A portuguese person, a spanish person and an english person meet in a bar…
… and start discussing developmental biology. This may sound like the beginning of a joke, but in fact happened during the Joint Meeting of the Portuguese, Spanish and British Societies for Developmental Biology, which took place in Algarve, Portugal, in early October. The meeting venue, besides having the aforementioned bar, was also closely located to the beach, which we were able to enjoy thanks to a pleasant weather. Some of the participants also took advantage of the beautiful and family-friendly location to bring their own families. Nevertheless, the scientific talks and poster sessions still managed to draw the participants away from the seaside.
A meeting by the sea. Photo by Catarina Vicente @the_node.
The meeting started with early development, with a plenary lecture on the principles of pluripotency presented by Austin Smith. The lecture focused on the ongoing quest to establish human naïve embryonic stem cells in vitro independently of pluripotency transgenes, showing the progresses achieved so far and presenting the challenges that still need to be overcome.
The transition from pluripotency to lineage commitment was explored by Sally Lowell, whose work identified some of the factors that prime cells for differentiation and revealed a role for adhesion molecules in the decision to differentiate. Berenika Plusa presented the advantages of using rabbit as an alternative model to study early mammalian development. Andrew Johnson showed that axolotl, an organism without extraembryonic tissues, can be used to study later roles of the pluripotency factor Nanog.
The regulation of neuronal differentiation was also the focus of several talks. Kate Storey showed how differentiating neurons in the chick neural tube undergo apical abscission and revealed new evidence for the involvement of microtubule dynamics and adhesion molecules in this process. Also in the chick neural tube, Elisa Marti presented work on the role of Shh signalling in the decision to proliferate or differentiate and showed that the subcellular localisation of several Shh pathway components contributes for this decision. Anna Philpott also talked about division/differentiation in the nervous system and the regulation of proneural factor activity by phosphorylation in Xenopus. François Guillemot highlighted the role of the proneural factor Ascl1 in adult brain neurogenesis and how modulation of Ascl1 stability affects the balance between quiescence and differentiation. The talk by Alexandre Raposo was also on Ascl1 and its function promoting chromatin accessibility during neurogenesis.
The link between adult neural stem cells and cancer was discussed by two drosophilists. Cláudia Barros is using a fly brain tumour model to identify new factors involved in tumour initiation, while Rita Sousa-Nunes is using this model to study the interaction between tumour cells and the microenvironment.
Moving away from neural lineages, we also heard about regulation of proliferation, differentiation and cell movement of presomitic mesoderm progenitors from Leonor Saúde and single cell oscillators as components of the segmentation clock during somitogenesis from Andrew Oates.
Later in development, the formation of the inner ear lumen in zebrafish was introduced by Berta Alsina, revealing that mitotic cell rounding and epithelial thinning regulate lumen expansion. Juan R. Martinez-Morales talked about optic cup morphogenesis in zebrafish, showing that both rim involution and basal constriction contribute to cup folding. Zebrafish embryos were also the stars in the beautiful movies shown by Claudia Linker, whose work combined live imaging with cell ablation to test the role of leader, follower and pre-migratory cells in the collective migration of neural crest cells.
At the chromatin level, Ana Pombo proposed that the priming of developmental genes for future expression in embryonic stem cells involves the Polycomb complex, a specific modification of the RNA polymerase II and local transcript degradation. Rui Martinho showed how chromatin remodelling is involved in the transcriptional reactivation of the Drosophila oocyte during meiosis. Javier Lopez-Rios presented his work on a limb-specific enhancer responsible for the spatial differences in Ptch1 expression between mice and bovine, which underlies their distinct limb anatomy.
The meeting ended with a plenary talk by Moisés Mallo, who presented his work on Gdf11 as the coordinator of the trunk to tail decision during vertebrate embryogenesis and revealed an unexpected role for a pluripotency gene in trunk specification.
The meeting included many other exciting talks that have not been reported here. Overall, the meeting programme showcased the diversity of the developmental biology field in terms of subjects and model systems. The meeting also achieved a perfect gender balance among speakers – 17 female and 17 male speakers. Outside the lecture hall, scientific discussions continued throughout the free afternoons and outdoor poster sessions while enjoying the warm weather. And, of course, in the bar.
As the meeting came to an end, the sunny weather turned into a rainy storm, which made the departure a little less sorrowful.
Coffee break al fresco. Photo by SPBD. (6 votes) Loading...
Mike Levine, director of the Lewis-Sigler Institute for Integrative Genomics at Princeton University, is a developmental biologist who has dedicated his career to understanding how gene expression is regulated during development. Some of his most significant research, such as the co-discovery of the homeobox genes and his work on even skipped stripe 2, was performed in Drosophila, but he has since branched out to Ciona intestinalis, which he is using as a model to understand how vertebrate features have evolved. We had a lively chat with Mike at this year’s Society for Developmental Biology (SDB) meeting, where he was awarded the Edwin Grant Conklin Medal.
Here at the SDB meeting you will be awarded the Conklin Medal by the society. What does it mean to you to receive this prize?
It is a really special honour for me, for a number of reasons. First, the list of people who got it before me is pretty awesome, so I am very proud to be among them. People like John Gurdon, Nicole LeDouarin, and some of my friends and peers like Richard Harland, Cliff Tabin, Marianne Bronner… The other reason why this award is special for me is because Conklin did his lineage-tracing studies in sea squirts, and half my lab has worked on this model system for 20 years. To my knowledge, I am only the second sea squirt guy to get the Conklin Medal, after my good friend Nori(yuki) Satoh. For those of us who work on the same material that Conklin himself studied, this is a very special honour. He was always one of my scientific heroes.
You were SDB president a few years ago. What do you think is the role of the society?
A field of study is only as good as its smartest young people. I think it is important for the society to reach out to the young, talented stem cell, computational and genomics researchers and say: ‘Hey, this is a really cool field of study’. We have one advantage over most other fields: we work on intrinsically beautiful material. What is more beautiful than a developing embryo? I remember when I was an undergraduate seeing for the first time movies of developing chick and frog embryos and I was just mesmerised. I just thought: ‘Oh man, that is what I want to study’. And it is not only visual, it is a highly integrated science. It really pulls together so many different disciplines. We have a lot to offer to the next generations of discoverers, and the SDB needs to reel these young men and women in.
