The BSDB committee are inviting applications and nominations for a post doctoral representative to join the committee. We already have a Graduate Student representative and feel that the post doctoral community should also be represented. This position is available immediately and would be for a two year term and we would like someone in place by the New Year ideally. The post doc rep would attend committee meetings twice a year, one at the annual Spring BSDB-BSCB meeting and one in London in the fall. They would act as a voice and conduit for the postdoc community within the BSDB. This a great opportunity to gain insight and understanding of how a learned Society is run and could serve as a valuable addition to a CV. Please send applications and nominations to the BSDB Secretary : secretary@bsdb.org
A fully funded studentship (fees and RCUK-level stipend) open to EU students is available under the supervision of Dr Thomas Butts in the School of Biological and Chemical Sciences at Queen Mary, University of London.
The studentship is to study the evolution of neural development within vertebrates, particularly in relation to the control of neurogenesis. Within this broad area of interest in the lab, the details of the project will be dependent upon the interests of the student. More info is available at:
We are recruiting researchers at different levels to join a 4-years international project, DEVODIVERSITY, funded by the French Agence Nationale de la Recherché (ANR) and the São Paulo Research Foundation (FAPESP). The Brown Lab at the Instituto de Biociências in Brazil (USP) and the Tiozzo Lab at the Villefranche-sur-Mer Developmental Biology Laboratory in France (CNRS-UPMC) will study the evolution of regeneration, asexual reproduction, and clonality in several species of ascidians (Urochordata), and examine how ecological factors affect distribution ranges, evolution of life cycles, and developmental strategies.
DEVODIVERSITY has the following aims:
To resolve the phylogenetic relationships and evolutionary transitions between strictly sexual reproduction to budding or high regenerative abilities among Styelidae (Ascidiacea) species.
To provide a morphological and ecological understanding of asexual propagation (budding). We will generate detailed anatomical and developmental descriptions of budding processes, and explore if environmental conditions are associated with the use of particular budding modes.
To compare gene pathways involved in cell function or trans/de-differentiation processes of budding and regeneration by in silico analysis of transcriptomic data.
To launch a comparative genomic approach if styelid ascidians to better understand the evolution of major life history transitions in marine chordates, in particular the evolutionary transition from sexual to asexual propagation.
Positions available:
2 Postdoctoral positions (a2-year postdoc in France and a 3-year postdoc in Brazil): (1)candidate with experience in cell and molecular biology, some genomic experience will be favored and (2) candidate with experience in ecology, phylogenetics, and systematics.
4 Doctoral positions (two 3-year positions both in France and Brazil respectively):highly motivated candidates should have an internationally recognized Master degree.
1 Bioinformatician position (4-year position, only available in Brazil): the candidate must have strong competence in computer sciences or related areas. S/he should have either completed his/her doctorate OR should have five years of proven experience in information technology.
Several Master positions (both in France and Brazil): candidates should have completed their undergraduate degrees, and preferably with some research experience.
The successful candidates will join either the Brown lab in São Paulo (Brazil) or the Tiozzo lab in Villefranche-sur-Mer (France) and will have the opportunity to do laboratory and fieldwork in both countries.
Research position salaries and applications procedures are distinct for Brazil and France; these follow the general guidelines of either FAPESP (Brazil) or ANR (France) respectively. Please submit your letter of interest, updated CV, and contact information of three references to either Federico Brown (Brazil) or Stefano Tiozzo (France). Review of applications will begin on December 2014 and continue until the positions are filled. Start date should ideally be in early or mid-2015.
There is a fully funded PhD studentship opening in Barbara Jennings’s group at Oxford Brookes University (starting September 2015) to investigate developmental gene regulation in Drosophila.
For further details about the studentship and how to apply please see
Applications are sought for the role of Reviews Editor on The Journal of Experimental Biology, based in Cambridge, UK.
Joining an experienced and successful team, this newly created role is an exciting opportunity for a talented scientific editor to make a significant contribution to the leading journal in comparative animal physiology.
The Journal of Experimental Biology publishes papers on the form and function of living organisms at all levels of biological organisation – from the molecular and subcellular to the integrated whole animal – in an evolutionary and environmental context. In addition to core primary research articles, the journal has a well-regarded and dynamic front section of review and news content, and continues to expand its services to authors and readers.
Applicants will have a PhD in a relevant scientific field. Post-doctoral and/or previous editorial experience are highly desirable, although we will provide on-the-job training.
Core responsibilities:
– Commissioning, handling peer review and developmental editing of review material for the front section of the journal.
– Representation of the journal at international conferences and within the wider scientific community.
– Creative involvement in the journal’s development.
The successful candidate will have a broad interest in science, the scientific community and publishing. Excellent interpersonal and literary skills, enthusiasm and commitment are also essential requirements for the position.
The Reviews Editor will work alongside an experienced in-house team, including the Publishing Editor and News & Views Editor, as well as with our international team of academic editors. Additional responsibilities may be provided for the right candidate.
The position represents a unique opportunity to gain experience on a highly successful life-science journal and offers an attractive salary and benefits. The position is full time (although part-time working may be considered) and will be based in our offices in Cambridge, UK.
The Company of Biologists is a not-for-profit organisation and publishes three internationally renowned, established journals – The Journal of Experimental Biology, Development and Journal of Cell Science –as well as two newer Open Access journals – Disease Models & Mechanisms and Biology Open. The organisation has an active programme of charitable giving for the further advancement of biological research, including travelling fellowships for junior scientists and contributions to academic societies and conferences.
