Our next Development presents… webinar is on the topic of germ cell development and will be chaired by Development Editor, Swathi Arur (MD Anderson Cancer Center).
Tuesday 5 December 2023 – 15:00 GMT
Gabriele Zaffagnini (Centre for Genomic Regulation) ‘Why don’t oocytes get Alzheimer’s?’
Diego Sainz de la Maza (University College London) ‘Somatic cells support germ cell survival by shuttling glycolytic products’
Güneş Taylor (Francis Crick Institute) ‘The role of FOXL2 in pregranulosa cell specification within the vertebrate ovary’
The webinar will be recorded to watch on-demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
What comes to mind when I say, “sea star”? For me, I think of easily accessible eggs that we can fertilized in vitro to make completely clear larvae that grow in a 6-well dish. Ah yes, I guess you were also thinking about snorkeling in a transparent ocean!
My name is Margherita Perillo and I am a Research Scientist at the MBL in beautiful Woods Hole right in Cape Cod. My research focuses mostly on understanding tissue and organ morphogenesis: How do individual cells group together to form complex organs? The system I chose to establish to investigate this question is the sea star Patiria miniata larva. In this short article, together with Zak Swartz (Assistant Scientist at MBL who also works with sea stars) and Jamie MacKinnon (Research Assistant from the Swartz Lab), we explain why we love this research animal.
Who works at the MBL?
The Marine Biological Laboratory is a vibrant year-round institute for research and teaching affiliated with the University of Chicago (Fig. 1). You may know us for our summer season, when we host advanced research training courses including the famous Embryology and Physiology courses, as well as visiting scientists and students from around the world, reaching a campus population of around 1,200 people. But throughout the year, MBL is home to over 30 resident faculty and laboratories across three departments, including Ecosystems Center, the Josephine Bay Paul Center for Comparative Molecular Biology and Evolution, and the Eugene Bell Center for Regenerative Biology and Tissue Engineering. Our research community spans different length scales and disciplines, from biomedical cell biology to ecosystem-level interactions. In addition, the MBL offers immersive undergraduate courses, including the Semester in Environmental Science and the new Semester in Biological Discovery, and a brand new Ph.D. program in conjunction with the University of Chicago.
Figure 1. The MBL Campus. Credit Marine Biological Laboratory
Patiria miniatain the wild
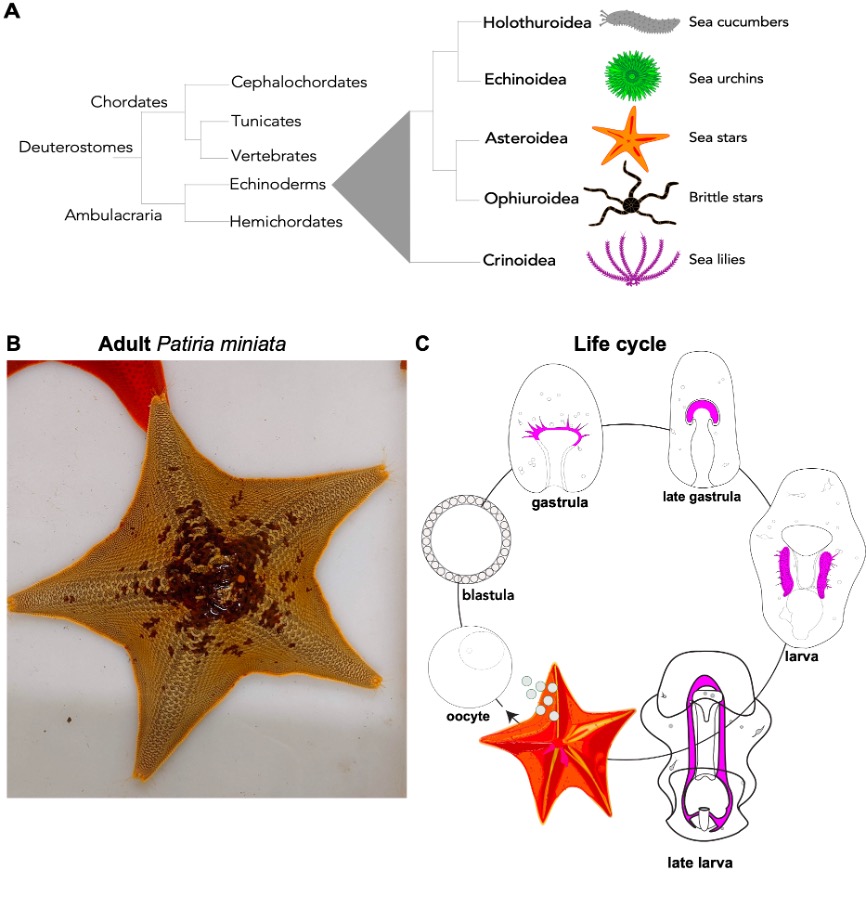
Sea stars are echinoderms, a group of bilaterian animals that includes sea urchins, sea stars, sea lilies, brittle stars, and sea cucumbers. Because of their close relationship with vertebrates, these animals are great models to ask biomedical questions, as the basic cellular and developmental mechanisms that we study in sea stars are conserved in vertebrates (Fig. 2A). The sea star Patiria miniata (Fig. 2B) can be found all along the Pacific Coast, from Alaska to Mexico in deep and shallow waters 1,2. We get our animals from divers in California who ship us sea stars that we keep in big tanks in the MBL Marine Resource Center. Here a team of sea star experts takes care of them to make sure they enjoy their stay and have the best possible accommodations in Cape Cod.
Life cycle: Females and male adult sea stars live in groups and when the season is right, they release their gametes out in the ocean where fertilization happens (Fig. 2C). There are gametes are in each arm and if we are lucky we find a female with six arms -extra ovaries for us! Embryos and larvae of P. miniata go through gastrulation and transform into planktonic larvae. After a few months, the larvae undergo metamorphosis to create a tiny, juvenile sea star. A remarkable feature of sea stars (and all other echinoderms) is that while their adult body has a pentameric plan, their larvae are bilateral, meaning that if we draw a line in the center of the larva there is a left and a right side, like us!
Figure 2. A) Phylogenetic tree showing relationship of echinoderms to other deuterostomes. There are five families of echinoderms and they all have a 5-fold symmetry (only a coincidence?). B) Adults Patiria miniata come in different colors that range from red, to purple, orange or beige (Picture of one of our sea stars at MBL, Credits Margherita Perillo). C) P. miniata life cycle, from eggs or sperm released from adults to larval stages that eventually undergo metamorphosis and transform into a tiny sea star juvenile (Cartoon modified from Perillo et. al, 2023).
Patiria miniatain the laboratory
One of the best parts about working with sea stars is that they are incredibly easy to culture and bring through metamorphosis. A normal week in the lab begins with a trip to the Marine Resources Center (MRC) to visit our adult sea stars, check their health, and collect gonads (Fig. 3A). We carefully make a 1mm ventral incision and extract a piece of ovary; these pieces are cultured in antibiotic-treated seawater and safely kept ex-vivo for weeks at 15°C3,4.
When we need to expand our larval cultures, we use a dissecting needle to tease open the ovary, remove any eggs we need for the day, and add hormone to induce maturation (Fig. 3B). After fertilization, the early-stage cell divisions will happen in just a few hours. Two days later they will have developed into swimming larvae which can be transferred into 500mL boxes and fed with a red and green algal cocktail. If we change the water biweekly and continue this feeding pattern, we can observe bipinnaria larvae beginning to metamorphose within a few months. At this time we begin to feed larvae small pieces of Aquanix kelp flakes, containing spirulina, and several sources of protein. The juvenile sea stars are very low maintenance and continue to grow larger and more motile day by day!
Figure 3. A) We keepthe sea starP. miniata in big tanks with sea water at 15°C in the MRC facility at MBL. B) Jamie working with sea star ovaries under a dissection scope.
P. miniata, an emerging system to understand organ morphogenesis
Our body is composed of many organs with diverse functions. What do they all have in common? Well, virtually all organs derive from epithelial tubes. During organogenesis these simple tubes grow, branch and elongate to make complex organs like lungs, kidneys, heart, pancreas and more. If this first step of making a tube goes wrong the embryo will develop with major birth defects with one or more organs that are shorter, have the wrong orientation in the embryo and do not function properly5-7.
Because of the fundamental role that epithelial tubes have in building our organs a key question is: What are the mechanisms that drive proper outgrowth and elongation of epithelial tube? And what can be a good model to address this question?
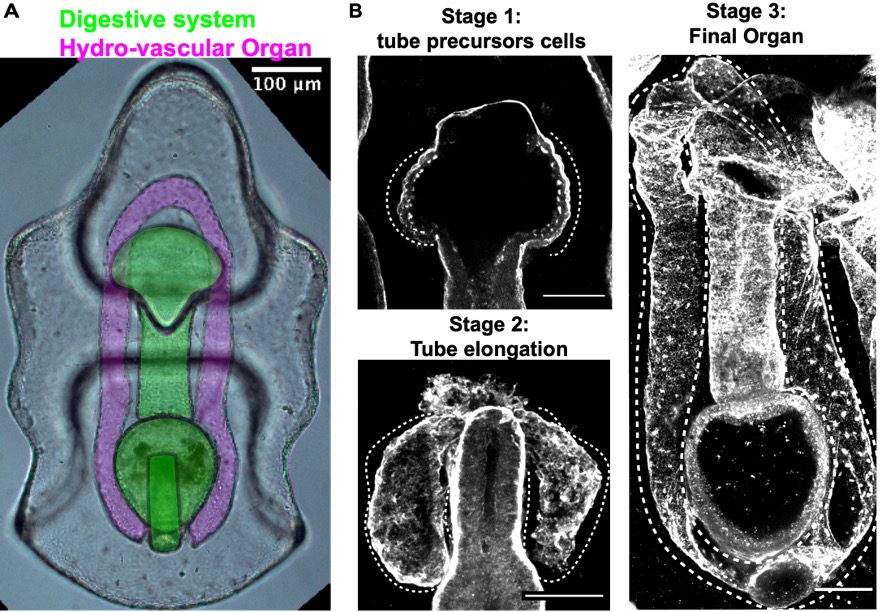
While vertebrates have many, complex and highly branched organs all tightly packed together, the sea star larva has only two simple and optically clear organs: a digestive system and the hydro-vascular organ, (HVO) (Fig. 4). In my recent work, I develop two important tools that allowed us to use this new system to study how tubes form: long-term live imaging (to look at cell movements) and I set up the first CRISPR Cas9 protocols for a sea star (to perturb gene function)8-10.