How did you first become interested in biology? I understand that you considered becoming a medical doctor…
I always had an interest in the life sciences, and enjoyed going to my backyard to dissect bugs with my little microscope. I came from a blue-collar family, so if you were good at biology, which I was, it was only logical that you should become a doctor and make some money. For a modest Jewish family, being a doctor is a big escalation in status. I tried to be a good boy and even took the medical school admissions test and went to a couple of interviews, but it really was not for me. I have always been a hypochondriac, so I can’t even imagine how many times I would have tested my own urine and blood for whatever disease I was learning about! So I had this ‘going to medical school’ thing hanging over me during my undergraduate studies, but I was lucky to discover the wonderful world of biological research.
It was really hard in Berkeley to find a lab where you can do research as an undergraduate. Fortunately, I had an amazing stroke of luck to get to work in Allan Wilson’s lab. He and Mary-Claire King had proposed that regulatory DNAs were really important in evolution and in distinguishing chimps and humans, and this definitely infiltrated my thinking.
During your scientific career you have examined how gene regulation is controlled. What excites you about this topic and why did you choose Drosophila as a model?
I love gene regulation. I love the process of transcription so much that I regard RNA as an unfortunate by-product of an otherwise elegant process! I think part of it is that when I was an undergraduate I must have learned about the lac operon in three different classes: genetics, molecular biology and protein biochemistry. It is an inherently beautiful mechanism. Who would have thought that a bacterium exposed to sugar would deploy this elaborate and elegant transcriptional response? The developmental biology classes by Fred Wilt also really stayed with me. So I had a strong sense that gene regulation was a cool process from my undergraduate studies. This was reinforced by my undergraduate research in Allan Wilson’s lab. There they were talking about regulatory DNA, but instead of bacteria they were looking at animal cells.
I first became interested in flies because of a Scientific American article written in 1975 by the Swiss molecular geneticist Ernst Hadorn on transdetermination. He took wing imaginal discs out of larvae and cultured them in the stomachs of recipient flies, so they proliferated for longer than they normally would. He then grafted these discs back into a recipient larva that underwent metamorphosis, and found that sometimes the grafted tissue didn’t become the original structure that it was slated for, but the whole thing transformed into a leg. I thought that was a really exciting discovery. Later on, I read about the homeotic mutants that Tom Kaufman and Ed Lewis were working on and figured: ‘It has got to be gene regulation, and it has got to be in flies’.
As you mentioned earlier, part of your lab now works on Ciona. Why this organism?
I was co-director of the embryology course at Woods Hole for a few years, and this gave me the chance to get exposed to a lot of different systems in developmental biology. When I heard Richard Whittaker and Nori Satoh talk about Ciona, I immediately loved the system. I don’t know if it triggered recollections about Conklin’s work, which I had been taught about as an undergraduate, but I just liked the simplicity. Embryogenesis is amazingly complex, and I really don’t think in 3D so well. But when I heard Whittaker and Satoh discuss Ciona, where the movements are not that complex, I thought: ‘This is a system I can understand’.
Our fly studies have always been pretty abstract, studying gene regulation but never connecting it to morphogenesis. I always thought we should be able to link the two, but at least for me it seemed hopeless to try it in flies. There are so many cells, the processes are complex and occur very late in development. But I thought Ciona might be a good place to attempt this and complement our fly studies. The thinking was: ‘Let us extend our studies from gene regulation in Drosophila to a model organism in which we can study gene regulation and the connection to cellular morphogenesis in development’.
Bob Zeller, who trained with Eric Davidson studying sea urchins but had done undergraduate work in sea squirts, came to my lab as a postdoc to set up this system. I thought we were nearing the end of the line with the flies, so the plan was to wind down and eventually just convert completely to sea squirts. But every time I think I am going to drop the flies, I just can’t. I like sea squirts, don’t get me wrong, but I am really a fly guy. I feel like Michael Corleone in The Godfather III: “Every time I try to get out, they keep pulling me back”. The early fly embryo is the sweetest system in the world for looking at gene activity in development. The last 10 years have been dominated by fantastic new technologies, such as single-molecule live imaging, and these just work like a charm in the early fly embryo. So I can’t leave it!
You did your postdoc with Walter Gehring at the University of Basel. How did your time there influence your career?
Where do I begin in describing my 15 months in Basel? Culturally, it was a defining experience for me. I had never been a political person. But in Basel people would stay up drinking and discussing politics, and I learned that not everyone agreed with American policy. I had never realised how parochial my experience was until I went there, so it was really eye opening and gave me a broader perspective. There was also a special camaraderie that I had never experienced before. Ah, and Europe! I had never been to Europe before, and it is like a giant museum, with its cathedrals and art… It was truly a mind-blowing, defining experience.
As for the lab, it was a hot and cold experience. The hot part was that I met some of my best friends and collaborators, like Markus Noll, Erich Frei, Bill McGinnis and Ernst Hafen. Also on the good side, everything I have done with Drosophila for the ensuing 30 years was a direct consequence of my time in Basel working on gene expression in the fly embryo. Unfortunately, Walter and I just didn’t get along, so I eventually had to leave. But as difficult as my personal relationship was with Walter, I would probably do it again, because I got an enormous amount from the experience, both culturally and scientifically.
What would you consider to be your most important discovery?
The work I did with McGinnis and Hafen on homeobox genes was pretty good, but I don’t like to think I did my best stuff in those 15 months as a postdoc. I think that I am proudest of my work on eve stripe 2. The project was launched by a student named Dusan Stanojevic who was very mercurial, very high maintenance, but absolutely brilliant. When he started the project he said: “This is going to be the lacoperon and the lambda switch of developmental biology!”. At the time I thought he was cracked, but 25 years or so later I would say that there is something to it! Our work on eve stripe 2 was less a single discovery than a war of attrition. It took 3 to 4 years of really hard work, doing DNA binding assays, targeted mutagenesis and transgenesis, which were harder methods then than they are now. Some amazingly talented scientists, including Tim Hoey and Steve Small, worked through that problem.
Which scientific questions would you like to tackle in the future?
TA few years ago Delsuc et al. (2006) showed that the urochordates (which include sea squirts) and not the cephalochordates, as most text books still say, are the closest living sister group to the vertebrates. This paper has been extremely influential in our thinking because it means that if you are interested in understanding the evolutionary origin of some of the major vertebrate innovations, such as the neural crest, neurogenic placodes and second heart field, Ciona tadpoles are a good place to look. Of course Ciona doesn’t have a neural crest, but it does have a cell type with some of the properties of neural crest. We also found that Ciona tadpoles have neurogenic proto-placodes, another feature of the vertebrate head.