Applicants should send a CV, along with a covering letter that summarises their relevant experience, why they are enthusiastic about the role, and their salary expectations, by email to hr@biologists.com no later than 3 December 2014. Please direct informal enquiries to Miriam Ganczakowski on +44 (0)1223 426164.
Applicants must be eligible to work in the UK and be able to travel internationally.
The 2014 Society for Developmental Biology (SDB) Lifetime Achievement Award was jointly awarded to Christopher Wylie and Janet Heasman in recognition of their outstanding and sustained contributions to the field. At the 73rd Annual SDB meeting, where they were presented with the award, we asked Chris and Janet about their careers and their advice for young researchers.
You both started off in medical school and then switched to a career in basic research. What inspired you to make the switch?
C: Actually,much of it was due to one of my professors, J. Z.Young, who did several things that enhanced basic science in medicine. Because students enter medical school directly from high school in England, they do not get exposed to research. J. Z. introduced a BSc-level science degree into the medical school curriculum. This was offered to students who did well in their medical examinations, and also served as entry into the MD/PhD program. Both Janet and I went with this route and just fell in love with biomedical research. A major part of the BSc course was a research project. I did mine with Ruth Bellairs and became fascinated by embryo development. I tried to make a movie of one of the experiments that we were doing but, in those days, making movies of embryos was a very complicated business. The movie (of a chick embryo) I was making went on all night, so I had to sit in the hot room for hours (there were no heated slide chambers in those days). Because of the heat, the grease in the microscope would melt and the body of the microscope would slowly slip down the column, so it had to be refocused after each image. By about 3 o’clock in the morning I was virtually naked and dripping with sweat! But I couldn’t believe my eyes as more and more embryonic structures appeared, as if by magic. Of course I had seen diagrams of development in my lectures, but I had never actually seen it happening. I was absolutely gobsmacked! That was my epiphany. And so I finished my BSc and was lucky enough to get offered a PhD place. Of course, the original objective was to then return to clinical medicine but I never did. I was offered a lectureship at University College, London, and that was the end of my clinical career.
J: I even did a year of clinical training (in those days medical school was two years of preclinical training and then three years of clinical school) but I didn’t enjoy it. I had the chance to go to Dartmouth College and work as a teaching assistant with Chris in developmental biology. While I was there I gave three lectures and I remember thinking to myself, if I can do this – if I can teach developmental biology successfully – then I am going to go back to England, pull out of my medical career and register as a PhD student. And that is what I did.
You’ve both been involved in setting up developmental biology institutes – in Cambridge, Minnesota and Cincinnati – and these are all continuing to flourish. What do you think is the key to success in a good research department or institute?
C: I think you have to have the right people…and money! If you recruit scientists who are willing to interact with each other and not compete, then you’ll probably have a successful institute, especially if it is well funded. Another thing that might seem trivial but proved to be very important was the tea room! It offered a centre where principals, group leaders, graduate students, postdocs and technicians could all meet and talk informally. Every morning at 10.30 at the Wellcome/CRC Institute in Cambridge (now the Gurdon Institute) we would all go up to the tea room for tea/coffee and chat. The things that we chatted about were not just ‘do you have an antibody against x that we can borrow?’, it was also ‘how do you think the institute could work better?’, ‘how can we improve the vivarium?’, ‘how is our financial situation?’. This day-by-day interaction was amazingly helpful for all aspects of the institute’s activities, not just our own research. It certainly made faculty meetings shorter because we had already discussed pretty well everything that came up.
J: Yes, the tea room worked really well in England.We tried very hard to make it work in the USA, but it was hard to persuade people to go to the tea room. No one would allow themselves enough time off to interact over tea! I also agree that the success of an institute depends on having the right people and being very careful in the job searches. It is absolutely essential to hire people who are interactive and who are going to collaborate with each other. It’s also important to have freedom – the freedom to be able to do science the way you want.
C: Yes, in Cambridge, the institute was funded by the Wellcome Trust and the Cancer Research Campaign, and they gave us a free hand. So we were largely buffered against Cambridge University politics and departmental structure. Similarly, in Minnesota I was just given the money and told ‘we need a centre for developmental biology’. And in Cincinnati I was given an even larger amount of money and told ‘build us a centre for developmental biology, Chris’. When you have such a financial advantage, it is not that difficult.
Chris, you have obviously been heavily involved with Development: you were Editor-in-Chief for a very long time and you made some pretty drastic changes to the journal when you started. How did you get involved with the journal and what prompted you to make those bold changes?
C: It all began when I was the Publications Officer of the British Society for Developmental Biology (BSDB). My job was to think up suitable symposium topics and identify people who could put a symposium together, edit the volume and to see it through to publication. At first, the papers presented at these symposia were published by Cambridge University Press as books, but the publication times were long and the costs were very high (and the profits to the society correspondingly small). So the BSDB moved to publishing their symposia as supplement volumes to The Journal of Embryology and Experimental Morphology (JEEM), which was owned and published by The Company of Biologists. The symposia attracted outstanding speakers (and therefore authors) from all over the world, and I couldn’t help noticing that the articles in the symposium supplements were generally more exciting than those in the parent journal. I thought, we are missing a lot of stuff in our journals in the UK. We need a journal that will capture the most recent advances in cellular, molecular and genetic approaches to developmental biology. So I went to several publishers and I said that I think there should be a journal (I had already called it Development in my mind) that captures all this stuff that the current British journals are missing. Lots of publishers took an interest in this. I was then approached by The Company of Biologists (who had heard about these discussions) and asked if I would take over the editorship of JEEM. I told them that I didn’t want to be the editor of JEEM but that I would consider being the editor of a different journal, one that had all the things that I thought should go into a topical developmental biology journal, and with a more modern format. Eventually they agreed. They said ‘Okay, do whatever you want.We will start a new journal, we will replace JEEM with this journal, and you can make whatever changes you want!’. And so I had a journal – Development – on my hands. It was absolute mayhem initially – it was probably June when we started and we had to have the issue out by January the following year – but, mercifully, the world’s developmental biologists reacted magnificently. I guess everyone else thought that there should be a journal like that, too.
