The 1st International Symposium on Women in Tunicate Biology, organized by Anna Di Gregorio and Marie Nydam, was held online on March 28-29, 2023. This global symposium was attended by 50 researchers from several countries, including Austria, Brazil, India, Italy, Japan, New Zealand, Turkey, and the United States. The 35 manuscripts in this collection include “tribute” papers that honor women scientists who pioneered and advanced the field of tunicate biology, as well as “In Her Words” letters, which provided a canvas for women scientists to freely describe their research and themselves. The proceedings are published in a Special Issue in genesis: The Journal of Genetics and Development. All of the articles in this Special Issue will be freely accessible on the genesis website for a period of 3 months.
This special Issue is expected to become a resource for all scientists interested in tunicate biology, a reference for early and contemporary work in this field of science, and an inspiration for all women scientists. All of the major fields of tunicate biology are represented, including developmental and stem cell biology, regeneration biology, ecology and taxonomy.
This is an excerpt of the Editorial written by James Briscoe and Katherine Brown, published in Volume 151, Issue 1 of Development.
The start of a new year is often a time to reflect and take stock. As researchers (and editors) we are usually so involved in the day-to-day challenges of our jobs that we lose sight of the bigger picture. But stepping back, it is extraordinary to see how much has changed over the last few years in scientific publishing and in the journals we are all familiar with. New business models, innovations in peer review and the rise of preprints are all having a huge impact, and the rise of Artificial Intelligence seems likely to revolutionise the research and publishing ecosystems (for good and bad) in ways we are only beginning to imagine. Of course, the primary role of journals continues to be to publish research findings and disseminate their conclusions, and broader developments in the field, to the wider scientific community. But the impact of journals like Development (and its sister journals at The Company of Biologists) extends further. By organising conferences and workshops, we help connect researchers and enable new collaborations. By awarding travelling fellowships and promoting the next generation of researchers, we support the next wave of innovation. By hosting and managing forums such as the Node, preLights and Focal Plane, we facilitate dialogue and the exchange of ideas and resources. These community-building efforts are possible only because we are a not-for-profit journal run by scientists for scientists. But, like all journals, we rely on our authors and the papers you send us. Without authors there would be no journal at all, and we couldn’t support the field in the ways that we do. Sometimes with all the demands and pressures on us, we lose sight of this broader perspective, and our choice of journal is driven by factors such as what we think would impress others, or where we think we might get an ‘easy ride’. However, where you choose to send your paper is not a neutral decision: publishing is political. By choosing to send your next paper to Development you are demonstrating your support for a not-for-profit scientist-led journal, and you are signifying your commitment to the field and to the next generation of researchers. It is only with your backing that we can continue supporting discovery in developmental and stem cell biology for years to come. So as you plan your 2024 submissions, we ask you to choose Development.
Last year saw the start of Development’s Pathway to Independence (PI) programme (Briscoe and Brown, 2022). We all recognise that finding a job in academia and setting up a lab are major challenges for which many postdocs feel underprepared. We established the PI programme with this in mind. It provides mentorship, training and networking opportunities to a group of postdocs about to apply for academic positions. It was a pleasure to meet the first cohort of fellows in October and hear their stories. You can read more about them on our website, and also find out more about the aims of the programme and the application and selection process. It was also satisfying to hear how the programme had helped them. One commented, ‘The programme has been a game changer for me as I search for an independent position, giving me unprecedented visibility in the community as well as amazing training that will help me in my transition to being a PI.’ We have now opened our second call for applications to this programme. We are keen to see applications from all corners of the world and from all areas of developmental and stem cell biology. If you know anyone that might benefit from the programme, please let them know about it.
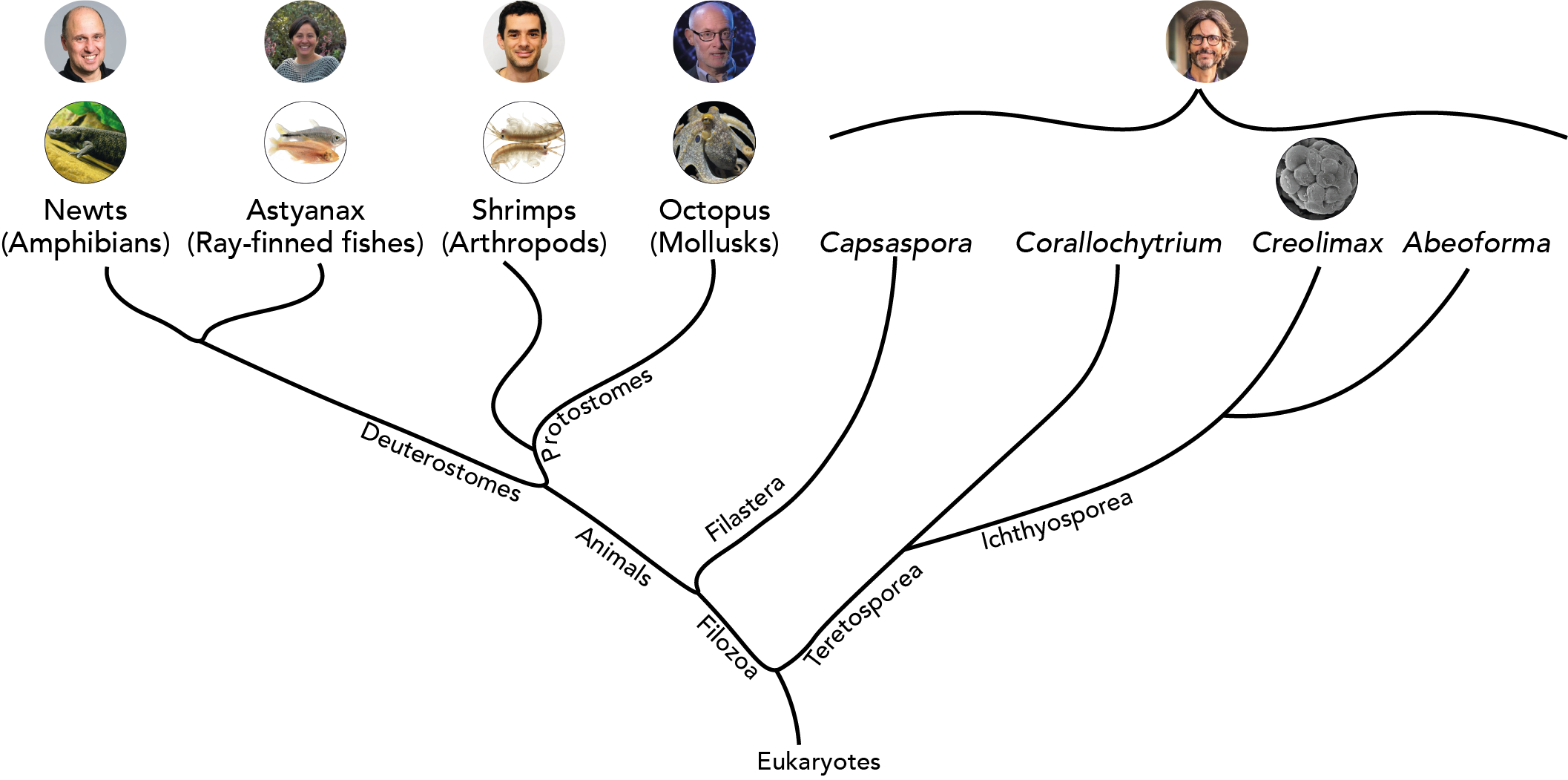
When the two of us, Brent and Alex, started our Node correspondent positions early last year, we both expressed an interest in non-model organisms (NMOs). While one of us (Brent) works with several NMOs, the other (Alex) is solidly in the mouse camp, arguably the ur-model organism of our days. Since we both declared a shared interest in NMOs, it was a matter of competing or collaborating (har har)! Combining our forces, we identified and individually interviewed five researchers who work with NMOs in their labs. But even in this group, some NMOs are more conventional than others.
