Hopefully some of you will have seen the recent editorial in Development on our approach to peer review. If you haven’t read it yet, please do take a look. In it, James Briscoe (the journal’s Editor-in-Chief) and I discuss some of the initiatives that the journal has taken to try and support authors through the peer review process – including, most recently, encouraging authors to include a ‘Limitations’ section in the discussion of their article, giving you an opportunity to lay out explicitly the scope and extent of your study and, where appropriate, to respond to referee concerns by acknowledging them rather than addressing them experimentally.
Off the back of this editorial, James has also written a blog post that I’d really encourage you to read. Entitled ‘In Praise of Peer Review‘, James sets out why he believes that peer review (in some form) is an invaluable and irreplaceable part of scholarly communication. Alongside the debate that’s been going on around eLife’s exclusion from Web of Science (and subsequent decision to send a partial feed of articles for indexing, the piece has generated some discussion on social media both around whether peer review actually works to guard against publication of fraudulent, sloppy or otherwise dubious papers, and around the degree to which it actually helps to improve papers. I think James has done a great job of setting out the ‘why’ of peer review, but here I thought I’d give my view on the ‘what’: what should a peer review report comprise?
But before I start, let’s remember that – in the majority of cases at least – peer reviewers are both 1) highly knowledgeable in the field of the paper they’ve agreed to review and 2) well-meaning. Yes we all know of cases where papers have been sent to referees that weren’t sufficiently expert or who set out to block publication for political or petty reasons. But these are in the minority – most reviewers are competent to do the job they’ve been asked to, and they want to do it well. And they do it for little or no reward, because they believe that it’s an important part of their responsibility as a member of the academic community. If or how they should be rewarded is a whole other topic that I won’t get into now, but I am incredibly grateful for their dedication.
So, what do I want a referee to do?
Firstly, I want a referee to be respectful. Remember that there are people behind the data and – before hitting the ‘submit’ button on their report – pause to consider the potential impact of your words on the authors, particularly the students and postdocs who’ve actually done the work. At Development, we’re very fortunate that the vast majority of referees do abide by this guidance, but that’s not to say that I’ve not come across the odd report that felt overly combative or dismissive in tone – and that’s not OK.
Secondly, I want the report to be reasonable in terms of the amount of additional work requested. Think about the amount of time (and money!) that might be involved in addressing any particular point and ask how important that point really is to the main story of the paper. Which leads me on to:
Thirdly, I’d ask the referee to focus primarily on addressing the question ‘do the data support the conclusions?’ and not ‘what could the authors do to make the conclusions more interesting?’. While it’s very useful to get expert opinion on how important/relevant/useful/important the paper will be for the community, it’s primarily the editor’s job to decide on whether the paper is – in principle – appropriate for the journal in question.
And finally, I want the referee to be honest about what aspects of the paper they can and can’t (or even did and didn’t) assess. Are you able to judge if the authors have used appropriate statistical analyses? (And if so, did you actually check?!) If the paper contains computational work, do you have the expertise to assess it fully? If the authors deposited data, did you look at it? We fully appreciate that referees can’t always be experts in every area of a paper – particularly an interdisciplinary one – and we try to recruit referees with complementary expertise, but it’s really useful to know what you did and didn’t review.
Most reports I read (and I read a lot!) do largely follow these guidelines, but there is still a definite tendency for a referee report to read a bit like a shopping list of potential experiments and textual revisions. Experienced authors can often read the nuance to decide which points to tackle experimentally, and good editors will (either pro-actively or in response to author queries) help to navigate the revision process. But referees can also do their bit to shepherd papers through the often all-too-painful process of publishing by remembering that there’s both a financial and a temporal limit to how much a group of authors can (and should) do to revise a paper, that a single paper can’t solve a whole research question, and that their opinion isn’t necessarily any more valid than that of the authors (or, for that matter, the other referees).
We could discuss ad nauseam the benefits and problems of pre-publication peer review in its current form (and I frequently do!), and alternative models are beginning to emerge that can act in parallel to, or even replace, our current system. But let’s also think about the little steps that we can take to make the current system less onerous and more constructive – thus easing the path to publication.
The massive presence of disorder and variability challenges the traditional metaphor of the developmental process as a perfectly executed program leading to precise mechanisms at every level [1,2]. Yet, the final outcome —the organism— remains both astonishingly complex and remarkably reproducible. This paradox piqued the interest of Dimitri Fabrèges and Takashi Hiiragi. Back then, around 2017, Takashi was research group leader at the EMBL Heidelberg, and Dimitri a postdoc in his group. They began to explore the idea of disorder and variability from a provoking viewpoint: instead of undermining the precision of the developmental process, randomness and variability might actually act as driving forces that ensure precision and reproducibility.
Motivated by this hypothesis, the researchers focused on the early stages of mammalian development; particularly, on the initial cleavage process of mouse, rabbit and monkey embryo, encompassing the first cell divisions post-fertilization up to the 16-cell stage. A first analysis showed that the division times of different cells in the same embryo were progressively being desynchronized. A pivotal moment in this sequence is the 8-cell stage —that is, after 3 division rounds— a moment in which cells already divided in a quite disorganized way. Due to this high variability, researchers found that the beginning of this stage was characterized by a highly heterogeneous set of cell packing configurations. However, such initial variability is smoothly but steadily reduced along the so-called compaction process, leading to a seemingly common, spherical-like structure at the end of the stage. Such a structure guarantees that, in the next round of divisions —i.e., at the 16-cell stage— there will be a suitable proportion of inner and outer cells. Achieving this correct proportion is essential: inner cells will lead to the organism itself, while outer cells form the placenta and extra-embryonic material. This observation raised a challenging question, namely, how can one support the intuitive claim that embryos begin highly heterogeneous but become remarkably similar with a more rigorous foundation.
This was the perfect challenge for Virginie Uhlmann, an expert on biological image processing who, at the time, had just started as research group leader at the EMBL-EBI, in Cambridge. She tackled this question by developing an advanced computational framework able to analyze and track the geometric changes in the embryonic shape in high detail. This approach conceptualized an embryo’s developmental path as a trajectory within a high-dimensional space whose coordinates captured relevant geometrical properties [3]. The key result was that, indeed, the trajectories exhibited significant initial disparity but converged surprisingly by the end of the compaction process in a particular region of the abstract space that characterized the embryo geometry.
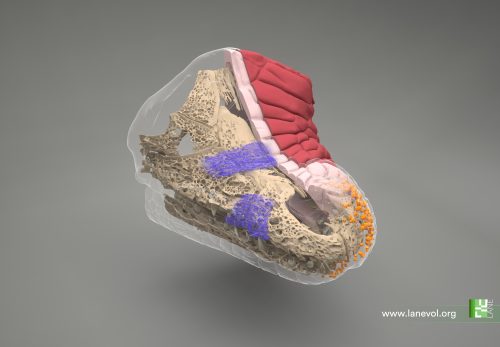
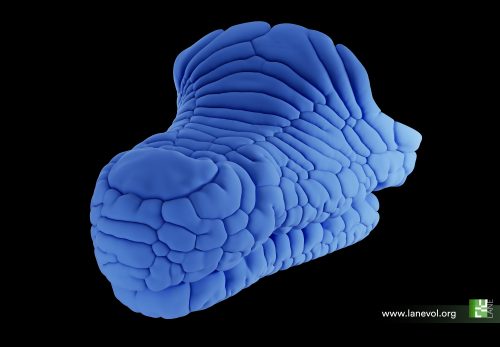
Which structure was represented in this region? Why did this particular structure seem to act as the developmental “target”? At the beginning of the winter of 2018, Takashi’s research group organized a retreat in the Catalan coastal town of Sitges, gathering several groups from the ISTA. Among the attendees were Edouard Hannezo, who just opened his research group as PI, and his first postdoc, Bernat Corominas-Murtra —both physicists working on biological problems. The evening was windy and stormy, and a little café was the refuge where they largely discussed with Dimitri and Takashi about the challenge of identifying and explaining the emergent structure. Although no immediate solution came out, Edouard and Bernat concluded that a deeper and simpler structural characterization was necessary —that is, complementing the geometric analysis with a topological one, stripping out all details but the raw structure. Weeks after, they stumbled upon a relatively recent publication showing a key mathematical finding: there are exactly different 13 ways to pack 8 spheres such that none of them exhibit independent movement. This result provided the key to define a classification scheme: either the embryos conform approximately to one of these 13 packing configurations, or they are floppy, meaning that some cells retain independent movement [4]. By establishing a suitable notion of “distance” among sphere packings, the researchers could classify embryos at various developmental time points. Their analysis revealed that, although variability was very high at the onset of the 8-cell stage, as the compaction process progressed, embryos consistently converged towards these similar packing structures along similar developmental pathways. At that point, the target structure was identified: the D2d packing of 8 spheres, in the Schoenflies notation.
Fig 1: 4-cell stage embryos give rise to many shapes at the beginning of the 8-cell stage, during which cell contractility triggers topological transitions. Ultimately, embryos are driven toward the most optimal packing (cyan). In parallel, the cell-autonomous desynchronization progressively increases temporal variability and helps to maintain topological optimality through generations, lowering spatial variability and promoting robustness. Picture taken from (Fabreges et al. 2024).
How does the embryo, without any external help, solve this kind of Rubik’s cube, i.e., transition from an arbitrary cell configuration to a specific optimal only one through successive cell rearrangements? Looking at the empirical data, one observable stood out above the other due to its clear trend: Adhesion was increasing along the compaction. Edouard suggested to challenge the simple hypothesis whether this slight change in the cell adhesion was enough to trigger all the topological rearrangements. The hypothesis has deep consequences. It implies that an increase on the cell adhesion could not only trigger deformations within the cells (i.e., increasing the contact surface, for example), but also qualitative reorganizations of the whole embryonic cell mass in a reproducible way. Computer simulations showed that such a genetically encoded slight increase in cell adhesion, coupled with significant random fluctuations in cell positions —disorder— was paradoxically facilitating the transition from any arbitrary packing of cells to the single optimal configuration. This hypothesis stands out as the simplest and, in the case of the mouse, it enabled even to reproduce in-silico the developmental trajectories of real embryos. In the case of rabbit and monkey, the role of other agents, like the zona pellucida —an external membrane that may exert a compressing force to the cell packing— could not be fully discarded.
At this point, the puzzle of the convergence towards a common, suitable embryo configuration was solved. However, the role of the temporal variability, which was experimentally observed at the starting point of the whole project and inspired it all, remained to be understood. Using several genetic perturbations, the results were surprisingly concluding that initial variability was actually required to achieve precise convergence. In particular, embryos in which cell divisions occurred more synchronously than in the wild-type ones showed a poorer convergence at the end of the compaction process, thereby hampering the further development of the embryo. The provoking hypothesis of Takashi and Dimitri on the role of stochasticity was thus proven to be fully consistent.