That was over 25 years ago. How do you think the field has changed since then?
C: Well, I think the science – especially in terms of techniques and approaches – is changing all the time. In just one year, since I’ve retired, I can see by coming back to this meeting that there are techniques that we weren’t using just last year when I was in the lab.
J: Another big change is that everything is now online. The fact that no one uses libraries anymore has changed publishing, and it has changed interactions between people and groups.
C: It also means that, to some extent, it doesn’t really matter anymore where you publish, because people don’t go to libraries and open journals and go through them. They use searches and keywords so it doesn’t really matter whether your paper is published in what was formerly regarded as a ‘second tier’ journal or a premier journal; if your paper is good, it is going to be read.
J: And it is going to be reviewed in a very informal sense too – online. There are big pluses to that but there are also some minuses because sometimes only the people with the loudest ‘voices’ will be heard. Distinguishing gossip from good science is becoming more and more difficult these days.
C: Another thing that has changed a lot is the number of model organisms studied. When I was a student there were at least a dozen experimental model organisms being used to study development.We needed to learn their anatomy and embryogenesis to keep up with the genetic, molecular and cellular data coming out. However, as both the time and money required to set up the appropriate protocols and reagents became so great, the number of model organisms used contracted dramatically. So, during the time that I was an editor at Development,we ended up going from a dozen or so model organisms that made regular appearances in its pages to just four or five.
J: But now it has expanded again, which is really exciting. Techniques such as CRISPR have allowed us to go the other way. It was very noticeable at this meeting that there are again a variety of model organisms being used. Evolution is at the centre again.
Science seems to be quite competitive these days, and it is getting increasingly difficult to get an academic tenure track position. What is your advice to young people who are starting out?
J: To recognise that it is very important to develop your own ideas of what research you want to do, even as a graduate student; don’t wait until the postdoc stage. Actively think about where you want your career to go and look for the labs that will fit what you want to do, rather than looking for the lab that will take you. You will be impressive in an interview if you know what you want to do. Rather than thinking ‘how am I going to finish my graduate studies?’, ‘how am I going to get my first fellowship?’ and so on, think ‘what am I really interested in answering?’, ‘what biological question do I want to answer?’ and ‘which model system should I use?’.
C: I think you have to choose the right postdoc position. This is really, really important. Also, you need to find good mentorship. That applies at all levels. If you don’t have a good PhD mentor, who will look after your next stage by being honest about labs that you should or should not go to as a postdoc, you are at a serious disadvantage. Similarly, you need a good postdoc mentor whowill advise you about which places you should think about going to for getting a job (and where you should not think about going to!). And then, when you are an assistant professor, you need really good mentorship for all aspects of your research life, including: research approaches, collaborations that will help you, grant applications, recruitment of lab personnel, disputes that occur in your lab, where you should send your papers, what committees you should (and shouldn’t) sit on.Young faculty are not ready for that kind of stuff, and they need senior faculty to act as good mentors and who can offer them advice, read papers and grants, and provide honest feedback.
You’ve both had very successful careers working as a husband-and-wife team while raising a family. What is your advice to younger people who are starting off in terms of work-life balance?
J: I think my advice is just to do it. There is no easy way. And that is true whether you are a scientist or working in banking or business. It is a balancing act that requires constant communication. I think scientists in some ways have it easy if they work together. I can always say I know where Chris is – he is in the lab! I know how to contact him there. I actually think that science is a good career for husband and wife teams. And don’t take yourself, and science, too seriously; at the end of the day you are a small cog in a big wheel and your kids carry on after you.
C: And be prepared to work shifts. We used to do that a lot.
J: And my advice to young women is: don’t put off having kids until the time is right. Because the time is never right.
C: The problem with putting off kids until the ‘right time’ is that you have to set criteria for the right time, and it is hard to know when these have been met. It is easy to say ‘we will have children when…’. But of course you never reach the ‘when’.
And, finally, what would people be surprised to find out about you?
C: I don’t think there is much about me that would surprise people. I think the basis for my success in editorship and leadership has been that I’m not a particularly complicated or devious person. So nobody ever needed to try to second guess me! I suppose that, given the public persona required to teach, lead a research centre, or edit a major journal, people might be surprised to know that I’mnot a gregarious person. The best part of my career has been working quietly in the lab, trying to discover things, reading about the discoveries others have made (I really miss the library), and trying to integrate it all into new knowledge.