Here, we have condensed each individual interview into one “super-interview” that compares and contrasts the questions, the approaches, and the problems these labs encounter in their research. The result is a regular pot pourri of NMO research. As they all understandably had many interesting things to say about their respective research areas, we’ll be uploading individual interviews as articles for The Node throughout the next couple of months.
It’s critical to establish systems facilitating investigations in novel organisms given the urgency to document and understand the rich biodiversity of our natural world and preserve it as best we can. But on top of that, we found these scientists are motivated by a universal human urge to explore something unusual, confusing, or sometimes just plain cool. They demonstrate the true spirit of research.
Although by no means a comprehensive or even representative list of NMOs, we hope this article will encourage people to become familiar with more obscure research models and questions, and perhaps provide the impetus for the field to explore beyond its established comfort zone.
With that, let’s introduce our researchers and their respective NMO(s) of choice:
.
.
.
Iñaki Ruiz-Trillo: works with single-celled eukaryotes (Institut de Biologia Evolutiva, Barcelona, Spain)
.
.
.
.
.
Cliff Ragsdale: works with octopus (University of Chicago, USA)
.
.
.
.
.
András Simon: works with newts (Karolinska Institutet, Stockholm, Sweden)
.
.
.
.
.
Michalis Averof: works with shrimps (Institut de Génomique Fonctionelle de Lyon, France)
.
.
.
.
.
Patricia Ornelas-García: works with Mexican cavefish (Universidad Nacional Autónoma de Mexico)
.
.
Could you give us a short summary of your research?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
What fascinates me is the evolutionary transitions. I did my PhD in one of the major evolutionary transitions—the original bilaterian, that transition from radial symmetry to bilaterally symmetrical animals. I was looking to do a postdoc on the origin of eukaryotes. My supervisor proposed to me, saying, ‘I have a very weird organism that seems to be closely related to animals. Would you be interested in the origin of animals?’ I said yes, I’d be interested, and then I started to work.
Cliff Ragsdale (octopus)
I did a post-doc on amphibian limb regeneration. In my own lab I moved into neuroembryology—essentially chick molecular experimental biology focusing primarily on midbrain development. And then about 2 decades ago, I switched to evolutionary neuroscience and was interested in two problems: one, the evolution of the neocortex and the other is on cephalopods, specifically octopus. I was interested in how a large brain could be organized apart from the vertebrate design.
Michalis Averof (shrimp)
In the lab we study regeneration using this small crustacean [Parhyale hawaiensis] as our model. Ever since my PhD, I was interested in comparative developmental biology, in what different organisms can tell us about mechanisms of development and how those mechanisms evolve. This is how we started to work with crustaceans. Then gradually regeneration, which was a side project, became our main focus.
Patricia Ornelas-García (Mexican cavefish)
I work mainly with the systematics and speciation mechanisms in freshwater fish species. Since my PhD, I have worked with Astyanax.In the beginning, I just wanted to [study] as many [Astyanax populations from various] caves as we could, so we could test these hypotheses of how many times the fish has been able to adapt to the caves. Nowadays, we are starting to move to some developmental analyses. So far, we have been able to reproduce five [Astyanax] populations, different from the common ones like Pachón or Tinaja. We are trying to compare the differences during early development. We are also exploring the phenotypic convergence with other cavefish in Mexico, particularly in some processes related with asymmetry.
András Simon (newt)
I started my lab 2002, and since then we have been working on newt regeneration. The reason we are interested in newts is because of their regenerative capacity. They can regenerate large body structures such as entire limbs, large parts of the brain, spinal cord, cardiac muscle. We think that [by studying newts] one can understand mechanisms that allow or disallow regeneration of complex body parts in vertebrates, but also use regeneration as sort of a way to address fundamental cell and molecular or biological questions that these animals are specialists in. In that context, dedifferentiation of cells is one of the main interests in my group.
.
What are some of the particular challenges of using this NMO/NMOs in general?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
There was a lot of missing information. Imagine you want to understand the origin of World War II. You need to have information of how it was before World War II, you know? It’s more or less the same—you need a comparison framework. And there was no information.
Cliff Ragsdale (octopus)
There are still many, many technical challenges. Octopus bimaculoides–the southern California two spot octopus–is kind of like a lab rat in that it has too long of a generation time to be appropriate for any kind of forward genetic studies. We really don’t have a cephalopod yet that’s appropriate for that. And you might say, “Oh, well, why can’t you just use CRISPR?” But the key thing is being able to inject embryos and to have the embryos survive. Transgenesis just hasn’t fallen yet.