The researched path was not easy: Big part of the project was carried out during the COVID-19 pandemic. In turn, during the project, Dimitri and Takashi moved to Utrecht, to the Hubrecht Institute, Virginie to the University of Zurich, and Bernat to the University of Graz. Researchers from several institutions1 provided their bits of knowledge in the multiple challenges that paved the achievement of the results, and, as in living organisms, the sum of different expertises —biology, physics, mathematics and computer science— ended up in something that was much more than the sum of its parts. As in any adventurous interdisciplinary research, moments of joy and concern alternated, sometimes without pause in between… All in all, this research provides a new, constructive interpretation of the striking amount of disorder observed along developmental stages: When coupled to the changes in cell mechanics, the interplay among them can lead to significant and precise reorganization events within embryos, paving the way for a new understanding on how complex geometries and, in general, organization patterns arise in living beings. Disorder, therefore, far from being a problem the system has to deal with, may be one of the leading forces driving the precision of organism development.
Publication:
Dimitri Fabrèges et al. Temporal variability and cell mechanics control robustness in mammalian embryogenesis. Science 386, eadh1145 (2024)
References:
[1] M. Carlson, W. Reeves, M. Veeman, Stochasticity and stereotypy in the Ciona notochord. Dev. Biol.397, 248–256 (2015).
[2] R. Dumollard, N. Minc, G. Salez, S. B. Aicha, F. Bekkouche, C. Hebras, L. Besnardeau, A. McDougall, The invariant cleavage pattern displayed by ascidian embryos depends on spindle positioning along the cell’s longest axis in the apical plane and relies on asynchronous cell divisions.eLife6, 1–23 (2017).
[3] R. Delgado-Gonzalo, N. Chenouard, M. Unser, Spline-based deforming ellipsoids for interactive 3D bioimage segmentation. IEEE Trans. Image Process.22, 3926–3940 (2013).
[4] N. Arkus, V. N. Manoharan, M. P. Brenner, Minimal energy clusters of hard spheres with short range attractions. Phys. Rev. Lett.103, 118303 (2009).
1Other institutions involved:
Institute for the Advanced Study of Human Biology (WPI-ASHBi), Kyoto University, Kyoto, Japan.
Department of Developmental Biology, Graduate School of Medicine, Kyoto University, Kyoto, Japan.
Research Center for Animal Life Science, Shiga University of Medical Science, Shiga, Japan.
INRAE, BREED, Paris-Saclay University, Jouy-en-Josas, France.
École Nationale Vétérinaire d’Alfort, BREED, Maisons-Alfort, France.
PhD position in the Denholm lab at the University of Edinburgh, UK
You will use modern techniques to study the development and/or physiology of one of the most powerful water-conserving systems in nature – the beetle cryptonephridial (or ‘buried kidney’) complex.
Insects can live and thrive in some of the most inhospitable environments on earth, including extremely desiccating conditions such as deserts. Many species possess a powerful water-conserving system called the cryptonephridial (or ‘buried kidney’) complex (CNC), which recovers water from the rectum and recycles it back to the body. This remarkable system even allows water vapour absorption from moist air, providing a novel physiological mechanism for water uptake. It is estimated that >400,000 insect species have a CNC, with CNCs being particularly common in beetles. The broad principles underpinning CNC physiology were laid down half a century ago, and the system has since become a staple textbook example of a countercurrent exchange system. Despite this, next to nothing is known about CNC development, molecular physiology, endocrinological regulation or evolution.
In this project you will use the model beetle species Tribolium and exploit enabling technologies including genomics, single-nuclei RNAseq, informatics, imaging and in-vivo analysis to identify how this system develops and functions. We have catalogued gene expression profiles (using snRNAseq) from the CNC of this species (in both embryo and adult), providing a window into its embryonic development and the molecular players involved in its physiological function at single-cell resolution.
The techniques you will use and be trained in include: (1) Bioinformatics. You will use this to prioritise key genes involved in the development and function of the system. (2) Hybridisation chain reaction fluorescent in situ hybridisation. You will use this to map expression of candidate genes in embryonic, larval and adult CNCs. (3) Gene knock-down: you will use RNAi to knock-down gene activity for each candidate and, use (4) Microscopy (both fluorescence confocal and electron microscopy) and simple physiological assays to establish roles for these genes in CNC development and function.
Results from this project will significantly expand our understanding of one of the most powerful water-conserving systems in nature, one that is fundamental to insect physiology, ecology and evolutionary success.
From the hard, protective scales of reptiles to the soft, insulating fur of mammals, amniotes are equipped with a remarkable range of skin appendages that help them thrive in diverse environments. To fulfil their various functions, such as mechanical protection, thermoregulation, and the provision of camouflage, skin appendages must be precisely arranged (or patterned). For instance, birds display a regular spatial arrangement of feathers which enable flight (1), whereas snakes exhibit a near-perfect hexagonal pattern of scales, which are aligned to their muscles and ribs (2), to aid locomotion.
All of these skin appendages develop from ‘placodes’ (3, 4) comprised of an epidermal thickening, an underlying aggregation of dermal cells, and conserved molecular signalling. It is well-established that the spatial patterning of placodes is self-organised through interactions between activatory and inhibitory morphogens, forming a so-called Turing reaction-diffusion system (5-7). In other words, the resulting self-organised “polka-dot” arrangement of gene expression provides a template defining where skin appendages will develop (1, 3, 4, 8).
Remarkably, the irregular polygon-shaped head scales of crocodiles present a fascinating exception to this paradigm, as they appear to emerge from a mechanical process, i.e., in the total absence of a genetic template (9). However, the exact nature of this process has remained elusive for more than a decade. In our new article published this week in Nature (10), we solve this mystery through a multidisciplinary approach combining experimental manipulations of developing crocodile embryos, cutting-edge light sheet microscopy, and state-of-the-art numerical simulations.
First, we imported a large number of Nile crocodile eggs from South Africa to our laboratory in Geneva, Switzerland. After incubating these embryos until their head scales begin to emerge, we performed precise in-ovo intravenous injections of epidermal growth factor (EGF) protein that, as its name indicates, increases epidermal growth while also promoting keratinisation. As we increased the dose of EGF, we observed the development of an extensive labyrinthine pattern of convoluted folds across the jaws of the crocodile embryo, instead of the normal polygonal scales. These results demonstrate that crocodile head scales develop through compressive folding and that EGF treatment exacerbates this folding. Note that tissue buckling from ‘constrained growth’ is thought to shape other biological forms, including the intricate folds of the human brain (11), villification of the human and chicken gut (12), the surface wrinkling of mucosa (13), and the small polygons on the glabrous skin of the rhinarium (or naked nose) of dogs, cows and ferrets (14). Interestingly, we have recently showed that, for this latter example, the polygonal form of the pattern is imposed by the underlying polygonal pattern of blood vessels that act as ‘mechanical positional information’ (14).
Remarkably, when we allow the crocodiles to hatch four weeks after the EGF treatment has finished, they no longer display a fully labyrinthine pattern, as it partially relaxes into a pattern of smaller polygons. Importantly, this pattern resembles that of another crocodilian species – the spectacled caiman. Therefore, simply by promoting skin growth and differentiation during embryonic development, we can alter the hatchling head scale pattern from that of a Nile crocodile to that of a caiman. Therefore, shifts in the molecular components of an intricate Turing reaction-diffusion system are not required to explain the evolution of head scale patterns in crocodilians. Instead, slight variations in the expression of a very small number of genes affecting skin growth and/or stiffness can clearly explain this macroevolutionary change.
We next used cutting-edge light sheet microscopy to build a realistic 3D computer model of the crocodile embryo’s head before the emergence of scales. This model incorporates the key elements that we suggest determine the mechanical patterning of crocodile head scales. This includes the geometry of the skin (such as the spatial variation of epidermal and dermal thicknesses) and underlying bone, the organisation of collagen fibres within the dermis, and the relative growth rates of the two skin layers. Remarkably, our computer growth simulations accurately reproduce the natural head scale arrangement of Nile crocodiles, which includes elongated units on top of the jaws and smaller polygons on the sides. In addition, increasing both skin growth and stiffness in our simulations replicates the labyrinthine brainy folding seen in our EGF-treated embryos. These results show that the polygonal arrangement of crocodile head scales can indeed emerge from a purely mechanical system of constrained skin growth, in the complete absence of self-organising gene interactions.
Overall, our findings show that the polygonal head scales of crocodiles emerge from compressive folding of the skin. Importantly, natural variations to the skin’s growth and material properties have allowed the evolution of diverse head scale patterns observed across different crocodilian species. Therefore, while gene interactions can drive many examples of embryonic patterning (7), self-organisation in development is not restricted to such chemical interactions. Indeed, self-organised physical processes also contribute to the emergence of intricate biological forms (15-17).
Check out our (hopefully appealing) 3D imaging data and numerical simulations in the following video summarising our results:
English version
———————————————————————
A shorter English version
———————————————————————
The French version
———————————————————————
Versions in other languages (Spanish, German, Japanese and Chinese) will be made available in January 2025.
References
1. W. K. W. Ho et al., Feather arrays are patterned by interacting signalling and cell density waves. PLoS biology17, e3000132 (2019).
2. A. C. Tzika, A. Ullate-Agote, S. Zakany, M. Kummrow, M. C. Milinkovitch, Somitic positional information guides self-organized patterning of snake scales. Sci Adv9, eadf8834 (2023).
3. N. Di-Poï, M. C. Milinkovitch, The anatomical placode in reptile scale morphogenesis indicates shared ancestry among skin appendages in amniotes. Science Advances2, e1600708 (2016).
4. R. L. Cooper et al., An ancient Turing-like patterning mechanism regulates skin denticle development in sharks. Science Advances4, eaau5484 (2018).
5. A. M. Turing, The Chemical Basis of Morphogenesis. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences237, 37-72 (1952).
6. S. Kondo, T. Miura, Reaction-Diffusion Model as a Framework of Understanding Biological Pattern Formation. Science (New York, N.Y.)329, 1616-1620 (2010).
7. M. C. Milinkovitch, E. Jahanbakhsh, S. Zakany, The Unreasonable Effectiveness of Reaction Diffusion in Vertebrate Skin Color Patterning. Annu Rev Cell Dev Biol39, 145-174 (2023).
8. S. Sick, S. Reinker, J. Timmer, T. Schlake, WNT and DKK determine hair follicle spacing through a reaction-diffusion mechanism. Science314, 1447-1450 (2006).