On the fly side I am very excited about the use of single-molecule live imaging. One of the big benefits of my recent move to Princeton is the close proximity to two of my favourite young Drosophilacollaborators: Thomas Gregor, a physicist who does live imaging in the fly embryo, and Stanislav Shvartsman, a chemical engineer studying signalling in fly eggs and embryos. Collaborating with these two labs is going to invigorate our studies. One line of research that I am most excited about is visualising enhancer-promoter communication directly. The human genome is just riddled with hundreds of thousands of enhancers. In other words, a typical gene in the human genome is regulated by up to 50 different enhancers. So all of a sudden you have to worry about trafficking: how do the right enhancers get to the right promoters at the right time? For all we know, this could be the rate-limiting determinant in the patterning of the Drosophila embryo. Thomas is devising strategies for directly visualising the interaction of remote enhancers with promoters in living embryos during key patterning events. That is very exciting.
You mentioned that you have moved to Princeton, where you are now the director of the Lewis-Sigler Institute for Integrative Genomics. What are you hoping to achieve in this new position?
The Lewis-Sigler is called a genomics institute but it really started as a systems biology institute, initially led by Shirley Tilghman and then David Botstein. Botstein was the first person I heard explain properly what systems biology is and the concept really turned me on. Systems biology is the systematic identification of every component of a complex process. You need the experimentalists to generate the big data, the computer scientists to handle the big datasets, and then quantitative biologists to model these datasets so that you can understand emerging properties of the process. I can know everything about a neuron in the neocortex but if I multiply that by a million I am not going to learn how consciousness works. You have to do something different. This is the philosophy of systems biology and I still believe in it. The Lewis-Sigler institute is like a scientific Noah’s Ark: it has a couple of computer scientists, a couple of high-throughput biologists, a couple of physicists, a couple of engineers. It is just the right mix of talents for systems biology, so I see no need to deviate from Botstein and Tilghman’s original vision. I just want to have some fun, and bring people together towards this enterprise of trying to learn the emerging properties of really complex processes, like the patterning of the fly embryo. I think there are wonderful challenges and opportunities, and with these new technologies we can take systems biology into the new millennium.
What is your approach to running a successful lab?
The alumni of my lab are an amazing group of people, and so many of them run their own labs now. I would love to take credit for it but, believe me, they came in pretty good! I have a reputation of being pretty demanding, a pretty tough boss. I have in me a bit of my Jewish uncle, who fought in World War II and had this warmth on the one hand and this tough ‘you are not quite good enough’ on the other hand. And I think I do a little bit of that in the lab.
I aim to keep my lab members excited about their project. I try to constantly look at the big picture and, if I have an idea, I try to give it to them when I am at my most enthusiastic. They might tell you that I am tough, but I hope they’ll also tell you that I do love science. It’s like with sports people getting towards the end of their careers: when you ask them what keeps them going they all say the same thing – they love the process. They like getting up in the morning, working out, training, they like the banter in the locker room. I really enjoy the process too. I like going in to the lab. I think whatever influence I have had in helping my lab members has been my enthusiasm for the process.
What is your advice for young scientists?
It is much harder now to find an identity for yourself in science. I was in the right place at the right time, I admit it. I got a great job when I was young and the field was wide open. It is much more crowded now. The whole ‘follow your passion and everything will work out’ may have been true 20-30 years ago, but it is not as true now. My hard-nosed advice to young scientists who want to continue being scientists (and you can do this in many capacities, it doesn’t have to be as the PI of a lab) is to learn technology. Go to a graduate programme or do a postdoc where you have access to the cutting-edge technologies. When I was a postdoc in the Gehring lab, Ernst Hafen and I helped develop the first in situ hybridisation methods with fixed tissues and I think it was that method that really got me a job. When you know a good technology, people are interested in it, even if they are not interested in the specific process you are working on. You increase your value. Discovery depends on technology now more than it ever did. The old guys did the easy stuff: we pillaged the low-hanging fruit a long time ago. I do believe the best is yet to come, but it requires technology. I would advocate imaging or genomics, or, best of all, somewhere in between! I also relate what I heard from many people over the years, including James Watson: “Don’t go straight up the middle in an established discipline. The action is at the cusps”. I think that is also very good advice.
What would people be surprised to find out about you?
There is the perception that I am a bit of an eccentric, and I think that even the people in my lab would be surprised to see how ordinary my private life really is. I am a family man, and I enjoy a tightknit relationship with my wife and two sons. We enjoy conventional suburban pleasures, such as going to the movies.
BONUS!: Hear Mike give his account of when he almost set one of his postdocs on fire!
Here are the highlights from the current issue of Development:
Atoh1: earmarked for differentiation
Atoh1 is a key regulator of the differentiation of hair cells, the sensory cells that support audition: it is upregulated during their differentiation and downregulated at postnatal stages. But what are the mechanisms underlying Atoh1 transcriptional regulation during inner ear development? To address this question, Neil Segil and co-workers (p. 3529) analysed the epigenetic status of Atoh1 in mouse hair cell progenitors. They report that histone H3 at the Atoh1 locus is bivalently marked by the repressive tri-methylation of lysine 27 (H3K27me3) and the permissive tri-methylation of lysine K4 (H3K4me3). In nascent hair cells, Atoh1 upregulation is accompanied by a reduction in H3K27me3 and requires the appearance of the permissive acetylation of histone H3 lysine 9. At postnatal stages, Atoh1downregulation is achieved by an increase in H3K9me3, which is a mark indicative of transcriptional silencing, and a reduction in histone H3 acetylation. In early postnatal supporting cells (a cell population that separates and surrounds hair cells and can regenerate them during the first postnatal week in mice), the bivalent marks are maintained, potentially explaining their latent regenerative capacity. This study suggests a mechanism for the epigenetic control of Atoh1 levels during inner ear development and reveals a potential target for future regenerative efforts to replace mammalian hair cells.