There are no invertebrate marine cell cultures. Let that settle in for a moment. You can do primary cultures, and like a science fiction film, these cells in culture or explants in culture will seem to live forever. But it’s very hard. To get proper cell culture of any marine invertebrate would open up a lot of cell biology if the techniques were general ones. There’s hardly anyone working on that, but that’s certainly a need.
Michalis Averof (shrimp)
[The Parhyale research community is] more isolated in terms of technology, in the sense that for every project we have to develop our own tools, there isn’t this big community behind you generating Gal4 drivers or Cre lines that are shared, like you have in other systems. When you start a project, you have to generate those tools by yourself. And that is a major limitation when working with our kind of peripheral models.
The other issue is, we haven’t yet figured out an easy way of sending these animals across the world without having problems with customs.
The benefit, on the other side, is that in almost anything you study you’re going to make new discoveries, because no one has studied that before. So you’re entering a virgin field. You have the opportunity to shape your research field to a larger extent.
Patricia Ornelas-García (Mexican cavefish)
Yeah, I think one of the challenges is that there are a very restricted number of people working with these [species]. And in a way, it’s fascinating, because you will find something new for sure. But [from another point of view], in research groups [studying established organisms] it is easier.
We were trying to characterise the microbiome of the fish. And it was a little challenging, because there was not a lot of information already published on protocols or how to treat the data. Or when we are trying to set up [experiments], for example, for physiology or for another kind of ecological analysis, it’s sometimes difficult.
András Simon (newt)
[Newts are] technically challenging for several reasons. One is the relatively long generation time, which can range from nine months to five years. When the molecular biology and the genetic era entered, these organisms became a bit obsolete. Most researchers said, “Well, if I can do all these things in mice, I won’t do that in salamanders, because it’s just too time consuming.” And funding agencies are not always very patient. They also have quite large genomes. Genome sequencing technologies were not there some years ago to make it possible to get a reasonable assembly of the genome.
.
What are some of the unique opportunities from using NMOs? Or what recent technical developments are aiding research progress?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
Most of the people who were working on the origin of animals were working with early branching animals, like cnidarians, ctenophores, and sponges. This is very important for early animal evolution, but you can never understand the transition because you are not considering the stuff before.
Cliff Ragsdale (octopus)
You know, it’s great to have a genome.
Michalis Averof (shrimp)
[Parhyale] is a very good system if you want to image what happens during regeneration. You can image the whole process at cellular resolution from beginning to end, [and] the reason is that these animals as adults are transparent. They’re small enough that we can just image through their legs with a confocal microscope. We can make transgenics so we can label the cells; and we can immobilise them, which can be a big challenge in other organisms [that regenerate as adults]. In our system, what allows us to image regeneration is the fact that arthropods are encased in the chitinous exoskeleton. We can use simple surgical glue to stick those animals onto a cover slip. And they will stay there, they can’t go away until they molt.
Patricia Ornelas-García (Mexican cavefish)
During my Bachelor’s, I was working with mice, the typical model in the lab, and somehow I think that the number of questions sometimes can be very restricted because there’s already so many studies in these animals, that it’s difficult to come up with something new.
And [working with Astyanax] I have a lot of things in my favour, I am from Mexico, I can work in the field, I can do a lot of in situ experiments. Even nowadays, there are very few Mexicans working with Astyanax. It just happened that there were a lot of things that made me realise that there was a lot of potential in the [Astyanax] system for me.
András Simon (newt)
For us, there were three things that enabled us to interrogate the problem at the molecular level. First, we introduced to the lab a new species, Pleurodeles waltl, which is relatively easy to breed. Secondly, sequencing technologies improved to the degree that now we can compile a good genome assembly. The third is genome editing technology. CRISPR revolutionised many fields, and ours as well. During the past 7-8 years we have been generating genetically modified animals for cell tracking experiments, for loss-of-function experiments, of different genes. Now we are really in the position to address the question of regeneration with molecular tools, and get some mechanistic understanding at the molecular level.
.
What is a particular study or finding in your NMO of choice that you think is really cool?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
The most important thing we have discovered is that the unicellular ancestor of animals was genetically much more complex than previously thought.
Cliff Ragsdale (octopus)
It’s in the genomes where I think the greatest surprises have appeared. Having large genomes kind of raises the possibility that, like the vertebrate lineage, you have whole genome duplications. That’s something we see in other animals as well. And that doesn’t show up at all in cephalopods. Lots of key gene families are duplicated. We’re very tempted to think, that even though bony fish, some clades have way, way too many genome duplications, we’d like to think that somehow the two rounds of genome duplication leading to jawed vertebrates is important for the innovations we see in structures of vertebrates, including us—there’s no indication of that in cephalopods. That raises the interesting possibility that gene regulatory mechanisms, cis regulatory mechanisms in particular, by disruption of this synteny by this blender effect might underlie many of the innovations seen in soft-bodied cephalopods.
Michalis Averof (shrimp)
One of the latest papers to come out is [by] people who study biological rhythms, and have studied how Parhyale regulate their daily activities in relation to tides. Our animal is an intertidal species, and it seems it has an endogenous clock that runs with the tidal cycle rather than with a day-night cycle. Well, they have both, but somehow the two interact in a complex way. People who study circadian rhythm had noticed that there were two peaks of activity, one in the morning and one in the evening. And the intertidal cycle is a little bit longer than 12 hours. So that might reflect the fact that they have a 12-hour cycle rather than a 24-hour cycle. But of course, in nature, the tidal cycle comes slowly out of sync with the day-night cycle. And that is not observed in the lab. [So] people are beginning to study phenomena that were not accessible before in the standard models, like tides and regeneration. There are new aspects of biology that become accessible once you have a new system.
Patricia Ornelas-García (Mexican cavefish)
In our most recent paper, by a Master’s student of mine, we assess [this] question [of how many times Astyanax has been able to adapt to caves independently]. In our results, we have at least three independent colonisation events of the caves, which for some is crazy, it’s not possible. But from our point of view, we are really relying on exhaustive sampling of the caves, and that is what we’re suggesting.
András Simon (newt)
There’s an interesting dichotomy: one would expect that the price newts pay for getting cells proliferating and dedifferentiating is that they are more prone to get cancer, but that’s actually not true. In fact, during regeneration some genes, which are heavily mutated in mammalian cancers, are actually downregulated during regeneration. And instead of everything just growing randomly, they create a structure.