9. M. C. Milinkovitch et al., Crocodile Head Scales Are Not Developmental Units But Emerge from Physical Cracking. Science339, 78-81 (2013).
10. G. N. Santos-Durán, R. L. Cooper, E. Jahanbakhsh, G. M. Timin, Michel. C., Self-organised Patterning of Crocodile Head Scales by Compressive Folding. Nature, 11 December 2024.
11. T. Tallinen et al., On the growth and form of cortical convolutions. Nature Physics12, 588-593 (2016).
12. A. E. Shyer et al., Villification: How the Gut Gets Its Villi. Science342, 212-218 (2013).
13. B. Li, Y.-P. Cao, X.-Q. Feng, H. Gao, Surface wrinkling of mucosa induced by volumetric growth: Theory, simulation and experiment. Journal of the Mechanics and Physics of Solids59, 758-774 (2011).
14. P. Dagenais et al., Mechanical positional information guides the self-organized development of a polygonal network of creases in the skin of mammalian noses. Curr Biol, (2024).
15. B. Li, Y. P. Cao, X. Q. Feng, H. J. Gao, Mechanics of morphological instabilities and surface wrinkling in soft materials: a review. Soft Matter8, 5728-5745 (2012).
16. A. J. Hughes et al., Engineered Tissue Folding by Mechanical Compaction of the Mesenchyme. Dev Cell44, 165-178 e166 (2018).
17. A. Bailles, E. W. Gehrels, T. Lecuit, Mechanochemical Principles of Spatial and Temporal Patterns in Cells and Tissues. Annu Rev Cell Dev Biol38, 321-347 (2022).
Matthew J. Stower, Richard C. V. Tyser, Shifaan Thowfeequ, Felix Zhou, Marta Portela, Konstantinos Miti, Jacintha Sugnaseelan, Xin Lu, Shankar Srinivas
Xiaofei Li, Rainy Wortelboer, Yi Song, Sahana Balasubramanian, Callie McLain, Alex Hernandez Manriquez, Joseph Suh, Brenton D. Hoffman, Adam V. Kwiatkowski, Glenn L. Radice
Ana R. Hernandez-Rodriguez, Yisha Lan, Fengtong Ji, Susannah B.P. McLaren, Joana M. N. Vidigueira, Ruoheng Li, Yixin Dai, Emily Holmes, Lauren D. Moon, Lakshmi Balasubramaniam, Fengzhu Xiong
Oguzhan F Baltaci, Andrea Usseglio Gaudi, Stefanie Dudczig, Weili Wang, Maria Cristina Rondon-Galeano, Ye-Wheen Lim, James Rae, Anne Lagendijk, Robert G Parton, Alison Farley, Benjamin M Hogan
Sarah Bowden, Magdalena Maria Brislinger-Engelhardt, Mona Hansen, Africa Temporal-Plo, Damian Weber, Sandra Hägele, Fabian Lorenz, Tim Litwin, Clemens Kreutz, Peter Walentek
Laura Lahti, Nikolaos Volakakis, Linda Gillberg, Behzad Yaghmaeian Salmani, Katarína Tiklová, Nigel Kee, Hilda Lundén-Miguel, Michael Piper, Richard Gronostajski, Thomas Perlmann
Hedda B. Somsen, Kristel N. Eigenhuis, Anne L. Kuijpers-Korporaal, Mariana Pelicano de Almeida, Roberto Montoro Ferrer, Kelsey Kats, Mike R. Dekker, Thuur Zuidweg, Zakia Azmani, Mirjam C.G.N. van den Hout, Wilfred F.J. van IJcken, Danny Huylebroeck, Maarten Fornerod, Raymond A. Poot, Debbie L.C. van den Berg
Daniel M. Snell, Wazeer Varsally, Aurélien Courtois, Sergio Menchero, Prabhakaran Munusamy, Richelle Rietdijk, Obah A. Ojarikre, Stephanie Strohbuecker, Haskan Kaya, Mahesh N. Sangrithi, James M.A. Turner
Victoria Osorio-Vasquez, Jan C. Lumibao, Kristina L. Peck, Kathryn Lande, Jonathan Zhu, McKenna Stamp, Shira R. Okhovat, Hyemin Song, Satoshi Ogawa, Jasper Hsu, Yang Dai, Angelica Rock, Chelsea Bottomley, Ethan Thomas, Alexandra Fowler, T’Onj McGriff, Siri Larsen, Muhamad Abdulla, Phil Greer, Jessica Gibson, Michael Downes, Ronald Evans, Jingjing Zou, Andrew M. Lowy, David C. Whitcomb, Rebekah White, Melena Bellin, Herve Tiriac, Dannielle D. Engle
Karen Bellec, Lynsey R Carroll, Kathryn AF Pennel, Yuanliangzi Tian, Yachuan Yu, Alexander R Cameron, Fabiana Herédia, Alisson M Gontijo, Joanne Edwards, Rippei Hayashi, Julia B Cordero
Angela Armento, Inga Sonntag, Ana-Cristina Almansa-Garcia, Merve Sen, Sylvia Bolz, Blanca Arango-Gonzalez, Ellen Kilger, Ruchi Sharma, Kapil Bharti, Rosario Fernandez-Godino, Berta de la Cerda, Simon J Clark, Marius Ueffing
Nedaa Al_Jezani, Asmaa Affan, Catherine Leonard, Nabangshu Das, Luiz Gustavo Almeida, Daniel Young, Anand O Masson, Antoine Dufour, Paul Salo, Pam Railton, James N Powell, Roman J Krawetz
Jose Blanco-Ameijeiras, Mar Garcia-Valero, Yara El Majzoub, Elena Rebollo, Javier Macho-Rendon, Jorge Corbacho, Juan Ramon Martinez-Morales, Elisa Marti
Anupama Rao, Andrew Russell, Jose Segura-Bermudez, Charles Franz, Rejenae Dockery, Anton Blatnik, Jacob Panten, Mateo Zevallos, Carson McNulty, Maciej Pietrzak, Joseph Aaron Goldman
Dorsa Toghani, Sanika Gupte, Sharon Zeng, Elmir Mahammadov, Edie I. Crosse, Negar Seyedhassantehrani, Christian Burns, David Gravano, Stefan Radtke, Hans-Peter Kiem, Sonia Rodriguez, Nadia Carlesso, Amogh Pradeep, Alexis Georgiades, Fabienne Lucas, Nicola K. Wilson, Sarah J. Kinston, Berthold Göttgens, Le Zong, Isabel Beerman, Bongsoo Park, Derek Janssens, Daniel Jones, Ali Toghani, Claus Nerlov, Eric Pietras, Marion Mesnieres, Christa Maes, Atsushi Kumanogoh, Thomas Worzfeld, Jin-Gyu Cheong, Steven Z Josefowicz, Peter Kharchenko, David T. Scadden, Antonio Scialdone, Joel A Spencer, Lev Silberstein
Davide Cinat, Rufina Maturi, Jeremy P. Gunawan, Anne L Jellema-de Bruin, Laura Kracht, Paola Serrano Martinez, Yi Wu, Abel Soto-Gamez, Marc-Jan van Goethem, Inge R. Holtman, Sarah Pringle, Lara Barazzuol, Rob P. Coppes
Soomin Cho, Emilia Servián-Morilla, Victoria Navarro Garrido, Beatriz Rodriguez-Gonzalez, Youxi Yuan, Raquel Cano, Arjun A. Rambhiya, Radbod Darabi, Robert S. Haltiwanger, Carmen Paradas, Hamed Jafar-Nejad
Mariana A. S. Artur, Robert A. Koetsier, Leo A. J. Willems, Lars L. Bakermans, Annabel D. van Driel, Joram A. Dongus, Bas J. W. Dekkers, Alexandre C. S. S. Marques, Asif Ahmed Sami, Harm Nijveen, Leónie Bentsink, Henk Hilhorst, Renake Nogueira Teixeira
Christine T. Nolan, Ian Campbell, Anna Farrell-Sherman, Bryan A. Briones Ortiz, Kerry A. Naish Ph.D, Veronica Di Stilio Ph.D, James E. Kaldy Ph.D, Cinde Donoghue Ph.D, Jennifer L. Ruesink Ph.D, Takato Imaizumi Ph.D
Salek Ahmed Sajib, Michał Bykowski, Pedro Barreto, Caroline Mauve, Etienne Delannoy, Sophie Blanchet, Rim Chamas, Markus Schwarzländer, Łucja Kowalewska, Bertrand Gakière, Livia Merendino
Silvia Basanta, Daniel J. Stadtmauer, Jamie D. Maziarz, Caitlin E. McDonough-Goldstein, Alison G. Cole, Gülay Dagdas, Günter P. Wagner°, Mihaela Pavličev
Sara Nolbrant, Jenelle L. Wallace, Jingwen Ding, Tianjia Zhu, Jess L. Sevetson, Janko Kajtez, Isabella A. Baldacci, Emily K. Corrigan, Kaylynn Hoglin, Reed McMullen, Matthew T. Schmitz, Arnar Breevoort, Dani Swope, Fengxia Wu, Bryan J. Pavlovic, Sofie R. Salama, Agnete Kirkeby, Hao Huang, Nathan K. Schaefer, Alex A. Pollen
Nikolaos Papadopoulos, Siddharth S. Kulkarni, Christian Baranyi, Bastian Fromm, Emily V.W. Setton, Prashant P. Sharma, Andreas Wanninger, Georg Brenneis
Alexandra T. Lion, Sophie M. Bodine, Kelley R. McCutcheon, Mayank Ghogale, Santhan Chandragiri, Deema Abayawardena, Bikram D. Shrestha, Abigail Descoteaux, Kathryn Alvarez, J’nesse A. Balkman, Breelyn Cocke, Athula H. Wikramanayake, Jennifer Schlezinger, Joyce Y. Wong, Vivek N. Prakash, Cynthia A. Bradham
Maddison L. Graffeo, Joseph Nguyen, Farin Yazdan Parast, D. Jo Merriner, Jessica E.M. Dunleavy, Denis Korneev, Hidenobu Okuda, Anne E. O’Connor, Donald F. Conrad, Reza Nosrati, Brendan Houston, Moira K. O’Bryan
Cláudia C. Oliveira, José Córdoba, John R. Pearson, Elizabeth Guruceaga, Ernesto Marín-Sedeño, Melissa García-Caballero, Juan Antonio Guadix, José M. Pérez-Pomares, Adrián Ruiz-Villalba
Majid Tarahomi, Miriam S. Zagers, Simin Zafardoust, A. Mohammadzadeh, Zohre Fathi, Hasti Sareban, Farnaz Fatemi, Soleimani Fakhr, Geert Hamer, Sjoerd Repping, Femke Schrauwen, Jan P. van Straalen, Frédéric M. Vaz, Madelon van Wely, Sebastiaan Mastenbroek
Alek G Erickson, Sergey Isaev, Artem Artemov, Jingyan He, Bettina Semsch, Aliia Murtazina, Jia Sun, Katrin Mangold, Anthi Chalou, Jonas Frisen, Michael Ratz, Emma Andersson, Peter V. Kharchenko, Igor Adameyko
Julia Naas, Meritxell Balmãna, Laurenz Holcik, Maria Novatchkova, Lina Dobnikar, Thomas Krausgruber, Sabrina Ladstätter, Christoph Bock, Arndt von Haeseler, Christopher Esk, Jürgen A. Knoblich
Cecilia Laterza, Elisa Cesare, Hannah T. Stuart, Martina D’Ercole, Alessia Gesualdo, Maria Grazia La Barbera, Sara Brignani, Carmela Ribecco, Roberta Polli, Roberta Frison, Silvia Angiolillo, Andrea Maset, Onelia Gagliano, Davide Cacchiarelli, James Briscoe, Elly M. Tanaka, Alessandra Murgia, Nicola Elvassore
Theo Perochon, Zeljka Krsnik, Marco Massimo, Yana Ruchiy, Alejandro Lastra Romero, Elyas Mohammadi, Xiaofei Li, Katherine R Long, Laura Parkkinen, Klas Blomgren, Thibault Lagache, David A Menassa, David Holcman
Claudia Feng, Elin Madli Peets, Yan Zhou, Luca Crepaldi, Sunay Usluer, Alistair Dunham, Jana M Braunger, Jing Su, Magdalena E Strauss, Daniele Muraro, Kimberly Ai Xian Cheam, Marc Jan Bonder, Edgar Garriga Nogales, Sarah Cooper, Andrew Bassett, Steven Leonard, Yong Gu, Bo Fussing, David Burke, Leopold Parts, Oliver Stegle, Britta Velten
Jelly HM Soffers, Erin Beck, Daniel Sytkowski, Marianne Maughan, Devarakonda Devasi, Yi Zhu, Beth Wilson, Yu-Chieh David Chen, Ted Erclik, James W. Truman, James B. Skeath, Haluk Lacin
Our ‘Featured resource’ series aims to shine a light on the resources that support our research – the unsung heroes of the science world. In this post, Connie Smith and Martin Ringwald introduce the data and functionalities available at Gene Expression Database and talk about the future directions of the database.