Here are the highlights from the new issue of Development:
Akt acts to reprogram germ cells
Primordial germ cells (PGCs) are unipotent – they go on to form germline stem cells and gametes. However, they are believed to possess latent pluripotency, allowing them to produce the next generation during the normal life-cycle, and to be reprogrammed to pluripotent embryonic germ cells (EGCs) upon experimental manipulation. Various protocols for EGC establishment have been reported, with varying efficiency. Yasuhisa Matsui and colleagues now report a highly efficient method for mouse PGC-to-EGC conversion, using Akt activation in concert with bFGF and LIF treatment (p. 4457). Using a relatively simple protocol starting from purified E10.5 PGCs in culture, the authors are able to achieve 60% reprogramming efficiency, which is significantly higher than any previous method. These reprogrammed EGCs readily contribute to mouse chimeras, including the germline. The authors suggest that Akt may act by inhibiting apoptosis of PGCs and/or by promoting signalling events (including via bFGF and LIF) that mediate PGC-to-EGC reprogramming, thus inducing the latent pluripotency of early PGCs.
Cerebral haemorrhage without leaky vessels
The vasculature of the central nervous system (CNS) is highly specialised, characterised by the formation of the blood-brain barrier that prevents leakage of vascular contents into the brain. Various molecules and pathways have been implicated in regulating angiogenesis in the CNS, including the αVβ8 integrin and members of the TGFβ pathway. It is thought that αVβ8 integrin expressed in the neuroepithelium regulates TGFβ signalling in the endothelium. On p.4489, Thomas Arnold, Louis Reichardt and colleagues set out to investigate this relationship and the effects of disrupting this signalling cascade on the CNS vasculature. The authors find that disruption of either the integrin or TGFβ pathway components leads to excess angiogenic sprouting, vascular dysplasia and cerebral haemorrhage. Importantly, however, these mutant mice display no signs of compromised blood-brain barrier formation and the vessels, although abnormal, are not leaky. Together, this work defines an important function for αVβ8 integrin-TGFβ signalling in limiting vascular sprouting in the CNS, and demonstrates that cerebral haemorrhage can result from abnormal angiogenesis rather than from blood-brain barrier breakdown.
Restricting liver progenitor potential
The liver possesses a remarkable capacity to regenerate, but what is the source of new cells during regeneration? The two epithelial cell populations – hepatocytes and cholangiocytes – can proliferate upon injury, but there is also evidence for the existence of an adult stem/progenitor cell, the liver progenitor cell (LPC), which resides in or near bile ducts and has both hepatocytic and cholangiocytic potential. Here (p.4448), Naoki Tanimizu and co-workers investigate the abundance and potential of LPCs from neonatal and adult mice. They find that the number of LPCs decreases during postnatal life, and also observe a change in their differentiation potential: adult LPCs are strongly biased towards the cholangiocyte fate and have very limited ability for hepatocytic differentiation, both in vitro and in vivo. Mechanistically, the authors show that the transcription factor grainyhead-like 2 (GRHL2), which is known to promote cholangiocyte differentiation, is more strongly expressed in adult LPCs than neonatal LPCs. GRHL2 in turn inhibits expression of miRNA122, which is important for the hepatocyte lineage. Thus, upregulation of GRHL2 in adult LPCs provides an explanation for their reduced hepatocytic potential, and its inhibition may help to produce functional hepatocytes in the adult liver.
The origin of the coronary vasculature
During heart development, the coronary vasculature forms by establishment of an endothelial plexus that expands around the heart. Current evidence suggests that the sinus venosus (SV), endocardium and proepicardium may all contribute to coronary development. However, the relative contributions of these sources and the molecular mechanisms regulating coronary angiogenesis are still unclear. In a detailed lineage-tracing analysis in mouse (p. 4500), Kristy Red-Horse and colleagues find that the SV and endocardium make spatially restricted and complementary contributions to the coronary vasculature, with dorsal and lateral vessels having primarily SV origin, while the endocardium contributes significantly to vessels of the ventral heart and ventricular septum. The proepicardium makes a minor, but non-spatially restricted, contribution. VEGFA has previously been shown to promote angiogenesis of a subpopulation of coronary vasculature, and the authors now find that VEGFC promotes growth of a complementary set of vessels – those derived from the SV. These data provide a comprehensive view of the sources of the coronary endothelium and help to unravel the mechanisms by which these vessels form.
PLUS:
An interview with Christopher Wylie and Janet Heasman
The 2014 Society for Developmental Biology (SDB) Lifetime Achievement Award was jointly awarded to Christopher Wylie and Janet Heasman in recognition of their outstanding and sustained contributions to the field. At the 73rd Annual SDB meeting, where they were presented with the award, we asked Chris and Janet about their careers and their advice for young researchers. See the Spotlight on p. 4415
How to make a cardiomyocyte
Kenneth Chien and colleagues discuss how insights into the molecular and cellular framework underlying cardiac development can be used to guide the in vitro specification of cardiomyocytes, whether by directed differentiation of pluripotent stem cells or via direct lineage conversion. See the Primer article on p. 4418
The cellular and molecular mechanisms of vertebrate lens development
Although lens induction has been studied for over 100 years, recent findings have revealed a myriad of signaling pathways and gene regulatory networks that are required for lens formation in vertebrates. Ales Cvekl and Ruth Ashery-Padan summarize recent progress in the field, emphasizing the interplay between the diverse regulatory mechanisms employed to form lens progenitor and precursor cells. See the Review on p. 4432
The Minnesota Center for Philosophy of Science in the College of Liberal Arts at the University of Minnesota invites applications for a full-time postdoctoral position in conjunction with the project “Integrating Generic and Genetic Explanations of Biological Phenomena,” sponsored by the John Templeton Foundation. This project brings together scholars in philosophy and biology to address the problem of integrating different types of explanations from scientific approaches to bridge the divide between genetic and generic or physical explanatory approaches for three domains of biological phenomena: development, evolutionary novelty, and evolvability (for more details, see: http://genericgenetic.umn.edu/). Position duration: 03/01/2015 to 08/31/2017.
