A postdoctoral Position is open in the lab of “Imaging and Regulation of Morphogenesis in Higher Vertebrates” at the Pasteur Institute in Paris, France. Our lab is interested in understanding morphogenesis of developing structures, at a cellular level. Using avian models we combine state-of-the-art live imaging microscopy, quantitative analyses, biophysical, cellular and molecular biology approaches to access the cellular dynamics of development.
This specific project aims at elucidating the cellular events underlying the initiation of limb bud formation and how such cell events are dynamically regulated at the molecular level, using the generation of transgenic avian lines and live imaging methodologies. For more information about projects and the lab please visit:
The position is a 4-year postdoctoral position funded by the ERC (European Research Council), available immediately. We are seeking highly motivated candidates with expertise in developmental and/or cellular biology. Experience in imaging and chick development will be positively considered.
The Pasteur Institute, located in the vibrant city of Paris, has a longstanding history of excellence in developmental biology and in science in general, with access to excellent core facilities.
Applicants should send a cover letter (describing briefly research interests), a C.V and contact information for up to 3 academic references to jgros@pasteur.fr.
My name is Tyler Square, and I am a PhD student in the Medeiros Lab at the University of Colorado at Boulder in the Ecology and Evolutionary Biology department. Also in the Medeiros lab is David Jandzik, a postdoc who was kind enough to provide most of the media for this post. Our lab primarily studies the evolution of development (evo-devo) in vertebrates, with a focus on structures and developmental processes in the head. I personally focus on the evolution of neural crest cells, which are a really cool (and really weird) cell type that contributes to a whole slew of diverse structures in vertebrates: neurons and glia of the peripheral nervous system, cartilage and bone of the head skeleton, smooth muscle of the heart, pigment cells, and more!
Tyler Square (left) demonstrating the suction power of a large adult.
David Jandzik with an adult lamprey
In order to have a complete view of vertebrate evolution, it is important to use a diverse sampling of vertebrates. This helps us infer when certain developmental processes evolved, or how particular lineages might have undergone specialization. To these ends, our lab uses zebrafish, Xenopus laevis, and the sea lamprey (Petromyzon marinus) as vertebrate models, and amphioxus (Branchiostoma floridae) as an invertebrate chordate outgroup (closely related to vertebrates).
Lampreys are surely our most charismatic model organism, and they are phylogenetically positioned in a very favorable spot for our purposes. Much work has already been done in jawed vertebrate (gnathostome) models like zebrafish, Xenopus, chicken, and mouse, so by understanding lamprey development we can get a more complete picture of how vertebrate development evolved. Sea lampreys are part of a vertebrate group called the cyclostomes (which comprise only lampreys and hagfish) that are well-supported as the sister group to gnathostomes; thus, all lamprey and hagfish are the vertebrate animals most distantly related to you!
A composite image showing different phases of the lamprey life cycle. The picture of the embryo (left) is stained by in situ hybridization for a gene marking migratory neural crest cells.
This lamprey is approximately one month old, and will soon run out of yolk and must rely on filter feeding.
Sea lampreys have a rather complex life history, and it helps to be familiar with this since it dictates how we are able to use them in a lab setting. Some aspects of their biology make them ideal for use as a developmental model, whereas other aspects are far from ideal. Their entire lifecycle takes somewhere between 5 and 11 years to complete. Most of this uncertainty comes from the duration of their filter feeding larval stage, called an ammocete. Once hatched, these small (~1 cm) ammocete larvae enter a blind, mostly burrowed phase of suction feeding on detritus and algae that lasts somewhere between 3 and 10 years depending on the locality. Metamorphasis occurs after this filter feeding stage when the lamprey are ~12 cm, which is when the animals develop eyes and teeth. These now parasitic juveniles swim out to a nearby ocean or lake and look for a series of hosts for 12-20 months of bloodmeals. By feeding on a series of hosts, they attain a wide range of final adult sizes from as small as 35 cm to as large as 90 cm. This range again reflects locality: populations confined to lakes tend to be smaller on average, while individuals in those populations connected to the ocean can attain much larger sizes. Since our breeding phase adults come almost exclusively from the Great Lakes, the majority of the adults in our lab are between 50 and 60 cm. Sea lampreys then enter their adult breeding phase lasting a few months where they completely stop eating, swim back upstream to build a nest, and spawn. Adult sea lampreys die usually within a week of spawning.
Due to the length of this lifecycle and the micropredatory feeding strategy during the juvenile stage, no laboratory or marine facility has ever housed a complete sea lamprey lifecycle. This means that no inbred or stable transgenic lines of sea lamprey (or any cyclostome, for that matter) exist anywhere, making some of the fancy tools developed in more common developmental models like zebrafish or mouse out of reach.
Group photo of a few adults in one of our holding tanks. Pictured here are just six, but these tanks can hold up to 18 adults.
