering work in translational research areas, providing the foundation for new medical treatments. The Institute currently comprises 29 research groups based across 6 sites in Cambridge. In 2018 all researchers will move to a new building on the Cambridge Biomedical Campus.
Applications are invited for a research associate to join Dr Laurenti’s group. We combine state-of-the-art experimental and computational methods to study the unique biological and molecular properties of human Haematopoietic Stem Cells (HSCs).
The post holder will join a new multidisciplinary project funded by the BBSRC. This collaboration between Dr Laurenti and Prof. Göttgens laboratories has as principal aim the investigation of the functional and molecular heterogeneity of HSCs throughout a human lifetime, with a particular focus on the effects of healthy ageing. The project will combine single cell, transcriptomics, epigenomics, flow cytometry, single cell functional assays in vitro and in vivo.
The successful candidate is expected to creatively and independently carry out their own research project, while collaborating on a regular basis with a team of experimentalists and computational biologists. They will also effectively communicate their work in writing and oral presentations at internal meetings and international conferences.
Candidates should hold a PhD in a relevant field. In particular, they will have a strong background either in stem cell biology, haematology, immunology or ageing. Extensive experience with flow cytometry, mouse models and tissue culture is required. The candidate should also possess molecular biology skills. Additional expertise with high-throughput sequencing data and the R programming language would be desirable. Finally, they will have demonstrated scientific achievement with an excellent publication record.
Once an offer of employment has been accepted, the successful candidate will be required to undergo a health assessment and a security check.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/11327. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
The closing date for all applications is the Thursday 03 November 2016.
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute, University of Cambridge
Reference: PS10545
Closing date: 05 January 2017
Studentships starting October 2017
Stem cells are defined by the dual capacity to self-renew and to differentiate. These properties sustain homeostatic cell turnover in adult tissues and enable repair and regeneration throughout the lifetime of the organism. In contrast, pluripotent stem cells are generated in the laboratory from early embryos or by molecular reprogramming. They have the capacity to make any somatic cell type, including tissue stem cells.
Stem cell biology aims to identify and characterise which cells are true stem cells, and to elucidate the physiological, cellular and molecular mechanisms that govern self-renewal, fate specification and differentiation. This research should provide new foundations for biomedical discovery, biotechnological and biopharmaceutical exploitation, and clinical applications in regenerative medicine.
Cambridge Stem Cell Community
The University of Cambridge is exceptional in the depth and diversity of its research in Stem Cell Biology, and has a dynamic and interactive research community that is ranked amongst the foremost in the world. By bringing together members of both the Schools of Biology and Medicine, this four year PhD programme will enable you to take advantage of the strength and breadth of stem cell research available in Cambridge. Choose from over 50 participating host laboratories using a range of experimental approaches and organisms.
Programme Outline
During the first year students will:
• Perform laboratory rotations in three different participating groups working on both basic and translational stem cell biology.
• Study fundamental aspects of Stem Cell Biology through a series of teaching modules led by leaders in the field.
• Learn a variety of techniques, such as advanced imaging, flow cytometry, and management of complex data sets.
Students are expected to choose a laboratory for their thesis research by June 2018, and will then write a research proposal to be assessed for the MRes Degree in Stem Cell Biology. This assessment will also be used to determine whether students continue on to a 3-year PhD.
Application Process
Visit http://www.stemcells.cam.ac.uk/study/wtprogramme/ for further details including how to apply. Please note: you will be required to complete and submit a departmental application form, provide two references and upload copies of your transcripts as part of the application process.
We invite applications from both EU and Non-EU applicants*.
Application Deadline: Thursday 5th January 2017 Interviews will be held on: 23rd, 24th and 25th January 2017
*Non-EU applicants, who need to apply for additional funding (which covers tuition fees at the ‘overseas’ rate), must also apply through the University Graduate Student Application Form ‘Applicant Portal’ by Wednesday 7th December 2016.
The University values diversity and is committed to equality of opportunity.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
We are looking for highly motivated and talented candidates for a 4yr-PhD programme in the laboratory of Dr Toki Takahashi at the University of Manchester. This studentship is funded by the BBSRC DTP and will start in September 2017. The overall aim of the project is to understand how our head is formed by combining a laboratory experimental approach with high-throughput bioinformatics analysis. This project is in collaboration with Dr Shane Herbert and Professor Magnus Rattray.
