In the latest issue, Disease Models & Mechanisms (DMM) launches a Special Collection that focuses on translational advances made using the Drosophila model, including insights into disease mechanisms, emerging technologies and therapeutic advances. This Collection, guest edited by Norbert Perrimon andNancy M. Bonini, includes a poster article on tools and assays for fly research (from Hugo Bellen’s lab), comprehensive Review articles, an update from Flybase, and original Research articles reporting new findings pertinent to understanding neurodegenerative diseases. An Editorial provides a summary of the Collection’s contents so far. The launch issue, which is dedicated to the memory of Marcos Vidal, can be accessed in full here.
Coming up soon is a Model for Life interview with Gerry Rubin, Review articles from the labs of Seung Kim and Heinrich Jasper, plus more original research.
Features and Reviews Editor, Journal of Cell Science
Based in Cambridge, UK
Applications are sought for the role of Features and Reviews Editor for Journal of Cell Science, our flagship journal serving the cell biology community.
Joining an experienced and successful team, including Academic Editor-in-Chief Michael Way, this is an exciting opportunity for a talented scientific editor to make a significant contribution to one of the major journals in the field of cell biology. Journal of Cell Science publishes outstanding primary research articles, reviews and topical comment, and continues to expand its services to authors and readers.
Applicants should have a PhD, and ideally postdoctoral experience, in cell biology or a relevant scientific field. The successful candidate will have strong communication, networking and interpersonal skills. We are also looking for enthusiasm, motivation, commitment, and a broad knowledge of and interest in cell biology and the scientific community. Editorial experience is preferable but not essential.
Core responsibilities include:
Commissioning, handling peer review and developmental editing of material for the front section of the journal
Writing content for the In This Issue section of the journal
Representation of the journal at local and international conferences and within the wider scientific community
Maintaining and developing the interviews section of the journal
Creative involvement in the journal’s development and marketing activities
Additional responsibilities may be available for the right candidate. The Features and Reviews Editor will work alongside both an experienced Executive Editor and Reviews Editor. This full-time position is offered for an initial 3-year term, and is based in The Company of Biologists’ attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Journal of Cell Science, Development, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV to hr@biologists.com quoting job reference JCSFRE0316, along with a 400-word summary of a recent ground-breaking development in cell biology, and a covering letter that states their current salary and explains their interest in the post.
Applications should be made as soon as possible and by April 8th, 2016.
Earlier this year we asked you to vote for your favourite image from a selection of 4 beautiful pictures taken by the students of the 2014 Woods Hole Embryology course. You chose a stunning image of a bat embryo, which features in the cover of the latest issue of Development.
It is now time to vote again, but this time we would like you to choose your favourite movie from the 4 below. The winning movie will feature on the homepage of Development, and a selected still will feature in the cover of the journal (click the links below each image to see what the cover would look like). The poll is set up to allow only one vote per person, so please stick to this rule to give all the images a fair chance!
Voting will close noon GMT on March the 30th.
Movie 1. Fly Embryo – Dorso-ventral Split
Drosophila melanogaster embryo stained for DAPI (blue, all nuclei), Elav (green, neural nuclei), anti-HRP (red, neural membranes and axons, and ring gland) and Fasciclin II (yellow, subset of CNS neuron cell bodies and motoneuron axons). Dorsal side at stage 16, ventral side at stage 14. Cover image. This movie was taken by Shane Jinson (MBL, USA) and Amber Famiglietti (Swarthmore, USA).
Movie 2.Fly Embryo – Sections
Drosophila melanogaster embryo, stage 16/17, stained for Tropomyosin (cyan, muscles), anti-HRP (magenta, neuron cell membranes and axons), Pax 3/7 (yellow, segmental patterns in ectoderm and CNS, MAb DP312), Engrailed (red, segmental patterns in ectoderm and CNS), and Twist (green, subset of neural and muscle nuclei). Imaged with a Zeiss LSM 780 Confocal. Cover image. This movie was taken by Carolyn Kaufman (Stowers Institute, USA).