Most labs like ours that study sea lamprey embryology get shipments of adult spawning phase lamprey starting in mid-June, which were trapped in streams. Most US labs, including ours, get lampreys from the Hammond Bay Biological Station in Millersburg, MI. We hold these animals in large tanks with an ample rate of water turnover, as the animals still exude waste despite that fact that they are no longer eating (and won’t eat again before they naturally die in our hands). Our lab is usually home to around 50 adults at any given time, split into three ~350 liter tanks. We like to have slightly more than half of our animals be females, but we don’t always have much of a choice. Usually by mid- to late August, all of the adults that came through our lab have completed their lifecycle, and have perished naturally.
Around half of the days during the ~10 week “lamprey season” are injection days. To get sea lamprey zygotes, gametes are manually stripped from the live adults and put on glass dishes with a little bit of system water in them. At any given time during lamprey season we typically have up to three females in their prime egg-laying stage while we wait for the others to ripen. So on a normal day in the height of the season, a typical fertilization bout will use one or two proven females to make a few big, injectable batches of eggs (~3,000), while also doing some small test fertilizations of another few females who might be ripe (~200 eggs). Some females never give us good eggs, and will instead tempt us with eggs that don’t fertilize, or ones that do but the resulting embryos most frequently die at day three or four (during or just after gastrulation). Some females will never even give us eggs at all before they die. Other females will give us 30,000-50,000 good eggs over the course of their two to five day prime. But even with these optimal females, eggs lain before or after the viable eggs will sometimes be poor. The males tend to be easier to work with, and can also last for much longer in their prime breeding stage; some males have lasted up to three weeks, cementing their legacy in our lab as the sire to more than 100,000 embryos! Once we have a good, proven male, we tend to use him alone to perform fertilizations, but before proving out a male we will combine the sperm of three or four males during one fertilization event (since sometimes males shed only very small amounts of sperm).
Movie 1: Fertilization. A short video on how we manually strip gametes from males
and females
Daily tasks during the lamprey season are split between the adult care, which is minimal, and the embryo care, which can be rather time consuming. The adults require daily checks, as dead animals need to be collected soon after they perish as to prevent system contamination. We also do ammonia tests periodically to ensure that the water changes are sufficiently removing waste from the holding tanks.
A close-up of our lamprey system. Pictured here are dishes holding ~1000 embryos.
The embryos command the majority of our attention. On a typical day with a good, fertile female, we spend four to six person hours injecting embryos, and two to eight person hours ‘sorting’ the previously-injected and wildtype embryos. We sort the embryos both to remove dead material, and also under a fluorescent microscope to select those individuals exhibiting visible lineage tracer, indicating that they were successfully injected. We usually inject up to 30-40 µl of injection solution into 6,000-8,000 zygotes on a good injection day.
Movie 2: Microinjection. A video showing the setup for injecting lamprey zygotes.
As you might imagine, working with lamprey is a dream when the eggs and embryos are healthy. In one sitting using CRISPR we can potentially generate hundreds or even thousands of sea lamprey that are mutant for a given gene of interest. If survival is good, this becomes enough material for both in situ hybridization (ISH) on multiple genes at multiple stages of development, and genomic DNA from presumed mutants for analysis of the targeted loci. However, our most recent 2015 season was a better example of the converse egg quality situation: while we did eventually have some successful rounds of injection, the first month or so of lamprey season was full of females giving us eggs that were almost unanimously destined for death on day four. This meant many, many hours injecting and sorting embryos that yielded little or no useable data. It’s not unusual for us to happen upon a female with poor egg quality every now and then even during a good season, but to spend so many hours under a microscope every day for weeks with nearly nothing to show for it can be demoralizing at times. But despite these high-stakes prospects, even just one or two good injection days can generate enough data to conduct a thorough study on the role of a given gene in sea lamprey development. So while it might be a bummer to look back on many weeks of time and supplies that yielded practically nothing, it can all be worth it for those few good days wherein enough material is generated to keep our lab busy for the rest of the year.
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series here and read other posts in this series here.
The Max Planck Institute for Molecular Biomedicine in Muenster, Germany has an opening for a
Postdoctoral Scientist
(position-code 19-2015)
The position is available in the DFG Emmy Noether junior group of Dr. Ivan Bedzhov that is focused on understanding the self-organization of the pluripotent lineage in mammalian embryos at the time of implantation (I. Bedzhov and M. Zernicka-Goetz, Cell, 2014). The successful candidate will investigate the mechanisms of self-organization of the pluripotent epiblast in the context of the developing embryo and in vitro using embryonic stem cell as a model system.
We are looking for a talented and highly motivated post-doctoral scientist. Previous research background in embryonic stem cells pluripotency or epithelial polarity, experience in in vitro systems of epithelial morphogenesis, gene-targeting and genome-wide data analysis will be an advantage.
The Max Planck Institute for Molecular Biomedicine offers dynamic, multidisciplinary environment with state-of-the-art transgenic, imaging, genomics and proteomics equipment and core facilities. The working language in the institute is English.