.
How does data analysis and sharing between labs employing NMOs differ compared to studies using established MOs?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
We have quite a lot of collaborations. Our lab is quite diverse. We collaborate a lot. So far most of the community is quite open. It’s usually not a problem.
Michalis Averof (shrimp)
We all use the same population of Parhyale. It’s a population that has been kept in the lab for more than 20 years. There are a few people beginning to isolate new populations from the wild. The funny thing about the population we share is that it was picked up in an aquarium in Chicago, about 25 years ago, and we don’t know which part of the world it came from originally. We all use it, the genome has been sequenced from and all our transcriptomic work is based on that population. For transgenesis and CRISPR, we more or less use the same protocols. We don’t share transgenic lines so often, but that’s mostly because we have different interests, and each of us develops our own lines for the particular questions we’re asking.
Patricia Ornelas-García (Mexican cavefish)
In my opinion, one of the things that the Astyanax model has is that [researchers working on it] are very open. For example, when we were trying to reproduce a fish, we were [asking for] a lot of information [from other groups].
András Simon (newt)
Personally, I’m very open with our data. If someone asks me about even unpublished data, I’m happy to share. In my group, we sometimes host guest researchers from the field who are welcome to participate in our lab meetings, where we discuss raw data. Of course, sometimes people are more cautious. But I would say in general, this is a super friendly field.
.
How large/connected is the community studying your particular NMO of interest?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
There’s quite a lot of research and quite a lot of labs in the region of multicellularity. My feeling is that every year there is more and more people interested. So now, we are kind of explosive. I see more and more labs connecting with us and being interested in our organisms or in our questions.
It’s one of the biggest transitions compared to all the other major evolutionary transitions. The origin of life, the origin of the eukaryotic cell, the origin of humans, the origin of bilaterian animals–there’s only one single origin of all of that. Animals acquired multicellularity from some kind of ancestor, which we are trying to get who they are, or how they were. Plants acquired multicellularity from a different ancestor from different parts of the tree of life. And algae acquired multicellularity from different ancestors with different genomes, different raw materials. And so on and so on. And the people working on those multicellular organisms—we don’t talk to each other. We go to different conferences, different journals. I think we have to talk to each other at some point. It was impossible to do that in the past because there was a lot of research in the origin of animals, but not so much in the origin of plants. But now there is more and more of these models. I think it’s a very good moment.
Cliff Ragsdale (octopus)
There’s a huge explosion with people who are interested in the brain or who are interested in development and other aspects of cephalopod biology. I kind of think people were always fascinated by cephalopods, captivated by the behaviors of octopus and the adaptive coloration system particularly pronounced in octopuses and cuttlefish.
Michalis Averof (shrimp)
It’s a very small community, there are maybe 20 or 30 people working on the animal. You would imagine that small communities are very well connected. We are connected, but not very tightly. I think it mostly has to do with the fact that we are on different continents and we study different questions. We talk to each other and we share tools and genetic resources. For example the genome sequencing and assembly was a collective effort.
Patricia Ornelas-García (Mexican cavefish)
An important thing to highlight in Astyanax is that we have the biennial meeting. Because it’s not a model organism, [everyone is] really [willing] to talk about the system in a very open way, and include new researchers. Particularly for me, when I was finishing my PhD, this was a very dramatic point, because I saw a potential in the system that I can be included.
Nowadays, a lot of people are trying to [investigate] this model [from] the eco-evo-devo perspective. They have realised [that it is important to distinguish between the different Astyanax lineages], because some of the results that they get are related with the lineage, and not particularly with the environment. In these terms, there is a growing number of labs wanting to work in the field, know more about the ecology in situ, learn more about the behaviour, the physiological adaptations.
András Simon (newt)
We are growing to the degree that we have a yearly salamander meeting. Also, there is the larger field of regenerative biology, which is at a decent size now. We just started a new organisation, the International Society for Regenerative Biology, and the inaugural meeting was in Vienna this year. That gives room to different regeneration model overlays, because regeneration at this scale is quite widespread evolutionarily but also randomly distributed. It doesn’t decline with the increase of [organismal] complexity. At these meetings we gather researchers who work with regeneration model organisms, in particular salamanders, including newts, planaria, zebrafish, but also Hydra, cnidarians.
.
Would you say it takes a particular sort of character to want to study NMOs?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
The most important thing is the question. Then the techniques and the approach vary depending on the moment.
Sometimes it has been hard to be alone. And at the same time, there is something beautiful about that, because you seem to be a pioneer. It takes something of the more human angle of being an explorer, you know? So I cannot complain. I mean, I complain because it was hard at some points and it would have been faster [in another organism]. But also it was nice to, for many years, say, ‘Oh, you should look at these organisms.’ And maybe some people didn’t believe you and now they say, ‘Oh yeah, that’s cool.’
Michalis Averof (shrimp)
Definitely, it takes a different kind of researcher. To work with these animals, you have to realise that research is going to move forward much more slowly, because you will have to start many things from scratch.
András Simon (newt)
If one is not super naïve, it must be that they are more dedicated to answering a question than just progressing in their career. But I like the field a lot. Because of the lack of tools, I found the field intellectually very mature. People had time to think, that is my impression, and I liked it. Now, of course, there are [more] tools. So the risk is that we’re going to do too many experiments. No, I’m just joking.
.
Do you have any additional or broader comments that you might like to share?
Iñaki Ruiz-Trillo (single-celled eukaryotes)
Work with people that are nice people, that are persons before scientists. Whether it’s a PhD, working for a supervisor or collaborators, whatever. You have to have people that you feel confident you can say a stupid thing. And you can laugh together and you can be interested together about the questions. Then everything will be much, much, much better.
Michalis Averof (shrimp)
Over the years, we have developed this idea that model organisms will reveal universal mechanisms, and that we can study most of biology through the model systems that we have chosen. But I am convinced that there is an enormous amount of biology that we are missing, if we rely only on the established models. There are biological phenomena which are not represented in this handful of organisms.
I see these like new continents of biology that are still unexplored. New model organisms will allow us to explore these. Of course, it’s going to be difficult, it’s going to take time, and it’s going to take development of tools. But that’s for me the major motivation for going into different systems, because I think there’s biology that we haven’t discovered yet.