The HVO is the perfect epithelial tube: we found that it starts as a sheet of cells that bud off the digestive system (stage 1) to form two parallel tubes (stage 2) that elongate, make one branch and eventually fuse to form a looped organ (stage 3). HVO functions might be related to larval buoyancy in the water column11 and I’m investigating if this is its only function.
We used the HVO as a model to define aspects of tube morphogenesis that were still poorly defined, like for instance: What drives tube elongation? We found that the FGF pathway is a major driver of tube outgrowth and that this pathway also controls branch point formation through the transcriptional factor Six1/2. Using live imaging we investigated the mechanics of tube elongation and found that cells of the growing tube actively migrate and at the same time divide to allow for tube extension and expansion. This is relevant from a biomedical perspective, as these steps are conserved with mammals 8.
Figure 4. A) A live sea star larva where digestive system and hydro-vascular organ (HVO) are highlighted. These are the only two organs of this organism and follow a stereotypical growth. B) Stages of HVO development (laminin staining).
Sea stars for fundamental reproductive biology
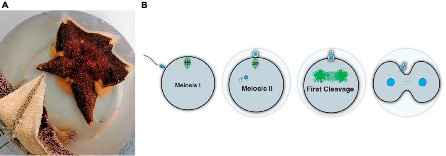
In the lab of Zak Swartz, we work with sea stars to explore fundamental reproductive processes from a cell biological perspective. In contrast to mammals, which undergo reproductive aging and have limited fecundity, the sea star produces millions of new oocytes throughout its (30 year+) lifespan through adult oogenesis (Figure 5A). This is a practical advantage, as having such abundant access to ovary tissue and oocytes lowers the barriers to doing our experiments. But it also fascinating biology: how do sea stars manage to continuously produce so many oocytes whereas humans are born with a limited set? Periklis Paganos is leading a project that uses single-cell genomics to define the cell type repertoire that drives this reproductive longevity, and cell biological approaches to understand how these cells interact with each other. Our goal is to define the signaling interactions and cellular states that support a long reproductive lifespan, which we hope will help inform human fertility treatments.
Another special aspect of working with sea stars is their status as ecologically important animals. As predators and keystone species, they have an outsized impact on food webs. Like many other marine invertebrates, sea stars release their eggs directly into the seawater, with minimal protection against any fluctuations in the environment. Yet, they are fertilized and must accurately perform meiotic and mitotic processes to form an embryo under these conditions (Figure 5B). Jamie MacKinnon is asking how resilient sea star reproduction is to climate change, including variables such as temperature. By comparing eggs from different species, we aim to identify predictive measures for how marine eggs and early embryos will respond to extreme climate fluctuations. We are also working developing new genetic tools for sea stars, an effort led by Akshay Kane in our lab, and Nat Clarke at MIT, that we hope will make sea stars and other echinoderms more accessible for the research community – stay tuned!
Figure 5. A) Adult female sea star spawning out thousands of eggs, visible as the orange material emanating from between the arms. B) A summary of cell division processes between fertilization and the first embryonic cleavage that we study in our lab.
Patiria miniata combines a biomedically relevant phylogenetic position, genetic tools for functional analysis and a lot of oocytes and embryos available year-round -we are excited to learn more from these model organisms in the future.
1 Ebert, T. A. Life-History Analysis of Asterinid Starfishes. The Biological Bulletin241, 231-242, doi:10.1086/716913 (2021).
2 Morris, R. H., Abbott, D. P. & Haderlie, E. C. Intertidal invertebrates of California. Vol. 200 (Stanford University Press Stanford, 1980).
3 Swartz, S. Z. et al. Quiescent cells actively replenish CENP-A nucleosomes to maintain centromere identity and proliferative potential. bioRxiv, 433391 (2018).
4 Pal, D., Visconti, F., Sepúlveda-Ramírez, S. P., Swartz, S. Z. & Shuster, C. B. Use of echinoderm gametes and early embryos for studying meiosis and mitosis. Mitosis: Methods and Protocols, 1-17 (2022).
5 Ely, D. M. & Driscoll, A. K. Infant Mortality in the United States, 2020: Data From the Period Linked Birth/Infant Death File. Natl Vital Stat Rep71, 1-18 (2022).
6 Baldwin, D. & Yadav, D. in StatPearls (StatPearls Publishing
7 Eitler, K., Bibok, A. & Telkes, G. Situs Inversus Totalis: A Clinical Review. Int J Gen Med15, 2437-2449, doi:10.2147/ijgm.S295444 (2022).
8 Perillo, M., Swartz, S. Z., Pieplow, C. & Wessel, G. M. Molecular mechanisms of tubulogenesis revealed in the sea star hydro-vascular organ. Nature Communications14, 2402, doi:10.1038/s41467-023-37947-2 (2023).
9 Oulhen, N., Pieplow, C., Perillo, M., Gregory, P. & Wessel, G. M. Optimizing CRISPR/Cas9-based gene manipulation in echinoderms. Dev Biol490, 117-124, doi:10.1016/j.ydbio.2022.07.008 (2022).
10 Perillo, M., Swartz, S. Z. & Wessel, G. M. A conserved node in the regulation of Vasa between an induced and an inherited program of primordial germ cell specification. Dev Biol482, 28-33, doi:10.1016/j.ydbio.2021.11.007 (2022).
11 Potts, W. T. The physiological function of the coelom in starfish larvae and its evolutionary implications. Physiol Biochem Zool76, 771-775, doi:10.1086/381463 (2003).
Come join us next year in France to discuss all things Neural Crest: From patient to model system and back agan. This conference is organised by the Ph.D students of the ITN ‘NEUcrest’ aiming to highlight the works of early career researchers. We have exciting speakers, a session on living with a Neurocristopathy and a great location. Most importantly, the first 30 students to sign up to our conference, get a 50% discount. We look forward to meeting you!Scan the QR code or follow this link to register on our website: https://neucrestfinalconference.org
In a recent Development paper, Wan-Sheng Liu and colleagues find that the cancer/testis antigen PRAMEL1 regulates spermatogonial development by inhibiting retinoic acid signaling, playing a crucial role in the proper establishment of the first and subsequent rounds of spermatogenesis. We caught up with first author Mingyao Yang to find out more about the story behind the paper.
Mingyao, what prompted you to join Wansheng’s lab at the Pennsylvania State University?
During my time at China Agricultural University, I nurtured a profound fascination for reproductive biology. My passion was further ignited through my involvement in a national reproductive lab, where I delved into researching female reproductive biology. As I contemplated pursuing graduate studies in the USA, I came across Dr. Liu’s lab, in the Center for Reproductive biology and Health (CRBH) at Penn State. Although Dr. Liu’s expertise centered on male reproduction, I saw this as a distinctive chance to expand my horizons into uncharted territory. I recognized this opportunity as a platform to enrich my knowledge and skills, providing a stimulating environment for my personal and academic growth. What struck me most was Dr. Liu’s personalized mentorship. He meticulously identified my weaknesses, provided direct guidance, and helped me address each of them individually. This tailored mentorship not only inspired me but also affirmed that I was on the right path to personal growth and a successful research career.
CRBH faculties direct a dynamic and interactive graduate and postgraduate training program and conduct research in diverse areas of reproductive biology and endocrinology. Within this atmosphere, young researchers benefit from interactive learning experiences facilitated by a cohesive team of reproductive experts, engaging in cutting-edge research. In this collaborative space, students, from various labs, use shared equipment to explore diverse scientific questions. The environment fosters extensive discussions, collaborations, and mutual support among our researchers, enriching our educational journey and enhancing our research endeavors. These reasons really attracted me to Dr. Liu’s lab.
How did the project get started?
PRAME (Preferentially Expressed Antigen in Melanoma) was first discovered in melanoma cells in 1997 (Ikeda et al., 1997). Subsequent research revealed that PRAME can multiply across different chromosomes during evolution, forming a multicopy gene family in eutherian animals (Chang et al., 2011). Human, mouse, and bovine genomes contain approximately 60, 90, and 60 copies of PRAME, respectively. Since its discovery, over 500 papers have been published on the Prame family, with most focusing on cancer biology and only a few on reproduction. Our laboratory contributes to unravel the roles of the Prame family in reproduction.
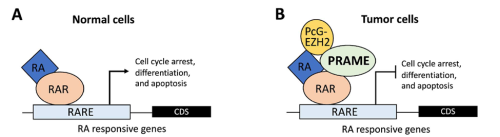
In cancer biology, PRAME serves as a biomarker for various types of cancers (Epping et al., 2005; Kern et al., 2021). Its molecular function involves inhibiting the retinoic acid receptor (RAR) signaling pathway, blocking differentiation, and promoting proliferation (Epping et al., 2005). In germline development, PRAME members (PRAMEL7 and PRAMEL19) counteract retinoic acid (RA)-dependent differentiation, maintaining naïve pluripotency in embryonic stem cells (Casanova et al., 2011; Graf et al., 2017; Napolitano et al., 2020). In spermatogenesis, PRAMEF12 is known to regulate the number of spermatogonia stem cells (SSCs), although its specific molecular functions remain unstudied (Wang et al., 2019).