Revisiting blastomere equality
The fertilised mammalian egg gives rise to seemingly equivalent blastomeres until the fourth cleavage division, when the first indication of lineage specification appears. At this point, certain blastomeres divide symmetrically and others asymmetrically. When do these apparently identical cells diverge and how do these differences arise? To answer this question, Enkui Duan and colleagues performed single-cell transcriptional analysis of human and mouse blastomeres (p. 3468). By studying the mammalian zygote, in which transcription – a known source of heterogeneity during mitosis – is mostly silent, the authors showed that small biases in gene expression arise after the first cleavage division from the unequal distribution of cellular substances between daughter cells, called ‘partitioning errors’. These are especially pronounced for transcripts present in small quantities, which are more likely to be asymmetrically distributed. As cleavage divisions progress, the activation of embryonic transcription minimizes or amplifies the initial biases through positive or negative feedback regulation. Furthermore, the authors show that lineage specification is driven by the relative ratio of pairs of competing lineage specifiers, such as Cdx2 and Carm1, the levels of which are determined by both cleavage history and de novo transcription. This study shows that symmetry breaking leading to lineage specification is a continuous process that emerges as early as the two-cell stage, before morphological differences between blastomeres are detectable.
Revising the origin of thyroid C cells
Thyroid C cells (or parafollicular cells) are neuroendocrine cells found in the thyroid gland that secrete calcitonin. To date, it has been thought that these cells arise from the neural crest but here, on p. 3519, Mikael Nilsson and co-workers overturn this view. Using lineage tracing, they show that Wnt1-positive neural crest cells do not give rise to C cells in the mouse embryonic thyroid gland. Instead, they reveal, thyroid C cells are derived from Sox17-positive anterior endoderm. The researchers further show that the transcription factors Foxa1 and Foxa2, which are known to play a role in the development of other endoderm-derived populations, are co-expressed in C cell precursors, where they play non-redundant roles. Finally, the authors also show that Foxa1 and Foxa2 are expressed and appear to play distinct roles in human medullary thyroid carcinoma (MTC) cells. Together, these findings disprove the current concept of a neural crest origin of thyroid C cells and argue that MTC tumours should be reclassified as neuroendocrine tumours with an endodermal origin, a change that, from a clinical perspective, may open up new avenues in the search for MTC treatments.
Lin-28: balancing stem cell divisions
The Drosophila intestine is known to undergo adaptive growth in response to feeding, and this growth is fuelled by the symmetric self-renewing divisions of intestinal stem cells (ISCs). But what controls ISC division patterns? Here, on p. 3478, Nicholas Sokol and colleagues reveal that the RNA-binding protein Lin-28 promotes symmetric ISC divisions and hence tissue growth in the Drosophila intestine. They first show that Lin-28 is highly enriched in adult Drosophila ISCs. They further report that, although lin-28 null mutants are viable, adult mutant animals exhibit reduced numbers of ISCs. Following on from this, the researchers demonstrate that Lin-28 is required in ISCs to promote food-triggered self-renewing divisions and expansion of the ISC pool. Finally, they report that Lin-28 acts independently of its well-known target let-7 and instead interacts with mRNA encoding the insulin-like receptor (InR), suggesting that Lin-28 modulates InR levels, and thus insulin signalling, to control cell division patterns. In summary, the authors propose that Lin-28 acts as a stem cell intrinsic factor that boosts insulin signalling in ISCs and promotes their symmetric division in response to nutrients.
PLUS:
An interview with Mike Levine
Mike Levine, director of the Lewis-Sigler Institute for Integrative Genomics at Princeton University, is a developmental biologist who has dedicated his career to understanding how gene expression is regulated during development. We had a lively chat with Mike at this year’s Society for Developmental Biology meeting, where he was awarded the Edwin Grant Conklin Medal. See the Spotlight article on p. 3453
Heparan sulfate proteoglycans: a sugar code for vertebrate development?
Heparan sulfate proteoglycans (HSPGs) have long been implicated in a wide range of cell-cell signaling and cell-matrix interactions, both in vitro and in vivo in invertebrate models. Here, Poulain and Yost provide a comprehensive overview of the various roles of HSPGs in these systems and explore the concept of an instructive heparan sulfate sugar code for modulating vertebrate development. See the Review on p. 3456
Featured movie
This issue’s featured movie shows mouse development from E11.5 to E14.0, as displayed in the 4D atlas of mouse development developed by Wong and colleagues. Read their paper on this useful resource: http://bit.ly/1NeEiAg
Department/Location: Wellcome Trust – Medical Research Council Cambridge Stem Cell Institute, University of Cambridge, UK
Salary: £24,775-£28,695
Reference: PS07303
Closing date: 12 November 2015
Fixed-term: The funds for this post are available until 30 June 2017 in the first instance.
The Wellcome Trust-Medical Research Council Cambridge Stem Cell Institute (SCI) comprises 200 researchers spanning fundamental science through to clinical applications. Our goal is to advance disease modelling, drug discovery and regenerative medicine through understanding the genetic and biochemical mechanisms that control stem cell fate.
Applications are invited for a computational biologist to join the SCI’s bioinformatics group. We apply state-of-the-art experimental and computational methods toward understanding the biological properties and biomedical potential of stem cells.
The vacant post is at Research Assistant level and would be suitable for individuals with either a computational or biological background. The post holder will bridge the research groups within the SCI and will analyse next generation sequencing data using cutting edge software tools on internally (SCI)-produced data. He/she will work in a team of bioinformaticians dedicated to the application of modern bioinformatics techniques to stem cell research.
Candidates should be able to work in a UNIX/Linux environment. Proficiency with a scripting language (e.g. Perl/Python) and statistical data analysis tools (R, Matlab) would be a strong advantage. Additional experience with analysis of high-throughput sequencing data is desirable. The post holder will be involved in the development and interpretation of multilayer genomic, transcriptomic and epigenomic data. Necessary training in specialist computational tools will be provided; the main criterion is an enthusiasm to use bioinformatic approaches to advance stem cell research.
To apply online for this vacancy and to view further information about the role, please visit: http://www.jobs.cam.ac.uk/job/8347. This will take you to the role on the University’s Job Opportunities pages. There you will need to click on the ‘Apply online’ button and register an account with the University’s Web Recruitment System (if you have not already) and log in before completing the online application form.
Please upload your Curriculum Vitae (CV) and a covering letter in the Upload section of the online application to supplement your application. If you upload any additional documents which have not been requested, we will not be able to consider these as part of your application.
The closing date for all applications is the Thursday 12 November 2015.
Interviews will be held week commencing 23 November 2015. If you have not been invited for interview by 23 November 2015, you have not been successful on this occasion.