What is the Gene Expression Database (GXD)?
GXD is a long-standing, freely available community resource that collects and integrates mouse gene expression data generated by biomedical researchers worldwide. Our primary emphasis is on endogenous gene expression during development, covering data from wild-type and mutant mice. Data types include RNA in situ hybridization, immunohistochemistry, knock-in reporter, RT-PCR, Western blot, and RNA-seq. Close to half a million expression images (mainly from in situ data) are readily available and accessible to searches. As an integral component of Mouse Genome Informatics (MGI), GXD combines its expression information with genetic, functional, phenotypic, and disease-oriented data, thus facilitating the study of the molecular mechanisms of development, health, and disease.
What inspired the development of GXD?
We began developing GXD in 1993 for two main reasons: (i) working in the field of mouse development, we realized that there was no database to store and integrate all the mouse expression data being published, and (ii) it was clear that gene expression information would provide a crucial link between the mouse genetic and phenotypic information already being collected by the emerging Mouse Genome Database (MGD). Our plan always was to combine MGD and GXD into the integrated resource which is now known as MGI.
The development of GXD was a pioneering effort because no comparable resources existed at the time. Our biological requirement analysis resulted in a publication in Science in 1994 and has served as a blueprint for our work ever since. The basic concepts developed there are still relevant today. This includes the discussion of the complementary nature of text-based (i.e. anatomy ontology-based) and spatial representation of expression patterns, and the notion that both approaches should be combined to integrate the data and to enable effective searching and reasoning over them.
What is available at GXD?
Since the beginning, GXD has collected classical types of mouse developmental expression data, i.e. in situ and blot data. These data are acquired through systematic curation of the scientific literature and by collaborations with large-scale expression projects.
Comprehensive literature survey. As a first step in our literature annotation work, our curators review new publications to find studies of endogenous gene expression during mouse development that use classical types of expression assays. They then index all genes that have been studied in the paper, the assay types used, and the ages analyzed. These data, combined with bibliographic information from PubMed, can be accessed using the Gene Expression Literature Search. These searches are more effective and complete than PubMed searches because the annotations are based on the entire article, including supplemental data. This index is up to date and currently includes over 33,000 publications.
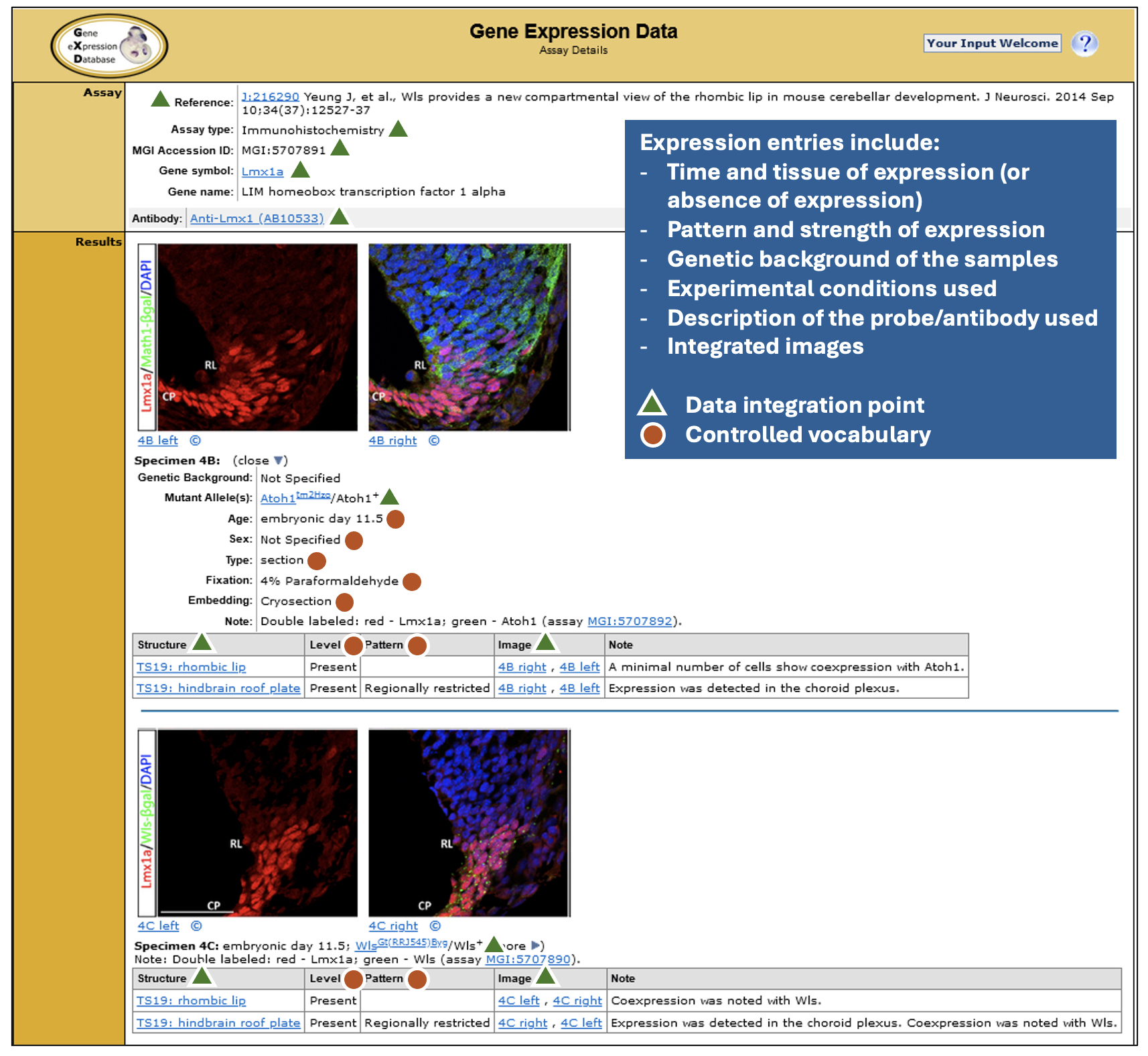
Detailed expression data that are easily searchable in many ways. GXD expression annotations are detailed, making extensive use of controlled vocabularies and ontologies, as illustrated. These standardized metadata enable the close data integration and the search and display capabilities that make GXD really shine. As of November 2024, for classical types of expression data, there are over 2 million annotated expression results for more than 16,000 genes with ~480,000 accompanying images.
Detailed Data: Expression entries include time and tissue of expression, pattern and strength of expression, genetic background of the samples, experimental conditions used, description of the probe/antibody used and integrated images.
Index of publicly available RNA-seq and microarray experiments. More recently, we created an index of RNA-seq and microarray expression experiments deposited in GEO (Gene Expression Omnibus) and ArrayExpress. Finding experiments of interest in these repositories can be difficult because the entry metadata consists of free text provided by the submitters. To address this issue, we identify studies of endogenous gene expression in wild-type and mutant mice and annotate the experiment and sample metadata using the detailed controlled vocabularies and ontologies used elsewhere in GXD and MGI. Our current index of 8,000 mouse experiments includes standardized annotations for the anatomical structure, developmental stage, mutated gene, strain and sex of the samples, as well as the study type and key parameters of each experiment. Searches using this indexed metadata can be combined with free text searching of experiment titles and descriptions to allow you to find experiments of interest more effectively and reliably.
RNA-Seq expression data. GXD-relevant bulk RNA-seq data are imported from the European Bioinformatics Institute’s (EBI) Expression Atlas. The Expression Atlas generates uniformly processed TPM-level data sets for a select set of high-quality bulk RNA-seq experiments. As part of its incorporation into GXD, these data are further processed and annotated, resulting in seamless integration with GXD’s in situ and blot data.
How can researchers access data in GXD?