Patricia Ornelas-García (Mexican cavefish)
The problem with non-model organisms is the conservation situation. Nobody will catch Mus musculus from the field, they have so many reproductive lines in captivity that they don’t have to. Non-model organisms are in the opposite situation. Most labs [working on them] want to have more wild lines, more related with what is really happening in the field. And if you have 200 labs working [on cavefish], imagine the impact that we can have on the natural population. We published, just at the beginning of the year, a paper [on] size estimation of the fish population in the caves, and it’s maybe around [a few] thousands of fish. And imagine, in the last 10 years, there have been around 200 fish extracted from the caves. So if you imagine a system that has to recover from 20% of the population being lost only because of scientific sampling, it’s problematic.
When you try to make researchers aware of the situation, [they] really believe that the main extinction drivers of this kind of population are not related with our sampling. Most of us really believe that it’s all global warming, or local people extracting water for drink. I’m very surprised, because normally you have to fight this kind of attitude outside the scientific [world].
Thus, it’s important to be part of the solution, not part of the problem. Maybe we should consider what we can do to solve the conservation situation. We need to be more aware about the impact of our research and do our best to guarantee the prevalence of this model for future generations. [Because] what makes these organisms amazing also makes them vulnerable, in a way.
András Simon (newt)
I was thinking about what makes an organism model or non-model? Is it a qualitative term, or quantitative? If enough numbers of researchers work on it, is it a model organism? If not, is it not a model organism? What is the definition? I would probably say that [the newt] is a model organism for regeneration, but it’s a non-model organism in the sense that not so many people work on it, although the community is growing.
.
A phylogenetic tree of the organisms our interviewees work on.
We couldn’t have succeeded in our effort to make this compilation article without the generous time these researchers were willing to share with us, so our sincere thanks go out to András, Michalis, Paty, Cliff, and Iñaki. We are really hoping that this is just the beginning of our collective exploration of NMOs, and of greater interspecies research collaboration. The more we investigate, the more we can hope to understand how the amazing tree of life is built, interconnected, and how to best attempt to preserve it in all its messy glory.
“I feel like I have the best of both worlds. I’m still very much focused on genes and genetics, which is what I spent a lot of my academic career doing, but with that added benefit of, okay, well, I can see the reason why we’re doing this work.“
Dr Louisa Zolkiewski
This latest episode of the Genetics Unzipped podcast is all about the next generation of geneticists, as we meet three up-and-coming scientists who have been supported by the Genetics Society, and learn all about Bermuda skinks, the Black Death and life after a genetics PhD.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
The Genetics Society would love to hear your thoughts to help them make those choices.
So please go to geneticsunzipped.com/survey and fill out our short questionnaire, telling us what you like and don’t like about the podcast and your thoughts on future changes.
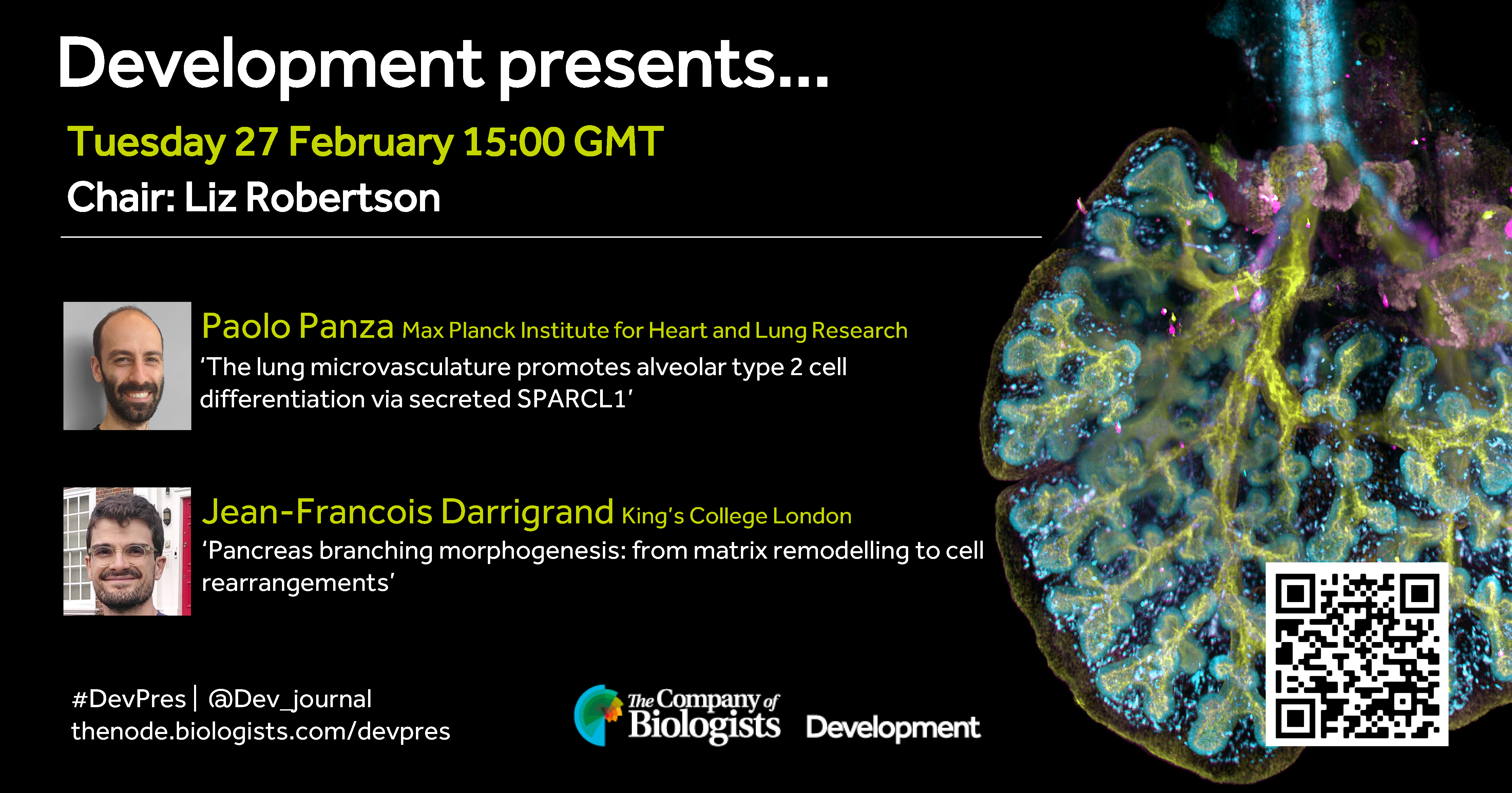
In the second 2024 webinar, Development Editor Liz Robertson (University of Oxford) hosts two early career researchers studying organogenesis.