Please quote reference PS07303 on your application and in any correspondence about this vacancy.
The University values diversity and is committed to equality of opportunity.
The University has a responsibility to ensure that all employees are eligible to live and work in the UK.
Evolution and development of shark skin teeth inspired surface design for global CO2 reduction
Dr Gareth Fraser, APS, University of Sheffield; Co-supervisors: Dr Mark Johnson, Department: School of Engineering, Centre for Engineering Dynamics, University of Liverpool;
Dr Zerina Johanson, Natural History Museum, London, Department of Earth Sciences
A Ph.D studentship is available for an interdisciplinary project focused on evolutionary developmental biology of shark skin teeth and engineering shark-inspired surface geometries for reduced drag leading to lower CO2 emissions across several industries. This is an ideal opportunity for an ambitious candidate to work at the interface of evo-devo and engineering. This project aims to explore novel methods to understand the evolution and development of shark tooth patterning and how these data can be modelled in silico for more energy efficient surface design that can be 3D-printed for functional models of drag reduction. This project would suit a candidate interested in skin tooth development in sharks and the diversity of these structures in pattern and morphology across species to understand the most efficient patterns and geometries in nature, and then how nature can help us develop solutions to environmental issues in the engineering industry.
Here is October’s round-up of some of the interesting content that we spotted around the internet!
News & Research
– Developmental Biologist Eric Davidson, who was based at CalTech, has sadly passed away. You can learn more about Eric and his career in this Q&A published in Current Biology a few years ago.
– Interesting piece in Nature News considers how biohackers (amateur biologists) are using CRISPR.
– Masayo Takahashi is the winner of the inaugural Ogawa-Yamanaka Stem Cell Prize, awarded by the Gladstone Institutes. Also don’t miss the chance to vote for the 2015 Stem Cell Person of the Year in Paul Knoepfler’s blog.
– How to unboil an egg and how to make a chicken walk like a dinosaur are just some of the winners of this year’s Ig Nobel prizes.
– How scientists fool themselves and how they can stop- preventing bias in science, in Nature.
In multicellular animals and plants, the single-celled zygote develops into the embryo. In the model plant Arabidopsis thaliana, the zygote divides asymmetrically to form a small cytoplasmic apical cell, which is the precursor of the proembryo, and a large vacuolated basal cell, which develops into the extra-embryonic suspensor (Figure 1). Communication between the intra-embryo (proembryo and suspensor) and the inter-embryo (embryo and surrounding tissues) is required for proliferation, pattern formation, and differentiation during embryogenesis1-3. Moreover, it has long been known that in several plant species, including Arabidopsis, disruption of the original embryonic tissue induces formation of newly embryonic cells derived from the extra-embryonic tissue4-6. However, the process by which an embryonic cell is initiated from the extra-embryonic tissue remains unclear. Angiosperm embryogenesis in vivo occurs within multiple layers of maternal tissues within the flower; therefore it is difficult to observe and analyze the intercellular communications in living material in real time. The development of a live-cell imaging system has been long-awaited to reveal how cell fate is specified by intercellular communication during early embryogenesis. Recently, we have developed a live-embryo imaging system to visualize cell division and cell-fate specification in Arabidopsis thaliana. We incorporated two advanced technologies—micro-engineering and optical manipulation—into this system.
Figure 1. Schematic representation of embryogenesis within Arabidopsis flowers. After double fertilization, the embryo and endosperm develop in the ovule. The zygote divides asymmetrically into an apical cell (embryonic cell) and a basal cell (extra-embryonic cell). The apical cell divides vertically to generate the proembryo, which ultimately gives rise to the adult plant. The basal cell divides horizontally to produce the suspensor (which acts like an umbilical cord in mammals), and finally undergoes programmed cell death.
First, we developed an in vitro ovule culture system so that time-lapse imaging could be performed. An in vitro culture system was necessary for real-time observation because embryogenesis could not be observed in vivo on account of the deep tissues. However, we could not observe any cell division in the isolated early embryo7. Therefore, we cultured the embryo within the maternal tissue—the ovules—in vitro. We used liquid medium to prevent the ovules drying up during long-term culture and placed the ovules close to the cover glass. However, stable observation of the ovules in the liquid medium was difficult because of their round shape, so we fabricated PDMS [poly(dimethylsiloxane)] microdevices to adjust the orientation and fix the position of the ovules using micro-engineering techniques.
PDMS has several advantages in the field of developmental biology, such as a high level of biocompatibility, including efficient gas exchangeability and a suitable transparency for observation. PDMS-based microfluidic devices have also been used for animal cell cultures8. Recently, plant researchers have also started to use PDMS-based microdevices for analyzing the growth of root tips, pollen tubes, and seedlings as a lab-on-a-chip9. Initially, we fabricated a PDMS microcage array to fix the ovules in the liquid medium10. To increase the trapping efficiency of the ovules, we modified the PDMS microcage array to a micropillar array11 (Figure 2). Micropillar arrays have a weaker effect on ovule growth because of the physiological flexibility of PDMS micropillars. Thus, we were able to conduct time-lapse imaging of cell division—from the zygote to the heart-stage embryo—for 3 days within the ovules in Arabidopsis embryogenesis.
Figure 2. Arabidopsis ovules on a PDMS micropillar array.
Another advanced technique is optical manipulation. To elucidate the mechanisms of cell–cell communications within the tissues, it is important to carry out a spatio-temporal analysis to determine when, where, and which cells are communicating. Optical manipulation is a powerful tool for such real-time analysis. Because lasers have high optical intensities at the focal point, laser ablation has long been used to disrupt cells. In plants, UV lasers have been used for laser ablation12,13. However, there are limitations to how UV lasers can be used. Short-wavelength lasers, such as UV lasers, are susceptible to refraction, absorption, and scattering from the tissues. Therefore, it is difficult to ablate the cell at the level of a single cell within deep tissues.
Recently, near-infrared pulse lasers have been gradually used to overcome the limitation of UV lasers. Near-infrared pulse lasers are tolerant to scattering within the tissues because of their longer wavelength. In addition, femto-second pulse lasers can induce nonlinear optical effects, such as plasma formation, at a sub-femto-liter volume. Thus it can induce damage even within a cell14. Hasegawa et al. performed the disruption of actin filaments within a tobacco-cultured cell using a femto-second pulse laser in real time15.