Movie 3.Fly Eye Disk
Drosophila melanogaster third instar eye disk. DAPI (dark blue, nuclei), anti-HRP (magenta, photoreceptor cell bodies and axons), Elav (cyan, photoreceptor nuclei), Repo (yellow, glia cell nuclei), Expanded (white). Disk also contains Tie-Dye clones – EGFP (green), RFP (red), and LacZ (medium blue). The EGFP clone is in patches of photoreceptors, while the RFP and LacZ clones are largely overlapping in a patch of peripodial membrane (RFP being mostly a subset of the LacZ clone). Imaged with a Zeiss LSM 780 Confocal. Cover image. This movie was taken by Jiajie Xu (University of Chicago, USA).
Movie 4. Fly Embryo – Seven Channels
Drosophila melanogaster embryo, stage 17, ventral view. DAPI (blue, nuclei), Elav (green, neuronal nuclei), Spalt (yellow, subset of neuron and muscle nuclei), BP102 (red, CNS axons), Even-skipped (magenta, subset of CNS nuclei, and ring of nuclei around anal pad), and anti-HRP (gray, neuronal cell bodies and axons). DIC image also collected during the scan. Music begins about 40 seconds into the video. Imaged with a Zeiss LSM 780 Confocal. Cover image. This movie was taken by Connie Rich (University of Cambridge, UK).
Here are the highlights from the current issue of Development:
Mak(or)in’ the switch to adulthood
The juvenile-to-adult (J/A) transition of many animals, from worms to humans, is regulated by the highly conserved RNA-binding protein LIN-28. In this and other contexts – including stem cell renewal versus differentiation decisions – LIN-28 acts to suppress the production of the microRNA let-7, which in turn inhibits a suite of downstream genes, most notably the translational regulator LIN-41. Now, using the J/A transition of C. elegans as a model, David Fitch and colleagues (799) identify a new player in this axis, the Makorin orthologue LEP-2. lep-2 mutant adults display a number of juvenile characteristics, including failure of male tail tip retraction, continued moulting into adulthood and defective male sexual behaviour. The authors provide evidence that LEP-2 acts to promote degradation of LIN-28 in larval stages, which is necessary for the J/A transition. The underlying molecular mechanism has yet to be resolved: as a putative E3 ubiquitin ligase, LEP-2 might directly target LIN-28 for degradation or it may act indirectly. Given the conservation of the Makorin family, along with data implicating mammalian Makorins in cell state transitions and in the timing of puberty onset, it is possible that this Makorin/LIN-28 interaction could control developmental switches in multiple contexts.
Maintaining and reprogramming sexual identity
Gonadal sexual identity is determined during development, but must also be maintained in adulthood. In Drosophila, the transcription factor Chinmo has been identified as a key regulator of male identity in the adult testis: it promotes expression of the male sex determinant DsxM (the homologue of which, DMRT1, is a key regulator of male identity in mouse testes). In chinmo mutants, somatic stem cells of the adult testis adopt female fate. Now (754), Erika Matunis and co-workers show that Chinmo is not only necessary but also sufficient to promote male fate. Overexpression of chinmo in adult ovarian somatic cells leads to severe oogenesis phenotypes. Marker expression analysis suggests thatchinmo-overexpressing somatic cells lose female identity and gain male fate. Strikingly, this also appears to affect the sexual fate of the germ cells: a proportion of ovary germ cells start to express male markers upon somatic chinmo expression. Unlike in testis, Chinmo in the ovary does not appear to promote DsxM expression, implying that there must be alternative mechanisms for masculinisation of somatic cells. Although these mechanisms have yet to be uncovered, these data provide strong evidence that sexual identity of the Drosophila adult ovary can be reprogrammed, and that sexual fate must be actively maintained throughout life.
At the heart of histone methylation
Mutations in the histone methyltransferase KMT2D are associated with Kabuki Syndrome – a haploinsufficient congenital, multi-organ syndrome that frequently includes severe heart defects. However, the role of KMT2D in the heart has not been analysed in detail. Benoit Bruneau and colleagues now address this (810), generating several conditional Kmt2d mouse mutants and analysing them both phenotypically and using genome-wide approaches. They find that cardiac deletion of Kmt2d causes embryonic lethality with defects in heart morphology and cardiomyocyte proliferation. Global gene expression analysis demonstrates dysregulation of genes associated with cell cycle regulation, ion homeostasis and hypoxia signalling. Functionally, ventricular calcium handling appears impaired. KMT2D is involved in H3K4 mono- and di-methylation; consistent with this, ChIP-Seq data demonstrate that Kmt2d depletion causes loss of H3K4me1 and me2 at specific loci. By correlating these data with the RNA-seq profiles and ChIP-Exo data for KMT2D chromatin binding, the authors are able to identify a small number of high-confidence targets for KMT2D, functions of which are consistent with the phenotypes observed upon Kmt2d deletion. As well as shedding light on the important role of KMT2D in mouse heart development, these data may have implications for the aetiology of the heart defects observed in Kabuki syndrome.