The vertebrate craniofacial development is a remarkably dynamic and intricate process, requiring the tightly regulated growth, migration and patterning of a particular cell population, cranial neural crest cells (CNC). For this reason, craniofacial malformations are among the most common, accounting for one third of human congenital defects. Whilst the causes of most cases are unknown, several clinical studies reported the mutations within Alx homeobox genes in recessive hereditary cases (ALX-related frontonasal dysplasia; ALX-FND). Our lab reported a new function of alx1 gene in frontonasal CNC migration, and demonstrated that alx1 is critical to zebrafish craniofacial development.
In this project, we will first establish a zebrafish model of ALX-FND by generating alx1 null mutant line with the CRISPR/Cas9 system. Using the real-time image analysis of this mutant, we will examine in details how alx1 controls the CNC migration. Furthermore, we will purify the migrating CNC from the mutant by FACS for deep sequencing of transcriptomes. Bioinformatics analysis from this targeted cell population will identify the downstream targets of alx1. By integrating the high-throughput data and the live image profiling of CNC migration into computer modelling, the project will elucidate the new alx1 gene regulatory network controlling vertebrate head development.
The Laboratory of Zebrafish Developmental Genomics headed by Cecilia L. Winata was established in partnership with the Max Planck Institute for Heart and Lung Research (MPI-HLR). The focus of our research is to elucidate the gene regulatory networks and epigenetic mechanisms regulating various aspects of developmental biology as the initial step towards understanding human genetic diseases. Our research employs state of the art methodology based on Next-Generation Sequencing as well as CRISPR/Cas9 genome editing technology.
We invite application from motivated and talented students to perform the research part of their Master thesis in the study of heart pacemaker development using zebrafish as a model. The project titled “Genomics dissection of the heart pacemaker in Zebrafish” will be funded by the First-TEAM grant from the Foundation for Polish Science (FNP) starting from January 2017 for 18 months. The First Team program of the Foundation for Polish Science co-financed from the European Union under the European Regional Development Fund.
The ideal candidate should be able to communicate in English language and possess good communication skills. A good understanding of any of the following subjects is essential: molecular biology, biochemistry, and/or genetics principles. Prior experience in lab work is highly desired.
We offer stipend in the range of 1500 – 2500 PLN/month depending on the candidates’ qualifications and time commitment, and the chance to work in a team of young and friendly scientists with interests in developmental biology, regulatory genomics, epigenetics, and biochemistry. The successful candidate will have the opportunity to work with the zebrafish model animal and learn and apply state of the art microscopy techniques on in vivo imaging of the heart, as well as other methods in zebrafish developmental biology.
Please forward your complete application including a CV, a motivation letter, university scores, and the contact of at least one referee to rekrutacjazdg[at]iimcb.gov.pl with the subject line “First-TEAM” before the 30th of April 2017. Thanking all applicants for their interest, only selected candidates will be invited for an interview.
Please include in your application the following statement: “In accordance with the personal data protection act from the 29th of August 1997, I hereby agree to process and to store my personal data by the Institution for recruitment purposes”.
Sanford Burnham Prebys Medical Discovery Institute, La Jolla, California
An NIH-funded postdoctoral position is available to investigate the signaling functions of retinoic acid (RA) during mouse embryo development. Our laboratory has reviewed recent advances in this field: Cunningham, T.J. and Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nature Reviews Molecular Cell Biology 16: 110-123 (2015).
We are searching for a Postdoctoral Associate to explore the mechanisms of RA signaling during the early stages of organogenesis and limb formation using CRISPR/Cas9 gene editing, RNA-seq transcriptome analysis, and embryo chromatin immunoprecipitation.
Candidates should have a strong background in developmental biology. Salary $47,476 with full benefits.
Interested applicants should email their CV and names of three references to:
Gregg Duester, Ph.D.
Professor
Development, Aging, and Regeneration Program
Sanford Burnham Prebys Medical Discovery Institute
So far in this series, we’ve featured fly nuclear pores,lizard tails, squid eyes and mouse digits, and heard from researchers working in Germany, the US and Canada. Today, we switch model system and geographical location once again. The work was published recently in Development, and uses timed inhibition of sonic hedgehog signalling during chick wing development to test models of digit patterning. The people behind the paper are its two authors: postdoc Joseph Pickering and PI Matthew Towers, MRC Career Development Fellow at The University of Sheffield in the UK.