Previous studies in Dr. Liu’s lab revealed that PRAMEL1 expression is enriched in the testes, particularly in spermatogenic cells ranging from spermatogonia to mature spermatozoa (Liu et al., 2021; Mistry et al., 2013). Based on this information, we hypothesized that PRAMEL1 might be involved in spermatogenesis by inhibiting the RA signaling pathway.
To test this hypothesis, we generated Pramel1 conditional and global knockout mice, forming the basis for this project.
Excerpts from previous paper (Kern et al., 2021)Excerpts from previous paper (Liu et al., 2021)
What was known about the role of retinoic acid signaling in spermatogenesis before your work?
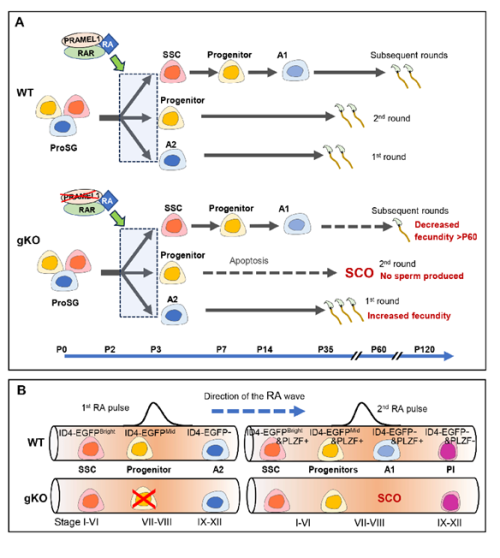
Retinoic acid (RA) signaling plays a crucial role in male reproduction and is essential for spermatogenesis (Griswold, 2016). Animals deficient in RA exhibit spermatogonia arrest and infertility. Retinoic acid drives at least four germ cell transitions during spermatogenesis (Endo et al., 2017; Griswold, 2016). In mice, the first transition occurs a few days after birth (around postnatal day 3 (P3)), transforming prospermatogonia into three subtypes of spermatogonia: SSCs (spermatogonial stem cells), progenitors, and A1 spermatogonia (Busada et al., 2014). A1 spermatogonia continue developing to initiate the first round of spermatogenesis, progenitors initiate the second round, while SSCs prepare for subsequent rounds (Law et al., 2019). The first pulse of RA initiates this initial germ cell transition.
Additionally, during each round of spermatogenesis, RA pulses stimulate spermatogonia differentiation, spermatocyte meiosis, spermatid elongation, and the release of spermatozoa from the seminiferous epithelium.
Can you summarize the findings in a paragraph?
In this study, we examined the underlying cellular and molecular mechanisms of PRAMEL1 during spermatogenesis. We reported findings on the involvement of PRAMEL1 in the initiation and maintenance of spermatogenesis by analyzing mouse models with either global or conditional Pramel1 inactivation. We found that:
Pramel1 plays a crucial role in regulating RA responsiveness of cell-fate committed prospermatogonia, maintaining a balance between undifferentiated and differentiating spermatogonia during the initial round of spermatogenesis.
Pramel1 has a more pronounced effect on progenitors than on other subtypes of germ cells in young males. It also plays a role in maintaining undifferentiated spermatogonial populations in mature mice.
PRAMEL1 affects progenitor homing process during the initiation of spermatogenesis in neonatal testis.
Pramel1 deficiency led to an increased fecundity in juvenile males and decreased fecundity in mature males.
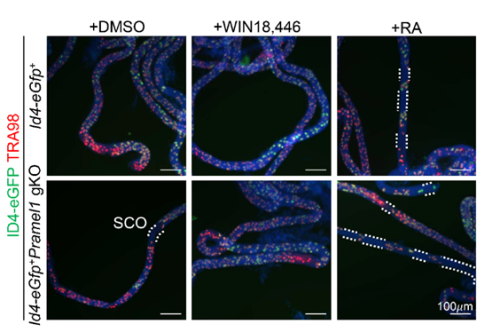
Pramel1 deficiency resulted in a regional Sertoli cell-only (SCO) phenotype during the first round of spermatogenesis, which was rescued by administration of the RA inhibitor WIN18,446, suggesting that PRAMEL1 functions as an inhibitor of RA signaling in germ cells.
Overall, our findings suggest that PRAMEL1 fine-tunes RA signaling, playing a crucial role in the establishment of the first and subsequent rounds of spermatogenesis.
The role of PRAMEL1 during the establishment of spermatogenesis. (A) A model for RA responsiveness in three germ cell lineages in the wild-type and Pramel1– deficient mice (for details, see paper). The critical time points during germ cell development in mice are indicated below the model. (B) A proposed model for the establishment of stages I to stage XII of the seminiferous epithelial cycle in a neonatal wild-type and Pramel1 gKO testis. A1 and A2, A1 and A2 spermatogonia; Pl, preleptotene spermatocyte; ProSG, prospermatogonia; SCO, Sertoli cell-only; SSC, spermatogonial stem cell.
Were you surprised to find that Pramel1 deficiency affected juvenile and mature mice differently?
Certainly, we were surprised by these findings, as we did not anticipate the divergent function of PRAMEL1 in young animals compared to older ones.
Interestingly, a novel concept has emerged indicating that the first round of spermatogenesis constitutes a distinct program separate from the subsequent rounds (Law et al., 2019; Yoshida et al., 2006). During the first round, sperms are produced at a juvenile age, whereas the subsequent rounds of spermatogenesis occur during mature age. The initial A2 spermatogonia, transitioning directly from prospermatogonia in response to the first RA pulse, drives the first round of spermatogenesis. In contrast, the subsequent rounds of spermatogenesis originate from spermatogonial stem cells (SSCs). Our results provide compelling evidence supporting the idea that the mechanisms underlying the first round and subsequent rounds of spermatogenesis are different.
Did you have any particular result or eureka moment that has stuck with you?
During this work, I often felt like walking in a maze. There are too many unsolved mysteries during spermatogenesis. However, fresh results and inventive ideas, whether derived from literature, expert insights, or our own discussions, served as beacons of guidance, illuminating our path, and bringing moments of clarity amidst the intricate complexity.
One of the exciting moments was when we obtained the whole-mount immunofluorescence (IFS) results following RA treatment. These results revealed that the RA inhibitor successfully rescued the regional SCO phenotype in the young gKO testis. This outcome strongly suggested that PRAMEL1 acts as an inhibitor of RA signaling during spermatogenesis. Typically, my advisor, Dr. Wansheng Liu, and I often have different interpretations or perspectives regarding my results. However, this time, he wholeheartedly agreed with me when we examined the original results, and our shared enthusiasm underscored the significance of our findings.
Whole-mount immunofluorescent staining with TRA98 (red) antibodies on seminiferous tubules. White dashed lines outline SCO regions. Nuclei are counterstained with DAPI (blue).
And the flipside: were there any moments of frustration or despair?
Certainly, graduate school, especially for international students like me, came with its fair share of frustrating moments. One such instance involved the extensive immunofluorescence staining required for our research paper. Initially, the staining procedures didn’t yield high-quality results, possibly due to issues with our protocol or the antibodies we were using. I persisted in optimizing our protocol, conducting the staining repeatedly in an attempt to improve the outcomes. Simultaneously, I experimented with numerous antibodies sourced from different companies. Complicating matters, our funding was limited at that time, requiring us to approach these companies and request free samples of antibodies, which we tested one by one. The journey to completing this project was arduous, but it was also incredibly motivating to witness the quality of results improving gradually with each attempt.
What’s next for this story? And personally, Mingyao, what is next for you after this paper?
In this study, we understand the role of PRAMEL1 during spermatogenesis while our previous study has revealed? the function of PRAMEX1 in testis. To gain a better understanding of the role of PRAME family during spermatogenesis, we have successfully generated a Pramel1/Pramex1 double knockout mice. Thus, the next of this story is to figure out the interaction of the two different members of Prame family during spermatogenesis.
For me, I will further explore the mystery during spermatogenesis in the lab of Dr. Oatley (one of our co-authors in this paper) in Washington State University. My career goal is to become an independent investigator researching the mechanisms that underpin spermatogenesis. My hope is that the outcomes of my research program will be translated to solutions for male infertility that impacts humans, domestic animals, and wildlife. Infertility is a significant concern that affects a substantial number of people worldwide, with approximately 20% of couples facing difficulties conceiving a pregnancy. Through my research in male reproductive biology, I aim to contribute to the development of innovative solutions and interventions to address the male side of infertility. This involves investigating the underlying causes of male infertility, treatment options, and improving assisted reproductive technologies. By gaining a deeper understanding of reproductive processes and disorders, I hope to make meaningful contributions to improving fertility outcomes and enhancing the quality of life for individuals and families facing fertility challenges.
Reference:
Busada, J. T., Kaye, E. P., Renegar, R. H., & Geyer, C. B. (2014). Retinoic acid induces multiple hallmarks of the prospermatogonia-to-spermatogonia transition in the neonatal mouse. Biology of Reproduction, 90(3), 1–11. https://doi.org/10.1095/biolreprod.113.114645
Casanova, E. A., Shakhova, O., Patel, S. S., Asner, I. N., Pelczar, P., Weber, F. A., Graf, U., Sommer, L., Bürki, K., & Cinelli, P. (2011). Pramel7 mediates LIF/STAT3-dependent self-renewal in embryonic stem cells. Stem Cells, 29(3), 474–485. https://doi.org/10.1002/stem.588
Chang, T., Yang, Y., Yasue, H., Bharti, A. K., Retzel, E. F., & Liu, W. (2011). The Expansion of the PRAME Gene Family in Eutheria. 6(2). https://doi.org/10.1371/journal.pone.0016867
Endo, T., Freinkman, E., De Rooij, D. G., & Page, D. C. (2017). Periodic production of retinoic acid by meiotic and somatic cells coordinates four transitions in mouse spermatogenesis. Proceedings of the National Academy of Sciences of the United States of America, 114(47), E10132–E10141. https://doi.org/10.1073/pnas.1710837114
Epping, M. T., Wang, L., Edel, M. J., Carlée, L., Hernandez, M., & Bernards, R. (2005). The human tumor antigen PRAME is a dominant repressor of retinoic acid receptor signaling. Cell, 122(6), 835–847. https://doi.org/10.1016/j.cell.2005.07.003
Graf, U., Casanova, E. A., Wyck, S., Dalcher, D., Gatti, M., Vollenweider, E., Okoniewski, M. J., Weber, F. A., Patel, S. S., Schmid, M. W., Li, J., Sharif, J., Wanner, G. A., Koseki, H., Wong, J., Pelczar, P., Penengo, L., Santoro, R., & Cinelli, P. (2017). Pramel7 mediates ground-state pluripotency through proteasomal-epigenetic combined pathways. Nature Cell Biology, 19(7), 763–773. https://doi.org/10.1038/ncb3554
Griswold, M. D. (2016). Spermatogenesis: The commitment to Meiosis. Physiological Reviews, 96(1), 1–17. https://doi.org/10.1152/physrev.00013.2015
Ikeda, H., Lethe, B., Baren, N. Van, Smet, C. De, Vitale, M., Moretta, A., Boon, T., Coulie, P. G., Istologia, I., & Biomediche, S. (1997). Characterization of an Antigen That Is Recognized on a Melanoma Showing Partial HLA Loss by CTL Expressing an NK Inhibitory Receptor. Immunity, 6, 199–208.