The institute is located in Muenster, a vibrant city with a highly international academic environment.
Starting date will be as soon as possible. All conditions for the employment will be according to the regulations of the contracts for the civil service (TVöD, Tarifvertrag für den öffentlichen Dienst) level 13 TVöD.
The Max Planck Society seeks to increase the number of women in those areas where they are underrepresented and therefore explicitly encourages women to apply.
The Max Planck Society is committed to employing more handicapped individuals and especially encourages them to apply.
Please send your application (with the position-code 19-2015), letter of motivation, CV including a complete list of publications and the contact information of 2 referees to:
The festive season is here, so it’s time to start thinking about gifts. Many of my friends are scientists, so I have been searching for science-themed gifts. I thought I would share some of my finds here on the Node, in case you are searching for something similar (or would like a science-themed gift yourself!)! Below are some of my finds. If you have any other good ideas feel free to leave a comment!
Biscuit cutters
Does your lab have the tradition of bringing cakes to lab meetings? Or maybe your department has a monthly happy hour? Give these cookie cutters to a friend or colleague and maybe you’ll benefit from this gift too! There are many other biscuit cutter options on this shop.
Science scarves
I spotted these beautiful silk scarves in the Artologica shop on Etsy (you can also follow the artist on twitter- @artologica). Check out their shop for other sciency gifts, including petri dish ornaments that would look great in any lab (or personal) Christmas tree!
Brain cells scarf
Mitosis scarf
Cuddly science
Giant microbes started off by creating cuddly versions of disease-causing microbes, but have since branched off to other types of cells and even a cuddly Drosophila!
Science jewellery
There is a lot of science-themed jewellery out there. The Vexed Muddler is a good place to start your search, with beautiful ceramic science jewellery such as these cool bracelets.
Embroidered notebooks
Some of these may be too fancy to be used as lab books, but they make a very special gift! Check out the Fabulous Cat Papers shop on Etsy for more designs.
Embryo pillow
A hard-working scientist needs a rest after a long day in the lab. So what best than a sciency pillow? The design of this pillow is particularly appropriate for developmental biologists, displaying an embryogenesis-inspired pattern. Check out Klara Acel’s shop if you like the design but would prefer it as a bag, rug, or a matching duvet cover!
A gift for a little scientist
It is never too early to start learning about cell division, right? Check out the shop Cognitive Surplus for more great sciency gift ideas, including DNA replication champagne glasses and a tardigrade tote bag! When I contacted the shop owners they told me they are keen to get ideas for new science-themed products, so get in touch with them if you feel inspired!
Developmental biology t-shirt
I spotted this t-shirt showcasing Haeckel’s embryos in the nonfinctiontees shop. They have a lot of other science-themed t-shirts to choose from!
Cute stocking fillers
There is always the need for small gifts this time of the year. How about some cute science badges? More to choose from at the Roocharms shop.
Do you have any other ideas for great science-themed gifts? Leave your suggestion as a comment!
In 2014, the British Society of Developmental Biology (BSDB) has initiated the Gurdon Summer Studentship program with the intention to provide highly motivated students with exceptional qualities and a strong interest in Developmental Biology an opportunity to engage in practical research. Each year, 10 successful applicants spend 8 weeks in the research laboratories of their choices, and the feedback we receive is outstanding. Please, read the student report, kindly sent to us by Isabella Watts(Oxford University) who was hosted in summer 2015 by Tatjana Sauka-Spengler at the Weatherall Institute of Molecular Medicine in Oxford..
A transcriptional analysis of the innate immune response to melanoma: the promise of the zebrafish
This summer I was privileged enough to receive a Gurdon Summer Studentship to work for 8 weeks in the laboratory of Tatjana Sauka-Spengler, at the Weatherall Institute of Molecular medicine. About to go into my third year of studying medicine at Oxford, I have spent the majority of the first two years of my degree learning about the science that underpins medicine. All our lecturers highlight the importance of understanding the scientific evidence that underlies the concepts that we learn. However, no matter how much time you spend trawling through the depths of PubMed to find a new and exciting paper for an essay, it’s not the same as getting hands on experience in a laboratory and learning about different scientific techniques.
My project involved helping my supervisor Amy Kenyon with the final steps of her PhD. Amy had generated a transgenic zebrafish model to study the innate immune response under conditions of inflammation, including tuberculosis and melanoma. This model system harnesses developmental biology and basic science to provide an invaluable tool to better understand the mechanisms of different disease states.