We performed laser disruption to our live-embryo imaging system using a femto-second pulse laser11. When an apical cell (embryonic cell) was disrupted by laser irradiation, the cell fate of the remnant basal cell (extra-embryonic cell) was converted, through changes in identity gene expression, into an embryonic cell (Figure 3). In terms of the disruption of basal cells, we found that apical cells were not converted into basal cells. Thus, we succeeded in performing real-time observation of cell-fate conversion for the compensation of an embryonic cell. We propose that the fate and behavior of each individual cell in Arabidopsis embryos is determined via intercellular communication with neighboring cells, and that such flexibility contributes to the robustness of plant embryogenesis.
Figure 3. Time-lapse images of embryonic marker, DRN::erGFP (green, ER localization) after laser irradiation of the apical cell. When an apical cell was disrupted (asterisk), the basal derivative cell underwent DRN expression and divided like an apical cell. Scale bar, 20 µm.
Our imaging system enables direct visualization of the dynamics of cell-fate specification during embryogenesis using micro-engineering techniques. In plants, cell fates are specified via positional information during development. Femto-second laser irradiation, as a form of optical manipulation, enables a spatio-temporal induction of cell-fate conversion that is impossible with genetic ablation. Thus, these techniques will enable novel insights into the molecular mechanisms of cell-fate specification via intercellular communication in plant development. Moreover, these techniques are highly suited to research into developmental biology as well as to plant research, so will be of interest to broad developmental biologists in addition to their applicability to live-cell analysis.
1 Kawashima, T., & Goldberg, R. (2010). The suspensor: not just suspending the embryo Trends in Plant Science, 15 (1), 23-30 DOI: 10.1016/j.tplants.2009.11.002
2 Javelle, M., Vernoud, V., Rogowsky, P., & Ingram, G. (2011). Epidermis: the formation and functions of a fundamental plant tissue New Phytologist, 189 (1), 17-39 DOI: 10.1111/j.1469-8137.2010.03514.x
3 Lafon-Placette, C., & Köhler, C. (2014). Embryo and endosperm, partners in seed development Current Opinion in Plant Biology, 17, 64-69 DOI: 10.1016/j.pbi.2013.11.008
4 Haccius, B. (1955). Experimentally Induced Twinning in Plants. Nature176, 355-356, doi:Doi 10.1038/176355a0
5 Yeung, E., & Meinke, D. W. (1993). Embryogenesis in Angiosperms: Development of the Suspensor Plant Cell, 5 (10), 1371-1381 DOI: 10.1105/tpc.5.10.1371
9 Sanati Nezhad, A. (2014). Microfluidic platforms for plant cells studies Lab on a Chip, 14 (17), 3262-3274 DOI: 10.1039/C4LC00495G
10 Park, J., Kurihara, D., Higashiyama, T., & Arata, H. (2014). Fabrication of microcage arrays to fix plant ovules for long-term live imaging and observation Sensors and Actuators B: Chemical, 191, 178-185 DOI: 10.1016/j.snb.2013.09.060
11 Gooh, K., Ueda, M., Aruga, K., Park, J., Arata, H., Higashiyama, T., & Kurihara, D. (2015). Live-Cell Imaging and Optical Manipulation of Arabidopsis Early Embryogenesis Developmental Cell, 34 (2), 242-251 DOI: 10.1016/j.devcel.2015.06.008
12 van den Berg C, Willemsen V, Hendriks G, Weisbeek P, & Scheres B (1997). Short-range control of cell differentiation in the Arabidopsis root meristem. Nature, 390 (6657), 287-289 PMID: 9384380
13 Higashiyama, T., Yabe, S., Sasaki, N., Nishimura, Y., Kuroiwa, H., & Kuroiwa, T. (2001). Pollen Tube Attraction by the Synergid Cell Science, 293 (5534), 1480-1483 DOI: 10.1126/science.1062429
14 Tirlapur, U., & König, K. (2002). Targeted transfection by femtosecond laser Nature, 418 (6895), 290-291 DOI: 10.1038/418290a
15 Hasegawa, J., Higaki, T., Hamamura, Y., Kurihara, D., Kutsuna, N., Higashiyama, T., Hasezawa, S. & Matsunaga, S. (2014). Vacuole subdivision in plant cell growth by the genotoxic stress inducing DNA double strand breaks. Cytologia79, 467-474, doi:10.1508/cytologia.79.467
This Sticky Wicket article first featured in Journal of Cell Science. You can read the Mole’s visit to the first 5 circles of Grant Hell here. Read other articles and cartoons of Mole & Friends here.
Mole was revising his rejected grant and moaning about how unfair it all is, when the shade of Francis Bacon, the Father of the Scientific Method, appeared to take him on a tour of Grant Hell. Sir Francis (‘Frank’) looks like Dumbledore but talks like a wanna-be hipster but, hey, maybe Mole is just dreaming all this. Anyway, they went to Grant Hell. There, they met lots of wretched souls who were not successful in revising their grants, and had lots of reasons why not.
Oh, one more thing. For some reason, the story is told in triplets that don’t rhyme. Probably they did in some other language, but it would have been too much trouble to translate it and make it rhyme. Mole may be getting lazy. Or maybe he’s up to something? No, most likely he’s just lazy. We join our travelers, having passed the Fifth Circle, and have continued up to the Fourth.
“Here is the Fourth Circle,” said Frank. This was Greed.
“But hey,” I said, “I thought greed is good. That’s what they tell us,
It makes the market work.” Frank shook his head.
“Wait,” I said, “Are you saying that these poor people are here
Because they wanted to do too much? They wanted too many grants?
I mean, what’s wrong with a little ambition if the science is good?
“Listen,” I went on, “I’m getting a little tired of hearing that somebody, who
I think is a really terrific scientist, has ‘too many grants.’ I mean, if they are
Getting the best work done, maybe they deserve the grants. You know?”
“And now,” I think I was on a roll. “Now you tell me that they are here,
In Grant Hell, because they work too hard.”1 “Easy, Big Guy,2” said Frank,
And I looked to see if he was talking to me. He was. Nobody calls me ‘Big Guy.’
He went on. “You’ve got it all wrong. They aren’t here because they wanted
Too many grants. They are here because they confused grant funding and power.
Their greed is not to do more great science but to amass influence.”