Kern, C. H., Yang, M., & Liu, W. S. (2021). The PRAME family of cancer testis antigens is essential for germline development and gametogenesis. Biology of Reproduction, 105(2), 290–304. https://doi.org/10.1093/biolre/ioab074
Law, N. C., Oatley, M. J., & Oatley, J. M. (2019). Developmental kinetics and transcriptome dynamics of stem cell specification in the spermatogenic lineage. Nature Communications, 10(1), 1–14. https://doi.org/10.1038/s41467-019-10596-0
Liu, W. S., Lu, C., & Mistry, B. V. (2021). Subcellular localization of the mouse PRAMEL1 and PRAMEX1 reveals multifaceted roles in the nucleus and cytoplasm of germ cells during spermatogenesis. Cell and Bioscience, 11(1), 1–18. https://doi.org/10.1186/s13578-021-00612-6
Mistry, B. V, Zhao, Y., Chang, T., Yasue, H., Chiba, M., & Oatley, J. (2013). Differential Expression of PRAMEL1 , a Cancer / Testis Antigen , during Spermatogenesis in the Mouse. 8(4). https://doi.org/10.1371/journal.pone.0060611
Napolitano, G., Tagliaferri, D., Fusco, S., Cirillo, C., De Martino, I., Addeo, M., Mazzone, P., Russo, N. A., Natale, F., Cardoso, M. C., De Luca, L., Lamorte, D., La Rocca, F., De Felice, M., & Falco, G. (2020). A novel member of Prame family, Gm12794c, counteracts retinoic acid differentiation through the methyltransferase activity of PRC2. Cell Death and Differentiation, 27(1), 345–362. https://doi.org/10.1038/s41418-019-0359-9
Wang, Z., Xu, X., Li, J. L., Palmer, C., Maric, D., & Dean, J. (2019). Sertoli cell-only phenotype and scRNA-seq define PRAMEF12 as a factor essential for spermatogenesis in mice. Nature Communications, 10(1). https://doi.org/10.1038/s41467-019-13193-3
Yoshida, S., Sukeno, M., Nakagawa, T., Ohbo, K., Nagamatsu, G., Suda, T., & Nabeshima, Y. (2006). The first round of mouse spermatogenesis is a distinctive program that lacks the self-renewing spermatogonia stage. 1505, 1495–1505. https://doi.org/10.1242/dev.02316
Thank you to all the talented people who submitted their artwork and to everyone who has visited the art exhibition so far. The exhibition is open until the end of November 2023.
We asked you, the Node community, to vote for your favourite artwork from the exhibition. We also asked a panel of judges from the BSDB and The Company of Biologists to choose their favourite — and the results are now in!
Category: Scientific images
Judges’ choice
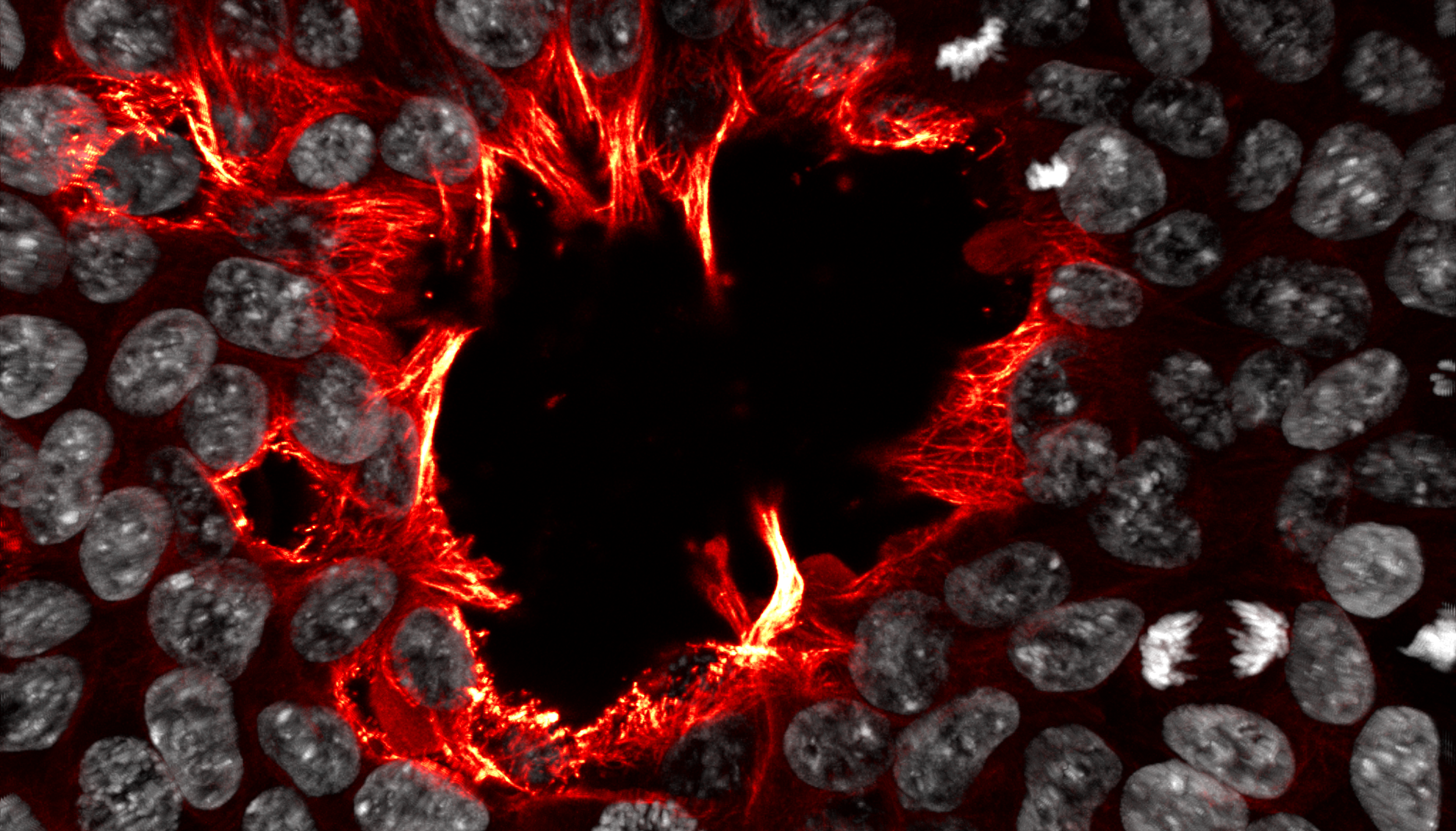
The Maw at Etna Oliver Anderson (Australian Regenerative Medicine Institute) In this image, microtubules are shown in red/yellow, and nuclei in white. Cells rush to fill an opening in the colony, with their jagged flame-like microtubules roaring into the centre like the devouring forge-flames of Cyclopean Etna. (Aeneid Book VIII: Lines 416-425) Human induced pluripotent stem cells, imaged using a Zeiss LSM780 confocal microscope. Cells are labelled with DAPI (white), and immunostained for alpha-tubulin (red-yellow).