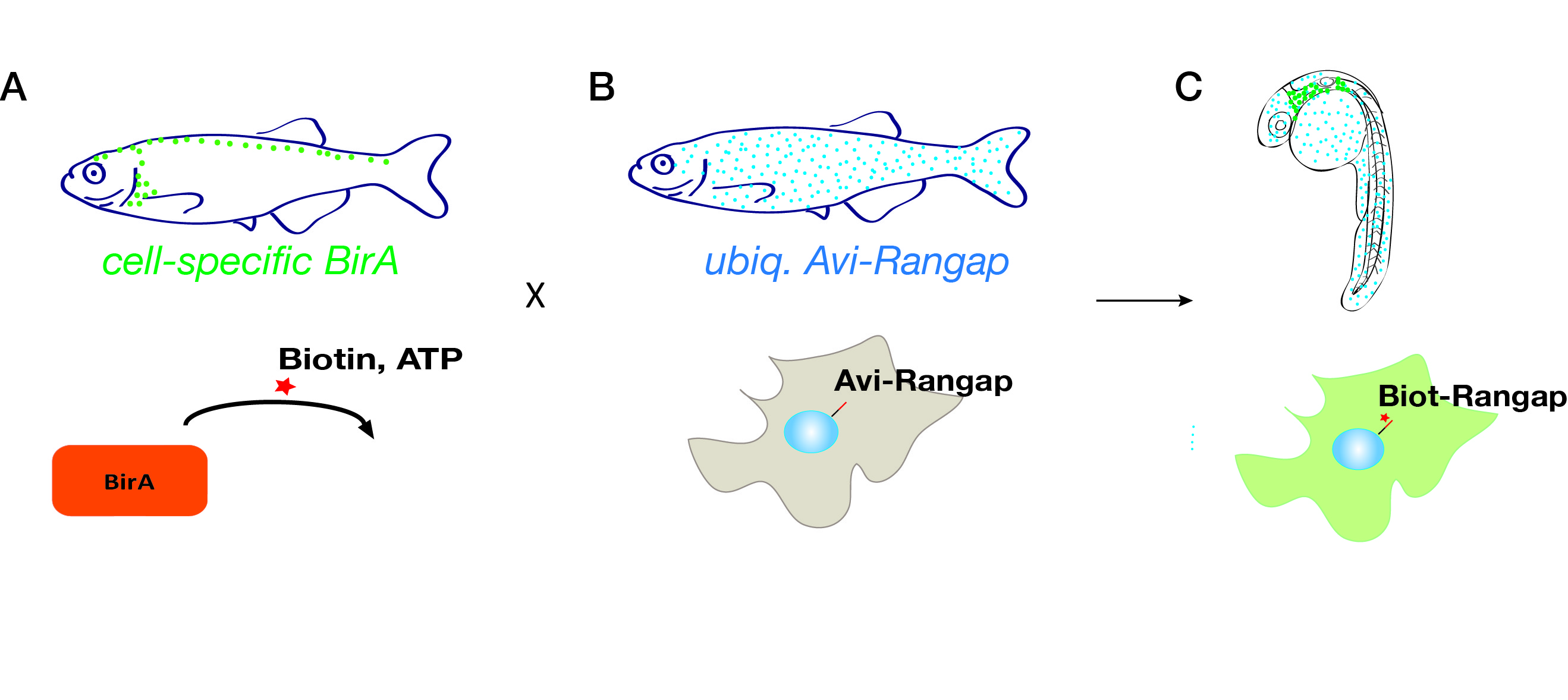
I was mainly focussing on the innate immune response to melanoma, which is an incredibly exciting area of research as macrophages and neutrophils form a major part of the tumour microenvironment. These cells have diverse roles in tumours and can both destroy transformed cells and also contribute to tumour development. However, it has proven difficult to fully examine the interactions of these cells and transformed cells in vivo due to limitations in current model systems. Amy developed a unique model system, pioneered in vivo by the Sauka-Spengler laboratory, which allows isolation of specific cell types and cellular compartments directly from the organism, via a genetically encoded, tissue-specific biotin tagging system.
Fig. 1. A genetically encoded, tissue-specific biotin tagging system.
The system involves fish that ubiquitously express the nuclear envelope-associated protein Rangap fused to an Avi-tag (Avi-Rangap). These have then been crossed with a separate transgenic line where E.coli biotin ligase (BirA) is selectively expressed under control of the mpeg or mpo promoter which are specifically active in macrophages or neutrophils, respectively. Therefore, in the macrophages or neutrophils, biotin is covalently added to the Avi-tag Rangap protein, resulting in the ‘biotinylation’ and tagging of the nuclei of these cells, which can then be isolated using highly stringent streptavidin-biotin affinity purification protocol.
To generate an inducible melanoma model, another binary system of transgenic fish has been used. In the driver line, the bacterial LexPR repressor is fused to a truncated form of the human progesterone receptor, all under the control of a melanocyte specific promoter. The effector line harbours the Lex Operon, controlling the expression of a Ras-oncogene fused to an mCherry fluorescent reporter. When the synthetic steroid mifepristone is added to the water of embryos carrying both alleles, it binds to the LexPR transactivator which, in turn, binds to the Lex Operon region and drives expression of Ras-oncogene specifically in melanocytes, resulting in their transformation and proliferation.