Search Forms. The GXD Home Page (http://www.informatics.jax.org/expression.shtml) provides the best entry point to all the features and resources provided by GXD. In addition to the Expression Literature and RNA-Seq and Microarray Experiment searches described above, of particular note are:
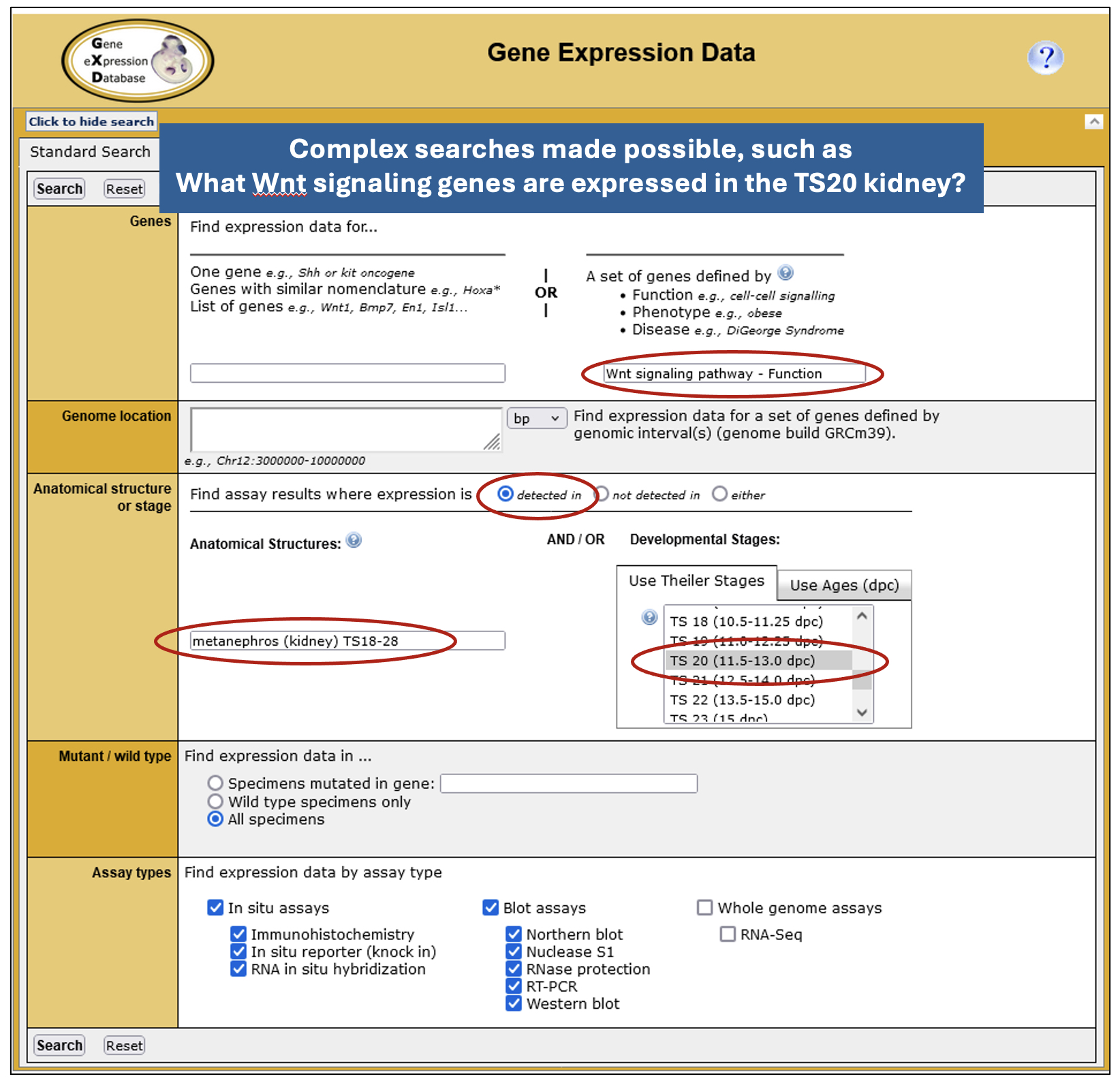
• Expression Data and Image Search – powered by GXD’s detailed annotations and data integration, this form provides the most fields, enabling basic or complex searches tailored to specific use cases.
Complex searches possible, such as “What Wnt signaling genes are expressed in the TS20 kidney?”
• Expression Profile Search – this allows you to search for genes by their expression profile. You can specify up to 10 anatomical structures and whether expression is present or absent in these structures. While currently limited to classical expression data, this search will soon be expanded to allow searching of RNA-seq data and the specification of developmental (Theiler) stages.
• Search Using Gene List – this permits you to retrieve GXD’s expression data for lists of genes and is, for example, useful for the further analysis of gene sets identified via high-throughput transcriptomic studies.
• Developmental Anatomy Browser – allows you to navigate through the extensive mouse developmental anatomy ontology used and maintained by GXD and provides links to the expression and phenotype data associated with those structures.
• MouseMine – provides programmatic access to GXD data.
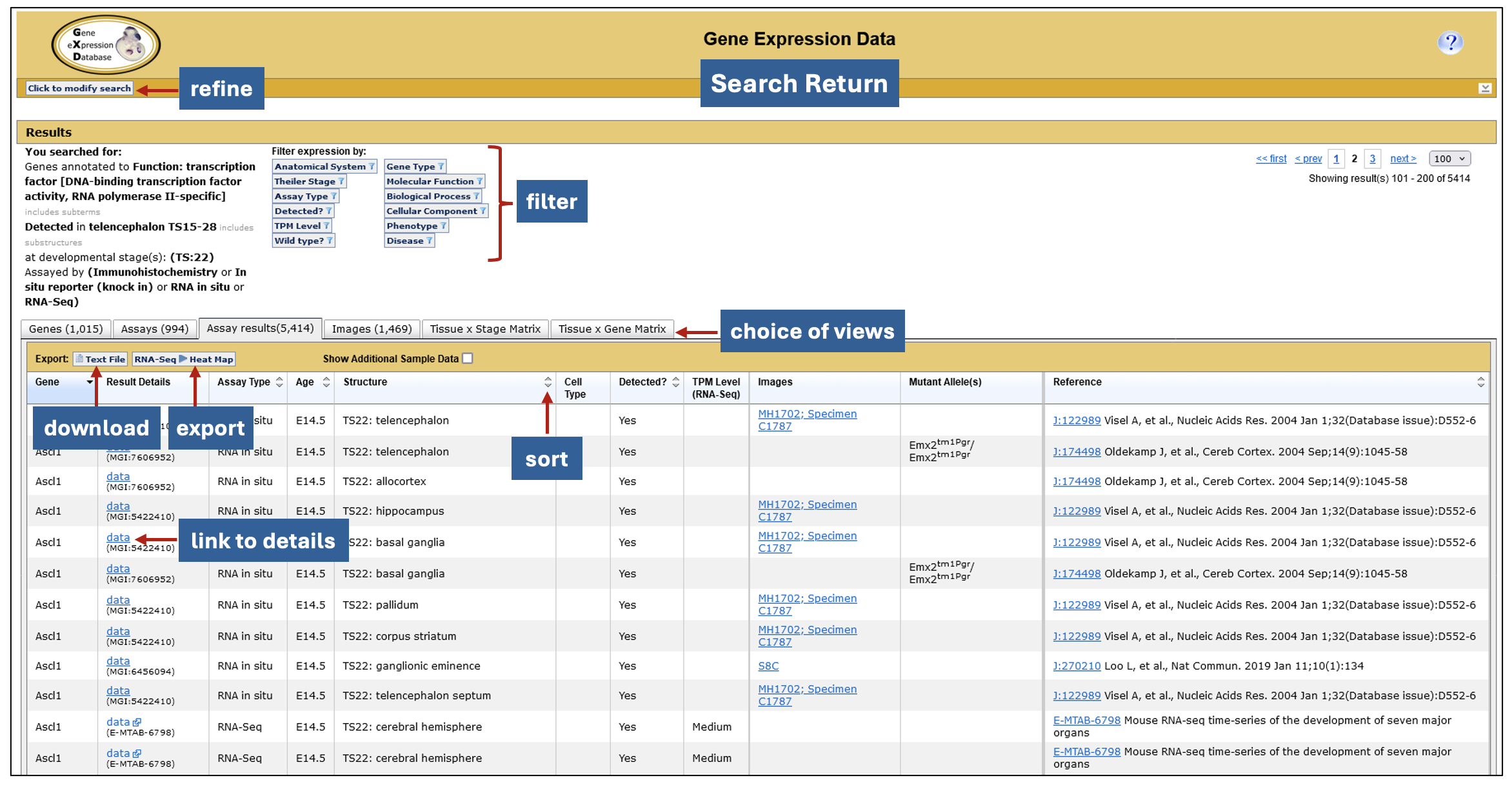
Interactive search returns. GXD expression data searches generate a six-tabbed data summary, allowing for different views of the search results. Filters that use the genetic, functional, phenotypic and disease-related information in MGI, as well as attributes of the returned expression data, allow you to tailor the return further. The data can be exported to other applications for further analysis including, in the case of RNA-seq data, Morpheus, a heat map visualization and analysis tool developed at the Broad Institute. Morpheus offers a myriad of utilities for further display and analysis, including sorting, filtering, hierarchical clustering, nearest neighbor analysis, and visual enrichment.
Interactive summary of search returns.
Any hidden gems that researchers might be less aware of?
Our gems are “hidden in plain sight” – the search forms described above. Most users of GXD/MGI use the Quick Search found in the upper right-hand corner of the pages. This provides a quick entry point into the data, but our search tools give you the capability to formulate queries using a wide variety of parameters derived from the integration of data in GXD and MGI. This allows you to successfully execute precision searches that can only be dreamt of at other resources.
Who are the people behind GXD?
GXD is located at The Jackson Laboratory in Bar Harbor, Maine. GXD scientific curators all hold Ph.Ds and have expertise in molecular and developmental biology. MGI has an integrated software team, supported by both the GXD and MGD grants, and integrated administrative and user support personnel. The following list represents our current staff. Many others have contributed to the success of GXD since 1993.
• Principal Investigator – Martin Ringwald
• Co-Principal Investigators (and co-leaders of the MGI Software Team) – Joel Richardson and Richard Baldarelli
• Scientific Curators – Jacqueline (Jackie) Finger, Terry Hayamizu, Ingeborg McCright, Constance (Connie) Smith, and Jingxia (Jing) Xu
• MGI Software Team – Jeffrey Campbell, Lori Corbani, and Pete Frost
• User Support – David Shaw
• Administrative Assistant – Janice Ormsby
Where does GXD’s funding come from?
GXD received startup funding from the W.M Keck Foundation. Since 1996 GXD has been supported by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD).
How can the community contribute to GXD?
We accept direct submissions. Review our Guidelines for submitting expression data and then contact us at gen@jax.org.
How can the community help GXD?
Cite GXD in your publications. This allows us to demonstrate our utility which helps us secure our funding.
What are the current and future directions of GXD?
We will be expanding our offerings to users who are interested in cell types. We have already begun to annotate expression data and samples from high-throughput expression experiments using terms from the Cell Ontology (CL). In the near future, users will be able to browse, search, and filter expression results and RNA-seq and microarray index entries by cell type.