Judges’ choice runner-up and People’s choice
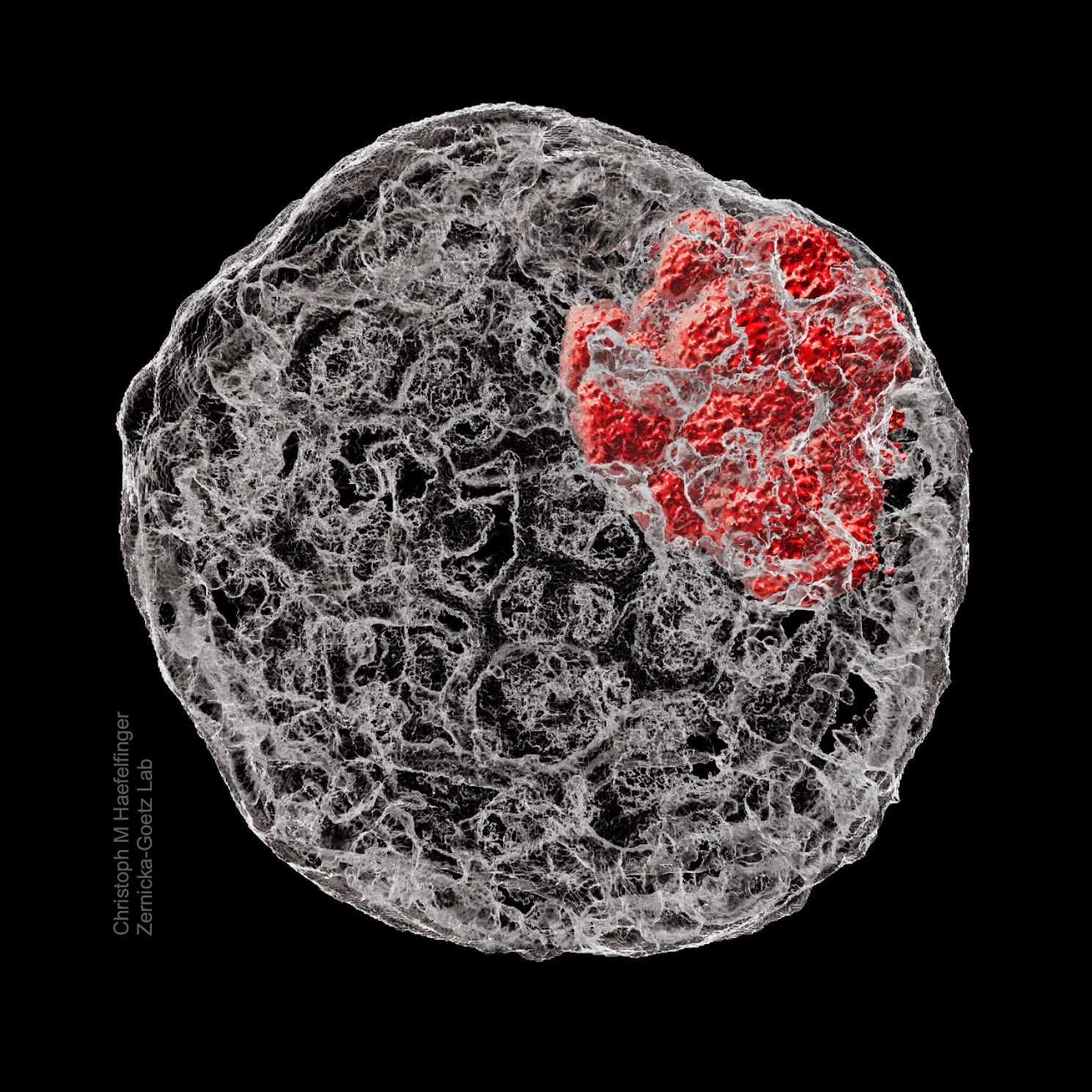
The backbone of stem cell derived embryos Christoph Markus Haefelfinger (California Institute of Technology) The cytoskeletal structure of preimplantation embryos demonstrated in a reconstruction of a stem cell derived mouse blastoid. After fixation, the structure was immunostained for f-actin (phalloidin, grey) and the inner cell mass (Oct4, red), then imaged.
Category: Science-inspired art
Judges’ choice and People’s choice
Klimt-olotl Elad Bassat (Research Institute of Molecular Pathology, IMP) The decision of Axolotls to stay in water rather than metamorphose. As I am in Elly Tanaka’s lab in Vienna I drew it in the style of the Austrian Gustav Klimt.
Judges’ choice runner-up
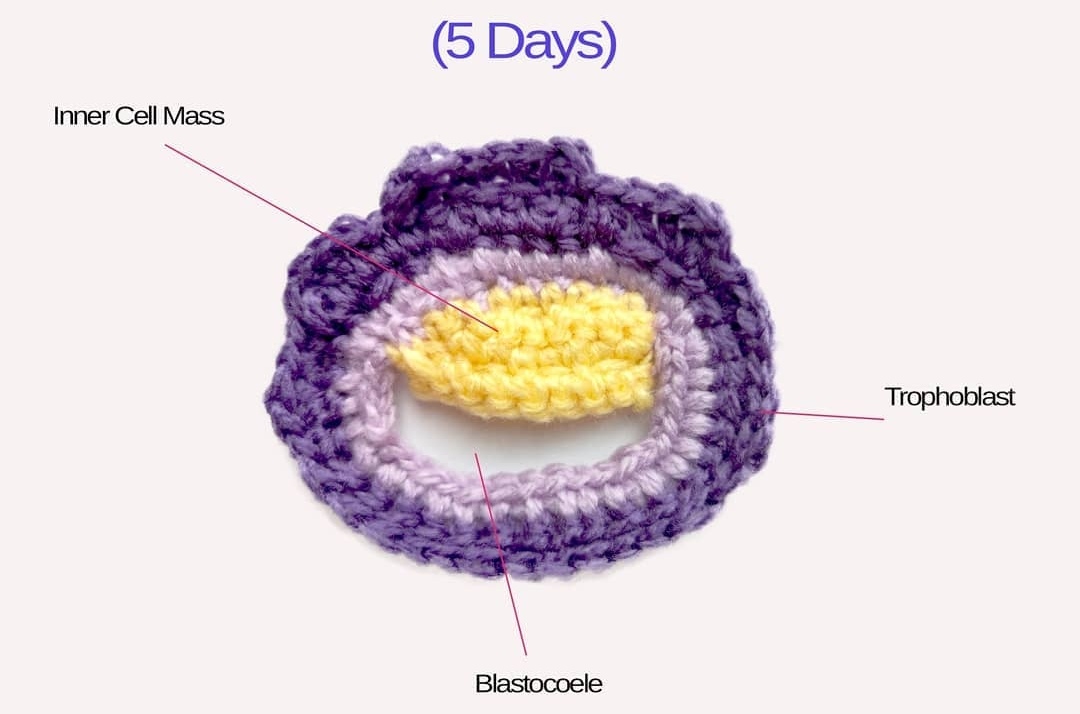
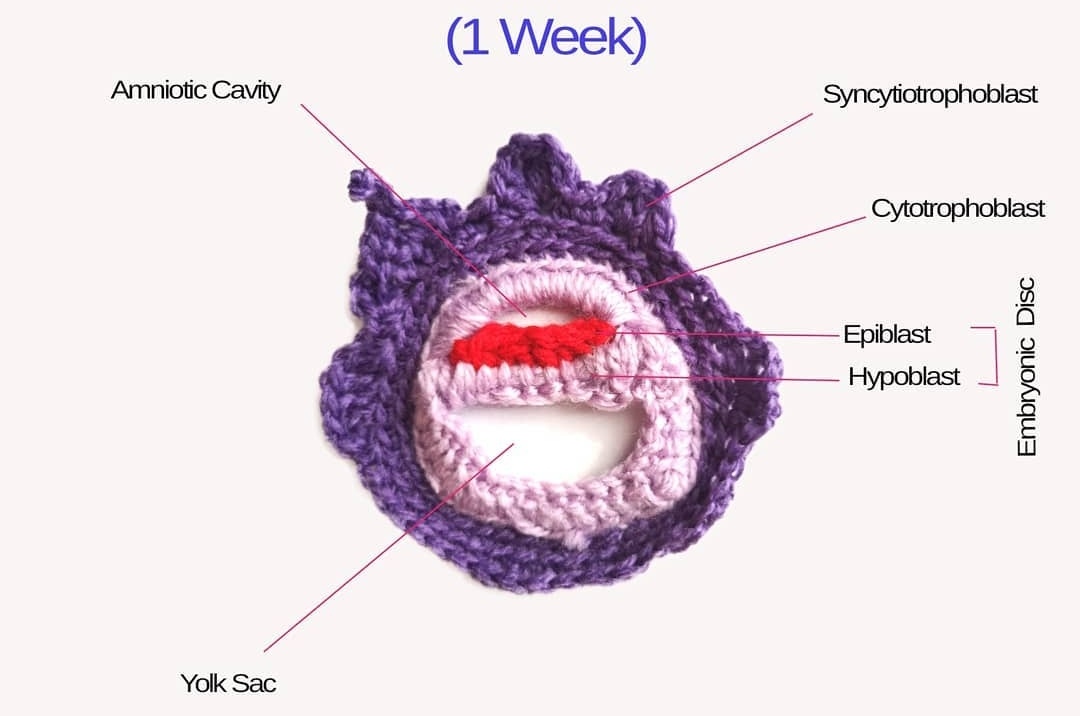
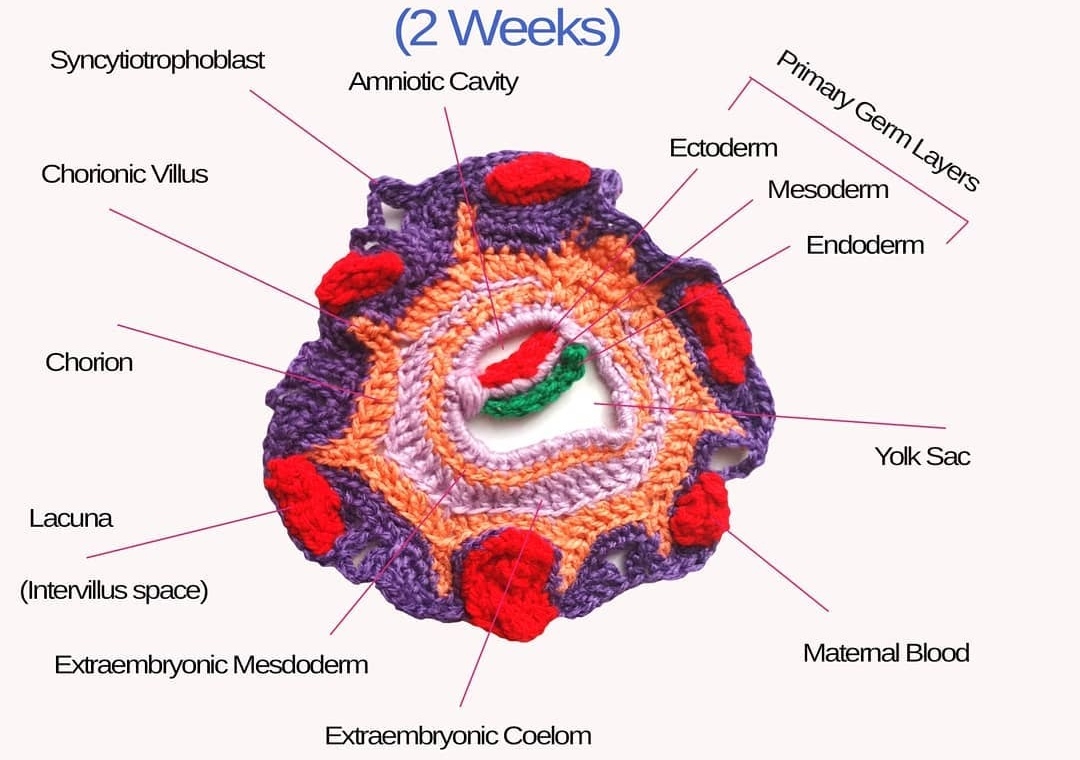
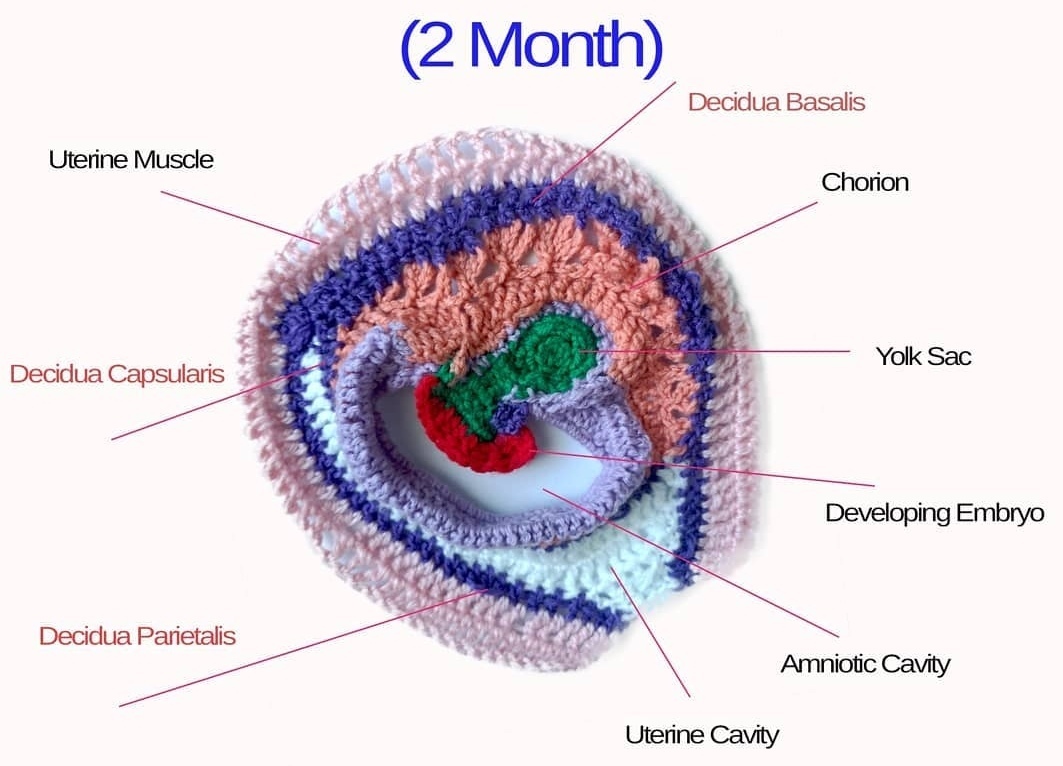
Crocheted models of embryonic development Tahani Baakdhah (Krembil Research Institute, University of Toronto) Crocheted model of 5 days, 1 week, 2 weeks and 2 months gestational embryonic development.
(Click on individual image to see the full size version)
Category: Art by Scientists
Judges’ choice
Living Water Brent Foster (Whitney Laboratory for Marine Bioscience) Ink drawing of an octopus. When I see these creatures glide across a surface, I almost think of them as living water.
Judges’ choice runner-up
Castle of Dreaming Dragons Morag Lewis (King’s College London) This began as a small pencil sketch in 2019, and grew into a multipage creation over the year until it was finished in June 2020. It was inked digitally, then printed in sections and painted using watercolours.
People’s choice
Nautilus Brent Foster (Whitney Laboratory for Marine Bioscience Chalk drawing of a nautilus.
The webinar on 25 October 2023 was chaired by Development Associate Editor Irene Miguel-Aliaga (Imperial College London) and features three early-career researchers studying metabolism and development. This webinar coincided with the completion of Development’s Special Issue: Metabolic and Nutritional Control of Development and Regeneration. Below are the recordings of the talks.
Maria Fernanda (Nanda) Forni (Yale University)
Talk and Q&A by Maria Fernanda (Nanda) Forni
Hidenobu Miyazawa (EMBL Heidelberg)
Talk and Q&A by Hidenobu Miyazawa
Siamak Redhai (DKFZ)
Talk and Q&A by Siamak Redhai (1 votes) Loading...
Luka Jarc, Manuj Bandral, Elisa Zanfrini, Mathias Lesche, Vida Kufrin, Raquel Sendra, Daniela Pezzolla, Ioannis Giannios, Shahryar Khattak, Katrin Neumann, Barbara Ludwig, Anthony Gavalas
John R. Klem, Tae-Hwi Schwantes-An, Marco Abreu, Michael Suttie, Raeden Gray, Hieu Vo, Grace Conley, Tatiana M. Foroud, Leah Wetherill, CIFASD, C. Ben Lovely
Surag Nair, Mohamed Ameen, Laksshman Sundaram, Anusri Pampari, Jacob Schreiber, Akshay Balsubramani, Yu Xin Wang, David Burns, Helen M Blau, Ioannis Karakikes, Kevin C Wang, Anshul Kundaje
Adam David Langenbacher, Fei Lu, Luna Tsang, Zi Yi Stephanie Huang, Benjamin Keer, Zhiyu Tian, Alette Eide, Matteo Pellegrini, Haruko Nakano, Atsushi Nakano, Jau-Nian Chen
Jeannine Basta, Lynn Robbins, Lisa Stout, Michelle Brennan, John Shapiro, Mary Chen, Darcy Denner, Angel Baldan, Nidia Messias, Sethu Madhavan, Samir V. Parikh, Michael Rauchman
Vi Pham, Livia Sertori Finoti, Margaret M. Cassidy, Jean Ann Maguire, Alyssa L. Gagne, Elisa A. Waxman, Deborah L. French, Kaitlyn King, Parith Wongkittichotee, Xinying Hong, Lars Schlotawa, Beverly L. Davidson, Rebecca C. Ahrens-Nicklas
Archana Kamalakar, Brendan Tobin, Sundus Kaimari, Afra I. Toma, Irica Moriarity, Surabhi Gautam, Pallavi Bhattaram, Shelly Abramowicz, Hicham Drissi, Andrés J. García, Levi B. Wood, Steven L. Goudy
Bongjun Kim, Yuanjian Huang, Kyung-Pil Ko, Shengzhe Zhang, Gengyi Zou, Jie Zhang, Moonjong Kim, Danielle Little, Lisandra Vila Ellis, Margherita Paschini, Sohee Jun, Kwon-Sik Park, Jichao Chen, Carla Kim, Jae-Il Park
Shoichiro Takeishi, Tony Marchand, Wade R. Koba, Daniel K. Borger, Chunliang Xu, Chandan Guha, Aviv Bergman, Paul S. Frenette, Kira Gritsman, Ulrich Steidl
Alice Santambrogio, Yasmine Kemkem, Thea L. Willis, Ilona Berger, Maria Eleni Kastriti, Louis Faure, John P. Russell, Emily J. Lodge, Val Yianni, Rebecca J. Oakey, Barbara Altieri, Stefan R. Bornstein, Charlotte Steenblock, Igor Adameyko, Cynthia L. Andoniadou
Elvira H. de Laorden, Diana Simón, Santiago Milla, María Portela-Lomba, Marian Mellén, Javier Sierra, Pedro de la Villa, María Teresa Moreno-Flores, Maite Iglesias
Ethan Tietze, Andre Rocha Barbosa, Bruno Henrique Silva Araujo, Veronica Euclydes, Hyeon Jin Cho, Yong Kyu Lee, Arthur Feltrin, Bailey Spiegelberg, Alan Lorenzetti, Joyce van de Leemput, Pasquale Di Carlo, Tomoyo Sawada, Gianluca Ursini, Kynon J. Benjamin, Helena Brentani, Joel E. Kleinman, Thomas M. Hyde, Daniel R. Weinberger, Ronald McKay, Joo Heon Shin, Apua C.M. Paquola, Jennifer A. Erwin
Manon Jaffredo, Nicole A. J. Krentz, Benoite Champon, Claire E. Duff, Sameena Nawaz, Nicola Beer, Christian Honore, Anne Clark, Patrik Rorsman, Jochen Lang, Anna L. Gloyn, Matthieu Raoux, Benoit Hastoy
Rosalie Sinclair, Minmin Wang, Zaki Jawaid, Jesse Aaron, Blair Rossetti, Eric Wait, Kent McDonald, Daniel Cox, John Heddleston, Thomas Wilkop, Georgia Drakakaki
David Bolumar, Javier Moncayo-Arlandi, Javier Gonzalez-Fernandez, Ana Ochando, Inmaculada Moreno, Ana Monteagudo-Sanchez, Carlos Marin, Antonio Diez, Paula Fabra, Miguel Angel Checa, Juan Jose Espinos, David K Gardner, Carlos Simon, Felipe Vilella