From the very first day of my project I learned a huge amount, and the learning curve was definitely vertical at times! I was incredibly lucky to be working on a very broad topic which meant that I was able to try my hand at a huge number of different techniques.
Some of the earliest work I did involved confirmation of the different model lines I was using. In my first experiment, I wanted to check whether the melanoma model we were using effectively mimicked the cancer over-proliferation phenotype. In the zebrafish embryo melanocytes are usually post-mitotic at 60 hours post fertilisation, thus I wanted to confirm that induction of our cancer system resulted in continued cell proliferation past this point. We injected EdU, a thymidine analogue incorporated into the DNA of dividing cells, into the pericardium of day 4 embryos, in order to label proliferating cells. Upon imaging we saw that, in fish where the LexPR system was activated, there was continued proliferation of melanocytes whilst in control fish there was no overlap between the melanocytes and the stain that labelled the dividing cells.
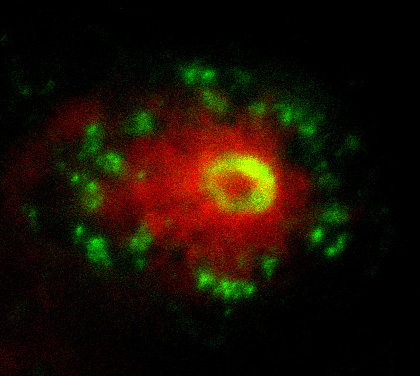
Fig. 2. This image shows the early interaction between the neutrophils (green) and a transformed melanocyte (red).
In the next step of my project I made use of the transparency of the early zebrafish embryo to image the live interaction between the neutrophils, macrophages and transformed melanocytes (Fig. 2). The final step of my project was a transcriptional analysis of the interactions between the innate immune cells and tumour cells. Collecting day 5 embryos where we had induced the oncogenic transformation in melanocytes, we selectively isolated the nuclei of macrophages and neutrophils from the fish using the streptavidin-biotin affinity purification previously mentioned. Whilst incredibly rewarding, this did involve spending entire days in the cold room, and I got a lot of strange looks as I carried 3 jumpers into work in the blazing sun and even drank hot chocolate in the institute’s cafeteria on the hottest day of the year! After isolating the RNA from the nuclei and making cDNA libraries, the data was sent off for sequencing and we were able to see which genes were up- and down-regulated in these cells in response to the cancer.
Looking back at my time in the lab I can’t quite believe how much I have learned! I have had an amazing time and every week has been packed with new things. I loved getting to learn more about the processes involved in laboratory science and I have been convinced that in the future I want to combine both clinical work and research. This has been a very valuable experience and I would really encourage other university students to apply for a Gurdon Summer Studentship. I also want to say a huge thank you to everyone in my lab, especially Tatjana and Amy for making my summer so interesting!
The Department of Neural Development and Physiology at Institute of Cell Physiology at the National University of Mexico (www.ifc.unam.mx), is seeking for outstanding candidates to apply for a tenure track position equivalent to Assistant Professor. Application deadline: February 29th , 2016
Candidates must fulfill the following requirements:
Ph.D. degree.
Three years (minimum) of postdoctoral experience.
Excellent academic and publication records (at least 8-10 publications in prestigious journals), showing a record of scientific contributions in areas related to neural development and physiology.
The proposal should include multidisciplinary experimental approaches and the use of cutting edge techniques.
To be under 37 years old for men and 39 years old for women at the time of the official incorporation date.
It is desirable that the candidates are eligible or awardees of grants to establish their own independent laboratory.
Duties:
To lead a research group with an innovative and independent project.
To be eager to integrate into a multidisciplinary team and collaborate.
To obtain external financial support for his/her research.
To train young researchers at bachelor and graduate levels.
To communicate his/her discoveries to the society.
To support the development of the institution.
The Institute will provide:
Laboratory space
Access to institutional equipment and service units.
Salary (depending on applicant qualifications).
Submit your application before February 29th, 2016 to secdir@ifc.unam.mx and addressed to Dr. Marcia Hiriart Urdanivia Director of the Institute of Cell Physiology, including:
Cover letter describing research experience and interests, a list of all publications highlighting the most relevant ones with a brief explanation of their importance and PDF of all publications.
Curriculum vitae
Three letters of reference or contacts of professionals providing reference letters.
Research plan (no more than 10 pages)
A copy of the full application should be submitted to seacad@ifc.unam.mx
Selection: a Selection Committee will evaluate Applicants. On April 1st, 2016, selected candidates will be asked to give a seminar at the Institute and will be interviewed by faculty members, any time between April 15th and May 20th, 2016.
Selection announcement: not later than June 14th, 2016.
Position available to start: Between August 16th and September 16th, 2016.