Matthew Towers (L) and Joseph Pickering (R, representing avian developmental biology in fancy dress)
So Matt, can you tell us how you came to form your lab, and what questions your team is aiming to answer?
MT From my days as a PhD student working on the functions of cell cycle genes during snapdragon leaf development, I have been interested in how growth and pattern specification are integrated. As a postdoctoral researcher I was fortunate enough to continue working on this problem – but on chick limb development – under the supervision of Cheryll Tickle, who has contributed greatly to this field. I set up my own lab studying limb development at Sheffield, first with the support of the university and my sponsor Marysia Placzek, and then with an MRC Career Development Fellowship. I recently obtained a Wellcome Trust Senior Research Fellowship.
“The key question my lab is addressing is how cells integrate their intrinsic developmental programmes with extrinsic signals during limb patterning.”
The key question my lab is addressing is how cells integrate their intrinsic developmental programmes with extrinsic signals during limb patterning. This work has shed light on some of the classic, but controversial models of limb development – in particular, how pattern is specified along the antero-posterior (thumb-little finger) axis, and, in collaboration with Marian Ros’ lab in Santander, the proximo-distal (shoulder-digit) axis. I also co-supervise two PhD students with Marysia who are studying how growth and pattern specification are integrated during the early development of the chick hypothalamus.
Sheffield seems like an exciting place for life sciences and developmental biology at the moment?
MT The Biomedical Science department is an exciting place to study the life sciences. We also have strong links with a number of excellent research centres that focus on a range of subjects including stem cell biology, sensory neuroscience and membrane biology. I am part of a very dynamic group of researchers in the Bateson Centre who use a variety of model organisms, including zebrafish, chickens, fruit flies, slime moulds and mice to investigate a range of questions in developmental biology. These include trying to understand the basic patterning process, to elucidating how pathogens subvert the normal development of an individual.
And Joseph, how did you come to join Matthew’s lab?
JP I completed my PhD with Anne-Gaelle Borycki in Sheffield studying the regulation of an extracellular matrix protein, Laminin α1, by Sonic hedgehog (Shh) signalling in zebrafish. In fact, my paper on this has just been published.
Matt started at the university just as I was writing up my thesis. I found his work very interesting, but he was not yet ready to hire a postdoc! I left the university for a role as a scientific sales rep, which gave me a good grasp of the kinds of developmental biology research going on around the country. I kept up to date with his research activities through discussions in the pub, and in 2014, I decided to take up a postdoc position in Matt’s lab. I had developed an interest in Shh signalling and patterning mechanisms since my days as an undergraduate student, and the chick limb seemed like the ideal model to study this further.
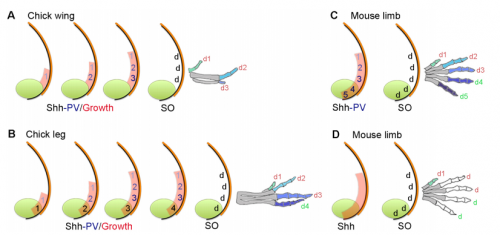
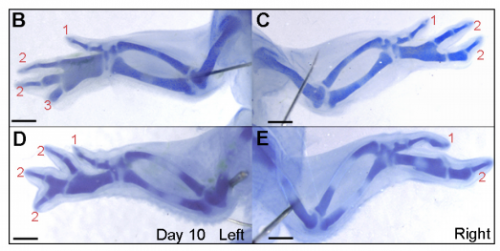
Models for digit patterning in different limbs of chick and mouse, from Figure 1 in the paper.
Your paper uses chicken wing development to test models of vertebrate digit patterning. What was the state of play before you started this work, particularly with regard to ideas of positional information and self-organisation?
MT&JP Positional information can explain how different digit identities (thumb vs. little finger) are specified in the chick wing. However, it has been apparent for a long time that digit number and spacing is determined by a self-organising mechanism that can be modelled by a Turing-like process. Before we started this work it was unclear how these two processes could act together to generate some of the diverse digit patterns found in nature. Thus positional information and self-organisation were often viewed as competing models of digit patterning.