TFAP2C: a key controller of placental growth
In mammals, proper placental development is essential for growth and viability of the embryo. The transcription factor TFAP2C is known to be important for specification and maintenance of trophoblast stem cells (placental progenitors), but whether this factor also plays roles at later stages of placental development is less well understood. On 787, Hubert Schorle and co-workers provide insights into the role of TFAP2C in a subset of placental progenitors, the TPBPA-expressing population that forms the junctional zone of the placenta. Loss of Tfap2c from this population leads to growth defects in the junctional zone, with reduced numbers of TPBPA+ cell-derived trophoblasts. Microarray analysis and follow-up experiments provide evidence that TFAP2C controls several key aspects of placental development: it inhibits Cdkn1a, a cell cycle inhibitor; promotes expression and activation of Akt to regulate glycogen synthesis; and promotes MAPK pathway activity – important for trophoblast proliferation and differentiation – by repressing the Dusp family of MAPK inhibitors. Importantly, this conditional mouse mutant provides a model for intrauterine growth retardation, as mutant embryos show lower foetal and birth weight. Preliminary data in a human trophoblast cell model suggests that this important role of TFAP2C may be conserved.
PLUS:
Building and re-building the heart by cardiomyocyte proliferation
Dissecting the cellular and molecular mechanisms that promote cardiomyocyte proliferation throughout life, deciphering why proliferative capacity normally dissipates in adult mammals and deriving means to boost this capacity, are primary goals in cardiovascular research. Here, Matthew Foglia and Kenneth Poss discuss the cellular and molecular mechanisms that control cardiomyocyte proliferation, during both heart development and regeneration across various species. See the Review article on p. 729
A comparative view of regenerative neurogenesis in vertebrates
Alessandro Alunni andLaure Bally-Cuif summarize the striking similarities in the essential molecular and cellular properties of adult neural stem cells between different vertebrate species, both under physiological and reparative conditions. They also discuss differences in the reparative process across evolution and how the study of non-mammalian models can provide insights into both basic neural stem cell properties and stimulatory cues shared between vertebrates. See the Review on p. 741
Featured movie
This movie shows the mating behaviour of wild type male C. elegans. This behaviour is disrupted in mutants of lep-2, a new heterochronic gene identified by Fitch and colleagues. Read the paper here.
DMM is looking for an enthusiastic intern who wishes to gain experience in science publishing.
Joining an experienced and successful team, including Academic Editor-in-Chief Monica Justice, the internship offers an ideal opportunity to gain in-depth experience on a growing Open Access journal in the exciting and fast-moving field of translational research. DMM publishes primary research articles and a well-regarded front section of reviews and topical comment. The intern will work alongside an established publishing team in our Cambridge offices.
Because the journal serves both basic biomedical researchers and clinicians, applicants will have a PhD or MD, ideally with some relevant research experience, and a broad knowledge of model organisms and disease issues.
The intern’s core responsibilities will include:
Representation of the journal at international scientific conferences, Company workshops and within the wider scientific community, with a view to promoting the journal and commissioning new front-section content.
Conducting interviews with high-profile scientists in the biomedical arena.
Support for the Academic Editors in their assessment and handling of primary research articles.
Outlining a strategy for journal social media activities and writing press releases.
Involvement in the journal’s development and marketing activities.
Contribute to special editorial projects on the journal.
Essential requirements for the job are enthusiasm, commitment, judgement, integrity and a mature attitude. Candidates should have excellent interpersonal skills and confidence, excellent oral and written communication skills, and a broad interest in research and the research community. They should also be willing to travel. Previous editorial experience is not required.
The internship will last for 9 months at a salary of £15,000 pro rata.
The Company of Biologists is based in attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Disease Models & Mechanisms, Development, Journal of Cell Science, Journal of Experimental Biology, and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
Applicants should send a CV by email to hr@biologists.com along with a covering letter that summarises their relevant skills and experience, and why they are enthusiastic about this opportunity. Candidates must be able to demonstrate their entitlement to work in the UK.
