The Biologists @ 100 conference, happening 24-27 March 2025 in Liverpool, UK, will have a dedicated half-day event specifically for early-career researchers (ECRs), in the afternoon of 24 March 2025.
The ECR event will focus on career opportunities both inside and outside of academia. We’re excited to have Richard Sever, the co-founder of bioRxiv and medRxiv, as the keynote speaker at the career session. You’ll also hear from professionals from a diverse spread of career paths to discuss how skills developed during your research career can be applied in a number of different roles.
There will also be food and drinks in the evening for ECRs following the main conference’s welcome reception for those who have attended the workshop.
Biologists @ 100 is a celebration of the 100th birthday of The Company of Biologists, bringing together different communities, including cell, developmental, experimental and disease biologists, and will incorporate the Spring Meetings of the British Society for Cell Biology (BSCB) and the British Society for Developmental Biology (BSDB).
No such thing as a standard career path – an interview with Christos Kyprianou
Christos Kyprianou is a postdoctoral researcher at the Institute of Molecular Biotechnology of Austria. Instead of doing a postdoc right after his PhD in developmental biology, Christos joined The Company of Biologists as FocalPlane’s Community Manager. After setting up and getting the community site for microscopy off the ground, Christos decided to return to academia. What motivated him to switch in and out of academia? And what’s next for him?
Have you always been interested in science?
I was always interested in science. As a kid, I had this picture in my mind that I would be doing something science or medical related. Going through high school, the only thing that stuck with me was biology, and physics to a lesser degree. Then at university, the first lecture I had was on genetics and I really enjoyed it. That’s when I realised I would like to do science in one way or another.
After your PhD, you worked as the Community Manager of FocalPlane. How did this job come about?
A lot of the times, academia leads you to believe there’s one way to do science: bachelor’s, maybe a master’s, PhD, postdoc, then group leader. I remember when I was asked why I wanted to do a PhD during my interviews, I was a bit baffled, because I thought that’s the only way to do science. But now I know you can still do science in many different capacities. As I was going through my PhD, this was becoming more and more obvious to me. Towards the end of my PhD, my PI got a position at Caltech, and I was offered to go to Caltech, which I was interested in. But then the FocalPlane job advert came up, and it made me realise I wanted to try something different, and not just go down the default route of doing a postdoc after a PhD. I applied and got the job.
What was your role at FocalPlane and the Journal of Cell Science (JCS)?
I was involved in setting up FocalPlane and getting it off the ground. I was putting together the content, inviting people, organising events, and participating in events to spread the message. At the beginning, it was unclear to me how was I doing science by building a community and a website. But there were a lot of principles from science that would apply in building FocalPlane. For example, identifying who are the people that are key in driving the microscopy field, and bringing together people who are interested in microscopy but don’t have access to resources, and working as part of a team to create FocalPlane. I was also part of the Journal of Cell Science (JCS) team, so I got to read articles and write Research Highlights, which was a very good exercise in science writing. It’s about taking something quite complicated and summarising it using 200 words. Unfortunately, because of the pandemic, I feel like I missed out on doing more things for JCS, like attending conferences.
After FocalPlane, you went on to do a postdoc at the Wyss Institute at Harvard University. How did that opportunity come about and what influenced that decision?
There were a few factors, but the pandemic played a big part. For me, it changed from being all day in the lab to being at home, as the office was closed. The other thing was when I was commissioning content and talking to people about their research, I started to miss doing the hands-on science myself. And thirdly, the lab I ended up joining as a postdoc was one of the labs that I really aspired to be part of during my PhD. Magda, my PhD supervisor, was meeting that PI and they talked about some of my PhD work, so I took that opportunity to reach out to him. They were working on organ-on-chip technology, which was new and interesting to me. I thought that was the right opportunity for me to go back to doing research. I also wanted to do something translational, because I felt like I missed out on that with my PhD project. Maybe because some people say once you leave academia, nobody is going to want to take you back, I was feeling a bit pressed with time to not miss that postdoc opportunity. But I now know this is not true; I’ve seen people do that, even after longer breaks from academia.
You’re currently a postdoc at the Institute of Molecular Biotechnology of Austria. What motivated the move from the US to Austria and what are you working on now?
My primary career goal right now is to get into industry. It’s a bit less going with the flow and I’m trying to approach my career in a more targeted way, by building up a portfolio and working with projects in the translational and technology areas. Coming back to Europe was also because my family and my wife’s family are in Europe. In terms of the lab, I was looking for something that would help build my portfolio. I’m now in Nicolas Rivron’s lab that is developing stem cell-based embryos for the purposes of using it as a tool to understand health and disease in the context of fertility. It’s a technology that you can build on top of other technologies. I’m not working on a particular biological process; instead, I’m focusing more on learning how I can apply all sorts of questions using these technologies.
Apart from your research, you said you’re trying to build your portfolio in industry. How are you doing that?
The Wyss Institute in Boston, where I did my first postdoc, is a place where academia meets industry. My work there was more industry-adjacent. We were collaborating with startups and more established pharma companies to validate drugs. That was when I decided I wanted to build my portfolio towards industry. After moving to Austria, I wanted to get more exposure to what industry is like here, so I joined the initiative ‘Vienna Bio-Centre Industry Insights’. I attended a couple of seminars, and I was impressed by the quality of speakers, from names like BioNTech, Merck, and a few consulting companies. The initiative was looking for more co-leads, so I applied for the position. I think my experience at FocalPlane really helped with me being selected. Being a co-lead is a good way for me to meet people within and outside of the Vienna Bio-Center and understand the industrial ecosystem in Vienna and the wider German speaking area of Europe. We’re trying to come up with different types of events to give people an idea of what it is like to work in industry and provide networking opportunities with people in industry.
How has your experience at FocalPlane help you in your subsequent roles?
People talk about the importance of having soft skills as a scientist. It’s like a muscle I got to practise during my time at FocalPlane, such as bring people together, organising events, and knowing who to talk to. My experience at FocalPlane and The Company of Biologists helped give me more credibility in these skills. What FocalPlane has become now is beautiful. I really enjoy the variety of its content and how popular it’s become. I even met someone random at the Max Perutz Labs, who mentioned they were planning to write something for FocalPlane. It’s great that people in the field know about it and interact with it.
Looking back your career path so far, would you have done anything differently?
I would definitely have done a bit more extracurricular activities during my PhD. I think I was overly focused on doing my project and not being as involved with other things. I would like to have been more aware of what other opportunities are available and how else I could do science, and not just necessarily going down the academia path. Nowadays there are so many opportunities to try out and learn about different things, like workshops and recruitment events from consulting and biotech companies. These are good ways to get to know people with those careers and get outside of your PhD bubble.
Christos reflects on what he would have done differently during his PhD.
Do you have any advice to someone thinking of switching career paths?
I think it’s a lot about informing and preparing yourself beforehand. If you’re thinking about making a career transition, it’s more about finding out exactly what that career is. Talk to as many relevant people as possible, and really find out if that’s for you. In many jobs, I feel like you can get trained and learn the specifics of it while on the job. For my role at FocalPlane, I didn’t do much science communication before, and it’s not like I had to build another community before. I was just another disgruntled microscope user who wanted to do more with his samples, but there weren’t resources out there easily accessible to people like me. But that was what Sharon, JCS’s Executive Editor at that time, said FocalPlane needed – someone who understood the target audience and their needs. But the rest, like organising events, commissioning content and talking to people, that’s all the stuff I learned on the job.
I’ve been seeing more opportunities for people to get into industry without a PhD. You mostly get trained on the job, and there are particular ways of doing things, so you don’t necessarily need a PhD. I think it’s important that undergrads and master students know about these opportunities before going down the PhD path. Equally I’ve seen people with years of postdoc experience who managed to get into industry.
Do you think you’ll stay in academia? Are there any other career paths you’ve considered?