We are also analyzing how we can represent and integrate results from newer types of expression analysis into GXD, namely single-cell RNA-Seq and spatial genomics data.
Following the plans outlined in our 1994 Science paper, we worked with the Edinburgh Mouse Atlas project (EMAP) to enable the spatial representation of expression data. Unfortunately, the EMAP project ended several years ago. However, similar projects and collaborations should hold big promise for dealing with spatial genomics data.
Ringwald M, Baldock R, Bard J, Kaufman M, Eppig JT, Richardson JE, Nadeau JH, Davidson D. 1994. A Database for Mouse Development. Science265:2033-2034. PubMed
Ringwald M, Mangan ME, Eppig JT, Kadin JA, Richardson JE, and the Gene Expression Database Group. 1999. GXD: A Gene Expression Database for the laboratory mouse. Nucleic Acids Res.27:106-12. PubMed
Smith CM, Finger JH, Hayamizu TF, McCright JJ, Eppig JT, Kadin JA, Richardson JE, and Ringwald M. 2007. The mouse Gene Expression Database (GXD): 2007 update. Nucleic Acids Res.35: D618-D623. PubMed
Finger JH, Smith CM, Hayamizu TF, McCright IJ, Xu J, Eppig JT, Kadin JA, Richardson JE, Ringwald M. 2015. The mouse gene expression database: New features and how to use them effectively. Genesis. doi: 10.1002/dvg.22864. PubMed
Finger JH, Smith CM, Hayamizu TF, McCright IJ, Xu J, Law M, Shaw DR, Baldarelli RM, Beal JS, Blodgett O, Campbell JW, Corbani LE, Lewis JR, Forthofer KL, Frost PJ, Giannatto SC, Hutchins LN, Miers DB, Motenko H, Stone KR, Eppig JT, Kadin JA, Richardson JE, Ringwald M. 2017. The mouse Gene Expression Database (GXD): 2017 update. Nucleic Acids Res.2017 Jan. 4;45 (D1): D730-D736. PubMed
Smith CM, Hayamizu TF, Finger JH, Bello SM, McCright IJ, Xu J, Baldarelli RM, Beal JS, Campbell JW, Corbani LE, Frost PJ, Lewis, JR, Giannatto SC, Miers DB, Shaw DR, Kadin JA, Richardson JE, Smith CL, Ringwald M. 2019. The mouse Gene Expression Database (GXD): 2019 update. Nucleic Acids Res. 2019 Jan. 8;47 (D1): D774–D779. PubMed
Smith CM, Kadin JA, Baldarelli RM, Beal JS, Blodgett O, Giannatto SC, Richardson JE, Ringwald M 2020. GXD’s RNA-Seq and Microarray Experiment Search: using curated metadata to reliably find mouse expression studies of interest. Database 2020 Mar. 4 PubMed
Baldarelli RM, Smith CM, Finger JH, Hayamizu TF, McCright IJ, Xu J, Shaw DR, Beal JS, Blodgett O, Campbell J, Corbani LE, Frost PJ, Giannatto SC, Miers DB, Kadin JA, Richardson JE, Ringwald M 2021. The mouse Gene Expression Database (GXD): 2021 update. Nucleic Acids Res.2021 Jan 8;49(D1):D924-D931. PubMed
Ringwald M, Richardson JE, Baldarelli RM, Blake JA, Kadin JA, Smith CL, Bult CJ. 2021. Mouse Genome Informatics (MGI): latest news from MGD and GXD. Mamm Genome.Oct 26, 2021 PubMed
The Biologists @ 100 conference, happening 24-27 March 2025 in Liverpool, UK, will have a dedicated half-day event specifically for early-career researchers (ECRs), in the afternoon of 24 March 2025.
The ECR event will focus on career opportunities both inside and outside of academia. We’re excited to have Richard Sever, the co-founder of bioRxiv and medRxiv, as the keynote speaker at the career session. You’ll also hear from professionals from a diverse spread of career paths to discuss how skills developed during your research career can be applied in a number of different roles.
There will also be food and drinks in the evening for ECRs following the main conference’s welcome reception for those who have attended the workshop.
Biologists @ 100 is a celebration of the 100th birthday of The Company of Biologists, bringing together different communities, including cell, developmental, experimental and disease biologists, and will incorporate the Spring Meetings of the British Society for Cell Biology (BSCB) and the British Society for Developmental Biology (BSDB).
No such thing as a standard career path – an interview with Christos Kyprianou
Christos Kyprianou is a postdoctoral researcher at the Institute of Molecular Biotechnology of Austria. Instead of doing a postdoc right after his PhD in developmental biology, Christos joined The Company of Biologists as FocalPlane’s Community Manager. After setting up and getting the community site for microscopy off the ground, Christos decided to return to academia. What motivated him to switch in and out of academia? And what’s next for him?
Have you always been interested in science?
I was always interested in science. As a kid, I had this picture in my mind that I would be doing something science or medical related. Going through high school, the only thing that stuck with me was biology, and physics to a lesser degree. Then at university, the first lecture I had was on genetics and I really enjoyed it. That’s when I realised I would like to do science in one way or another.
After your PhD, you worked as the Community Manager of FocalPlane. How did this job come about?
A lot of the times, academia leads you to believe there’s one way to do science: bachelor’s, maybe a master’s, PhD, postdoc, then group leader. I remember when I was asked why I wanted to do a PhD during my interviews, I was a bit baffled, because I thought that’s the only way to do science. But now I know you can still do science in many different capacities. As I was going through my PhD, this was becoming more and more obvious to me. Towards the end of my PhD, my PI got a position at Caltech, and I was offered to go to Caltech, which I was interested in. But then the FocalPlane job advert came up, and it made me realise I wanted to try something different, and not just go down the default route of doing a postdoc after a PhD. I applied and got the job.
What was your role at FocalPlane and the Journal of Cell Science (JCS)?
I was involved in setting up FocalPlane and getting it off the ground. I was putting together the content, inviting people, organising events, and participating in events to spread the message. At the beginning, it was unclear to me how was I doing science by building a community and a website. But there were a lot of principles from science that would apply in building FocalPlane. For example, identifying who are the people that are key in driving the microscopy field, and bringing together people who are interested in microscopy but don’t have access to resources, and working as part of a team to create FocalPlane. I was also part of the Journal of Cell Science (JCS) team, so I got to read articles and write Research Highlights, which was a very good exercise in science writing. It’s about taking something quite complicated and summarising it using 200 words. Unfortunately, because of the pandemic, I feel like I missed out on doing more things for JCS, like attending conferences.
After FocalPlane, you went on to do a postdoc at the Wyss Institute at Harvard University. How did that opportunity come about and what influenced that decision?
There were a few factors, but the pandemic played a big part. For me, it changed from being all day in the lab to being at home, as the office was closed. The other thing was when I was commissioning content and talking to people about their research, I started to miss doing the hands-on science myself. And thirdly, the lab I ended up joining as a postdoc was one of the labs that I really aspired to be part of during my PhD. Magda, my PhD supervisor, was meeting that PI and they talked about some of my PhD work, so I took that opportunity to reach out to him. They were working on organ-on-chip technology, which was new and interesting to me. I thought that was the right opportunity for me to go back to doing research. I also wanted to do something translational, because I felt like I missed out on that with my PhD project. Maybe because some people say once you leave academia, nobody is going to want to take you back, I was feeling a bit pressed with time to not miss that postdoc opportunity. But I now know this is not true; I’ve seen people do that, even after longer breaks from academia.
You’re currently a postdoc at the Institute of Molecular Biotechnology of Austria. What motivated the move from the US to Austria and what are you working on now?
My primary career goal right now is to get into industry. It’s a bit less going with the flow and I’m trying to approach my career in a more targeted way, by building up a portfolio and working with projects in the translational and technology areas. Coming back to Europe was also because my family and my wife’s family are in Europe. In terms of the lab, I was looking for something that would help build my portfolio. I’m now in Nicolas Rivron’s lab that is developing stem cell-based embryos for the purposes of using it as a tool to understand health and disease in the context of fertility. It’s a technology that you can build on top of other technologies. I’m not working on a particular biological process; instead, I’m focusing more on learning how I can apply all sorts of questions using these technologies.
Apart from your research, you said you’re trying to build your portfolio in industry. How are you doing that?
The Wyss Institute in Boston, where I did my first postdoc, is a place where academia meets industry. My work there was more industry-adjacent. We were collaborating with startups and more established pharma companies to validate drugs. That was when I decided I wanted to build my portfolio towards industry. After moving to Austria, I wanted to get more exposure to what industry is like here, so I joined the initiative ‘Vienna Bio-Centre Industry Insights’. I attended a couple of seminars, and I was impressed by the quality of speakers, from names like BioNTech, Merck, and a few consulting companies. The initiative was looking for more co-leads, so I applied for the position. I think my experience at FocalPlane really helped with me being selected. Being a co-lead is a good way for me to meet people within and outside of the Vienna Bio-Center and understand the industrial ecosystem in Vienna and the wider German speaking area of Europe. We’re trying to come up with different types of events to give people an idea of what it is like to work in industry and provide networking opportunities with people in industry.
How has your experience at FocalPlane help you in your subsequent roles?
People talk about the importance of having soft skills as a scientist. It’s like a muscle I got to practise during my time at FocalPlane, such as bring people together, organising events, and knowing who to talk to. My experience at FocalPlane and The Company of Biologists helped give me more credibility in these skills. What FocalPlane has become now is beautiful. I really enjoy the variety of its content and how popular it’s become. I even met someone random at the Max Perutz Labs, who mentioned they were planning to write something for FocalPlane. It’s great that people in the field know about it and interact with it.
Looking back your career path so far, would you have done anything differently?
I would definitely have done a bit more extracurricular activities during my PhD. I think I was overly focused on doing my project and not being as involved with other things. I would like to have been more aware of what other opportunities are available and how else I could do science, and not just necessarily going down the academia path. Nowadays there are so many opportunities to try out and learn about different things, like workshops and recruitment events from consulting and biotech companies. These are good ways to get to know people with those careers and get outside of your PhD bubble.
Christos reflects on what he would have done differently during his PhD.
Do you have any advice to someone thinking of switching career paths?
I think it’s a lot about informing and preparing yourself beforehand. If you’re thinking about making a career transition, it’s more about finding out exactly what that career is. Talk to as many relevant people as possible, and really find out if that’s for you. In many jobs, I feel like you can get trained and learn the specifics of it while on the job. For my role at FocalPlane, I didn’t do much science communication before, and it’s not like I had to build another community before. I was just another disgruntled microscope user who wanted to do more with his samples, but there weren’t resources out there easily accessible to people like me. But that was what Sharon, JCS’s Executive Editor at that time, said FocalPlane needed – someone who understood the target audience and their needs. But the rest, like organising events, commissioning content and talking to people, that’s all the stuff I learned on the job.