Our group has an opening for a postdoctoral fellow to work on NIH-funded projects involving FGF signaling in cochlear development, with a focus on auditory supporting and sensory cell differentiation in the mouse model. Candidates should have >2 years of graduate or postdoctoral research experience in inner ear development, including anatomic, molecular and functional analyses, with strong publications (can be pending) in these areas. Individuals with expertise in confocal microscopy of other developing and mature sensory systems with an interest in studying the ear are also encouraged to apply. Experience with mouse genetics is not absolutely required, but is a big plus. The stipend will follow NIH guidelines. Benefits include health and dental insurance. The successful candidate will be mentored through the process of applying within two years of the initial appointment for individual support (for stipend and/or newly developed project) as appropriate. Presentation and networking opportunities include a weekly Human Genetics Research-in-Progress series and a monthly meeting of the Inner Ear Research Group. There will also be an opportunity to attend at least one appropriate scientific conference per year.
If interested, please contact suzi.mansour@genetics.utah.edu and provide a very brief (<1 page) current research description, a Biosketch, and the names and contact information of two individuals (current PI and one other), who can be contacted to comment on your suitability for this position. The position can start as early as Jan. 4, 2016, but other start dates are negotiable.
In 2014, the British Society of Developmental Biology (BSDB) has initiated the Gurdon Summer Studentship program with the intention to provide highly motivated students with exceptional qualities and a strong interest in Developmental Biology an opportunity to engage in practical research. Each year, 10 successful applicants spend 8 weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Please, read the student report which was kindly sent to us by Aran Shaunak. Arun studies at Cambridge University and was hosted in summer 2015 by Jose Silva at the Cambridge Stem Cell Institute. During hisproject, entitled “The molecular mechanisms of Oct4”, Arun aimed to create new cell lines for the study of Oct4 and optimise a protocol for P-STAT3 western blots to test its functional involvement. Arun’s report does not focus on the actual project but rather presents an entertaining opinion piece about the emotional roller coaster of his project, which provides good insights into the realities of laboratory life.
What every potential academic needs to learn about science
Six days into my first real taste of scientific academia, I found myself in the tissue culture room at 9.15 on a Saturday morning. I had been the willing recipient of my own batch of stem cells, which I was charged with growing up, looking after and eventually experimenting upon to hopefully provide data of some use to my PhD supervisor. Excited to be flying solo already, my cell line was given the 5 star treatment.
Apparently stem cells are so demanding to grow that they often require feeding every day once their flask is well populated, meaning that I had no choice but to come in on my supposed weekend to give them their meal of serum and LIF. And so initially I was thoroughly disillusioned with the glamorous cutting-edge-of-science academic lifestyle which seems to be the usual interpretation of “I work in a stem cell lab”; to me, it just seemed like I had unsuspectingly taken on millions of pets that were higher maintenance than my ex-girlfriend and more productive than teenage rabbits.
However, everything changed over the course of the next week. Working closely with my supervisor, a time-course experiment done on those very cells which had consumed my weekend resulted in the production of a gel and a western blot. The lab were sceptical; people had spent months attempting to get out a clean western for the protein we were looking at, all to no avail, hence why a large part of my 6 week project was expected to be taken up with western after western after western. But lo and behold, a protocol based on ‘estimated’ dilutions and getting bored before the timer went off produced a near-perfect western blot – a testament to the experience of my supervisor. Furthermore, there was actually an interesting result there too, one which we had hoped to see (although, obviously, no-one believed it until we could show them the loading control – as a born and bred cynic I feel science may be the perfect career path after all).
Thus the majority of my planned placement was completed in the first 10 days. And as I fed my cells yet again I realised something had changed. It wasn’t a chore to change their media, or passage them into a new flask to give them room to grow. I wanted to freeze down stocks, so that if disaster struck I wouldn’t lose them. The result of our experiment had both given me a taste of the success that an academic feels when they finally look down the microscope and see that they were right all along, and earned me the respect of my older, more experienced colleagues. And I have come to realise that I had attributed some of that success to the cells themselves; that they had earned the time and trouble it took me to look after them.
Real life hit like a hammer in the second half of my placement, when a year’s worth of work disintegrated in front of my supervisor’s eyes, nearly forcing us to resort to the emergency gin tucked away in the second drawer. Cloning experiments that were going beautifully suddenly collapsed and we were forced to begin again from scratch. I finally felt the disappointment that comes from unexplained failure, and found true respect for the resilience of your average Joe PhD; after a day, week, month or year of two-steps-forward-one-step-back, they still come into the lab with the idea that just maybe today is the day for a breakthrough. But I also felt the pride that comes with success; a week of western blot optimization resulted in the development of a protocol that my lab will use in order to quantify levels of pSTAT3 in stem cell experiments, something that had previously been impossible to reliably achieve.
However, it seems to me that my first ever cell line was responsible for my fundamental change of opinion on academic life. I realised that scientists don’t spend weekends in the lab and nights with a pad of paper by their bed because they have to. The life isn’t as glamorous as it sounds: stem cells may not be growing people new kidneys in time for Christmas this year. We do it because the cells become yours, and you care about them ; your results become your badge of honour, and you show them off with pride; and most importantly, you absolutely, definitely will find out how Oct4 functions on a molecular level, and you’re damn well going to do it before anyone else.