Applicants should have a Ph.D. or equivalent terminal degree in the biological sciences with a background in cellular and molecular development, demonstrated interest in theoretical issues in development and evolution, and evidence or promise of scholarly achievement. Familiarity with computational modeling methods is preferred. The successful candidate will: (a) work with the core team members (http://genericgenetic.umn.edu/people/core-team) on conceptual aspects of integrating genetic and generic or physical explanatory approaches, and (b) carry out an experimental program focusing on the relationship between the gene expression networks and physical dynamics involved in limb pattern formation with the aim of identifying integrated models that provide necessary conditions for the development and evolution of distinctive digit phenotypes in vertebrates.
We recently demonstrated an improved method for homologous recombination (HR)-mediated genome editing using TALEN (Transcription activator-like effector nuclease) in zebrafish (Shin et al., 2014). In the study, we identified that a total of 3kb of homology in the targeting construct, 1kb of one arm and 2kb of the other arm, is sufficient to induce HR-mediated knock-in. Importantly, our results suggest that a specific targeting construct configuration (a double stranded break in the short homology arm) can enhance the efficiency of HR-mediated knock-in. I believe that the method is highly valuable due to its fundamental advantage, which is the ability to manipulate in vivo genome precisely. In this post, I discuss several potential applications using HR-dependent genome engineering.
Potential applications using HR-mediated genome engineering
1. Tagging fluorescent protein
HR-mediated gene targeting technology allows us to precisely deliver a relatively large DNA fragment encoding a fluorescent protein into a specific locus of the chromosome. For example, HR dependent GFP knock-in alleles will be useful to monitor the spatiotemporal localization of a target protein in vivo. Although there is a potential caveat such as altered protein functions due to tagging, this method allows visualization of tagged proteins in the physiologically relevant condition rather than overexpression context. Tagging a flouorescent protein along with biotinylation tag, such as avitag, will have an additional capacity to study protein-protein interactions. Because in vivo BirA-Avitag system (de Boer et al., 2003) can carry the advantage of mass spectrometry (MS) analysis, the resulting knock-in lines will be useful for tracing the tagged target gene product as well as screening for novel protein binding partners.
2. Mirroring gene expression
As shown in our paper, the expression of a target gene can be monitored by precisely inserted sequences encoding a fluorescent protein that is linked with ‘self-cleaving’ 2A peptides just in front of the a target gene’s stop codon. I believe this strategy will allow tracing of specific cells, which express both a target gene and a fluorescent reporter protein, without functional alterations of the gene products. In addition to this, the physical separation of a target gene product and a fluorescent reporter protein allows for independent protein behaviors, which is important for diversifying methodologies. For example, application of this method with Cre recombinase instead of a fluorescent reporter will result in Cre knock-in lines that can be used to generate tissue specific Cre driver lines for conditional knock-out mutants. Alternatively, specific cell types can be permanently labeled by crossing the ubi:Switch transgenic line (Mosimann et al., 2011) with Cre knock-in lines. Moreover, replacement of the bacterial biotin ligase BirA by a fluorescent reporter will serve as a biotinylation driver for tissue or cell specific gene expression profiling (Housley et al., 2014).
3. Generation of floxed alleles
To create conditional knock-out lines, a gene of interest must be modified by the insertion of two loxP sites to ecise the floxed exon(s) via Cre-mediated recombination. Although the insertion of a loxP site in the target locus can be accomplished by using loxP site containing single-stranded oligonucleotides (ssONs), insertion of two loxP sites can pose a challenge due to low efficiency of ssONs knock-in. Therefore, using a combination of two loxP site containing targeting constructs with TALEN is an useful and efficient strategy to obtain floxed alleles.
I believe that many other applications using HR-mediated genome engineering are already developed or will be invented for innovative research. Hence, our efficient HR-dependent knock-in method will allow many laboratories to generate multi-purpose knock-in lines and serve as a platform for development of more sophisticated genome manipulation methods.
Reference article:
de Boer, E., Rodriguez, P., Bonte, E., Krijgsveld, J., Katsantoni, E., Heck, A., Grosveld, F., & Strouboulis, J. (2003). Efficient biotinylation and single-step purification of tagged transcription factors in mammalian cells and transgenic mice Proceedings of the National Academy of Sciences, 100 (13), 7480-7485 DOI: 10.1073/pnas.1332608100
Housley, M., Reischauer, S., Dieu, M., Raes, M., Stainier, D., & Vanhollebeke, B. (2014). Translational profiling through biotinylation of tagged ribosomes in zebrafish Development, 141 (20), 3988-3993 DOI: 10.1242/dev.111849
Mosimann, C., Kaufman, C., Li, P., Pugach, E., Tamplin, O., & Zon, L. (2010). Ubiquitous transgene expression and Cre-based recombination driven by the ubiquitin promoter in zebrafish Development, 138 (1), 169-177 DOI: 10.1242/dev.059345
Shin, J., Chen, J., & Solnica-Krezel, L. (2014). Efficient homologous recombination-mediated genome engineering in zebrafish using TALE nucleases Development, 141 (19), 3807-3818 DOI: 10.1242/dev.108019
The morphological evolution of limbs and external genitalia were both essential adaptions to a life on land. While the former deals with the novel locomotory challenges facing an animal invading a terrestrial environment, the latter is concerned with something even more essential: reproduction! Living on land means that gametes can no longer be fertilized externally simply by releasing them in water, e.g. as frogs do. Male and female gametes need to be brought together as well as protected from dehydration. Internal fertilization, utilizing specialized external genitalia, solves this dilemma, delivering sperm to their target and hence providing protection inside the animal’s body.