“Positional information and self-organisation were often viewed as competing models of digit patterning.”
Could you talk us through the basic experimental rationale behind the paper?
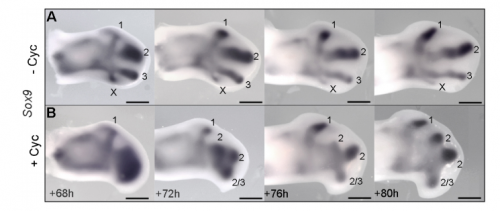
JP: I was investigating another problem using the drug cyclopamine to inhibit the Shh pathway in the chick wing bud. Unexpectedly, I noticed that at a specific stage of development, the cyclopamine-treated wings produced an extra digit. Matt had occasionally seen this phenotype when he was a Postdoc in Cheryll Tickle’s lab (MT – it gave me a headache because it did not seem to make any sense) and we talked about how similar it looked to patterns of digits produced by self-organisation in the absence of positional information. The study of self-organisation in the limb has had a renewed interest, especially in recent papers from James Sharpe and Marian Ros. Therefore, since it looked like we had transformed a chick wing pattern requiring positional information, to one not requiring it, I decided to study it further. I was able to routinely obtain the phenotype and this allowed us to characterise how the temporal interplay between positional information and self-organisation can produce diverse digit patterns.
Sox9 expression in untreated and treated wings, from Figure 4 in the paper
Could you sum up the key results of your paper in a paragraph?
MT&JP Previously, as a postdoc I had shown that Shh signalling integrates antero-posterior growth with the specification of cells with three antero-posterior positional values that give rise to the three chick wing digits (1-2-3). In our current Development paper we showed that Shh inhibition during a very precise point during this process could unexpectedly result in wings forming with four digits (1-2-2-2, based on the pattern of phalanges). We showed that this occurred because the specification of antero-posterior positional information is truncated. However, a switch to antero-posterior growth mediated by the overlying epithelium then occurs. This expansion allows for cells specified with a digit 2 positional value to give rise to up to three digit 2s by self-organisation. Interestingly, one of these digits 2s unexpectedly arises from the Shh-producing cells of the polarizing region – an ability lost in the dinosaurian ancestors of birds.
And to expand on this theme, what does this work suggest about the evolutionary history of digit patterning?
MT&JP Previously, we had presented a model for how positional information specifies the four different positional values of the chick leg digits (1-2-3-4). As the chick leg has remained unchanged during the evolution of amniotes in terms of phalangeal pattern, we suggest that it represents the patterning mechanism for four of the five digits of the common ancestor of birds and mammals. We speculate that the ancestral mechanism has been maintained in the bird wing, apart from the loss of the most-posterior digits that arise from the polarizing region (digits 4 and 5). However, in light of our new findings, we propose that the specification of positional information has been curtailed in the mammalian limb, but that, based on phalangeal pattern, self-organisation then results in the formation of several digit 2s (1-2-2-2-2).
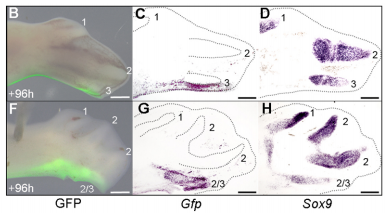
The GFP grafting experiment, from Figure 5 in the paper
When doing the research, was there a particularly exciting result or eureka moment that stayed with you?
JP Successfully grafting a polarising region from a GFP-expressing chicken into a wild-type chicken felt like quite an achievement by itself, but did not come close to the excitement of discovering that the graft contributes to the fourth digit of cyclopamine-treated wings! This is the first piece of work to clearly show that a digit can arise from the polarizing region of the chicken wing bud, and gave important insights into how mammalian digits may be patterned. We were also surprised because we had expected an additional central digit to arise in the cyclopamine-treated wings, not an extra posterior digit. I also once managed to juggle 3 eggs for at least 10 seconds.
And what about the flipside: any particular moments or frustration and despair?
JP Performing the experimental work in this paper was very repetitive and laborious due to the sheer number of embryos that would die. This was particularly frustrating when it involved difficult experiments such as tissue grafting.
Removal of the apical ectodermal ridge after cyclopamine treatment, from Figure 7 in the paper
So what next for you, Joseph?