Disease Models & Mechanisms invites you to submit original research for consideration for an upcoming Special Collection named Spotlight on Rat: Translational Impact scheduled for publication in autumn 2016. This ongoing collection* will focus on translational advances made using Rat as a model organism, including insights into disease mechanisms and therapeutic targets, new resources and technologies, and drug discovery and development.
We invite you to showcase your breakthrough Rat research in this Special Collection. Submissions should describe original research in the form of a Research or Resource article. Please read the author guidelines for information on preparing a manuscript for DMM, and submit your manuscript via our online submission system. Please highlight that your submission is to be considered for the Special Collection in your cover letter. For rapid feedback on the suitability of a paper for inclusion in the Special Collection, please send us a presubmission enquiry. The Collection will also contain invited Review articles covering topics of broad interest to the Rat community and interviews with leading scientists in the field. The launch issue of the Special Collection will be widely marketed and will be distributed at relevant conferences worldwide, providing prominent exposure for your work.
Submission deadline: 1st March 2016
Key benefits of publishing in DMM include:
High visibility and impact (2014 Impact Factor 5)
Open Access (CC-BY licence) and PMC deposition
Rapid peer review and publication
Indexed in Medline, ISI and Scopus
Not-for-profit publisher
*Please note that not all papers accepted for publication after peer review will be included in the launch issue for the Special Collection; some will be published in later issues and added to the compiled online Special Collection. Contact us for further details.
In 2014, the British Society of Developmental Biology (BSDB) has initiated the Gurdon Summer Studentship program with the intention to provide highly motivated students with exceptional qualities and a strong interest in Developmental Biology an opportunity to engage in practical research. Each year, 10 successful applicants spend 8 weeks in the research laboratories of their choices, and the feedback we receive is outstanding. Please, read the student report, kindly sent to us by Anna Klucnikawho studies at Cambridge University and was hosted in summer 2015 by Aziz Aboobaker in Oxford.
Immortal worms
The Gurdon Summer Studentship has allowed me to undertake a fascinating summer project in the laboratory of Aziz Aboobaker in Oxford, working with the planarian Schmidtea mediterranea, an exciting model for stem cells, ageing and cancer1. These flatworms have the capacity to regenerate completely from the smallest fragment from almost any body part, owing to the widespread abundance of neoblasts in their mesenchyme. Neoblasts, or at least a subset, have been demonstrated to be pluripotent by single cell injection into lethally irradiated worms resulting in complete rescue2.
Since vertebrates do not have adult stem cells that are pluripotent, planarians provide a unique opportunity to study the mechanisms of stem cell maintenance, induction of differentiation, whole body regeneration, as well cancer related stem cell behaviour.
An interesting feature of neoblasts is the presence of chromatoid bodies, which strikingly resemble germ granules found in germ cells across metazoa. These are electron dense structures made up of RNPs involved in posttranslational gene regulation in the germline. Chromatoid bodies and germ granules contain many homologous proteins, which suggests a conserved germline multipotency program3. Except for Nanos, neoblasts express most germ line specific genes, such as homologues to Bruli, Piwi, and Tudor, and RNAi depletion of these proteins blocks regeneration and indicates involvement in neoblast maintenance and differentiation4.
Does Tudor reign over pluripotency?
The focus of my project was on the Tudor homologue in S.mediterranea, Smedtud-1. RNAi in a related species, S. polychroa, results in neoblast depletion, indicating a role in long-term neoblast maintenance5. From studies in other organisms we know that the Tudor domains of Tudor bind symmetrically dimethylated arginines of Piwi proteins and glycolytic enzymes in the germ granules6,7. These studies and knockdowns indicate that Tudor is involved in piRNA synthesis required for gametogenesis and stem cell maintenance. The role that Tudor may have as a pluripotency factor made me keen to investigate this mysterious protein.
What makes the picture even more interesting is that RNP granules are also found in planarian neurons and Smedtud-1 is expressed in the CNS as well as in germ cells and neoblasts8,9. However it is not known in which specific neurons Tudor is expressed or when Tudor expression is switched on and/or off.
Elucidating the Tudor lineage
I wanted to find out what is the pattern of Tudor expression in stem cell progeny as they differentiate into neurons. To do this I carried out fluorescence in situ hybridization (FISH) for several markers (tudor, coe, th, tph, ChAT10) to visualize their expression on top of an immunostain for Tudor protein.
Coe is a marker for neural progenitors and neurons and so I synthesised antisense probes to detect coe to see whether Tudor is co-expressed continuously throughout the neuronal lineage or not.
To establish where in the CNS Tudor is expressed, various markers for specific neuronal subtypes were used- th for dopaminergic, tph for serotonergic and ChAT for cholinergic neurons. Anish, a DPhil student, was thus able to help me show that Tudor protein is not expressed, for example, in dopaminergic neurons but in proximal neurons.