Daniel Frank, Maria Bergamasco, Michael Mlodzianoski, Andrew Kueh, Ellen Tsui, Cathrine Hall, Georgios Kastrappis, Anne Kathrin Voss, Catriona McLean, Maree C Faux, Kelly Rogers, Bang Tran, Elizabeth Vincan, David Komander, Grant Dewson, Hoanh Tran
Martina Jabloñski, Guillermina M. Luque, Matías D. Gómez-Elías, Claudia Sanchez-Cardenas, Xinran Xu, Jose Luis de la Vega-Beltran, Gabriel Corkidi, Alejandro Linares, Victor X. Abonza Amaro, Dario Krapf, Diego Krapf, Alberto Darszon, Adan Guerrero, Mariano G. Buffone
Bernard Y. Kim, Hannah R. Gellert, Samuel H. Church, Anton Suvorov, Sean S. Anderson, Olga Barmina, Sofia G. Beskid, Aaron A. Comeault, K. Nicole Crown, Sarah E. Diamond, Steve Dorus, Takako Fujichika, James A. Hemker, Jan Hrcek, Maaria Kankare, Toru Katoh, Karl N. Magnacca, Ryan A. Martin, Teruyuki Matsunaga, Matthew J. Medeiros, Danny E. Miller, Scott Pitnick, Sara Simoni, Tessa E. Steenwinkel, Michele Schiffer, Zeeshan A. Syed, Aya Takahashi, Kevin H-C. Wei, Tsuya Yokoyama, Michael B. Eisen, Artyom Kopp, Daniel Matute, Darren J. Obbard, Patrick M. O’Grady, Donald K. Price, Masanori J. Toda, Thomas Werner, Dmitri A. Petrov
Zhisong He, Leander Dony, Jonas Simon Fleck, Artur Szałata, Katelyn X. Li, Irena Slišković, Hsiu-Chuan Lin, Malgorzata Santel, Alexander Atamian, Giorgia Quadrato, Jieran Sun, Sergiu P. Paşca, J. Gray Camp, Fabian Theis, Barbara Treutlein
Michael J Cotton, Pablo Ariel, Kaiwen Chen, Vanessa A Walcott, Michelle Dixit, Keith A Breau, Caroline M Hinesley, Kasia Kedziora, Cynthia Y Tang, Anna Zheng, Scott T Magness, Joseph Burclaff
Nusayhah Hudaa Gopee, Ni Huang, Bayanne Olabi, Chloe Admane, Rachel A Botting, April Rose Foster, Fereshteh Torabi, Elena Winheim, Dinithi N Sumanaweera, Issac Goh, Mohi Miah, Emily Stephenson, Win Min Tun, Pejvak Moghimi, Ben Rumney, Peng He, Sid Lawrence, Kenny Roberts, Keval Sidhpura, Justin Englebert, Laura Jardine, Gary Reynolds, Antony Rose, Clarisse Ganier, Vicky Rowe, Sophie Pritchard, Ilaria Mulas, James Fletcher, Dorin-Mirel Popescu, Elizabeth FM Poyner, Anna Dubois, Andrew Filby, Steven Lisgo, Roger A Barker, JONG-EUN PARK, Roser Vento-Tormo, Phuong Ahn Le, Sara Serdy, Jin Kim, CiCi Deakin, Jiyoon Lee, Marina T Nikolova, Neil Rajan, Stephane Ballereau, Tong Li, Josh Moore, David Horsfall, Daniela Basurto Lozada, Edel A O’Toole, Barbara Treutlein, Omer Bayraktar, Maria Kasper, Pavel Mazin, Laure Gambardella, Karl Koehler, Sarah Teichmann, Muzlifah Haniffa
Henry Tat Quach, Spencer Farrell, Kayshani Kanagarajah, Michael Wu, Xiaoqiao Xu, Prajkta Kallurkar, Andrei Turinsky, Christine Bear, Felix Ratjen, Sidhartha Goyal, Theo J Moraes, Amy Wong
Patrice Pottier, Malgorzata Lagisz, Samantha Burke, Szymon M. Drobniak, Philip A. Downing, Erin L. Macartney, April Robin Martinig, Ayumi Mizuno, Kyle Morrison, Pietro Pollo, Lorenzo Ricolfi, Jesse Tam, Coralie Williams, Yefeng Yang, Shinichi Nakagawa
“Shift work might be a real driver of health inequality; that these people are, as a function of their job, being forced to effectively mess with their biology”
Dr Priya Crosby
In the latest episode of the Genetics Unzipped podcast, we’re clocking in to chat about the genetics of circadian rhythms. How can molecules tell the time, why don’t we have drugs for jet lag yet and could a midnight snack stop malaria in its tracks?
Our November webinar will be chaired by Development’s Reviews Editor, Alex Eve, and features three early-career researchers studying the intersection between development and disease. The webinar will be held using Zoom with a Q&A session after each talk.
Tuesday 14 November 2023 – 14:00 GMT
Mauricio Rocha-Martins (Instituto Gulbenkian de Ciência) ‘How tissues orchestrate growth and morphogenesis: Lessons from the vertebrate retina’
Nicole Edwards (Cincinnati Children’s Hospital Medical Center) ‘Discovering the developmental basis of trachea-esophageal birth defects: evidence for endosomeopathies’
Cecilia Arriagada (Rutger’s University) ‘Role of mesodermal fibronectin in mechanotransduction during cardiac development’
Writing in Nature, Bill Jia, Sean Megason, Adam Cohen, and colleagues ask the question: how does the heart go from silent to beating during embryonic development? Using genetically encoded sensors and optogenetic tools, they captured the very first heartbeat of a zebrafish embryo and dissected the biophysical basis for its timing and spatial structure. We chatted with first author Bill Jia to hear the story behind the paper.
How did you come to join the labs of Adam Cohen and Sean Megason?