“Oh,” I said. “Then they get what they deserve.” Because being ‘powerful’
As a research scientist is a bit like being the King or Queen of Cheese.3
It isn’t worth very much and can smell pretty terrible. Frank cracked up at that one.
We had moved to the Third Circle, where the wretched never stopped writing,
Except to eat the paper they were generating. Or electrons, but I couldn’t see that.
Then they would write some more. “Here is Gluttony,” whispered Frank.
I stopped one poor soul who was eating a page as it came out of a printer.
“Why are you doing this?” I asked. “Oh,” he moaned. “You know how it is,
I don’t get my grants funded, so I write more and more and more.
“I was putting in a grant every cycle, but that didn’t work, so I went to two,
Then three. I don’t have anything new to say, and I spend all my time
Regurgitating my ideas and sending them in. Eventually I’ll get one.” He belched.
“But wait,” I cried, “Maybe you need to stop and actually think about what
You are writing! Maybe if you took some time out, you’d frame the work
In a way that reviewers will see why it is important?” He wolfed down a page.
“No, this is how I do it,” he sputtered between mouthsful. “This is how I roll.”
“How’s that working out for you?” I asked, but he wasn’t interested.4
“Let’s go up,” said Frank. “Not much more to see here.”
We found the Second Circle. Souls were flying in the winds that blew around us.
“This is Lust,” said Frank, watching them flit around us. “Their only crime, I think,
Is to want their grant too badly. They don’t want to change it, because they love it.”
I could understand. Maybe this was my own circle. I really, really want my grant.
I lust for it. The work is lovely, and elegant, and important. But it wasn’t funded.
I could feel myself starting to float off the ground and my clothes began to flap.
“Whoa, there, Compadre,5” said Frank, grabbing my leg and bringing me to earth.
“Get a grip. I mean, its just a proposal for what you want to do. There’s nothing
of value in a grant, except as a way to get the support for the work, right?”
I wasn’t so sure. I mean, when I write a grant, I think about the project,
Really think about it, often for months. The act of writing helps me to formulate,
Consolidate my thoughts. It’s a plan for the next few years of research.
But yes, he was also right. The act of writing a grant, really, is just an approach
To supporting the work that has been formulated. Yes, thinking about it is good,
But actually doing the work is what this is all about. I needed to think.
And found that we had entered the First Circle. Limbo. Nothing to do but wait.
And while we all waited to hear the results of our submission, we were happy.
Because until we get the bad news, we can imagine that we’ll get it.
We emerged in the light of Hope. And there, before us, was a mountain.
“We’ll climb that in a while,6” said Frank. “But first, I know a good place
Near here to get some lunch. You down for that?” Me, I didn’t know down from up.
Footnotes
↵1 Mole is correct here, and despite FB’s words to the contrary, many experienced investigators have their grants fail because of a perception that “they have too much money.” Indeed, funding institutions sometimes place limits on total amounts a program may have, and in those that do not, the reviewers may take this position themselves. But Mole feels that this is inappropriate; some investigators are simply more productive than others and, given the opportunity, more support can result in substantially more impactful research from such investigators. Of course, the opposite is also true – there are investigators who will not be productive, regardless of how much support they have.
↵3 (il re o la regina di formaggio) It is possible that Mole is referring to either King Camembert (1411–1489) or, perhaps, Queen Gouda (1536–1590). Or he may be referring to his friend Weasel who is, indeed, very fond of cheese.
↵4 This approach, of flooding granting institutions with applications in hopes that one may “stick” is becoming much too common. Mole doubts that it is a viable strategy, unless such multiple applications can be done with the same care that goes into only one. As this is rarely the case, the strategy might, at least, be reconsidered. Given that the result is an avalanche of hastily prepared applications converging at any one competition, an exceptionally well-crafted (and well-reasoned) application might stand out and be favored. It’s just a thought.
↵6 The mountain, Grant Purgatory, eventually leads to Grant Paradise, which lasts only until the period of the grant runs out, and a renewal applied for. Mole points out that it may be interesting to note that, whenever a grant is rejected and we find ourselves in Grant Hell, we blame the process (“they killed me”). When it happens that we emerge into the light of funded Grant Paradise, though, we did it all by ourselves (“yes, I got the grant”).
Xenbase (www.xenbase.org) is the Xenopus bioinformatics and genomics resource. Xenopus is a major model for fundamental cell and developmental biology and a model for human disease. Xenbase is a totally free, and globally accessible database, used by Xenopus researchers worldwide, and is funded by the National Institute of Child Health and Human Development. Xenbase has two performance sites: the curation team is based in Cincinnati, OH (PI: Dr. Aaron Zorn) and the developer/database management team is based at the University of Calgary in Canada (PI: Dr. Peter Vize).
Xenbase is seeking to fill 2 full time Curation positions to join the curation team at the Division of Developmental Biology, Cincinnati Children’s Hospital, Cincinnati, OH, USA. Curation positions offer a challenging job away from the wet-lab and research bench, where interpreting, annotating and displaying complex data is our main task. Curators also develop strategies to improve data curation; work to improve data display/querying on the website; interact with our user community at research conferences; develop programming skills; and contribute to Xenbase publications.
Job Description:
Curation and annotation of published Xenopus scientific literature, focusing on gene expression and the extraction of other research data: genes, transgenics, antibodies, morpholinos, phenotypes, genetic interactions, gene product functions and models of human disease.
Import and annotate data from large-scale screens (e.g., loss-of-function morpholino screens, gain-of-function mRNA screens).
Help develop new features: curation and processing of public and directly submitted RNA-seq and ChIP-seq NGS data from Xenopus experiments, curation of mutant phenotypes and transgenics; expanding our an anatomy atlas; implementing GO annotations.
Co-author reports and publications, and give presentations at national and international meetings and workshops.
Qualifications:
MSc or PhD degree in bioinformatics and/or developmental biology, genomics, genetics, molecular biology, zoology, anatomy or related field.
Demonstrated ability to produce scientific papers, reports and presentations
Demonstrated ability to work in a team as well as independently, efficiently (i.e both quickly and accurately) and be self-motivated
Strong interpersonal and communication skills, including excellent written and spoken English
Preference will be given to applicants with:
Experience with a bioinformatics, genomics or model organism database
Experience in data annotation/biocuration, knowledge of relational databases, and familiarity with ontologies.
Experience in a Xenopus or other vertebrate (mouse, zebrafish or chick) developmental biology lab.