I’ve been in academia for quite a long time, and I don’t think it’s for me in the long run. I’m exploring different options outside of bench work that allow me to serve science in a different capacity. Business development is something I’m interested in. I’ve been talking to friends who have done this switch and seeing what it’s like. There’s no one particular path of doing science, and it really does come in many different ways. Anything that contributes to science is valid, and everybody should feel comfortable exploring the different options.
Christos encourages people to explore different options.
Finally, what do you like to do in your spare time?
Because we just recently moved to Austria, we’re still trying to get into a routine. I’m just starting to get back to things like playing squash and going on hikes. A more recent hobby is 3D printing. I have my own entry level 3D printer that I’ve been modifying. I try to design and print my own things, like a coffee funnel so I don’t make a mess when I put coffee grounds into my Moka pot. I’ve been more focused on modifying the printer, so it performs better. I think that happens to a lot of people – they get sidetracked by building the printer rather than the actual printing. You can get a pretty cheap printer to start with, because the idea is if you get a bad printer, almost certainly every single part is going to break, so you get to learn what each part does as you’re fixing it. Pretty much like I think a lot of us are figuring out our careers as we go!
Our lab studies the emergence of collective cell behavior and its connection to biological functions in development and reproduction. We focus on the regulation of cellular mechano-chemical coupling in mammalian tissues, using mouse models and cell lines. By integrating live cell imaging with mathematical modeling, we aim to uncover the design principles governing multicellular dynamic systems.
Specifically, through the use of FRET cell imaging to detect kinase activity, we have demonstrated that extracellular signal-regulated kinase (ERK) activity is mechano-sensitive and generates active cellular forces. These forces are fundamental to collective epithelial migration and tissue morphogenesis in developing murine organs, such as the lung and cochlear duct. We will further investigate this regulatory mechanism by integrating tissue geometry and topology to gain insights into not only morphogenesis but also tissue homeostasis and repair.
Another area of interest in our research relates to reproduction. Despite its physiological importance, the origin and role of germ cell dynamics in the reproductive tract remain poorly understood. To address this, we have employed intravital microscopy to study murine sperm and reproductive tracts, allowing us to observe cellular and molecular activities in living tissues. In particular, we aim to explore the biophysical mechanisms underlying collective sperm dynamics and its relationship to reproductive functions.
Lab roll call
Sound Wai Phow: I am a research assistant and the lab safety officer of the lab. My duties include lab logistics and maintaining the mice colonies used by our lab for experiments. I am currently working on a project investigating tissue fluidity and migration.
Veronica Lee Xi Min: I am a PhD student and I am investigating the effects of sperm flow on epithelial cells in a male reproductive organ known as the epididymis. For my study, I am using an in vitro microfluidic system integrated with live cell imaging!
Ankita Nayak: I am a PhD student in the lab. I work on unravelling the impact of substrate curvature on the extrusion of DNA-damaged cells from a tissue-like environment.
Nicole Ng Shu Ying: I am a research assistant and am mostly in charge of the lab’s finances and general maintenance, with a current project involving characterising the murine epididymis using various tissue clearing and imaging methods.
Thanh Phuong Nguyen: I am a research fellow in the lab. I investigate the causative relationship between mechanobiology of epididymal epithelium and sperm development.
Emmanuel Vikran S/O Puspa Nathan: I am an Undergraduate FYP student working on the interaction of aberrant cells and healthy cells with regard to ERK wave propagation.
Chen Xingyu: I am a master student doing my capstone project. And I’m learning and exploring the structure of the epididymis.
Cao Gefei: I am a new PhD student looking into a potential project investigating the dynamics of transzonal projections and ERK signaling within follicles under the context of aging.
Lee Yuan Zhe: I’m a Year 0 PhD student, hoping to look into how collective sperm motion emerges consistently, despite the heterogeneity in environmental and individual sperm cell parameters.
Favourite technique, and why?
My favorite technique is using light microscopy on cells and tissues—simply because it’s fun! I also enjoy quantifying the data and extracting information to build mathematical models. For me, creating a smooth workflow from live imaging to modeling is the ideal approach. It’s so satisfying to turn what we see under the microscope into something that can predict or explain what’s happening.
Apart from your own research, what are you most excited about in developmental and stem cell biology?
I am excited about how complex biological forms—both in shape and size (morphology) and how they’re connected (topology)—self-organize and how they are linked with physiology. Exploring the mechanisms behind these is incredibly exciting!
PI: How do you approach managing your group and all the different tasks required in your job?
We use Slack for daily communication, hold weekly lab meetings to discuss research progress, and have one-on-one meetings for project updates. While there is no strict management style, responding promptly to questions and concerns from lab members is my top priority. Since our lab is young, I make it a point to take initiative and personally demonstrate anything new, like setting up equipment or conducting experiments. My motto is: think, feel, and move!
PI and others from the lab: What is the best thing about where you work?
Tsuyoshi: My colleagues at the MBI have different background in broad disciplines, which is nice to receive great stimuli for a new idea.
Sound: The people and the facilities. Everyone (even from other labs and cores) is very friendly and willing to teach/share their expertise and knowledge without hesitation. The facilities available makes this conducive environment very easily achievable.
Veronica: Definitely the people. We are all very different in terms of work styles and personalities, yet we blend well together. The fact that almost all of us sit together also makes it easier to discuss our work, and nurture bonds beyond regular coworkership. Other than our abundance of microscopes (we have three just for our lab!), I also love how we each have our own comfortable work space. If we could give a lab tour, you would see that each member has some sort of plushie/ figurine/ ornament on our tables that truly reflects our unique personality (e.g. I have two alpaca plushies mounted on my monitor screen).
Ankita: The open-lab system allows us to interact with people from all labs at MBI. It makes for a stimulating environment with people from different backgrounds who are always willing to lend a hand. Apart from the open-lab system and core facilities which helped us a lot while setting up a new lab, I love how organised we are here. Our lab’s databases are easily accessible to all members, and up to date, which helps me plan my experiments easily. The institute also organises informational seminars and conducts training sessions for the equipment which enables us try new avenues in our work.
Nicole: The people here are all very willing to help each other out, even if we are not in the same lab team. Support teams that we call core facilities are a blessing. From IT support to daily lab amenities, as well as usage of a plethora of devices available to everybody is always well coordinated and efficient. The support teams are very willing to conduct training sessions for any device that one may want to try using, so I learnt a lot about various microscopes available in the institution in such a short period of time which is amazing.
Thanh: The setting of open-lab creates a cross-link communication between researchers across different lab with different background. Lots of new ideas and approaches come during those across-the-lab scientific discussion. Facilities of supporting cores play substantial roles in ensure that research works going smoothly and with high efficiency.
Emmanuel: The diversity of people and good facilities. Discussing plans and data with people around me within and beyond my lab is enriching, where I can receive ideas that propel success in approaching research.
Xingyu: THE ATMOSPHERE! I can feel the harmonious and positive energy working with lab members . Everyone is keen to share and help. Also working in our lab is a great opportunity to try these advanced technologies. All the advice I’ve got and accessibility to the lab will help me figure out how scientific research is carried out, and build my own academic research map.
Yuan Zhe: The fact that people don’t take things seriously. On a clownish scale from 0 to 10, I rate myself 11. So, it’s great to be at a place where I can be free to blurt out any nonsensical thoughts I have all the time.
PI and others from the lab: What’s there to do outside of the lab?
Tsuyoshi: Singapore has many parks and green spaces. Watching various birds, like hornbills, kingfishers, and even chickens, is relaxing. Running in the parks is also wonderful—you might even come across huge lizards and snakes along the way.
Sound: Weekly inter-lab dinner and games night.