Tuesday 27 February – 15:00 GMT
Paolo Panza (Max Planck Institute for Heart and Lung Research) ‘The lung microvasculature promotes alveolar type 2 cell differentiation via secreted SPARCL1’
Jean-Francois Darrigrand (King’s College London) ‘Pancreas branching morphogenesis: from matrix remodelling to cell rearrangements’
At the discretion of the speakers, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
In line with preLights’ ongoing postLights initiative, one of the preLights Ambassadors, Martin Estermann, has decided to create and host a new webinar series focusing on the journey of the preprint. This involves a series of virtual webinars featuring different researchers (often early-career researchers) who posted their manuscripts as a preprint before or during the peer review process.
The goal of this webinar series is to highlight the benefits of preprinting to the research community and to discuss relevant examples both within and outside the preLights community.
Three webinars were chaired by Martin in December 2023, which featured three preprints that have all been covered by preLights.
Back in early 2013, Olivier Pourquie (Development’s then Editor-in-Chief) and I were working hard to integrate stem cell biology into Development. Reading papers and attending conferences, much of the focus at the time was on how principles of development could be harnessed to improve stem cell differentiation protocols, often with the goal of using those derived cells for more medically-relevant purposes. But we were also beginning to see a new trend emerging: the desire to use in vitro stem cell approaches to really understand human development. Organoids had only recently been developed, and the range of tools available was limited (single cell sequencing was in its infancy), but we could clearly see the potential of this burgeoning field.
From this, the idea for Development’s first Journal Meeting “From Stem Cells to Human Development” was born, and in September 2014, around 120 scientists gathered together at Wotton House in Surrey. The event enabled researchers working on a wide range of organ systems and questions, but all facing a common set of difficulties associated with working with human cells and tissues, to come together and discuss both the challenges and their potential solutions. The event also provided an opportunity to discuss the ethical considerations around working with human stem cells and primary tissues; looking back at my notes from that first meeting, and reading the articles we published as a result of it (Hyun, 2015 and Hermeren, 2015), it’s striking to me that the ethical debate at the time focussed largely on human-animal chimeras – the idea and potential consequences of generating stem cell-derived integrated embryo models were barely discussed. A decade on, and such models are very much at the forefront of the community’s mind.
At the time, there were no other conferences focussed exclusively on human development, and the enthusiasm for bringing this growing community together was clear. To answer this desire, Development has turned this event into a biennial meeting, and we’re delighted to announce that registration for the sixth ‘From Stem Cells to Human Development’ meeting is now open. Having attended every iteration since the meeting series’ inception, it’s been a real pleasure to see how both the field and the community have grown. Work with human stem cells and primary tissues holds huge promise both to understand our own developmental origins and to develop therapeutics, and the rate of progress has been impressive. I’m excited to see what we will learn at this September’s meeting, and I invite those of you with an interest in this dynamic and important field to apply to join the conference!
All flavours of a conference – in-person, online, or maybe hybrid?
Thinking back to my first conference, it was a confusing experience for me. It happened well into my PhD around the end of the second year. Before that, I would prepare the posters, but it was my supervisors who would go to the conference and present them. The only directions I got for my first conference were to stay at my poster and answer questions. It took me a couple of conferences, talking with friends, and internet surfing to crack the enigma of these scientific gatherings. I realised that conferences are much less about diligently listening to everything that is presented and much more about wondering, sharing, and learning.
Once I understood the rules, conferences felt like an ocean of information, with unexpected treasure findings waiting in the middle of a dive into a topic you have chosen on a whim. I can’t even count how many times I would go into a room, listen to a series of talks, and suddenly hear the solution to the problem I had been having for some time or a confirmation of my idea that sounded crazy.
I was in the process of figuring out how to network at conferences when COVID-19 turned everything online. And I was all in for this new experience. Now, after having a fair share of both online and in-person conferences I am ready to summarise what is the difference and what works or not so much.
The structure – what is easy to transfer to online and what is challenging
Online conferences mostly follow the same structure as the in-person conferences:
opening remarks
plenary lectures
multiple parallel oral talk sessions
poster session (not all online conferences)
industry booths/workshops (not all conferences)
closing remarks
Lectures and talks are easy to transfer to online settings; the talks are usually prerecorded or live, depending on the technical abilities of a conference. The part which is difficult to transfer to online settings is poster presentations. In-person conferences usually have many printed posters (sometimes e-posters displayed on a monitor) in one place and let people wander around with a book of abstracts. In online conferences, I saw two approaches.
Try to replicate the experience of the onsite conference by creating a virtual space (like gathertown) where attendees can move their virtual selves to wander around.
Give each poster presenter a very short talk or flash presentation (generally within 5 min) with a couple of minutes for questions.
In my experience, the first approach works well for small conferences, where you can easily read abstracts for all the posters in a short period of time. The positive point of this setup is that you can stay with the poster you like for as long as you want, giving you the freedom to have interesting conversations on the topic you like. When the number of posters is huge, it becomes difficult technically, as not everyone has a super-fast internet connection, and it is just difficult to navigate on a computer screen.
For some people, the second approach may be more logical. It allows attendees to hear the content of all posters in a structured manner, so you don’t miss anything interesting. However, having only several minutes for questions is rarely enough, and if you want to connect with a speaker, you will need to try catching them online afterward. So, in the case of the second approach, I wonder if there is a need to make a poster or just turn it into a short presentation with slides (which, to be fair, some conferences do).