In addition to the adaptive value of limbs and external genitalia in the context of the transition to land, there is also a striking similarity in the patterning genes expressed during the development of these two structures. For both these reasons, a potential co-evolution of limbs and genitals has been discussed in the past1-4. In our recent Nature paper we have identified another, unexpected link between the limbs and external genitalia that may help to explain some of the molecular similarities between the two5.
From Limbs to Genitals
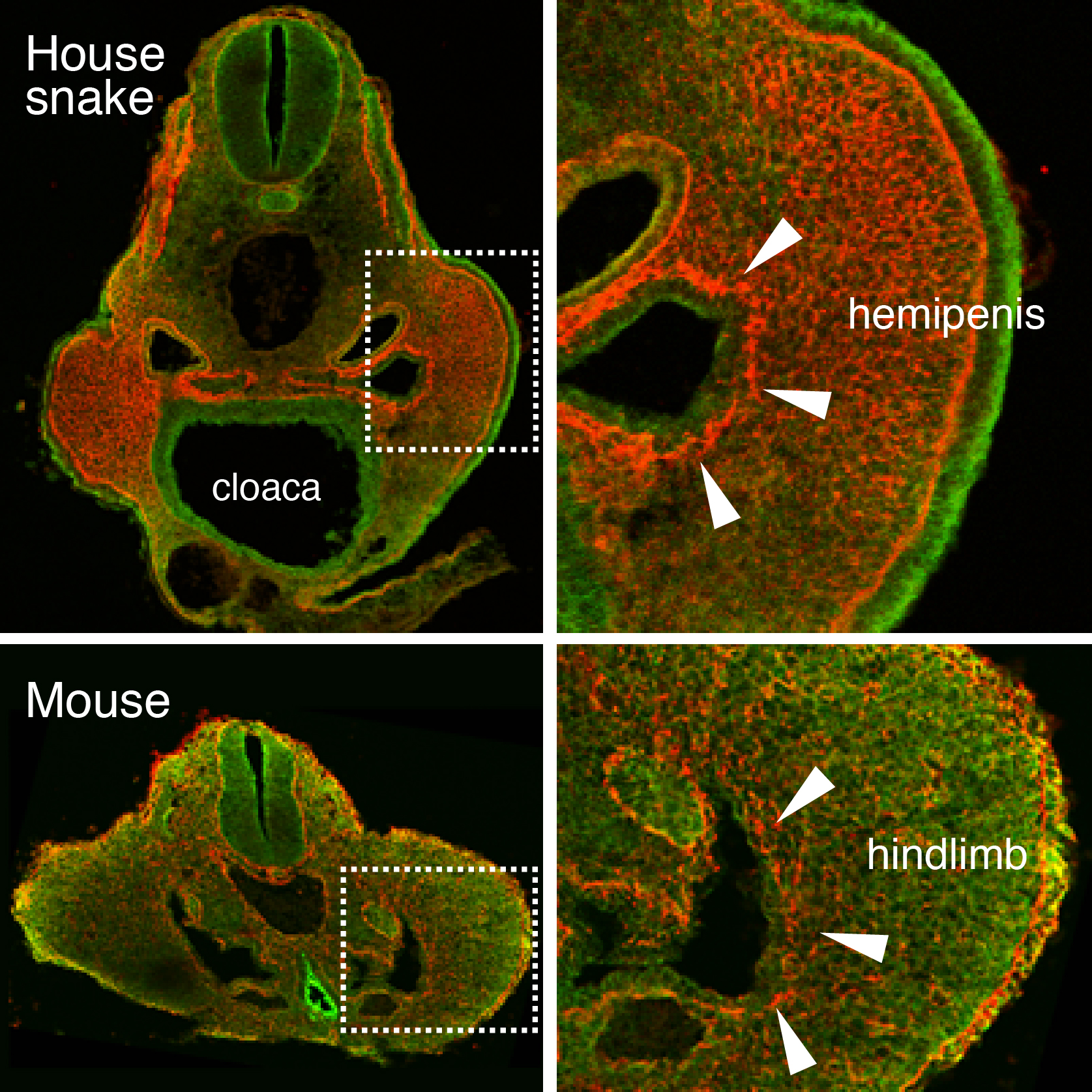
As might be expected for Cliff Tabin’s lab, this project also started out as a study focusing on limb development. At the time, Jérôme Gros (a former postdoc in the lab, now PI at Institut Pasteur, Paris) was investigating the earliest steps of limb initiation in vertebrate embryos. He found that in both mouse and chicken embryos the mesenchyme of the growing limb buds originates from the epithelial lateral plate mesoderm (LPM), through a process called epithelial-to-mesenchymal transition, or EMT6. Given the interest of our lab in evolutionary questions, former graduate student Jimmy Hu suggested looking for the presence or absence of the same process in the limb-less snake….of which he just happened to have a few embryos in his freezer – courtesy of Olivier Pourquié who had previously brought to our attention the presence of outgrowths at a hindlimb-like position. There was no evidence for such EMT at the level where forelimbs once were supposed to form. However, when looking at the hindlimb level, a clear EMT was visible – only in snakes the resulting mesenchymal cells of this “limb-like” bud contributed to their budding external genitalia, the so-called hemipenes (Figure 1). Such similarity in limb and genital bud initiation in squamates led us to question the developmental origin of external genitalia in other amniotes.