Veronica: I can’t boast enough about our game night! While other labs tend to meet up over dinner, we host both dinner AND games. Members of other labs are also welcome since our lab is quite small. Recently, each of us have also delved into various sports in an attempt to keep fit/become fitter. From what I know: Tsuyoshi climbs the stairs often, Thanh runs, Sound and Ankita gym, Nicole and I swim, Gefei does all three (run, gym, swim), and Yuan Zhe probably did some sort of sport that led to his spinal cord injury (get well soon!).
Ankita: Singapore may be a small country, but it boasts remarkable cultural diversity. I love exploring foods from different traditions, all conveniently accessible through the excellent public transport system. The city is also very walkable and features impressive green spaces. Plus, with Singapore’s easy access to other parts of Southeast Asia, travelling around the region is a real pleasure!
Nicole: There is a huge variety of food from a vast array of cultures in Singapore so it is always cool to visit new food places all around the country. New cafes, eateries and old school shops pop up once in a while and it’s always an interesting experience to try them out.
Thanh: Singapore is a modern city and therefore having a lots of way to have fun outside of the working time. People are colleagues during working hours and switch to friends after work for hanging out and explore Singpore’s unique food and culture.
Emmanuel: Singapore, being well-lit and even more beautiful at night, makes nighttime runs exciting. The island features stunning paths that stretch for several kilometers, allowing uninterrupted runs without the need to stop for traffic.
Xingyu: I like the small talks and lunch with people. I’m quite shy now but I’ll try to engage more in the future!
Yuan Zhe: I enjoy vertical marathons and late-night cycling. Singapore has enough skyscrapers for the former, and enough large green parks for the latter. When the crowd is sparse late at night, the parks are pretty much Autobahn for cycling.
No such thing as a standard career path – an interview with Maria Rostovskaya
Maria Rostovskaya is a senior research scientist at the Babraham Institute, Cambridge, UK, studying development using human pluripotent stem cells. Between her undergraduate degree and her PhD, Maria was a dancer and dance teacher for a few years. What made her decide to switch careers and eventually follow an academic career path? How did her experience in dancing shape her subsequent career? We chatted to Maria to find out more.
How has your career path been so far?
I did my undergrad in molecular biology at the Moscow State University. After university, I decided to take a break from the academic career. I became a dance teacher. I also did some lab work during that time, but I did not think of really continuing a career in academia. Then, three years later, I decided to go back to academia and pursue a PhD. I was accepted to the PhD programme at the Max Planck Institute in Dresden. I had two main directions during my PhD – my main project was about bone marrow stem cells and my side project was on human pluripotent stem cells. After my PhD, I continued with a postdoc at the Cambridge Stem Cell Institute. Now, I’m a senior scientist at the Babraham Institute at Cambridge, working on human pluripotent stem cells in the context of human development.
After your university degree, you decided to become a dancer and dance teacher. Could you talk us through that decision-making process?
I was curious about many things, and I had always done dancing and science in parallel. During my time at the university, I focused a lot on my studies, so after that, I wanted to expand my other interests. Another reason is, at that time, I was not ready for the transition from studying to doing research. My undergraduate degree involved a lot of theory, and I enjoyed learning about concepts and principles. I think people don’t always realize the difference between learning the theory and doing the research: there is a huge difference in terms of the intellectual load. How we learn from a day spent with textbooks is very different from how we learn from a day working in a lab, and I just wasn’t ready for this difference in dynamics.
How was your experience as a dancer and a dance teacher? What type of dance did you specialize in?
The style of dance is called the Hustle dance. It’s a social dance, and it’s a fusion of salsa, mambo, and west coast swing. It’s a partner dance and can be danced with different music. There is no prepared scheme. It’s a lot about leading, following and improvising with the music. Its uniqueness comes from its freedom and flexibility. You can add various styles and elements. You can style it according to your own feelings.
My dance partner and I also created a unique dance style and fusion. We also designed a teaching course, and we were invited to run workshops in other cities and even abroad. We won several competitions. It was also rewarding to see the achievements and results of my students, because they also won competitions. Some of them went on to become dance teachers. It was a really exciting time, because I had the space for creativity and the opportunity to try various things.
You were enjoying dancing, but then you decided to pursue a PhD. What motivated that decision?
I think I had the feeling that I achieved a lot in dancing, and I thought that I needed to develop further. By that time, I have gained more self-awareness, and I understood what I actually need. For me, it’s important to have opportunities for further development, for professional growth. I like to have a sense of purpose and be intellectually stimulated. With this new-found awareness, I realized that probably I could try going back to academia and pursue a PhD.
Maria reflects on why she decided to switch from dancing to doing a PhD.
How did you approach applying for a PhD and choosing which area you wanted to work in?
At that time, I wasn’t entirely sure that I wanted to go back to academia. I wanted to try, so I chose one PhD programme and applied. I thought if I did not get it, I would just continue with dancing. But I did get it, so I decided to change my career path. It was a real match – it was the right time, the right place, and the right project. I was always excited about molecular biology and cells, and it was a really the right combination of topics for the project.
Was it difficult going back to science after a break?
At that stage, not really, because I was still at the very beginning of a research career. I did not feel very different from everyone else. But because I had already tried another career, I knew a PhD was what I wanted. I think that made the difference for me.
After your PhD, you decided to stay in academia and went to Cambridge to do a postdoc. You’re currently a senior research scientist at the Babraham Institute, UK. What is your research about?
When I came to Cambridge, I was first at the Cambridge Stem Cell Institute. Then I made the transition to the Babraham Institute. Now, I use human pluripotent stem cells to model early human development. I study the transcription and epigenetic control of the first lineage decisions. I’m interested in understanding how cells become different at the right time and the right place, and what molecular mechanisms control these decisions. A few years ago, I decided that I also need to add computational skills to my portfolio. There were a lot of big data in my project, and I always had to rely on other people to do the analysis, so I decided to learn bioinformatics. Now in my work, I combine wet lab and computational approaches.
Did being trained as a dancer and teaching dancing help your scientific career? Do you see any parallels in your dancing and scientific experiences?
I think it really shaped my approach to teaching. I do a lot of teaching, and I’ve been teaching for the university for 10 years already. I supervise undergraduate students, and I find a lot of similarities in that respect. I like finding a clear way to explain and structure the knowledge, training skills, identifying key points and filling gaps. I enjoy interacting with my students and I’m very excited seeing them progress. There is definitely a parallel there. Dancing and doing research are also about having a space for creativity, trying out new things, and not being afraid of going to the unknown. Both require being open to something unexpected.
Do you still dance and teach dancing?
I don’t teach dancing anymore, because when I moved to Germany, I had to focus on my PhD. I did not speak German, and I needed to settle in a different country. But I do dance a lot and go to socials events. I often go to festivals or boot camps in different countries. I try to be a part of the dancing community. With the social dance, you don’t need to have a specific partner. You just go and start dancing with any partner.
What’s next for you? Are you planning to continue in academia?
I had another decision point in my career two and a half years ago: my PI left the institute to join a biotech company. By that time, I was already a very senior postdoc and very independent in my research. I knew that I wanted to continue in academia, so I stayed at the Babraham. Throughout my postdoctoral years, I have managed to create my own research niche and approach. I work very independently now, and occasionally I supervise research assistants and summer students. I’m planning to continue the same line of research when I start my own independent group, and I’d like to keep teaching as a part of my career too.
Anything you’ve learned from switching career paths? Looking back, would you have done anything differently?
I certainly learned a lot from this switch. I was not just following the flow of doing a PhD after a science undergraduate degree. I took a break, and I think at that time, that was the right decision at that period of my life. I do not regret at all, not even a second. I don’t know how it would have been otherwise, but if I did not make that decision to take that break and do something else, I wouldn’t have been here now. I gained self-awareness, and it made me value what I have now. Pursuing career in academia is a conscious choice – I have a dream, and I want to follow this dream, and it is worth it. My career switches made me confident that right now, this is the right place for me. But I am open to changes in the future; we don’t know where life brings us.
Maria talks about not regretting switching career paths.
Do you have any advice to someone thinking of switching career paths?