JP More chicks and drugs and rock ‘n’ roll! I’m currently trying to understand how the four-digit phenotype that I have characterised occurs at the molecular level. I am testing various candidate genes for their role in digit formation from the polarising region using chicken and mouse embryos, and also validating an RNA-sequencing screen that I have performed on cells of the developing chick limb.
More chicks and drugs and rock ‘n’ roll!
And what is the Towers lab currently working on?
MT Our recent research has highlighted the importance of cells measuring time to intrinsically execute their developmental programmes (Chinnaiya et al, Nat Comm 2014, Saiz-Lopez et al 2015). Now we are particularly interested in the molecular nature of intrinsic timing mechanisms, and how they could be used to scale pattern formation both within, and between different species.
29 March – 11 April 2017 Cold Spring Harbor Lab Xenopus course:
Xenopus is remarkable for modeling human diseases including birth defects, cancer, and stem cell biology. Xenopus has and continues to make a major impact in our understanding of cell and developmental biology.
Students are encouraged to target their own genes of interest using CRISPR technology and then analyze phenotypes using the diverse array of assays available in Xenopus. Specifically, techniques covered include microinjection, and various molecular manipulations including, CRISPR knockouts, morpholino based depletions, transgenics, and mRNA overexpression. In addition, students can combine these techniques with explant and transplant methods to simplify or test tissue level interactions. To visualize subcellular and intercellular activities, we will introduce a variety of imaging methods including time-lapse, fluorescent and confocal microscopy. Additional methods include mRNA in situ hybridization and protein immunohistochemistry as well as basic bioinformatic techniques for gene comparison and functional analysis. Biochemical approaches such as proteomics and mass spectrometry will also be discussed. This course runs in close association with two other courses: Quantitative Imaging and Protein Complexes.
This course is designed for those new to the Xenopus field, as well as for more advanced students who are interested in emerging technologies. We encourage students to bring their own genes of interest and will tailor aspects of the course to enable them to initiate studies on their specific projects. GENEROUS SCHOLARSHIPS AVAILABLE.
 (No Ratings Yet)
(No Ratings Yet)
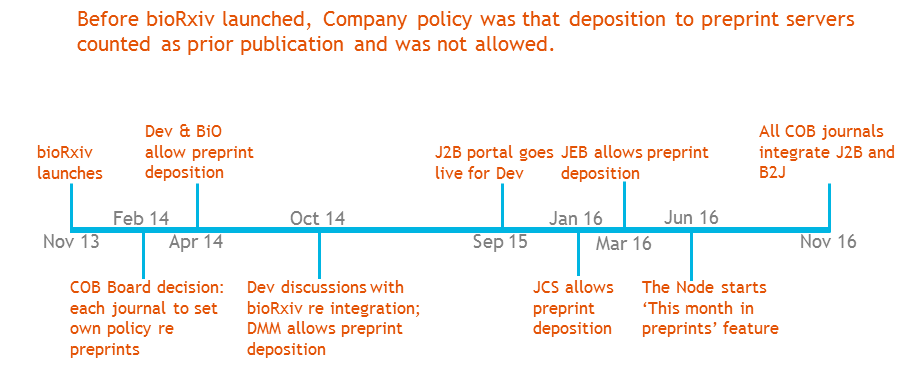 J2B portal is the system allowing simple transfer from the journal submission site to bioRxiv for immediate posting; B2J is the reverse portal whereby authors uploading a file to bioRxiv can easily submit to an integrated journal.
J2B portal is the system allowing simple transfer from the journal submission site to bioRxiv for immediate posting; B2J is the reverse portal whereby authors uploading a file to bioRxiv can easily submit to an integrated journal.  (9 votes)
(9 votes) eostatic cell turnover in adult tissues and enable repair and regeneration throughout the lifetime of the organism. In contrast, pluripotent stem cells are generated in the laboratory from early embryos or by molecular reprogramming. They have the capacity to make any somatic cell type, including tissue stem cells.
eostatic cell turnover in adult tissues and enable repair and regeneration throughout the lifetime of the organism. In contrast, pluripotent stem cells are generated in the laboratory from early embryos or by molecular reprogramming. They have the capacity to make any somatic cell type, including tissue stem cells.