I’ve been seeing more opportunities for people to get into industry without a PhD. You mostly get trained on the job, and there are particular ways of doing things, so you don’t necessarily need a PhD. I think it’s important that undergrads and master students know about these opportunities before going down the PhD path. Equally I’ve seen people with years of postdoc experience who managed to get into industry.
Do you think you’ll stay in academia? Are there any other career paths you’ve considered?
I’ve been in academia for quite a long time, and I don’t think it’s for me in the long run. I’m exploring different options outside of bench work that allow me to serve science in a different capacity. Business development is something I’m interested in. I’ve been talking to friends who have done this switch and seeing what it’s like. There’s no one particular path of doing science, and it really does come in many different ways. Anything that contributes to science is valid, and everybody should feel comfortable exploring the different options.
Christos encourages people to explore different options.
Finally, what do you like to do in your spare time?
Because we just recently moved to Austria, we’re still trying to get into a routine. I’m just starting to get back to things like playing squash and going on hikes. A more recent hobby is 3D printing. I have my own entry level 3D printer that I’ve been modifying. I try to design and print my own things, like a coffee funnel so I don’t make a mess when I put coffee grounds into my Moka pot. I’ve been more focused on modifying the printer, so it performs better. I think that happens to a lot of people – they get sidetracked by building the printer rather than the actual printing. You can get a pretty cheap printer to start with, because the idea is if you get a bad printer, almost certainly every single part is going to break, so you get to learn what each part does as you’re fixing it. Pretty much like I think a lot of us are figuring out our careers as we go!
Our lab studies the emergence of collective cell behavior and its connection to biological functions in development and reproduction. We focus on the regulation of cellular mechano-chemical coupling in mammalian tissues, using mouse models and cell lines. By integrating live cell imaging with mathematical modeling, we aim to uncover the design principles governing multicellular dynamic systems.
Specifically, through the use of FRET cell imaging to detect kinase activity, we have demonstrated that extracellular signal-regulated kinase (ERK) activity is mechano-sensitive and generates active cellular forces. These forces are fundamental to collective epithelial migration and tissue morphogenesis in developing murine organs, such as the lung and cochlear duct. We will further investigate this regulatory mechanism by integrating tissue geometry and topology to gain insights into not only morphogenesis but also tissue homeostasis and repair.
Another area of interest in our research relates to reproduction. Despite its physiological importance, the origin and role of germ cell dynamics in the reproductive tract remain poorly understood. To address this, we have employed intravital microscopy to study murine sperm and reproductive tracts, allowing us to observe cellular and molecular activities in living tissues. In particular, we aim to explore the biophysical mechanisms underlying collective sperm dynamics and its relationship to reproductive functions.
Lab roll call
Sound Wai Phow: I am a research assistant and the lab safety officer of the lab. My duties include lab logistics and maintaining the mice colonies used by our lab for experiments. I am currently working on a project investigating tissue fluidity and migration.
Veronica Lee Xi Min: I am a PhD student and I am investigating the effects of sperm flow on epithelial cells in a male reproductive organ known as the epididymis. For my study, I am using an in vitro microfluidic system integrated with live cell imaging!
Ankita Nayak: I am a PhD student in the lab. I work on unravelling the impact of substrate curvature on the extrusion of DNA-damaged cells from a tissue-like environment.
Nicole Ng Shu Ying: I am a research assistant and am mostly in charge of the lab’s finances and general maintenance, with a current project involving characterising the murine epididymis using various tissue clearing and imaging methods.
Thanh Phuong Nguyen: I am a research fellow in the lab. I investigate the causative relationship between mechanobiology of epididymal epithelium and sperm development.
Emmanuel Vikran S/O Puspa Nathan: I am an Undergraduate FYP student working on the interaction of aberrant cells and healthy cells with regard to ERK wave propagation.
Chen Xingyu: I am a master student doing my capstone project. And I’m learning and exploring the structure of the epididymis.
Cao Gefei: I am a new PhD student looking into a potential project investigating the dynamics of transzonal projections and ERK signaling within follicles under the context of aging.
Lee Yuan Zhe: I’m a Year 0 PhD student, hoping to look into how collective sperm motion emerges consistently, despite the heterogeneity in environmental and individual sperm cell parameters.
Favourite technique, and why?
My favorite technique is using light microscopy on cells and tissues—simply because it’s fun! I also enjoy quantifying the data and extracting information to build mathematical models. For me, creating a smooth workflow from live imaging to modeling is the ideal approach. It’s so satisfying to turn what we see under the microscope into something that can predict or explain what’s happening.
Apart from your own research, what are you most excited about in developmental and stem cell biology?
I am excited about how complex biological forms—both in shape and size (morphology) and how they’re connected (topology)—self-organize and how they are linked with physiology. Exploring the mechanisms behind these is incredibly exciting!
PI: How do you approach managing your group and all the different tasks required in your job?
We use Slack for daily communication, hold weekly lab meetings to discuss research progress, and have one-on-one meetings for project updates. While there is no strict management style, responding promptly to questions and concerns from lab members is my top priority. Since our lab is young, I make it a point to take initiative and personally demonstrate anything new, like setting up equipment or conducting experiments. My motto is: think, feel, and move!
PI and others from the lab: What is the best thing about where you work?
Tsuyoshi: My colleagues at the MBI have different background in broad disciplines, which is nice to receive great stimuli for a new idea.
Sound: The people and the facilities. Everyone (even from other labs and cores) is very friendly and willing to teach/share their expertise and knowledge without hesitation. The facilities available makes this conducive environment very easily achievable.
Veronica: Definitely the people. We are all very different in terms of work styles and personalities, yet we blend well together. The fact that almost all of us sit together also makes it easier to discuss our work, and nurture bonds beyond regular coworkership. Other than our abundance of microscopes (we have three just for our lab!), I also love how we each have our own comfortable work space. If we could give a lab tour, you would see that each member has some sort of plushie/ figurine/ ornament on our tables that truly reflects our unique personality (e.g. I have two alpaca plushies mounted on my monitor screen).
Ankita: The open-lab system allows us to interact with people from all labs at MBI. It makes for a stimulating environment with people from different backgrounds who are always willing to lend a hand. Apart from the open-lab system and core facilities which helped us a lot while setting up a new lab, I love how organised we are here. Our lab’s databases are easily accessible to all members, and up to date, which helps me plan my experiments easily. The institute also organises informational seminars and conducts training sessions for the equipment which enables us try new avenues in our work.
Nicole: The people here are all very willing to help each other out, even if we are not in the same lab team. Support teams that we call core facilities are a blessing. From IT support to daily lab amenities, as well as usage of a plethora of devices available to everybody is always well coordinated and efficient. The support teams are very willing to conduct training sessions for any device that one may want to try using, so I learnt a lot about various microscopes available in the institution in such a short period of time which is amazing.
Thanh: The setting of open-lab creates a cross-link communication between researchers across different lab with different background. Lots of new ideas and approaches come during those across-the-lab scientific discussion. Facilities of supporting cores play substantial roles in ensure that research works going smoothly and with high efficiency.
Emmanuel: The diversity of people and good facilities. Discussing plans and data with people around me within and beyond my lab is enriching, where I can receive ideas that propel success in approaching research.
Xingyu: THE ATMOSPHERE! I can feel the harmonious and positive energy working with lab members . Everyone is keen to share and help. Also working in our lab is a great opportunity to try these advanced technologies. All the advice I’ve got and accessibility to the lab will help me figure out how scientific research is carried out, and build my own academic research map.
Yuan Zhe: The fact that people don’t take things seriously. On a clownish scale from 0 to 10, I rate myself 11. So, it’s great to be at a place where I can be free to blurt out any nonsensical thoughts I have all the time.
PI and others from the lab: What’s there to do outside of the lab?
Tsuyoshi: Singapore has many parks and green spaces. Watching various birds, like hornbills, kingfishers, and even chickens, is relaxing. Running in the parks is also wonderful—you might even come across huge lizards and snakes along the way.
Sound: Weekly inter-lab dinner and games night.
Veronica: I can’t boast enough about our game night! While other labs tend to meet up over dinner, we host both dinner AND games. Members of other labs are also welcome since our lab is quite small. Recently, each of us have also delved into various sports in an attempt to keep fit/become fitter. From what I know: Tsuyoshi climbs the stairs often, Thanh runs, Sound and Ankita gym, Nicole and I swim, Gefei does all three (run, gym, swim), and Yuan Zhe probably did some sort of sport that led to his spinal cord injury (get well soon!).
Ankita: Singapore may be a small country, but it boasts remarkable cultural diversity. I love exploring foods from different traditions, all conveniently accessible through the excellent public transport system. The city is also very walkable and features impressive green spaces. Plus, with Singapore’s easy access to other parts of Southeast Asia, travelling around the region is a real pleasure!
Nicole: There is a huge variety of food from a vast array of cultures in Singapore so it is always cool to visit new food places all around the country. New cafes, eateries and old school shops pop up once in a while and it’s always an interesting experience to try them out.
Thanh: Singapore is a modern city and therefore having a lots of way to have fun outside of the working time. People are colleagues during working hours and switch to friends after work for hanging out and explore Singpore’s unique food and culture.
Emmanuel: Singapore, being well-lit and even more beautiful at night, makes nighttime runs exciting. The island features stunning paths that stretch for several kilometers, allowing uninterrupted runs without the need to stop for traffic.
Xingyu: I like the small talks and lunch with people. I’m quite shy now but I’ll try to engage more in the future!
Yuan Zhe: I enjoy vertical marathons and late-night cycling. Singapore has enough skyscrapers for the former, and enough large green parks for the latter. When the crowd is sparse late at night, the parks are pretty much Autobahn for cycling.
No such thing as a standard career path – an interview with Maria Rostovskaya
Maria Rostovskaya is a senior research scientist at the Babraham Institute, Cambridge, UK, studying development using human pluripotent stem cells. Between her undergraduate degree and her PhD, Maria was a dancer and dance teacher for a few years. What made her decide to switch careers and eventually follow an academic career path? How did her experience in dancing shape her subsequent career? We chatted to Maria to find out more.
How has your career path been so far?
I did my undergrad in molecular biology at the Moscow State University. After university, I decided to take a break from the academic career. I became a dance teacher. I also did some lab work during that time, but I did not think of really continuing a career in academia. Then, three years later, I decided to go back to academia and pursue a PhD. I was accepted to the PhD programme at the Max Planck Institute in Dresden. I had two main directions during my PhD – my main project was about bone marrow stem cells and my side project was on human pluripotent stem cells. After my PhD, I continued with a postdoc at the Cambridge Stem Cell Institute. Now, I’m a senior scientist at the Babraham Institute at Cambridge, working on human pluripotent stem cells in the context of human development.