I would say, have the courage to try. If you feel like you want to try different activities, just give it a go. You will never know unless you try, and perhaps it’s better not to regret about not trying.
You’ve always had dancing and science in your life. But have you ever thought about other career paths?
Maybe a dog walker… I’m just joking. I love dogs, but with an academic career, it’s really hard to have a dog. I already have dancing and science. I think two is a good number!
How do you find time for dancing, despite your busy work schedule?
I think it’s really important to have physical and socializing activities in your life. I think this is why I try to keep dancing in my life as much as I can. It’s about prioritising, organising yourself and getting into a routine. It’s a lot about our mindset and making space for activities outside of work.
Finally, what do you like to do in your spare time?
Well, dancing, of course! I also enjoy winter sports like ice skating and skiing, but the UK is not really the best place to live for these activities. So, to compensate, I take flying lessons. I like progress and I need the space for challenge and creativity. I started flying a few years ago, and so far, I’ve logged about 20 hours of flying. It’s been a bit on and off because flying requires time and focus. When I’m applying for a grant, I take a break from flying, because it’s best not to have the grant application on your mind when you’re flying a plane!
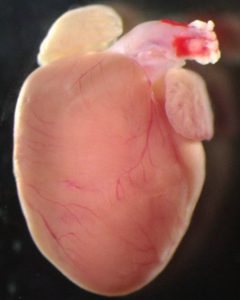
Brief summary: Congenital heart defects are the most common human birth defect, affecting around 1% of newborns. People with heart defects often show low levels of a protein that helps transport a key metabolite, citric acid, in their cells. Now, researchers in China have shown that although this protein does not directly affect heart development in mice, its loss causes problems with placental growth, leading indirectly to heart defects. The researchers also suggest potential treatments.
Image of a healthy 40-day-old mouse heart. Image credit: Wenli Fan
Press release: Congenital heart defects are the most common form of human birth defect, but we still don’t fully understand what causes them. Previous research had suggested that some heart defects could be triggered by problems with the placenta, the organ that provides oxygen and nutrients to the developing embryo. Now, researchers at Nanjing University, China have confirmed this link by focusing on a protein whose levels are reduced in many patients with congenital heart defects, called SLC25A1. SLC25A1 plays a key role in transporting citric acid, an important metabolite whose derivatives can affect gene expression, to different regions of our cells. However, it was unclear how the protein’s loss might be linked to congenital heart defects. By disrupting this protein in different tissues in developing mice, the researchers have shown that loss of SLC25A1 does not affect the developing heart directly. Instead, it leads to problems with placental growth and this, in turn, causes heart defects in the mice. The researchers publish their study in the journal Development on 26 November 2024.
The researchers used gene editing tools to produce mouse embryos that completely lack the SLC25A1 protein. As expected, these embryos developed heart defects. However, they also had problems with their placentas, which were thinner than usual. “A rapidly increasing number of studies in mice have suggested that the placenta is involved in the regulation of embryonic heart development,” explained Professor Zhongzhou Yang from Nanjing University Medical School, whose research group carried out the study. “However, the molecular basis of this relationship was unclear.”
The research team wanted to explore this potential relationship between placental defects and heart defects. To identify where in the embryo the protein is needed, they decided to remove SLC25A1 from specific tissues, rather than just disrupting it across the whole embryo. First, they made mouse embryos that lacked the SLC25A1 protein only in their developing heart. Surprisingly, these mice did not develop heart defects, suggesting that SLC25A1 is not playing an important role in heart cells. In contrast, when the researchers generated mice that lacked the SLC25A1 protein only in their placental tissue, they found that the mice developed both placental defects and heart defects. This suggested that SLC25A1 plays a key role in development of the placenta and, if placental development goes wrong, heart defects can occur as a result. Indeed, measurements showed that loss of SLC25A1 alters the citric acid balance in placental cells, resulting in changes to the cells’ DNA that disrupt placental development.
The researchers next explored how these changes to the placenta might cause problems with heart development. They found that the placentas lacking SLC25A1 had low levels of PSG1, a protein that is produced by the placental cells and is known to help regulate development of endothelia (sheets of cells that line the insides of certain structures in the body, including blood vessels). “We showed that administration of human PSG1 to pregnant mice improves placental and heart defects in embryos lacking SLC25A1,” Professor Yang explains. “PSG1 might therefore become a potentially effective drug to help improve placental and heart development of the foetus in the uterus.” While further work is needed, these findings could lead to the development of a new treatment strategy to prevent congenital heart defects in foetuses that are exhibiting placental problems.
Fan, W., Li, Z., He, X., Wang, X., Sun, M., Yang, Z. (2024). SLC25A1 regulates placental development to ensure embryonic heart morphogenesis. Development, 151, dev204290. doi: 10.1242/dev.204290
No such thing as a standard career path – an interview with Sara Morais da Silva
Sara Morais da Silva is currently a Reviews Editor at Journal of Cell Science, but publishing wasn’t her first calling. In fact, Sara has had years of experience in the lab, and then tried out careers in teaching and the industry. How did she end up as a journal editor? Here, we chatted to Sara about her research experiences, her ventures into various other careers, and why she ultimately decided to choose a career in publishing.
Could you talk briefly about your research experience and what you worked on?
I did my PhD in the MRC National Institute for Medical Research (NIMR), Mill Hill, London, in the mid-90s with Robin Lovell-Badge, working on a protein called Sox9. I found it was involved in sex determination, acting downstream of the sex determining gene Sry, and playing a key role in the differentiation of Sertoli cells. Afterwards, I moved to regeneration research, and did a postdoc with Jeremy Brockes at University College London. I worked on the proximal distal axis during the regeneration of the newt adult limb, and found that a protein called Prod 1 was responsible for cells determining their position within this axis as the regeneration proceeded. Later, I wanted to return to embryology, and I was interested in morphogenesis. I joined Jean-Paul Vincent lab in Mill Hill, where I worked on the germ band extension of the fruit fly embryo, specifically focusing on how the cell division axis contributed to elongation during extension.
At this point, I had my first child, and living in London was becoming very expensive. My husband, who wanted to transition to the pharma industry, received a good job offer in Porto, Portugal. I planned to start building my independent research there, but there was an economic crisis in 2009, and I had the opportunity to join Claudio Sunkel’s lab at Instituto de Investigação e Inovação em Saúde (I3S), working on the spindle assembly checkpoint in fruit flies. However, the economic crisis forced my husband’s company to close its Porto site. He found another job in Aberdeen, Scotland. Aberdeen offered limited opportunities in my field, so I decided to take a break, when my children were still young. During my time in Aberdeen, I did a part-time photography degree in college, which I really enjoyed. But once again, one day, my husband came home and told me his company was relocating to the southeast, to Stevenage. We moved to live in between London and Cambridge, and we’ve been settled there since.
After a break, you received a Wellcome Trust Research Career Re-entry Fellowship. What motivated you to apply and how was the application process?
By the time we moved near Cambridge, I had been away from science for almost four years, and I really missed it. I applied and succeeded in getting a Wellcome Trust Career Re-entry Fellowship to return to the lab. My project was a follow up from the spindle assembly checkpoint project that I started in Porto. I generated quite a bit of data that I am currently tidying up.
To secure a fellowship, you need a good project with substantial previous research experience. You need lab space and someone that will support you. In my case, I was fortunate to have two excellent mentors: Sarah Bray and Peter Lawrence both at Cambridge University. To be eligible, you had to be out of research for at least two years. I ticked all these boxes but applying for this re-entry fellowship was one of the hardest things I’d ever done. Writing the project proposal on my own was challenging, and the interview involved presenting to a panel of 15 people, sat around a U-shape table. When I was awarded the fellowship, I was very happy, because it gave me the possibility to re-enter science research.
How was your experience going back to the lab after a few years’ break?