Figure 1. An epithelial-to-mesenchymal transition (EMT) underlies the developmental initiation of the house snake hemipenis and the mouse hindlimb bud. Breakdown of the basement membrane (laminin staining in red) is seen in both embryos (arrowheads).
Tracing the Origin of External Genitalia
Squamates (snakes and lizards) are interesting in this regard, as within their clade partial or complete loss of limbs occurred among several species, yet they all keep their hemipenes. Using micro-computed tomography in collaboration with Emma Sherratt (now at University of New England, Australia), we realized that all squamate embryos formed their hemipenis buds at the same level where hindlimbs would form, whereas the mouse genitalia were emerging more posteriorly, towards the tail (see video). Moreover, we were able to visualize the internal location of the cloaca, an endodermal signaling center known to be important for genital outgrowth. Intriguingly, the cloaca seems similarly shifted in all squamate embryos, into the presumptive hindlimb field. This prompted us to determine which cell populations actually give rise to the different species’ genitalia – a question that seemed far from settled, when consulting the available literature.
Using a lentiviral lineage tracing system we were able to demonstrate that, indeed, important differences exist in the developmental origin of external genitalia among different species: whereas the mouse genital tubercle is built mostly of tailbud-descendant cells, the Anolis lizard hemipenis consists of cells from the same embryonic lineage that gives rise to its hindlimbs. In both species the external genitalia thus seemed to “follow” the localized signaling of the cloaca. This suggested the possibility that the evolutionary change in cell populations forming the external genitalia could, at least in part, be attributable to a shift in the relative position of the cloaca.
ADeep Homologyof Vertebrate Genitalia
An organ’s transcriptional signature is influenced by its developmental origin, yet can also give hints about evolutionary relations to other tissue types7,8. We therefore performed comparative RNA-seq analyses on early and late budding stages of limbs and genitalia, in both lizard and mouse embryos. Working in distantly related species, while considering similar tissue types, comes with its own set of problems when performing comparative transcriptomic studies – however, after a somewhat rugged start, it soon became clear that the Anolis limb and genitalia transcriptomes show a much higher degree of overall similarity, than was the case for the mouse samples. Also, at early stages, the Anolis hemipenis transcriptome is virtually indistinguishable from a generic limb molecular signature, and only later differentiates into a genitalia-like state. This confirmed, at a molecular level, the relatedness of the cells building limbs and genitalia in the Anolis lizard. Moreover, by grafting the cloacal signaling center into chicken limb buds, we were able to partially induce transcriptional changes reminiscent of early genitalia development, demonstrating the conserved ability of limb cells to respond to these cloacal signals, and supporting the idea that change in the location of the cloaca would have induced a similar genetic program in a different target tissue.
This study offers a potential explanation for the still striking similarities in gene expression in species that develop limbs and genitalia from discrete cell populations4 – namely, that a limb-derived state could represent the ancestral condition for the emergence of external genitalia. As such, a limb-like gene regulatory network for genitalia growth might have become hardwired in a putative ancestral genome. The genitalia of mice and lizards, while not homologous to one another sensu stricto, might thus represent an example of Deep Homology9: with homology in the genetic programs being executed and induced by the same ancestral signaling source, the cloaca.
1. Kondo T, Zákány J, Innis JW, & Duboule D (1997). Of fingers, toes and penises. Nature, 390 (6655) PMID: 9363887
2. Yamada, G., Suzuki, K., Haraguchi, R., Miyagawa, S., Satoh, Y., Kamimura, M., Nakagata, N., Kataoka, H., Kuroiwa, A., & Chen, Y. (2006). Molecular genetic cascades for external genitalia formation: An emerging organogenesis program Developmental Dynamics, 235 (7), 1738-1752 DOI: 10.1002/dvdy.20807
3. Cohn, M. (2011). Development of the external genitalia: Conserved and divergent mechanisms of appendage patterning Developmental Dynamics, 240 (5), 1108-1115 DOI: 10.1002/dvdy.22631
4. Lin, C., Yin, Y., Bell, S., Veith, G., Chen, H., Huh, S., Ornitz, D., & Ma, L. (2013). Delineating a Conserved Genetic Cassette Promoting Outgrowth of Body Appendages PLoS Genetics, 9 (1) DOI: 10.1371/journal.pgen.1003231
5. Tschopp, P., Sherratt, E., Sanger, T., Groner, A., Aspiras, A., Hu, J., Pourquié, O., Gros, J., & Tabin, C. (2014). A relative shift in cloacal location repositions external genitalia in amniote evolution Nature DOI: 10.1038/nature13819
6. Gros, J., & Tabin, C. (2014). Vertebrate Limb Bud Formation Is Initiated by Localized Epithelial-to-Mesenchymal Transition Science, 343 (6176), 1253-1256 DOI: 10.1126/science.1248228
7. ARENDT, D. (2005). Genes and homology in nervous system evolution: Comparing gene functions, expression patterns, and cell type molecular fingerprints Theory in Biosciences, 124 (2), 185-197 DOI: 10.1016/j.thbio.2005.08.002
8. Wagner, G. (2007). The developmental genetics of homology Nature Reviews Genetics, 8 (6), 473-479 DOI: 10.1038/nrg2099
9. Shubin, N., Tabin, C., & Carroll, S. (2009). Deep homology and the origins of evolutionary novelty Nature, 457 (7231), 818-823 DOI: 10.1038/nature07891
 (2 votes)
(2 votes) (No Ratings Yet)
(No Ratings Yet)
 Primordial germ cells (PGCs) are unipotent – they go on to form germline stem cells and gametes. However, they are believed to possess latent pluripotency, allowing them to produce the next generation during the normal life-cycle, and to be reprogrammed to pluripotent embryonic germ cells (EGCs) upon experimental manipulation. Various protocols for EGC establishment have been reported, with varying efficiency. Yasuhisa Matsui and colleagues now report a highly efficient method for mouse PGC-to-EGC conversion, using Akt activation in concert with bFGF and LIF treatment (p.