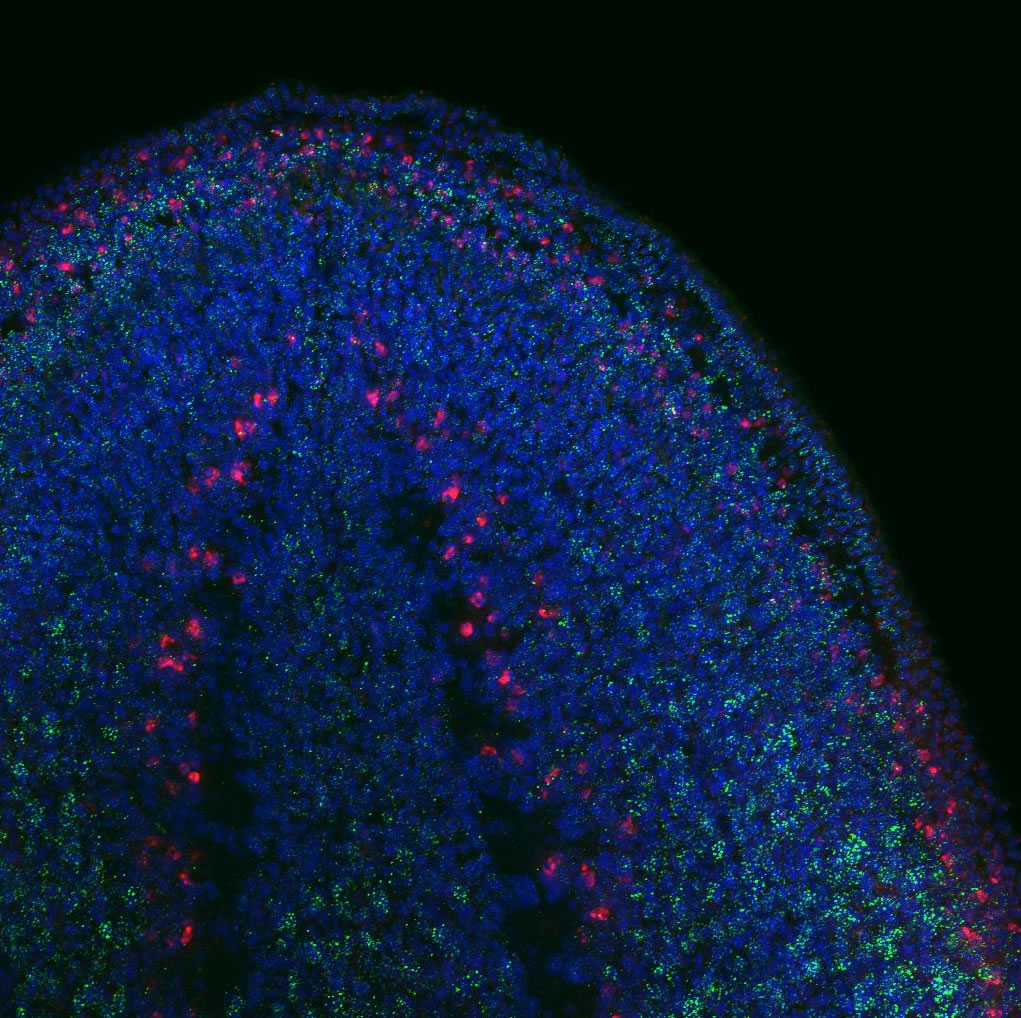
FISH and immunohistochemical staining showing the expression of a domapinergic neuron marker in (red), Tudor (green) and nuclei (blue). Courtesy of Anish Dattani.
To find out whether the expression pattern of the transcript matches that of Tudor protein. I used the genome data available to clone Smedtud-1 to synthesise antisense probes that I used for FISH jointly with immunostaining for the protein using the Tudor antibody.
Although I wasn’t able to collect all of my data in the short time that I was in the lab, I’ve achieved so much. I’ve learnt how to cut worms, microinject, clone genes, synthesise probes and carry out immuno and in situ protocols. I’ve learnt to always ask when in doubt. I’ve learnt to be scrupulous. I’ve learnt that it can be very frustrating when experiments don’t work out as planned and when time runs out. But those little setbacks showed me just how determined I am to do science.
The Aboobaker lab was extremely welcoming and supportive. Thank you to Aziz, Natasha, Dani, Nobu, Prasad, Anish, Sounak, Yuli, Sam, Damian, Alvina, Holly, Ben and Alex.

 (No Ratings Yet)
(No Ratings Yet)





 This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series
This post is part of a series on a day in the life of developmental biology labs working on different model organisms. You can read the introduction to the series  (10 votes)
(10 votes)









 (5 votes)
(5 votes)