I worked in the Physiology, Development and Neuroscience department in Cambridge for seven years. The re-entry fellowship was officially for four years, but I was part-time, and it was extended because of the pandemic. Overall, I loved it, but I didn’t find it as exciting as it used to be. I think the fact that I stopped for so many years gave me a bit more perspective about other things I could do. My drive to commit 100% to research was no longer there. With family, I started thinking of the pros and the cons of continuing with a career that is very insecure. All these factors that maybe didn’t affect me when I was younger, made me think twice about a career in academia. That’s why I decided to stop when the fellowship ended.
Sara reflects on her experience going back to the lab after a career break.
You said you stopped when the money ran out, but were you already starting to think about other potential careers before that point?
One consistent thread throughout my career was my husband, who is a scientist and always seemed happy and fulfilled in his career. He always had opportunities to progress and move to new challenges. I wanted a job still linked to science but that offered more stability. So just before the fellowship ended, I started applying to pharmaceutical companies, and I got a job in one of them.
How was your experience in industry?
I have to stress that it was my personal experience. But I didn’t like it and I didn’t last long – resigning after six months. There were lots of reasons. I worked for a contract research organization (CRO) that supported pharma and biotech companies. The clients came with a project and when it got to my hands it was already developed and designed. All I had to do was to go to the lab and do the experiments. I had little ownership over the work, and I could be reassigned at short notice to a new project while in the middle of an existing one. I was bored and busy at the same time and after a while, I resigned. Luckily, my husband’s stable career meant we didn’t have to panic about finances and he was very supportive all the way through.
Your stint in industry didn’t work out, what other careers did you consider?
I thought about where else I could use my experience and apply my knowledge. I considered teaching and found a retraining programme designed for people like me to go into teaching. I shadowed teachers in several secondary schools, and I saw everything – from the ideal class to a class where the teacher was screaming at the children, and the children were screaming back. From these experiences, my admiration for teachers grew. They lack adequate support, have a heavy workload and low pay. Teaching requires a calling and a talent that not everyone has and I decided it wasn’t for me. I also tried something completely different – I explored a career in the police force but while I was physically capable, I couldn’t see myself fitting into that role either.
Throughout your career, you’re very quick at deciding that something is not right for you. How did you manage to do that?
I think that comes with age and experience. Over time, I’ve learned what I like and don’t like. It’s also part of my personality – I don’t like doing things halfway. I like to have decisions made quickly. I need to feel happy and fulfilled in my job. If I’m stressed, it affects the people close to me. And life is short. If you dwell too much doing something you don’t enjoy, you’re just wasting time and postponing the inevitable.
You’re currently the Reviews Editor at Journal of Cell Science. What does your role entail?
As a Reviews Editor, I commission reviews on topics relevant to the journal’s readership. I attend conferences to stay updated on emerging fields and identify knowledge gaps. I also edit commissioned articles, ensuring they are of the highest quality, and write Research Highlights, which are one paragraph summaries of recently accepted papers in the journal. Additionally, I interview early career research scientists for JCS.
What attracted you to the world of publishing?
I have a few friends who work in publishing and once, whilst having dinner with a friend and laughing with her about all my career experiences, she said, “I don’t understand why you haven’t applied to be an editor yet! With your background, the amount of people you know and the number of scientific subjects you studied, it’s a no brainer!” So, I decided to give it a try. And luckily enough, the first advert I came across was the position that I have now!
In this role, I engage with science every day. It’s like being in a sweet shop – one day, I can be working on an article about mitochondria; the next day, I can be working on microtubules. I’m never bored. I meet a lot of people as well, which is something that I really like. I think my current role as an editor at JCS is the joining of all the experiences I had before – talking to scientists independent of the level of seniority, knowing the topics that could be interesting for the journal. This is the ideal position to consolidate all my past knowledge and experience.
Sara talks about how her past experiences led her to her current role as a journal editor.
Career breaks and two-body problem are challenges that many scientists face. How did you navigate these challenges, and do you have any advice to those who are going through them?
This is a really hard question because it’s a very personal experience. For every decision I make, I consider all the possible outcomes. My advice is to have a very, very thorough consideration about what can happen once you make a decision. Think hard about your priorities and the consequences of your choices.
If I may dig a bit deeper, I think the main reason underlying challenges like the two-body problem and career breaks in science is because the academic career structure is very poor. If there were more career opportunities and support, especially for postdocs, then maybe fewer people would leave academia.
Any advice you would give for people thinking of going into a new career?
For me, it has to be something that fits with my personality. Your job takes the largest part of your day, so you should be doing something you enjoy. It’s also important to find something that fits your experience and abilities. You need to find out which aspect of your scientific career you like the most – do you like interacting with other scientists or do you like pipetting in a lab? There are so many possible careers out there, so just choose according to your personality, and don’t be afraid of changes and getting out of your comfort zone.
I get approached a lot at conferences as a journal editor, and I volunteer to go to ‘meet the editor’ or career events at conferences. I think talking to people is very important when finding a new career. If I can offer my experience, then people can pick and take whatever they find useful.
Where do you find out more about different careers that you don’t know much about?
A good thing is to find people who are in jobs you are interested in and talk to them. I also went to a few career fairs to see what I could do. That was where I found the teaching retraining programme. And I researched loads of websites. As soon as I saw an interesting advert, I would do some research on that area and tried to talk to someone with a similar role.
Finally, what do you like to do in your spare time?
I spend most of my time with my family. I am also training to run the London Marathon, which takes up a lot of time! I love photography and painting – they are my escape. My home is like a private gallery, with my paintings and photographs everywhere. I also love going to the theatre and music gigs. I hope I’ll still be going to gigs in my 90s, listening to my favourite bands!
In this SciArt profile, we meet Petra Korlević, a scientist at the Wellcome Sanger Institute, interested in retrieving DNA from historic mosquito collections.
Can you tell us about your background and what you work on now?
I work on human malaria transmitting mosquitoes; population genetics, insecticide resistance, and currently trying to marry all that up with landscape genomics. My background is on getting DNA from difficult samples, for my PhD I worked on method development for ancient DNA extraction from bones and teeth, for my postdoc I developed a method for minimally morphologically destructive DNA extraction from museum pinned insects.
I was going to be way too many things, a veterinarian, a cartoon artist, an astronomer, a physicist… and also a nature researcher, so being a biologist fits at least that box. Though thinking of plans “for retirement” having a rescue for all the abandoned cats and dogs back home in Croatia wouldn’t be bad either.
And what about art – have you always enjoyed it?
My first ever recorded and preserved doodle was crayons on my dad’s arm at 11 months old, so I had a pretty quick start. Next was the living room wall. Then paper. I was lucky I grew up in an environment where art was encouraged together with, not against, a scientific pursuit. Then it got a bit tricky in school because I would still doodle things in my notebooks which not all teachers appreciated. I had a pretty harsh art-block during my undergrads and masters, but in the end art persevered!
Huevember (drawing of one Croatian animal per day/hue)
What or who are your most important artistic influences?
I grew up on Franco-Belgian and Italian Topolino comics. Back then my main influence was my dad since he, my sister and I would sit down and paint/draw together. There are just too many artists whose work I admire to list here, but a few: Vincent van Gogh (grew up with the starry night on the wall), Michael Whelan (his sci fi and fantasy realism is breathtaking, we used to sit down and make stories from his paintings without knowing the books attached to them), Joshua from False Knees (the ways he keeps animal drawings mostly realistic but with such beautifully woven personalities in nothing but simple pencil strokes is such an inspiration), Chen Zha (chentomology) and Nicole (fossilforager) (the two of them have some of the cutest invertebrate art out there, and invertebrates always need more love) and so so many more I am so sorry I can’t put you all in here!
While I do still enjoy the occasional traditional media project (and do embroidery as a hobby), I primarily use Procreate on my iPad, and Inkscape on my laptop. One thing I wish is to have more time to actually learn new things, I would love to learn animation and other new software.