Experience in analyzing genomics data, using GRN software, genome browsers and common bioinformatics tools.
How to Apply:
Please submit your application, to: aaron.zorn[at]cchmc.org with the following information:
A cover letter , including a statement of interest/purpose
CV/Resume.
Copy of your degree(s).
List 3 references/referees whom we may contact (please include their postal address, email and phone number).
Salary and Start Date:
Salary will be commensurate with qualifications and experience. Start date is negotiable, but expected to be in early 2016.
The successful applicants will be employees of Cincinnati Children’s Hospital and will undergo background checks, orientation and a 3-month probationary period. Employees are required to receive an annual flu vaccination.


 (6 votes)
(6 votes)
 Atoh1 is a key regulator of the differentiation of hair cells, the sensory cells that support audition: it is upregulated during their differentiation and downregulated at postnatal stages. But what are the mechanisms underlying Atoh1 transcriptional regulation during inner ear development? To address this question, Neil Segil and co-workers (p.
Atoh1 is a key regulator of the differentiation of hair cells, the sensory cells that support audition: it is upregulated during their differentiation and downregulated at postnatal stages. But what are the mechanisms underlying Atoh1 transcriptional regulation during inner ear development? To address this question, Neil Segil and co-workers (p. 


 Mike Levine, director of the Lewis-Sigler Institute for Integrative Genomics at Princeton University, is a developmental biologist who has dedicated his career to understanding how gene expression is regulated during development. We had a lively chat with Mike at this year’s Society for Developmental Biology meeting, where he was awarded the Edwin Grant Conklin Medal. See the Spotlight article on p.
Mike Levine, director of the Lewis-Sigler Institute for Integrative Genomics at Princeton University, is a developmental biologist who has dedicated his career to understanding how gene expression is regulated during development. We had a lively chat with Mike at this year’s Society for Developmental Biology meeting, where he was awarded the Edwin Grant Conklin Medal. See the Spotlight article on p. 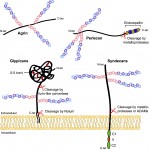 Heparan sulfate proteoglycans (HSPGs) have long been implicated in a wide range of cell-cell signaling and cell-matrix interactions, both in vitro and in vivo in invertebrate models. Here,
Heparan sulfate proteoglycans (HSPGs) have long been implicated in a wide range of cell-cell signaling and cell-matrix interactions, both in vitro and in vivo in invertebrate models. Here,  (No Ratings Yet)
(No Ratings Yet)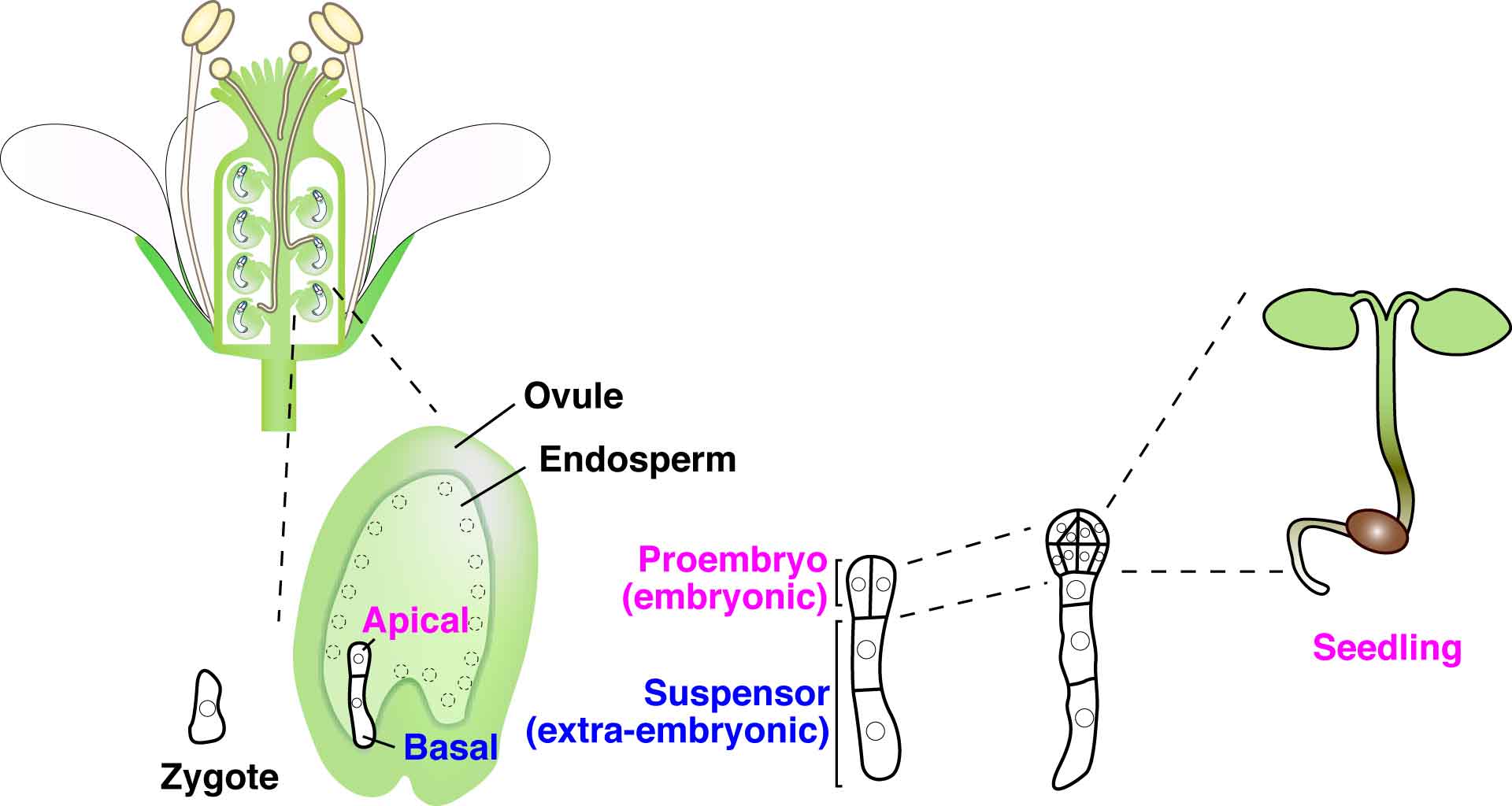



{kind=link}