Applications should be made as soon as possible and by March 21st 2016.
– Two different applications of optogenetics were highlighted on the Node this month. Giorgia wrote about how optogenetics can be used to control morphogenesis in Drosophila. Meanwhile, Clare and Rachel described how they adapted the phytochrome system to live zebrafish embryos, allowing proteins to be moved within a live embryo using light!
– What is the relationship between ESCs and cells in the mouse embryo? And how does naïve pluripotency differ in mouse and primate species? Thorsten posted about his latest paper and introduced a mouse and marmoset pathway expression atlas for early development.
– Malkiel presented mouse-human neural crest chimeras, and the applications of this system for studying human neural crest development and disease in vivo.
This month saw two new additions to our ongoing series!
– Helena, who was doing her PIPS internship with us until recently, wrote about what it is like to do cell culture and work with ESCs.
– And the McGregor lab told us about what is like using house spiders to study developmental biology and development!
Discussion
– Have you ever deposited your paper in a pre print server like bioRxiv? What would persuade you to? Share your thoughts with the latest Question of the Month!
– Why are international collaborations important? James wrote about his visit to India to launch a new EMBO partnership.
– Two book reviews featured on the Node this month. Helena reviewed the book “Decoding the Language of Genetics” by David Botstein, while Heather considered the brave new world of human germline genetic modification by reading “GMO Sapiens” by Paul Knoepfler.
Would you like to run the Node?Last month we also announced that we are recruiting a new Node community manager! The deadline for applications is the 14th of March.
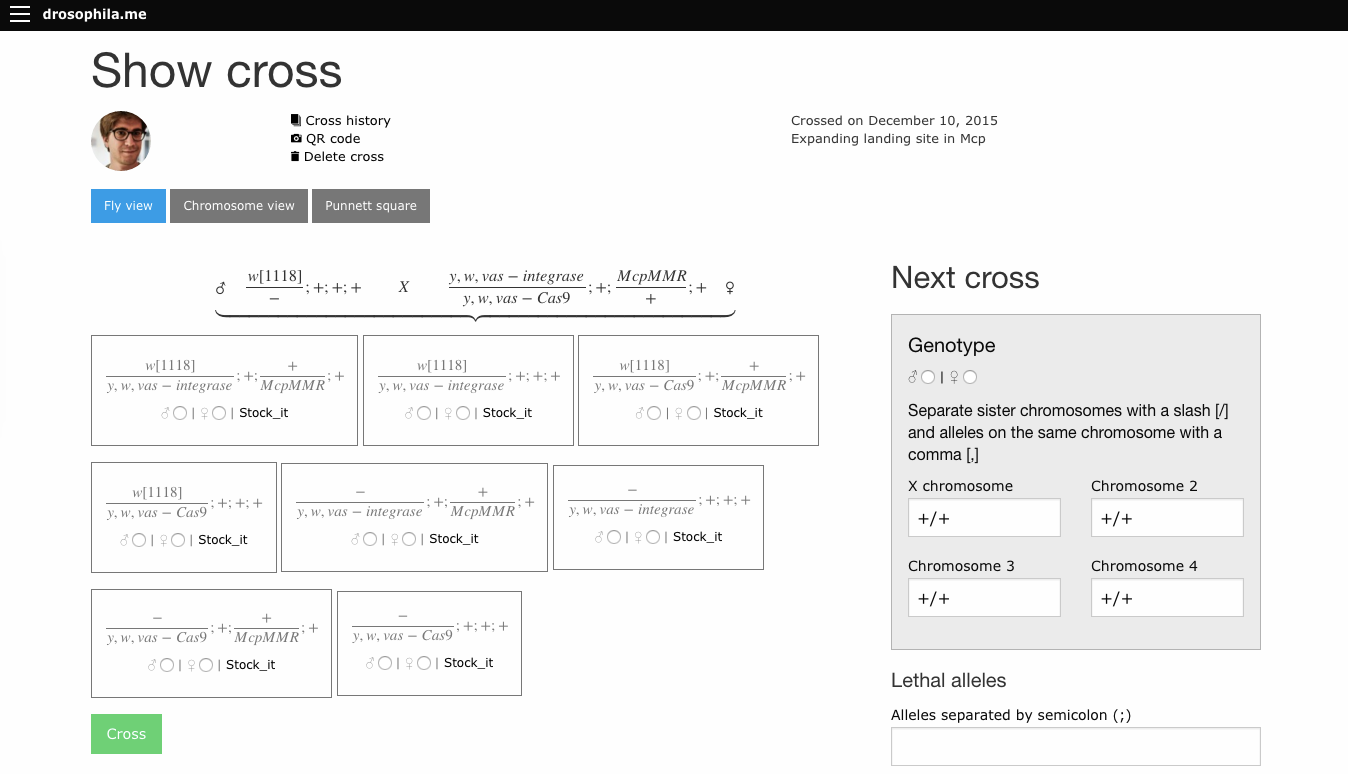
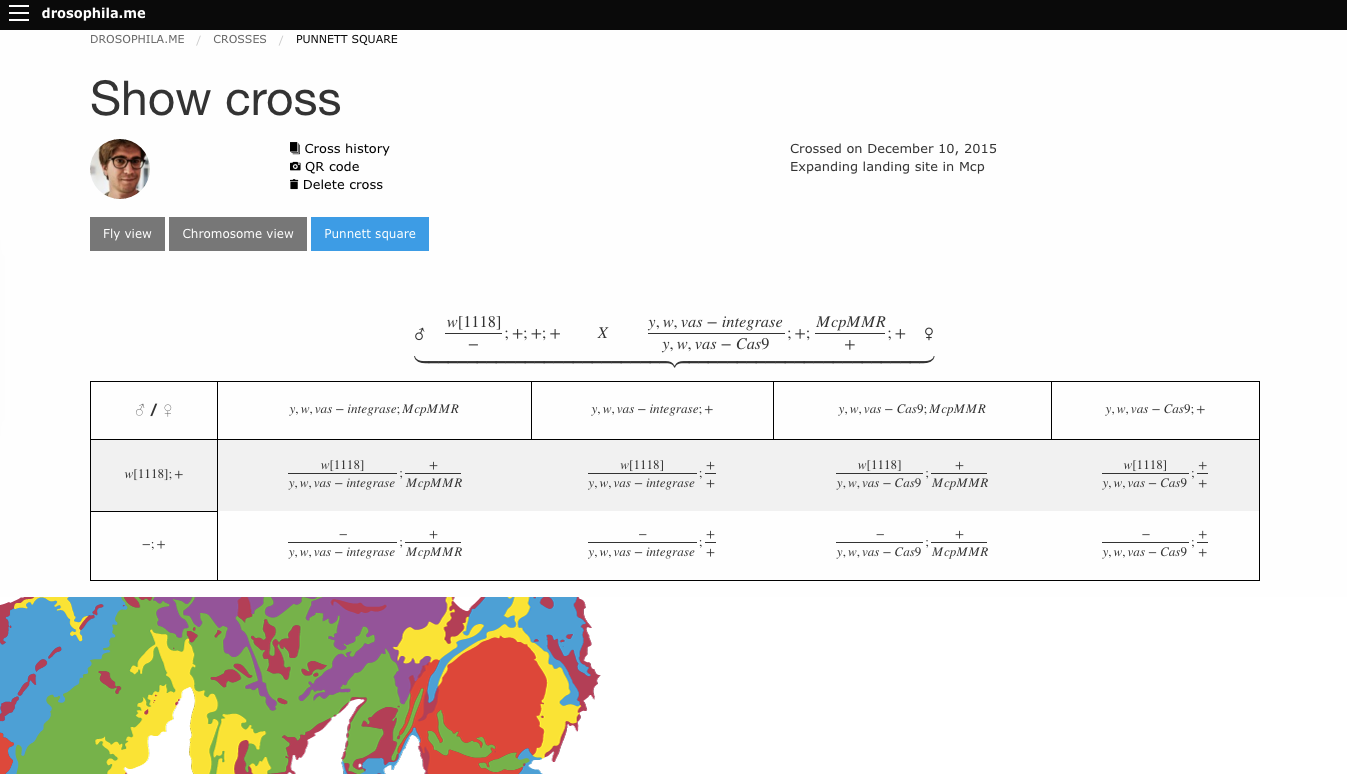
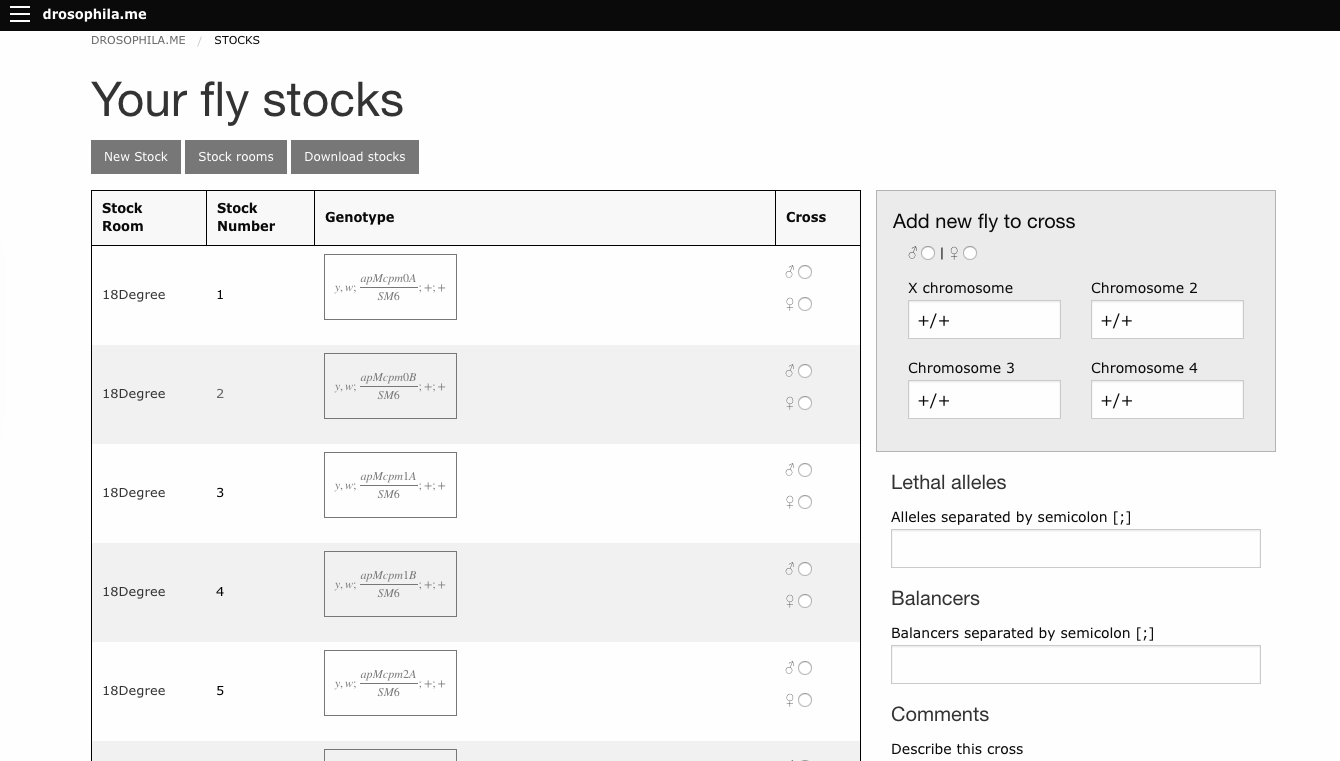
In that past post, I’ve promised a new version of the software, and I’m happy to announce it here.
The software can be reached under the address http://drosophila.me. Many many things changed from the previous version, some features:
User accounts
Your crosses are saved by default, you can use the software as a sort of digital genetic notebook
Crosses can be shared with colleagues (or with a qr code on a poster during a conference)
Cross history can be followed over generations
User can establish an own stock list and cross directly from it
Several visualisations of a cross: punnett square, genotypes, curly bracket view
Smartphone friendly
A web app as complex as drosophila.me is quite a code monster, and as I’m the only developer (for now), there might still be some glitches. I usually can solve them quickly, so let me know about it :)
I hope this tool will be useful for the community.
Places are still available for early career scientists interested in attending The Company of Biologists workshop ‘Metabolism in Development and Disease’. This is a unique opportunity to interact with the leaders in this field in a small and informal meeting. Apply until the 16th of March! For more information follow this link.