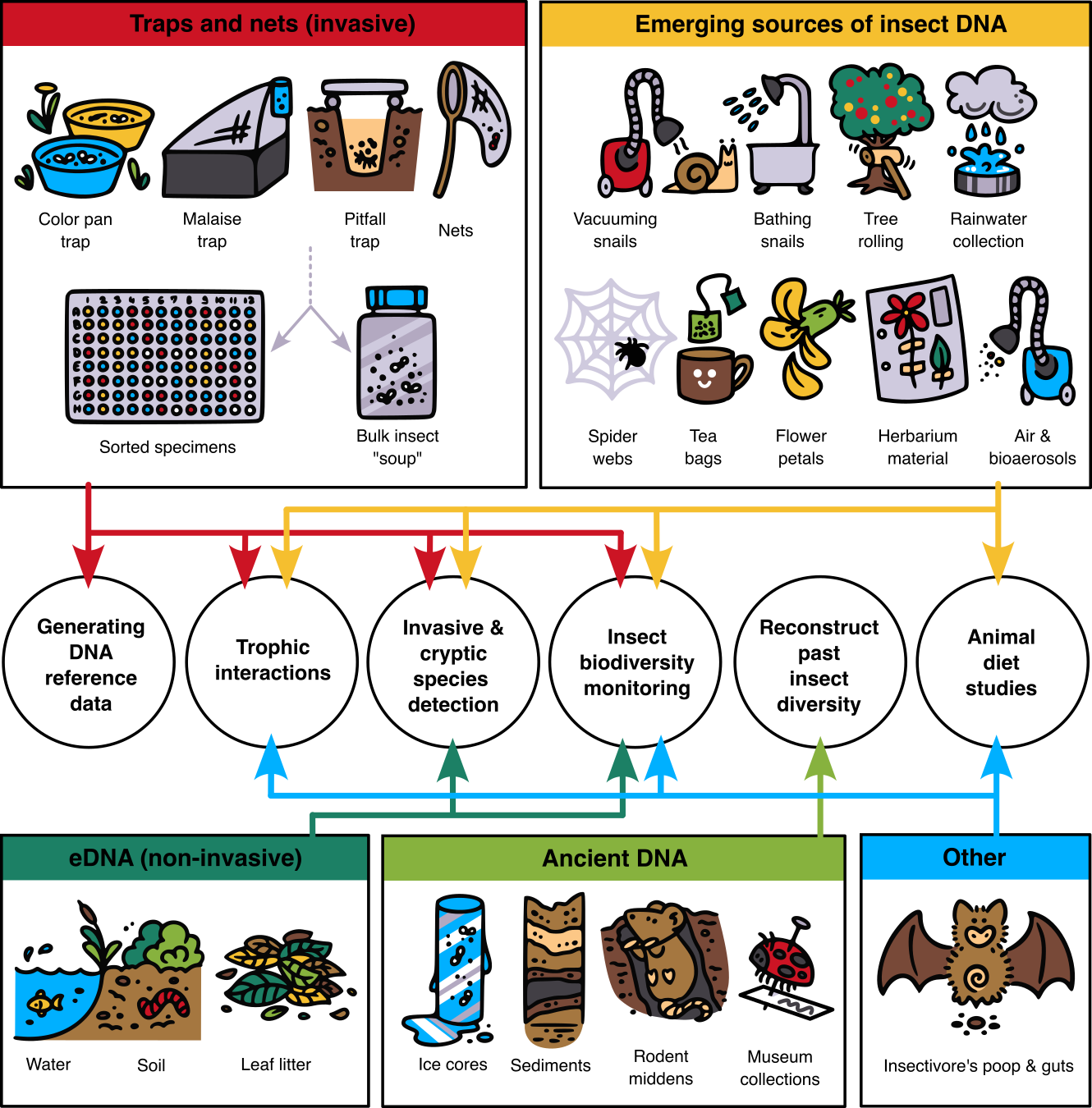
Fig2-v1 (schematic from the paper in the above Trends in Genetics issue, “Future of DNA-based insect monitoring”; really I just love the little snail getting pampered by vacuuming and bathing)
Does your science influence your art at all, or vice versa, or are they separate worlds?
It is a big symbiotic organism, both are amazing outlets for my creative process. And it definitely helps with creating engaging talks (for scientists or the general public) and having pretty paper figures.
Vector Kolymbari Meeting 2024 (live sketching of the event, with some little embellishments like a tick eating an olive and the EURO 2024 final recreated with mosquitoes)
What are you thinking of working on next?
Science-wise I need to get a wishlist of new Anopheles reference genome and get to mapping a bunch of understudied malaria vectors, then putting them into their habitat/climate context.
This year, we proudly celebrate the 30th anniversary of the Spanish Society for Developmental Biology (SEBD)—a milestone that marks three decades of passion, dedication, and collaboration among Spanish scientists worldwide, committed to uncovering the mechanisms that dictate the development of living organisms. Since its founding the 5th of May of 1994 by the visionary Antonio García-Bellido, SEBD has evolved into a vibrant and inclusive community, uniting established researchers with a growing number of emerging scientists eager to contribute to this dynamic and ever-evolving field. The integrative nature of developmental biology remains essential for understanding processes ranging from regeneration to evolution, and the influx of young talent continues to enrich our Society with fresh ideas and perspectives, ensuring that Spanish science remains at the forefront of innovation.
To commemorate this special occasion, we have gathered interviews with SEBD members who, over the years, have significantly contributed to the Society’s growth and success. These conversations reflect on personal journeys, celebrate key scientific advancements, and emphasize what makes SEBD unique: a combination of scientific excellence, creativity, and a collaborative spirit. They also underscore the Society’s role as a supportive environment where established scientists and early-career researchers work together to expand the boundaries of our understanding.
Through these interviews, we honour not only our rich history but also the exciting future that lies ahead. As we celebrate this milestone, we continue to foster connections, inspire creativity, and build upon the legacy of this extraordinary community. We hope you enjoy these reflections, which celebrate the individuals and the collective achievements that have shaped SEBD over the past 30 years.
We are confident that these qualities, which have made SEBD so special, will continue to drive its success in the future.
Eloisa Herrera is the current SEBD president. Eloisa’s research focuses on axon guidance and the formation of the nervous system. She investigates the mechanisms by which neurons establish precise connections, shedding light on the processes that shape brain wiring and their implications for developmental disorders. By combining molecular, genetic, and imaging techniques, Eloisa’s work aims to unravel the complexity of neuronal connectivity, providing critical insights into how proper brain architecture is established and maintained during development.
Sofia J. Araújo is the current SEBD secretary. Her research focuses on the genetic and cellular mechanisms that control branching morphogenesis and cell migration. Her work uses Drosophila as a model to investigate the processes involved in the development of organs like the trachea and nervous system, particularly how single-cell branching mechanisms contribute to organ formation. In addition to her research, Sofia is a keen science communicator and SciArt enthusiast. She is committed to exploring creative ways to make science more accessible, helping to bring complex biological concepts to a wider audience in an engaging and understandable manner.
Cristina Pujades specializes in vertebrate neural development, using zebrafish as a model organism to investigate the genetic and cellular processes that govern brain and spinal cord patterning. Her research centers on key processes such as cell migration, differentiation, and the establishment of neural circuits, offering fundamental insights into nervous system development and its evolutionary adaptations. Through her work, she contributes to a deeper understanding of how the nervous system forms and functions across species.
Former SEBD president Fernando Casares is a leading researcher in developmental genetics, with a particular focus on eye development and evolution. His work explores the genetic networks that regulate eye size and shape, primarily using Drosophila as a model organism. Through his research, Fernando has deepened our understanding of how alterations in developmental pathways contribute to morphological diversity, providing valuable insights into the fundamental processes of organogenesis.