The networking – difficult both in-person and online
For me, the biggest structural difference between in-person and online conferences comes from the way networking is organised. In-person conferences usually provide tea and coffee breaks when you can catch up with the person you were listening to right outside of the door or bump into some acquaintances, not to mention that there are some social events like dinners or workshops, which are specifically created for networking. Not that it makes networking easy; it just gives you much higher chances to strike up a conversation. There will always be an extroverted person who will go around and stir up some conversations. Hopefully, it will be your friend, and you will tag along (it was my strategy for networking in the early days of being a researcher 😉).
Thanks to the pandemic, I had a fair share of online and hybrid events (conferences, workshops, panel discussions), some of which I organised myself. My conclusion is that networking online is always challenging. In the case of a hybrid setting, when the conference is happening both in-person and online, the in-person and online participants are often not quite aware of each other’s presence, which creates two different audiences and the need to work with them separately. A lot of organisers naturally focus on an in-person audience, with no facilitators for online participants, resulting in a lack of networking and a feeling of being left out. Even in 100% online conferences, where you would usually have chats, forums, or virtual spaces, the absence of facilitators or specifically designed networking activities usually results in difficulties with networking.
The main reason, I guess, is that people are used to listening to something online, but not to communicating. Even in person, it may be difficult to master the courage to go to a person, introduce yourself, and start a conversation. It is several times harder when you don’t see their face and have no visual cues to rely on to see their expression and reactions so that you can understand whether they are up for a conversation, do they like where this is going, are they bored, do you need to finish, or can you continue. As a result – you give up even before trying.
One of my greatest experiences with online communication was Zeroverse’s online workshop on carbon literacy. The structure of the workshop included lectures, which were divided into several parts, and breakout sessions after each part. Breakout rooms had only 4-5 people, and all cameras were on. The participants also had a specific task for the breakout session and a general script of how to start a conversation announced by the host (introduce yourself, say your opinion on the matter, summarise what everyone said, and choose a representative to present). The tasks and groups were designed in a way that each person would need to talk. I found that it was easier to talk and overcome the first shyness when there was a system of communication.
Why not just participate in-person?
Because it seems that it is difficult to organise hybrid events, and there are many troubles for online participants, I want to talk about the reasons why you may choose online over in-person participation. And why I think that, as a community, we need more hybrid conferences rather than returning to in-person only.
You can always leave your cat on duty when you need a break ;)
Financial reasons. There are some obvious reasons. Skyrocketing prices for everything: participation fee, airplane/train tickets, hotel stay. Now I often hear that, for example, in Europe, people try to encourage participating in local conferences or traveling by train. It is a good initiative if you live in Europe or North America. But I live in Japan, which is only at the beginning of its journey to create an international scientific space. Not many international conferences choose to be in Japan because it is not an easy destination. Many local conferences are only in the Japanese language, which means that you can participate only if you have quite a high Japanese language ability. In many cases, even if there is no requirement to present your work in Japanese if you present in English, people will generally try to avoid you, as they don’t feel comfortable speaking English. So, if you want to be part of the bigger international scientific community, you would be forced to participate in conferences that are not in your country and would need to fly there. And the prices of airplane tickets from Japan to Europe at the moment are astonishing, especially with the weakening yen.
Prices are one of the reasons the conference world seems extremely unequal. So, it is always great to see that some conferences choose destinations like Africa, Asia, or Latin America as places for their international conferences. But another obvious answer to the problem – is to host hybrid conferences with lower participation fees for online participation.
Financial reasons were also one of the main reasons I decided to join my latest conference – EMBL Symposium “Organoids: modelling organ development and disease in 3D culture” online, rather than in person in Germany. But were there any other reasons? For me – yes.
Communication reasons. Online participation allows you to be in a comfortable place. Usually, I would participate from home, where I can sit comfortably (which sometimes looks like half-lying on the sofa or snacking at the dinner table or sitting down on the floor and petting my cat), be relaxed, which creates an atmosphere where I personally can concentrate easier and feel more comfortable asking questions. In my experience, I ask many more questions in online settings than in onsite ones, and I pinpointed the reason – the ability to type it rather than the necessity to hold your hand up and become the focus of the whole room’s attention is what makes it easier for me. Now, I do understand that this is not true for everyone. I do know that for some people, it is difficult to concentrate at home. I just want to point out that people like me exist, and we also want to attend conferences, feel included, and communicate with others. I would actually go even further and propose that even on-site conferences have a chat option where you can type the questions during the talk and opt for the session chair to read your question rather than ask it yourself.
One of the nice touches of the Organoid Symposium was that panel administrators treated both online and onsite questions equally and tried to ask them in the order of appearance. They also transferred all the unanswered questions in the chat due to time constrains to the designated forum and actively encouraged speakers to go check and answer the remaining questions on forum.
Effort reasons. Conferences are an intense experience. I am quite perplexed about how we came to the programs that include multiple parallel sessions and the schedule from 8 am to 8 pm. I probably need to do some research in that direction, too 😉. Maybe it reduces the price of the event, but at what cost for our brains? Even if you consider conferences fun, a 12-hour time frame is exhausting. But in reality, conferences counted as work. It requires a lot of listening, thinking, understanding, and concentration. I guess not all people are participating all day long, but in my experience, there are so many interesting things going on that when you are done marking all the sessions you are interested in, you have a very packed schedule. And at the end of the day, you feel overwhelmed with information. Online conferences allow you to listen to at least part of the content on demand! Which allows you to plan in a much healthier way. You check all the sessions you want to listen to and then check which of them are available on demand. Attend only the ones that are not available, and then listen to others in your free time. This is exactly what I did last month when I participated in an Organoid conference online. I’ve listened to what was available only in live streaming and then spent three weeks listening to everything else that was available on demand. Why so long? Because I was listening to one to two presentations a day and not every day, only when I knew I was ready for the new information.
Space reasons. Continuing on the comfort and overwhelming experiences — there are no overwhelming crowds of people when you join online. I know that many people love to be in a bustling place with everyone talking and mingling, but I am one of those people who get tired very quickly in these environments. The flashing slides in a dark room, people talking everywhere, especially during poster sessions, and the constant flow of information with often no planned activities for relaxation or proper rest (let’s be honest, 10 min break between 1 h sessions which you are supposed to spend on connecting with people or checking and going to the next session is hardly a rest). It is often difficult to find a quiet spot to recharge and have peace when attending in-person conferences. On the other hand, online conferences easily solve this problem. You have a 5- or 10-minute break – mute your laptop and have your quiet time. You can have a short breathing session, go talk with your partner, spend several moments with your pet, or just stare into a wall if that is what you need. Which, in my case, leads to enjoying the whole process much more. I can take a break when I feel that I need it. Going back to my last online conference on Organoids, I think that what they did great was give more time for breaks. The coffee break was 30 minutes, and the lunch was 1 hour and 30 minutes. The online participants would also see a banner that proposed yoga or some exercise to do during the breaks. I think that this kind of planning shows respect for our body’s needs. It takes into consideration that the intense brain work is still demanding on our bodies, and we need our rest.