Professors Graham Williams and Duncan Bassett have 6 full-time posts (5 years each) and 2 shared appointments (with Dr Jacques Behmoaras, Medical Research Council, 4 years each) to fill. You will be joining an established laboratory with state-of-the-art equipment and a strong track record in molecular endocrinology. The lab is a founder member of the Origins of Bone and Cartilage Disease (OBCD) international phenotyping consortium. Please see our website for this exciting opportunity to join our cutting-edge research program based at Imperial College London.
The neural crest arises between neural and non-neural ectoderm and represents a somatic cell type with unique properties of multipotency. The neural crest cells (NCCs) migrate throughout the body and differentiate into a diverse array of cell types and tissues including the peripheral nervous system, enteric and sensory nervous system, Schwann cells, skin melanocytes, as well as connective tissues. The neural crest has major clinical relevance since it is disproportionately involved in both inherited and acquired developmental abnormalities termed neurocristopathies.
NCCs migration, development, and differentiation into various tissues have been studied in vivo in avian embryos, in studies pioneered by Nicole Le Douarin. Donor quail neural crest tissues were grafted into similar regions of stage-matched developing chick embryos, to generate quail-chick neural crest chimeras in ovo. The cells of the two species are easy to distinguish based on the ability to identify interphase nucleus of the quail, in which a large amount of heterochromatic DNA is found in the nucleoplasm, and different from that seen in chick where heterochromatin is dispersed within the nucleoplasm nucleolus. This chimeric system provided reliable information on the fate and ontogeny of the engrafted cells.
Few years later, Rudolf Jaenisch developed a mouse chimeric system that allows the assessment of the developmental potential and migration of mouse neural crest in vivo. Primary neural crest cells isolated from C57BL/6 mouse embryos and microinjected in utero into neurulating E8.5 albino BALB/c embryos were shown to contribute efficiently to pigmentation in the host animal. The resulting neural crest chimeras showed, however, different coat pigmentation contribution-patterns depending on the genotype of the host embryos. Whereas BALB/c neural crest chimeras showed limited donor cell pigment contribution, restricted largely to the head and limbs, KitW-sh/KitW-sh mutant mice used as hosts for chimeras, displayed extensive pigmentation throughout, often exceeding 50% of the coat. The KitW-sh/KitW-sh murine model carries an inversion that spans a 2.8 Mbp segment proximal to the c-Kit locus that disrupts its regulatory sequences and leads to a deficit of melanocytes, with no change in viability or fertility. This mouse line allows for an empty melanoblast niche in which the transplanted NCCs can incorporate without competition from the host endogenous cell populations. In contrast to BALB/c chimeras, where the donor melanoblasts appeared to have migrated primarily in the characteristic dorsoventral direction, in KitW-sh/KitW-sh mutants the injected cells appeared to migrate into the longitudinal direction as well, as if the cells were spreading through an empty niche. This is consistent with the absence of a functional endogenous melanoblast population in KitW-sh/KitW-sh mutants, in contrast to BALB/c mice, which contain a full complement of melanocytes.
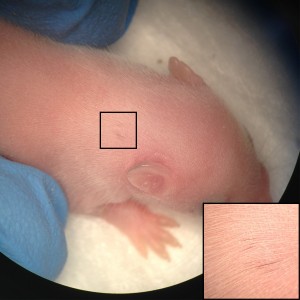
Human pluripotent stem cell-derived NCCs labeled with GFP migrate and contribute to the mouse embryo cranial region development at day E10.5. (Image by Malkiel Cohen)
The human induced pluripotent stem cell (hiPSCs) technology provides patient specific pluripotent cells that carry all genetic alterations that contributed to the disorder and thus represent a genetically defined cell system to study the respective disease. The greatest promise of hiPSCs is its potential to study human diseases in the Petri dish. In this approach patient-derived cells are differentiated into the cell types, which is affected in the patient with the goal to uncover a disease relevant phenotype in the dish. This method was also applied for modeling neurocristopathies such as Familial Dysautonomia, by differentiating patient-derived cells hiPSCs to neural crest and its derivatives, which presented disease manifestation in vitro. However, numerous human diseases originate already in embryogenesis, i.e. are caused by disturbances of developmental processes, therefore such an approach cannot recapitulate the developmental aspects of a disease because the test cells are not incorporated into the developing embryo and do not participate in normal developmental processes. Thus, a major challenge of the “disease in the dish” approach is establishing model systems that, using patient-derived hiPSCs, allow for the investigation of human disease under long-term in vivo conditions.