Former SEBD president Miguel Torres’s research centers on the mechanisms underlying embryonic morphogenesis and tissue regeneration. He has made significant contributions to our understanding of cardiac development, employing cutting-edge tools to construct a dynamic 3D atlas of heart formation. This work has offered valuable insights into the origins of congenital heart defects and holds potential implications for regenerative medicine. Additionally, he investigates endogenous regeneration pathways in the heart, drawing inspiration from highly regenerative species such as amphibians and zebrafish.
Former SEBD treasurer Paola Bovolenta focuses on key signaling pathways, including Wnt and Shh, that regulate brain development. Her research investigates the genetic and cellular mechanisms involved in brain patterning, with a particular emphasis on congenital diseases and brain malformations. Paola’s work has been instrumental in advancing our understanding of the molecular basis of developmental disorders, providing crucial insights into how disruptions in these pathways contribute to neurological conditions.
Former SEBD secretary Miguel Manzanares investigates transcriptional regulation during early embryogenesis, focusing on how molecular mechanisms drive lineage specification and organogenesis. His research aims to uncover the regulatory networks that govern the differentiation of cells and the formation of organs, providing valuable insights into fundamental processes of early development. Additionally, his work has important implications for regenerative medicine, as understanding these processes may contribute to strategies for tissue regeneration and repair.
Former SEBD treasurer Pilar Cubas is an expert in plant developmental biology, with a focus on the regulation of axillary bud dormancy and shoot branching. Her research delves into the genetic and molecular networks, including the role of TCP genes and strigolactone signaling, that control these processes. By employing model systems such as Arabidopsis thaliana, tomato, and potato, her team investigates how these regulatory mechanisms influence plant adaptation and productivity, providing insights that could enhance crop yield and agricultural sustainability.
Former SEBD president Ángela Nieto is widely recognized for her pioneering work on epithelial-to-mesenchymal transitions (EMT), processes that are crucial in development, fibrosis, and cancer metastasis. Her research has significantly advanced our understanding of how these cellular transitions regulate embryonic development and the pathological consequences when they are dysregulated. Ángela has received numerous prestigious national and international awards in recognition of her transformative contributions to the field of developmental biology.
This year’s SEBD Science Communication prize winner Luisma Escudero is a researcher best known for his discovery of the scutoid, a geometric shape important for understanding tissue formation and organ development. His work explores how geometry influences cellular packing and tissue organization, contributing to advancements in developmental biology and tissue engineering. Alongside his research, Luisma is an enthusiastic science communicator, and has recently published a children’s book “Papá, ¿cómo se enroscan las caracolas?”, which explains complex scientific concepts like scutoids in an accessible way. Through his research and outreach efforts, he aims to make science more accessible and engaging for the public.
Luisma Escudero, winner of the 2024 SEBD Science Communication Award
Former SEBD board member Isabel Fariñas is a leading expert in neural stem cell biology, investigating their role in brain development and adult neurogenesis. Her research aims to understand how neural stem cells maintain their potential for regeneration and differentiation throughout life. Isabel’s work has implications for understanding brain plasticity, repair mechanisms, and neurodegenerative diseases. Isabel has received this year’s Ramon y Cajal National Science Award from the Spanish Science Ministry.
Is there such thing as a standard career path for a scientist? Some people find fulfilment in pursuing a PhD, postdoc, then starting their own research group; others may have second thoughts or may be drawn by other interests and aspirations.
In this new series, we chatted to several developmental biologists who have had vastly different career trajectories. This interview series was inspired by this 2013 post from Kara Cerveny, who went back into academia as an assistant professor after a stint as a scientific editor.
Below is a sneak peak of the career stories.
One common thread throughout all the interviews is that there is no right or wrong when choosing a career path. Every decision is profoundly personal.
We hope that whichever career stage you are at, and whatever career journey you are embarking on, these stories can help demonstrate that many valid career paths exist. It is okay to take breaks, make U-turns and go on tangents – you might end up doing something you’ve never dreamed of!
I’d like to thank everyone featured in this series so far, who’ve kindly taken time out of their busy schedules to chat to me. I appreciate everyone’s honesty and willingness to share their personal stories.
I hope you enjoy reading these interviews.
Joyce Community Manager, the Node
Do you have an unconventional career path and want to share your story? Or do you know someone who does? Get in touch at thenode@biologists.com
Sneak peak of the career stories
Eve Seuntjens is currently the Principal Investigator of the Developmental Neurobiology Lab at KU Leuven, Belgium. Originally trained as a pharmacist, Eve decided to embark on an academic career but spent almost 16 years as a postdoc before landing an independent position. What is her advice to people currently in the endless postdoc period? Find out more from our interview with Eve.
Originally trained as an architect, Karen Liu made an active decision in her late 20s to pursue a scientific career, starting from washing the dishes as a lab technician. How was Karen’s journey towards becoming a professor in developmental biology? Read the interview to find out.
“I don’t think I would’ve been better off being a biologist to begin with”
Bill Hinchen has always wanted to go down the academia route, but an unfortunate series of events led him to pivot his career to science writing. When he was given a second chance to do a PhD, what made him decide to turn down the offer? How is he finding life as a freelancer? Read our conversation with Bill.
“I never wanted to leave academia, but now I love being a freelance science writer”
Before becoming the Reviews Editor of Journal of Cell Science, Sara Morais da Silva has dipped her toes in many different careers – from teaching, to pharma, and even a brief stint in the police! What has she learned along the way? Check out the interview with Sara.
“Choose what fits with your personality”
To Maria Rostovskaya, science and dance have always been equally important in her life. Between her undergraduate and PhD, Maria was a dancer and taught dancing. What made her switch careers and follow an academic path? How did her experience in dancing shape her subsequent career? Read the interview.
“I made a conscious choice to switch from dancing to research, and I don’t regret it at all“
Instead of doing a postdoc right after his PhD, Christos Kyprianou joined The Company of Biologists as FocalPlane’s Community Manager. After setting up the community site for microscopy, what made Christos decide to return to academia as a postdoc? We interviewed Christos to find out more.
“There’s no one particular path of doing science – explore different options”
No such thing as a standard career path – an interview with Karen Liu
Karen Liu is Professor of Genetics and Development at King’s College London, UK. She did English and architecture as an undergrad and worked in an architectural firm for a couple of years before deciding to pursue a scientific career. We had a chat with Karen to find out more about her switch from architecture to science, and how her own experience has influenced how she guides early career scientists through their career journeys today.
Can you briefly tell us about your career path so far?
I’m currently Professor of Genetics and Development at King’s College London, and I’ve been here my entire independent career. I started here as a lecturer in 2007 and before that, I was at Stanford for three years in the labs of Michael Longaker and Gerald Crabtree as a joint postdoc. Prior to that, I did a PhD at UC Berkeley with Richard Harland. Before my PhD, I was a technician at Columbia for two years, in a mouse lab with Argiris Efstratiadis and that was where I started in science. I was the only tech in the lab, so my duties included washing the dishes, lab management and a lot of mouse work. I barely knew what DNA was! I’m still astounded that I now have this job as a biology professor, because as an undergraduate, I went to Columbia University and studied architecture and English.
Karen talks about making an active decision to switch from architecture to biology.
Let’s wind back the clock: you started out training as an architect. How was that experience?
The architecture degree I did was very theoretical. I knew during the degree that I didn’t want to be an architect. Like science, architecture is a creative field but it’s very much about the funding pressures. To be a famous architect, you need a lot of resources backing you up, and the field comes and goes with the vagaries of the economic situation. I was at university in the early 90s, when there was a major economic crash. When I got out of university, I managed to get a job at I. M. Pei. This was a very exciting period in architecture – they had just done the Louvre Pyramid, the Rock and Roll Hall of Fame and other amazing buildings. But I didn’t work as an architect. I had an English degree as well, so I worked in the office where we put out press releases and wrote grant applications. I did a lot of architectural writing and putting together proposals.
What influenced your decision to pursue a scientific career?