After your university degree, you decided to become a dancer and dance teacher. Could you talk us through that decision-making process?
I was curious about many things, and I had always done dancing and science in parallel. During my time at the university, I focused a lot on my studies, so after that, I wanted to expand my other interests. Another reason is, at that time, I was not ready for the transition from studying to doing research. My undergraduate degree involved a lot of theory, and I enjoyed learning about concepts and principles. I think people don’t always realize the difference between learning the theory and doing the research: there is a huge difference in terms of the intellectual load. How we learn from a day spent with textbooks is very different from how we learn from a day working in a lab, and I just wasn’t ready for this difference in dynamics.
How was your experience as a dancer and a dance teacher? What type of dance did you specialize in?
The style of dance is called the Hustle dance. It’s a social dance, and it’s a fusion of salsa, mambo, and west coast swing. It’s a partner dance and can be danced with different music. There is no prepared scheme. It’s a lot about leading, following and improvising with the music. Its uniqueness comes from its freedom and flexibility. You can add various styles and elements. You can style it according to your own feelings.
My dance partner and I also created a unique dance style and fusion. We also designed a teaching course, and we were invited to run workshops in other cities and even abroad. We won several competitions. It was also rewarding to see the achievements and results of my students, because they also won competitions. Some of them went on to become dance teachers. It was a really exciting time, because I had the space for creativity and the opportunity to try various things.
You were enjoying dancing, but then you decided to pursue a PhD. What motivated that decision?
I think I had the feeling that I achieved a lot in dancing, and I thought that I needed to develop further. By that time, I have gained more self-awareness, and I understood what I actually need. For me, it’s important to have opportunities for further development, for professional growth. I like to have a sense of purpose and be intellectually stimulated. With this new-found awareness, I realized that probably I could try going back to academia and pursue a PhD.
Maria reflects on why she decided to switch from dancing to doing a PhD.
How did you approach applying for a PhD and choosing which area you wanted to work in?
At that time, I wasn’t entirely sure that I wanted to go back to academia. I wanted to try, so I chose one PhD programme and applied. I thought if I did not get it, I would just continue with dancing. But I did get it, so I decided to change my career path. It was a real match – it was the right time, the right place, and the right project. I was always excited about molecular biology and cells, and it was a really the right combination of topics for the project.
Was it difficult going back to science after a break?
At that stage, not really, because I was still at the very beginning of a research career. I did not feel very different from everyone else. But because I had already tried another career, I knew a PhD was what I wanted. I think that made the difference for me.
After your PhD, you decided to stay in academia and went to Cambridge to do a postdoc. You’re currently a senior research scientist at the Babraham Institute, UK. What is your research about?
When I came to Cambridge, I was first at the Cambridge Stem Cell Institute. Then I made the transition to the Babraham Institute. Now, I use human pluripotent stem cells to model early human development. I study the transcription and epigenetic control of the first lineage decisions. I’m interested in understanding how cells become different at the right time and the right place, and what molecular mechanisms control these decisions. A few years ago, I decided that I also need to add computational skills to my portfolio. There were a lot of big data in my project, and I always had to rely on other people to do the analysis, so I decided to learn bioinformatics. Now in my work, I combine wet lab and computational approaches.
Did being trained as a dancer and teaching dancing help your scientific career? Do you see any parallels in your dancing and scientific experiences?
I think it really shaped my approach to teaching. I do a lot of teaching, and I’ve been teaching for the university for 10 years already. I supervise undergraduate students, and I find a lot of similarities in that respect. I like finding a clear way to explain and structure the knowledge, training skills, identifying key points and filling gaps. I enjoy interacting with my students and I’m very excited seeing them progress. There is definitely a parallel there. Dancing and doing research are also about having a space for creativity, trying out new things, and not being afraid of going to the unknown. Both require being open to something unexpected.
Do you still dance and teach dancing?
I don’t teach dancing anymore, because when I moved to Germany, I had to focus on my PhD. I did not speak German, and I needed to settle in a different country. But I do dance a lot and go to socials events. I often go to festivals or boot camps in different countries. I try to be a part of the dancing community. With the social dance, you don’t need to have a specific partner. You just go and start dancing with any partner.
What’s next for you? Are you planning to continue in academia?
I had another decision point in my career two and a half years ago: my PI left the institute to join a biotech company. By that time, I was already a very senior postdoc and very independent in my research. I knew that I wanted to continue in academia, so I stayed at the Babraham. Throughout my postdoctoral years, I have managed to create my own research niche and approach. I work very independently now, and occasionally I supervise research assistants and summer students. I’m planning to continue the same line of research when I start my own independent group, and I’d like to keep teaching as a part of my career too.
Anything you’ve learned from switching career paths? Looking back, would you have done anything differently?
I certainly learned a lot from this switch. I was not just following the flow of doing a PhD after a science undergraduate degree. I took a break, and I think at that time, that was the right decision at that period of my life. I do not regret at all, not even a second. I don’t know how it would have been otherwise, but if I did not make that decision to take that break and do something else, I wouldn’t have been here now. I gained self-awareness, and it made me value what I have now. Pursuing career in academia is a conscious choice – I have a dream, and I want to follow this dream, and it is worth it. My career switches made me confident that right now, this is the right place for me. But I am open to changes in the future; we don’t know where life brings us.
Maria talks about not regretting switching career paths.
Do you have any advice to someone thinking of switching career paths?
I would say, have the courage to try. If you feel like you want to try different activities, just give it a go. You will never know unless you try, and perhaps it’s better not to regret about not trying.
You’ve always had dancing and science in your life. But have you ever thought about other career paths?
Maybe a dog walker… I’m just joking. I love dogs, but with an academic career, it’s really hard to have a dog. I already have dancing and science. I think two is a good number!
How do you find time for dancing, despite your busy work schedule?
I think it’s really important to have physical and socializing activities in your life. I think this is why I try to keep dancing in my life as much as I can. It’s about prioritising, organising yourself and getting into a routine. It’s a lot about our mindset and making space for activities outside of work.
Finally, what do you like to do in your spare time?
Well, dancing, of course! I also enjoy winter sports like ice skating and skiing, but the UK is not really the best place to live for these activities. So, to compensate, I take flying lessons. I like progress and I need the space for challenge and creativity. I started flying a few years ago, and so far, I’ve logged about 20 hours of flying. It’s been a bit on and off because flying requires time and focus. When I’m applying for a grant, I take a break from flying, because it’s best not to have the grant application on your mind when you’re flying a plane!
 (5 votes)
(5 votes)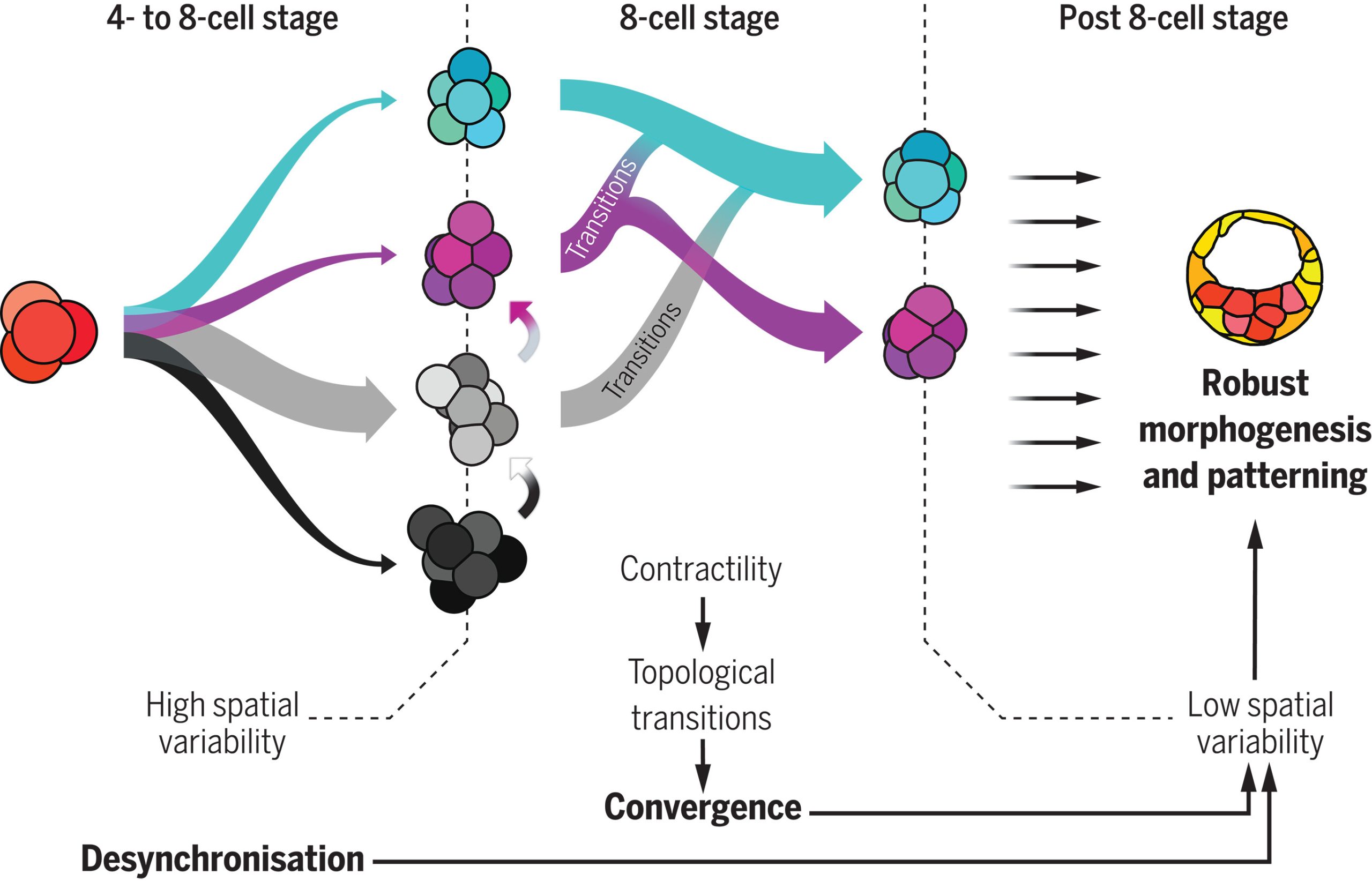
 (No Ratings Yet)
(No Ratings Yet)