Inspired by the ability to generate neural crest chimeras, we aspired to use the human pluripotent stem cells as a cellular source of human NCCs and to generate mouse-human neural crest chimeras. This platform would expand the potential of using human pluripotent stem cells for studying human neural crest development and disease in vivo. In our recent paper we have shown that human NCCS derived from human pluripotent stem cells when microinjected into post-implantation KitW-sh/KitW-sh mouse embryos could participate in normal embryonic development and provide functional neural crest contribution to the host murine model. We tracked the implanted NCCs, which had been GFP labeled, thru their migration paths, and found that the human cells exhibited similar migration patterns as would normally be found in mice. As a result, about 30% of the implanted embryos showed human NCCs during development, and later in adult mice pigmentation, which is similar to what we found in mouse-mouse neural crest chimeras.
Human NCCs were microinjected into E8.5 mouse embryos to generate postnatal mouse–human neural crest chimeras. Coat color contribution in a mouse, injected as embryo with human NCCs, is indicated as single, dispersed, dark pigmented hairs (boxed area). (Images by Malkiel Cohen).
Our results are one of the first evidences of contribution of human embryonic cell population with functional evidence in adult postnatal mammal. Interestingly, we have observed that the largest the evolutionary distance between the NCCs and the host mice, the less the contribution is noticed. While a widespread pigmentation was observed using primary mouse NCCs as donor cells, only localized pigmented hairs were found in KitW-sh/KitW-sh host mice, using rat NCCs and minimal for in vitro derived human NCCs. The limited level of contribution we observed using human NCCs, presumably represents the over 90 million years evolutionary distance between mouse and human, suggesting that the host environment might limit the maturation and differentiation of the injected human NCCs.
Our work serves as proof of concept and an important first step toward the goal of generating chimeric mice that carry disease-relevant human cells in the relevant tissue. Resulting mouse-human chimeras would fill an important gap in disease research, as existing models do not accurately mimic certain diseases and disease states. Cancer is frequently studied using xenografts, however this approach fails to provide insight into tumor initiation and progression. Moreover, complex diseases with long latencies, such as Alzheimer’s and Parkinson’s disease, can only be partially modeled using induced pluripotent stem cells in vitro. Mouse-human chimeras would be used to overcome these limitations and could be used for regenerative medicine as well.
 Nancy M. Bonini, includes a poster article on tools and assays for fly research (from Hugo Bellen’s lab), comprehensive Review articles, an update from Flybase, and original Research articles reporting new findings pertinent to understanding neurodegenerative diseases. An Editorial provides a summary of the Collection’s contents so far. The launch issue, which is dedicated to the memory of Marcos Vidal, can be accessed in full here.
Nancy M. Bonini, includes a poster article on tools and assays for fly research (from Hugo Bellen’s lab), comprehensive Review articles, an update from Flybase, and original Research articles reporting new findings pertinent to understanding neurodegenerative diseases. An Editorial provides a summary of the Collection’s contents so far. The launch issue, which is dedicated to the memory of Marcos Vidal, can be accessed in full here. (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)




 Dissecting the cellular and molecular mechanisms that promote cardiomyocyte proliferation throughout life, deciphering why proliferative capacity normally dissipates in adult mammals and deriving means to boost this capacity, are primary goals in cardiovascular research. Here,
Dissecting the cellular and molecular mechanisms that promote cardiomyocyte proliferation throughout life, deciphering why proliferative capacity normally dissipates in adult mammals and deriving means to boost this capacity, are primary goals in cardiovascular research. Here,  Alessandro Alunni and
Alessandro Alunni and – Two different applications of optogenetics were highlighted on the Node this month. Giorgia wrote about how optogenetics can be used to
– Two different applications of optogenetics were highlighted on the Node this month. Giorgia wrote about how optogenetics can be used to 
 – Have you ever deposited your paper in a pre print server like bioRxiv? What would persuade you to? Share your thoughts with the latest
– Have you ever deposited your paper in a pre print server like bioRxiv? What would persuade you to? Share your thoughts with the latest  – What’s the
– What’s the 


 (11 votes)
(11 votes)