Some of my favourite talks at the Organoid conference. I also wanted to talk a bit more about the Organoid conference, as it is because of this conference I decided to write about online participation.
The Organoid conference was also the first small conference I attended. Usually, as I said, there are a lot of multiple parallel sessions, but for the Organoid conference, there was only one. What I’ve realized is that this way, organizers can control the program much better, which means that the quality of the talks was fantastic! There were so many interesting results, ideas, and inspiration! Of course, conferences that allow more participants, even though the results may not be as groundbreaking, are also important; they just have different goals. But now I fully understand why several senior professors would recommend attending specific small conferences to keep in touch with the latest trends.
It was so good to hear Madeline Lancaster addressing the problem of reproducibility in biological research. The usual approach to the problem is – just to check several lots of cells and choose the ones that work. Which is a kind of “treat symptoms” versus “treat the cause” kind of solution. It does not answer the question, “Why do some cells work, and others don’t?” Can we really call it reproducible if you need to try several makers and lots of cells until you find the one that allows you to reproduce the results? It was refreshing to see a person who decided to answer the question: “Why are some iPS cells allowed to form organoids and others not? Is there a way to turn a bad iPS cell line into a good one? What is the mechanism behind all this?”. I do hope that more researchers will go into such details. As a researcher from the tissue engineering field, I feel that too many articles are concentrated on their techniques and know-how rather than on the fundamental process underlying all of it. This sometimes creates bizarre situations when some techniques work only in some labs, with certain reagents, and in the hands of certain people and practically cannot be replicated.
Another one of my favourite takeaways from the conference – is the video of the T-cell killing cancer from Anne C. Rios group. And I love it for completely unscientific reasons. It often feels futile to try and change something. There are so many inequalities around us. And sometimes it feels like nothing can be changed, nothing can be done. But looking at how one brave and energetic “Super engager” T-cell destroys the whole clump of cancer cells and, in the process, influences and triggers another not-so-energetic T-cell to go and do the same… Sometimes, it’s baffling how such human stories can happen even at the level of cells😊 You can change things; it is tough to do it on your own, but what you are doing can inspire others, and with time, you may even get help from unexpected sources, who will notice what you are doing and will be like – this is cool, this is what we need, let’s try to find a way to help.
This is a movie produced in 1964, which describes the procedure to achieve nucleo-transfer in the frog egg (nowadays referred to as ‘cloning’). The demonstration is performed by Dr Tom Elsdale, a distinguished embryologist who mostly worked at the MRC Human genetic unit in Edinburgh. Elsdale had spent some time in the USA to learn this experiment after the founding paper of the field was published by Briggs and King in 1952. This technology was further used by John Gurdon to carry out the first and Nobel prizewinning cloning of animals by using nuclei from adult differentiated cells. This movie echoed a publication describing the approach and published in 1960 by Elsdale, Gurdon and Fischberg in JEEM, the former name for ‘Development’. It was likely used for teaching purpose, as well as to facilitate the learning of this difficult manipulation by fellow scientists.
Where was this found?
This movie was in the archives of the Geneva Zoological Institute (Station de Zoologie Expérimentale) in Switzerland. In the 70s, this department was chaired by Michael (Mishka) Fischberg, who had been John Gurdon’s PhD supervisor in Cambridge, UK, and senior author of the paper mentioned above. Fischberg was a great scientist and a good man who had collected and developed the largest collection of Xenopus species in Europe to study developmental and evolutionary questions. In 2006, as a chair of this department, I had the difficult task to close this Institute (where I had been a student) and move it to more modern buildings downtown. Fortunately, the movie had been kept by Charles Thiebault, a close collaborator of Mishka Fischberg.
How was this taken?
This is a 16mm film, which was recently transferred onto a CD format. The 16mm camera must have been positioned on top of binocular objectives, yet it is not shown as to how this was done in the recorded demonstration and hence the exact recording set up is unknown (any comment on this?).
Why should people watch this movie?
This movie illustrates how classical embryology was carried out 60 years ago and gives an idea of how pioneers in the field could overcome the many difficulties caused by experiments as fundamental as nucleo-transfer, for example. You will appreciate the many small tricks used here and there, the extraordinary timer, the pre-AI animations and summary, as well as Elsdale’s experimental precision, despite being one-eyed due to an accident when he was a kid. A special mention is given to his pullover and to the brilliant soundtrack starting with croaking frogs.
References
Briggs R, King TJ. Transplantation of Living Nuclei From Blastula Cells into Enucleated Frogs’ Eggs. Proc Natl Acad Sci U S A. 1952 May;38(5):455-63. doi: 10.1073/pnas.38.5.455. PMID: 16589125; PMCID: PMC1063586.
Elsdale TR, Gurdon JB, Fischberg M. A description of the technique for nuclear transplantation in Xenopus laevis. J Embryol Exp Morphol. 1960 Dec;8:437-44. PMID: 13726553.
In the latest episode of the Genetics Unzipped podcast, we’re exploring the impact that Gregor Mendel and his populariser William Bateson have had on the past century of genetics, and asking whether there could have been a more Weldonian view of the world.
“It wasn’t a foregone conclusion that we would end up with our knowledge of inheritance organised around the idea that in the first instance genes come in these two versions, dominance and recessive, and they’re associated with these unit characters, you know, yellowness, greenness, roundness, wrinkledness, cystic fibrosis, black… That categorical thinking I think is problematic, it’s problematic scientifically and it’s problematic socially.”
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
 (No Ratings Yet)
(No Ratings Yet)












































 (3 votes)
(3 votes)