Because of the economic situation, it was not a stable job. I didn’t want to stay in architecture, but I had to think hard about what to do next. I had a fair amount of science background in high school. When I was working at I.M. Pei, I was ill for a while. I thought I needed to do something useful for humans, but I also want to do something fun. With biology, there’s an infinite number of questions out there. I thought if I could become a biologist, there will always be something I could be excited about. I didn’t know anyone who was a research scientist; it just seemed like a fun thing to go to grad school for.
Your entry into the research world was a lab technician job. How did that come about?
I didn’t take any biology in college, so initially I was looking around for night school. Then I found out that at Columbia, where I’d done my undergrad, offered employees two courses a term for free. So, I decided to find a job at Columbia. I was talking to the HR person at Pei, and she mentioned she had a friend who ran the sequencing facility at Columbia. After talking to them, they in turn mentioned that the lab of Argiris Efstratiadis was looking for a technician. I went to talk to Argiris, and amazingly, he gave me a job based on nothing. I think he liked interesting people who had done different things and who genuinely wanted to be in biology.
I started out washing pipettes and bottles and making media in the lab. In the meantime, I took some undergraduate classes, like genetics, developmental biology, and biochemistry. Biochemistry included organic chemistry, which totally kicked my ass! By the second year I worked there, they said I could take the first year PhD seminar course in the genetics department. It was during this course that I read about all these famous and wonderful genetics and development papers. The tutors included Gary Struhl and Iva Greenwald, and Ginny Papaioannou taught us mouse genetics. They made a developmental biologist out of me!
You’re currently a Professor of Genetics and Development at the Centre for Craniofacial and Regenerative Biology, King’s College London. What is your research about?
My lab works on the neural crest. It’s a vertebrate-specific population that arises in the embryo. The neural crest is multipotent in the embryo. The neural crest cells are born at the neural crest border, and then migrates. We are interested in the migratory properties of these cells and how they interact with their surrounding environment as the embryo develops. Because the embryo is changing dramatically during those stages, the neural crest cells need to crawl through tight spaces and navigate complicated environments. We’re interested in the molecules and the proteins that are important for human development and the phosphorylation events that control the interactions between cells. That’s the fundamental basis of the lab. On the other end, we’re identifying new gene variants from human patients who have congenital neural crest anomalies. Our focus is on head structures because the crest forms most of the skeleton in the head and some cardiac tissues. We collaborate with people who work on other neural crest derivatives, like the enteric nervous system. From a migration perspective, we’re also interested in a neural crest cancer called neuroblastoma, which arises from the sympathoadrenal system. We try to keep our work rooted within human anomalies, and that’s an active decision that I made when I started my lab. I wanted to make sure the things I do are relevant to the human condition.
Are you involved in other research initiatives?
I lead the Congenital Anomalies Cluster of the MRC National Mouse Genetics Network. Our overarching goal is to improve the way we model novel gene variants identified from patients with congenital anomalies. There is an infinite number of gene variants, but if you can’t attribute functional changes to them, then it’s meaningless data. The initiative is aimed at modelling these variants in mice. We are loosely linked with the Rare Diseases UK network, because a lot of the neural crest anomalies are rare diseases. We’ve just finished a Horizon 2020 training network called NEUcrest, which is a clinical to basic science PhD training programme spanning 15 students in 10 countries across Europe. That was exciting and rewarding to have a network of people focused on modelling neural crest anomalies. For me, at this stage of my career, I enjoy enabling these kinds of scientific interactions to have a broader impact. I’m also the academic lead for the Crick. I have oversight over all the Crick PhD students who are registered at King’s. I’m stepping down now after seven years. I run a PhD programme at King’s called Multiscale Models for Life, which is a new multidisciplinary programme where the two supervisors are from different disciplines. I also have a role at Kings as a postgraduate coordinator. In general, I think a lot about the different aspects of being a PhD student.
How did your previous roles help you get to where you are now? Do you see any similarities in architecture and science?
I think everyone winds up being the person they are like based on their route. From the architectural perspective, a lot of the themes still go through the research that we’re interested in now – how things are built, how they work together, how they function and what happens when things go wrong. I think the aesthetics of architecture really made me intolerant of bad figures or posters. I like schematics to be clean and figures to be aligned. I’ve not done any bench science myself in a long time, but I still love sketching out ideas and making schematics of our hypotheses. We’re also interested in 3D modelling now, like AlphaFold modelling of variants. Computational modelling in biology has matured over the years. A lot of that arose from architectural drawing and 3D renderings. I appreciate architecture from a distance much greater now than from within.
There’s also a similarity in terms of the collaborative nature of the work. The architectural field is very much about group work. People have their own expertise, but they have to bring all that together to make a building. The same is true in biology. I have people in my lab who love biochemistry and will only run western blots, and people who specialise in dissecting embryos. We need people to come together to build a magnificent story. I like seeing how things fit together and I like helping my people see that.
Karen compares architecture and biology.
Anything you’ve learned from switching career paths? Looking back, would you have done anything differently?
It’s a big question! I don’t think I would’ve been better off being a biologist to begin with. I don’t regret the long way to where I am now. For me, being a biologist is an active decision I made when I was in my 20s. I’m really fortunate that I was able to choose that and then move into it and wind up being a professor now. I was pretty terrified when I went into the PhD – I was 28 years old, and I knew zero biology going in. I think I did not take enough advantage of my undergraduate education. I was a really bad student, but one thing I would say to students now is, even if you’re a really bad student, you can come out of that and figure out what you really like, and then slowly work your way there. I don’t know if I could switch careers like I did now. I probably won’t be brave enough. It’s also helpful if you have a safety net. I think a lot of people couldn’t do that. Back then, perhaps I was a bit naïve. I figured if I could pay my rent, I’d be okay. If grad school didn’t work out, I could always get another job.
Do you have any advice to someone thinking of switching career paths? Has your own experience influenced how you give career advice?
I think people who haven’t switched careers can’t appreciate how difficult it is. Some people think that because they’ve just finished a PhD, switching feels like they’re failing. It’s not. You just did this PhD where you became the world’s expert on your topic! I think the hardest part is to get over that feeling that it’s a failure. Then you need to do a bit of self-reflection. What are you good at? Some people think, I’m not good at anything other than running Western blots. That’s so not true. You just wrote a 200-page thesis. You’ve presented your data again and again, coordinated with many people, and put together an entire project with many moving parts. You can ask yourself, am I good at the people part? Or the writing part? Would I prefer to be by myself all day, or would I prefer to spend a lot of time with others? Those simple questions are a really good start.
Then you can talk to all kinds of people and find people who are supportive. This can be difficult, but people are more supportive than one thinks. Looking back, I found that people really made an effort to point out particular opportunities and help me move to the next phase. Like that woman from the architectural firm, she didn’t have to tell me about her friend at Columbia’s sequencing facility, but she was helpful and was excited about my career prospects (I should really write to her and thank her!). So now in my own lab, I always try to point out opportunities to students.
I did work hard to get to where I am now, and I think people should own that. Of course you have to work hard, but it’s easy to work hard if you’re in a nice environment. But certain work environments are not so great, and if you could tell it isn’t the right place for you, then it’s time to take a step back and try to figure out how to get yourself out of that situation. We read these articles about changing careers and how you have to be brave etc. I think people in more senior positions need to appreciate that being able to change is a privilege, and I feel strongly about this. You cannot change if you don’t have a safety net, so we need to be providing that safety net, within the university and within the PhD programme.
Karen stresses the importance of providing career support for PhD students.
Finally, what do you enjoy doing outside of research?
I like good food and I like traveling. I like to do long-distance walking. My husband, who is also a scientist, and I have been trying to do things that are a little bit outside of our comfort zone. We took a pottery class recently. I’m terrible at it, but it’s fun!

 (No Ratings Yet)
(No Ratings Yet)
 (2 votes)
(2 votes)