Early in my PhD, I developed a fascination with how cells compute as a collective. Embryonic development jumped out to me as an incredible example of this phenomenon – cells must talk to each other and then make their own decisions based on the inputs they receive, yet the outcomes of these decisions are often patterns and functions at a level of organisation higher than the individual. The Megason lab’s goal is to identify the conceptual rules by which this comes to be, so it was a perfect fit. However, to understand the complex messages that cells send to each other, it is critical to find the channels through which this information is passed – what exactly are cells measuring? In developmental biology, a major emphasis has traditionally been placed on signal transduction of chemical morphogens. But this is just one of many processes that constitute cellular physiology, each with its own time and length scales over which it can transmit or process information. The Cohen lab has pioneered optical approaches to measure and perturb different aspects of cellular physiology. It also focuses on neuroscience, another discipline concerned with complex and collective computation which happens to exemplify the idea that patterns with different spatiotemporal scales can carry information in biology. I wanted to bring together the distinct strengths and shared interests of these labs. This has put me in a unique position to study developmental cell-cell communication and pattern formation in media that have historically been relatively inaccessible to biological inquiry – bioelectricity, second messenger signalling, and others.
Movie 1: Calcium dynamics across zebrafish gastrulation, convergence, and extension, recorded using the sensor jGCaMP6s.
How did the project get started?
We started out with a broad survey of different physiological dynamics in zebrafish development, using biosensors to look for patterns that might be correlated between cells across time and space. Our efforts imaging cellular calcium were particularly successful (Movie 1)! Consistent with previous work from the Miller lab and others (1), we found an enormous diversity of calcium transients in different cell types over early development. We struggled for a while to decide which of these patterns would be worth following up on because there were simply so many. Adam proposed looking at the first heartbeats because the purpose of the calcium dynamics in this system was known, so it would just be a matter of figuring out how they appeared. At first, I was concerned that too much would have been previously studied, but the literature suggested that there were still some major mysteries to be solved.
What was known about the emergence of the first heartbeat in vertebrate embryos before your work?
The question of how the heartbeat emerges can be broken down into a few pieces. At what time in development does it happen? What are the roles of different cells in the process? How does the heart first gain its regular timing, and how is its function coordinated across the entire tissue? It was already known that periodic electrical and calcium activity emerges in the heart much earlier in development than it is needed (e.g. for blood flow) (2). It was also previously suggested, based on surgical separation, dissociation, or genetic perturbation of heart morphogenesis, that most early cardiomyocytes had the capacity to beat on their own and that the fastest cell would synchronise the tissue (3–5). But reports differed on the exact details of these observations, which we think mainly owes to two reasons.
First, individual heartbeats are very fast compared to the progression of development – it is difficult to ascertain from a short recording of a few beats how far along the process any given animal is. As we saw in our measurements, there could be very large changes in the dynamics over very short developmental periods. Second, disruption of the tissue geometry by the methods described is also highly disruptive to electrophysiology, making any observed activity preparation-dependent. One simple and instructive example is to imagine a cell that expects to be coupled to its neighbour by gap junctions – after dissection of that interface, it now has hemichannels exposed to the extracellular fluid instead, which might completely change its electrical behaviour. Notwithstanding these potentially contradictory previous results, there was also no proposed mechanism for how the heart gains its timing, whose quantitative properties are integral to the organ’s function.
Can you summarize the findings of the paper?
Movie 2: Multiplexed recordings of the heart’s transition from silent to beating in zebrafish embryos, taken with the sensor jGCaMP7f (6).
First, we used long, multiplexed recordings of genetically encoded calcium sensors to efficiently capture the transition from silent to beating (including the very first heartbeat!) in many zebrafish hearts (Movie 2). This was a dramatic and sudden switch, which in combination with subsequent experiments to probe biophysical and molecular mechanism gave us confidence that something important had happened at that very moment. We then looked at the statistics of the beats after the first one and found that the trajectories of average beat rate and beat rate variability were quite stereotyped. This lent itself to a simple mathematical abstraction describing how oscillations can emerge from an apparently silent system, called a stochastic saddle-node on invariant-circle (SNIC) bifurcation.
This model has two important predictions – first, highly periodic oscillations can be driven by random noise; second, big changes in fast dynamics (beats) can emerge from very small changes in an underlying control parameter describing a decreasing distance between a resting state and a “threshold” for a single oscillation. This second idea, of small changes in one quantity resulting in big changes in another, is analogous to a miniscule change in temperature across the freezing point turning water into ice – which is why we describe it as a phase transition. Similar phenomena have been observed in tissue mechanical properties in development (7, 8), and perhaps more await discovery.