Primordial germ cells (PGCs) are unipotent – they go on to form germline stem cells and gametes. However, they are believed to possess latent pluripotency, allowing them to produce the next generation during the normal life-cycle, and to be reprogrammed to pluripotent embryonic germ cells (EGCs) upon experimental manipulation. Various protocols for EGC establishment have been reported, with varying efficiency. Yasuhisa Matsui and colleagues now report a highly efficient method for mouse PGC-to-EGC conversion, using Akt activation in concert with bFGF and LIF treatment (p.  The vasculature of the central nervous system (CNS) is highly specialised, characterised by the formation of the blood-brain barrier that prevents leakage of vascular contents into the brain. Various molecules and pathways have been implicated in regulating angiogenesis in the CNS, including the αVβ8 integrin and members of the TGFβ pathway. It is thought that αVβ8 integrin expressed in the neuroepithelium regulates TGFβ signalling in the endothelium. On p.
The vasculature of the central nervous system (CNS) is highly specialised, characterised by the formation of the blood-brain barrier that prevents leakage of vascular contents into the brain. Various molecules and pathways have been implicated in regulating angiogenesis in the CNS, including the αVβ8 integrin and members of the TGFβ pathway. It is thought that αVβ8 integrin expressed in the neuroepithelium regulates TGFβ signalling in the endothelium. On p. The liver possesses a remarkable capacity to regenerate, but what is the source of new cells during regeneration? The two epithelial cell populations – hepatocytes and cholangiocytes – can proliferate upon injury, but there is also evidence for the existence of an adult stem/progenitor cell, the liver progenitor cell (LPC), which resides in or near bile ducts and has both hepatocytic and cholangiocytic potential. Here (p.
The liver possesses a remarkable capacity to regenerate, but what is the source of new cells during regeneration? The two epithelial cell populations – hepatocytes and cholangiocytes – can proliferate upon injury, but there is also evidence for the existence of an adult stem/progenitor cell, the liver progenitor cell (LPC), which resides in or near bile ducts and has both hepatocytic and cholangiocytic potential. Here (p. During heart development, the coronary vasculature forms by establishment of an endothelial plexus that expands around the heart. Current evidence suggests that the sinus venosus (SV), endocardium and proepicardium may all contribute to coronary development. However, the relative contributions of these sources and the molecular mechanisms regulating coronary angiogenesis are still unclear. In a detailed lineage-tracing analysis in mouse (p.
During heart development, the coronary vasculature forms by establishment of an endothelial plexus that expands around the heart. Current evidence suggests that the sinus venosus (SV), endocardium and proepicardium may all contribute to coronary development. However, the relative contributions of these sources and the molecular mechanisms regulating coronary angiogenesis are still unclear. In a detailed lineage-tracing analysis in mouse (p.  The 2014 Society for Developmental Biology (SDB) Lifetime Achievement Award was jointly awarded to Christopher Wylie and Janet Heasman in recognition of their outstanding and sustained contributions to the field. At the 73rd Annual SDB meeting, where they were presented with the award, we asked Chris and Janet about their careers and their advice for young researchers. See the Spotlight on p.
The 2014 Society for Developmental Biology (SDB) Lifetime Achievement Award was jointly awarded to Christopher Wylie and Janet Heasman in recognition of their outstanding and sustained contributions to the field. At the 73rd Annual SDB meeting, where they were presented with the award, we asked Chris and Janet about their careers and their advice for young researchers. See the Spotlight on p. 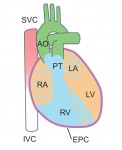 Kenneth Chien and colleagues discuss how insights into the molecular and cellular framework underlying cardiac development can be used to guide the
Kenneth Chien and colleagues discuss how insights into the molecular and cellular framework underlying cardiac development can be used to guide the  Although lens induction has been studied for over 100 years, recent findings have revealed a myriad of signaling pathways and gene regulatory networks that are required for lens formation in vertebrates. Ales Cvekl and Ruth Ashery-Padan summarize recent progress in the field, emphasizing the interplay between the diverse regulatory mechanisms employed to form lens progenitor and precursor cells. See the Review on p.
Although lens induction has been studied for over 100 years, recent findings have revealed a myriad of signaling pathways and gene regulatory networks that are required for lens formation in vertebrates. Ales Cvekl and Ruth Ashery-Padan summarize recent progress in the field, emphasizing the interplay between the diverse regulatory mechanisms employed to form lens progenitor and precursor cells. See the Review on p.