The abstract mathematical resting state and threshold described have a biological basis in the membrane voltage dynamics of many types of electrically excitable cells, including cardiomyocytes. We tested whether this was relevant in the earliest heartbeats by performing voltage imaging and perturbing various ion transporters. In a panel of several genes important in the adult spontaneous cardiac action potential, we found that only the L-type voltage-gated calcium channel (VGCC) was required for both voltage and calcium oscillations. This is consistent with the model – the voltage threshold for activation of the VGCC could be the threshold described in the equations.
We then used light-activated ion channels (channelrhodopsins) to see if we could optogenetically cross the predicted threshold before any endogenous activity could be observed. As the model predicted, the heart became increasingly easy to excite as the first spontaneous beat approached. Furthermore, local excitations could propagate across the tissue, explaining how many cells seemed to turn on all at once from the first beat (Movie 3) – the cells are primed and ready to respond to their neighbours.
Movie 3: Tissue-scale initiation of calcium activity (cyan) in the zebrafish heart cone (red), taken with calcium sensor jGCaMP7f.
Our observations on how the heart’s earliest temporal patterns were driven by noise and how the spatial patterns were always synchronised demanded a more careful analysis of functional specialisation in individual cells. If the threshold crossing is random, what permits a particular cell to do it? We know that the adult heart has a defined pacemaker region, and we knew from previous work where these cells ought to be at the time of the first heartbeat. Does the beat start in the same place every time, and is this where the pacemaker will be? How does the previously observed widespread spontaneity play into all of this? We performed imaging of the calcium activity together with markers defining cardiomyocyte subpopulations, showing that the spatial origin was indeed variable, but tended to start away from the future pacemakers. Using optogenetics we were able to establish that a fast intrinsic beat rate was in fact the biophysical mechanism that set the spatial origin.
Together our results provide a picture of how many aspects of heartbeat initiation fit together. Random noise allows the heartbeats to emerge from variable positions, but the cells are synchronised by frequency-based competition. This is likely implemented molecularly by a gradual increase in activation sensitivity of VGCCs. This process does not seem to be tightly coupled to the earliest steps of pacemaker development, but the biophysical mechanisms are sufficient to ensure early coordination and periodicity of the heartbeat.
Were you surprised to find that the individual heart cells abruptly start beating all at once?
Yes! Before we did the experiments, the analogy I had in my head was a swarm of fireflies. I was expecting to see individual cells blinking separately, with some sort of synchronisation process occurring gradually over time. How the heartbeat initiation could hypothetically play out depends on the relative ordering of the development of excitability, electrical coupling, and spontaneous oscillations – in my picture, the coupling would have had to emerge last. There is evidence that connexins are expressed very early on in heart development (9), so maybe the tissue-scale initiation ought to have been less surprising. But the dependence of the spatiotemporal dynamics on each of these features is quantitative, so someone who had started with that prior assumption could have just as likely been wrong – we had to do the measurements to find out!
Do you think this bioelectrical phase transition model is found across all developing vertebrate hearts?
It is very possible, given that many features of the transition have been suggested by other observations in different vertebrate models. The same L-type calcium channels are known to be required for early cardiac activity in mice (10), and similar observations of lability in the spatial origin of the heartbeat have been seen in chicks and mice (3, 10). One major way other species might differ is in the length scale of synchronisation. In animals with larger hearts, it may be possible to observe multiple regions of synchronisation with their own distinct origin points.
The paper involves optogenetic manipulations and live functional imaging, producing a lot of beautiful videos. Can you tell us more about the techniques and tools you used?
Many of our conclusions rested on measuring how the dynamics of the heartbeat changed in their response to fast perturbations over the slow timescales of development. To do this, it was essential to simultaneously manipulate and image the electrical activity without disrupting the heart’s developmental trajectory. A key enabler of these experiments was FR-GECO1c, a novel far-red calcium sensor developed by the labs of Robert Campbell and Yi Shen (11). This sensor offered spectral compatibility with several blue light-activated ion channels (channelrhodopsins), which allowed us to target the heart as accurately as light can be patterned. Using these tools, we reversibly stimulated or silenced the heart at scales of milliseconds and micrometres, while watching its response in real time. There has been a recent explosion in new optogenetic tools for manipulating and visualising cellular physiology beyond membrane potential, including activation of canonical signalling pathways (12). I think that these will push the field of developmental biology forward tremendously, as they open up causative studies of the spatial and temporal couplings that ultimately underlie the progression of organ patterning.
Can you postulate the function of early electrical activity even before the heart is connected to the circulatory system?
It could be a type of developmental checkpoint for later steps in heart development – now that the heart has established its basic electrical function, it is ready to perform later steps like specification of cell subtypes or the complex steps of morphogenesis involved in chamber formation. The distribution of spontaneity could be a mechanism of robustness in this process – if one cell that was supposed to drive the beat fails to develop, another one can take its place. One particularly interesting hypothesis is that calcium signalling downstream of the beats supports development (10, 13). Given that calcium is an important second messenger molecule for many cellular processes, it is unclear whether it is the calcium fluxes downstream of the beat that matter and whether these must be deconvolved from some other cellular calcium dynamics. This is a problem that applies to all second messenger signalling pathways, so the developing heart might be a useful model to teach us more about the general mechanisms of cellular computation.
Did you have any particular result or eureka moment that has stuck with you?
The success of the quantitative fit of our data by the SNIC bifurcation model was very gratifying. It’s really satisfying to me when important features of a complex biological process can be distilled down into a model that is simple, yet precise enough to be written as a mathematical formula. But one of my favourite things about this story was how interdisciplinary the findings are – I really believe there is a little something for everyone in there. It’s super fun to see feedback from developmental biologists, cardiologists, and physicists expressing excitement about completely different parts of the results.
And the flipside: were there any moments of frustration or despair?
At times I thought that our findings were too simple or obvious given what was already known about the developing heart. But something I’ve come to appreciate is that this is very subjective and depends strongly on how much one is willing to make assumptions based on what is already known.
What’s next for this story? And what’s next for you personally?
I think there are two major future directions for this story. The first is to understand the implications of the early self-organising activity for the heart’s development, as we discussed. The second is to understand in more depth the molecular basis of the predicted biophysical mechanism. How are the VGCCs being sensitised? Is it because of changes in channel expression levels, relative intracellular and extracellular calcium levels, changes in resting potential by modulation of other ion pumps and channels, or something else? What is the source of the noise that seems to drive the timing? Personally, I am nearing the end of my PhD. I hope to continue along the lines of the first direction – how do properties of biological electricity instruct cell decision making and tissue patterning in development? I want to do so by opening my own lab but am also thinking about postdoctoral positions that might allow me to pursue these questions.
References
1. S. E. Webb, R. A. Fluck, A. L. Miller, Calcium signaling during the early development of medaka and zebrafish. Biochimie. 93, 2112–2125 (2011).
2. W. Rottbauer, K. Baker, Z. G. Wo, M.-A. P. K. Mohideen, H. F. Cantiello, M. C. Fishman, Growth and Function of the Embryonic Heart Depend upon the Cardiac-Specific L-Type Calcium Channel α1 Subunit. Dev. Cell. 1, 265–275 (2001).
3. T. Sakai, T. Yada, A. Hirota, H. Komuro, K. Kamino, A regional gradient of cardiac intrinsic rhythmicity depicted in embryonic cultured multiple hearts. Pflüg. Arch.437, 61–69 (1998).
4. J. F. Reiter, J. Alexander, A. Rodaway, D. Yelon, R. Patient, N. Holder, D. Y. R. Stainier, Gata5 is required for the development of the heart and endoderm in zebrafish. Genes Dev.13, 2983–2995 (1999).
5. R. L. DeHaan, Synchronization of pulsation rates in isolated cardiac myocytes. Exp. Cell Res.70, 214–220 (1972).
6. B. Z. Jia, Y. Qi, J. D. Wong-Campos, S. G. Megason, A. E. Cohen, A bioelectrical phase transition patterns the first vertebrate heartbeats. Nature, 1–7 (2023).
7. A. Mongera, P. Rowghanian, H. J. Gustafson, E. Shelton, D. A. Kealhofer, E. K. Carn, F. Serwane, A. A. Lucio, J. Giammona, O. Campàs, A fluid-to-solid jamming transition underlies vertebrate body axis elongation. Nature. 561, 401–405 (2018).
8. N. I. Petridou, B. Corominas-Murtra, C.-P. Heisenberg, E. Hannezo, Rigidity percolation uncovers a structural basis for embryonic tissue phase transitions. Cell. 184, 1914-1928.e19 (2021).
9. N. C. Chi, R. M. Shaw, B. Jungblut, J. Huisken, T. Ferrer, R. Arnaout, I. Scott, D. Beis, T. Xiao, H. Baier, L. Y. Jan, M. Tristani-Firouzi, D. Y. R. Stainier, Genetic and Physiologic Dissection of the Vertebrate Cardiac Conduction System. PLOS Biol.6, e109 (2008).
10. R. C. Tyser, A. M. Miranda, C. Chen, S. M. Davidson, S. Srinivas, P. R. Riley, Calcium handling precedes cardiac differentiation to initiate the first heartbeat. eLife. 5, e17113 (2016).
11. R. Dalangin, M. Drobizhev, R. S. Molina, A. Aggarwal, R. Patel, A. S. Abdelfattah, Y. Zhao, J. Wu, K. Podgorski, E. R. Schreiter, T. E. Hughes, R. E. Campbell, Y. Shen, bioRxiv, in press, doi:10.1101/2020.11.12.380089.
12. H. M. McNamara, B. Ramm, J. E. Toettcher, Synthetic developmental biology: New tools to deconstruct and rebuild developmental systems. Semin. Cell Dev. Biol.141, 33–42 (2023).
13. N. D. Andersen, K. V. Ramachandran, M. M. Bao, M. L. Kirby, G. S. Pitt, M. R. Hutson, Calcium signaling regulates ventricular hypertrophy during development independent of contraction or blood flow. J. Mol. Cell. Cardiol.80, 1–9 (2015).

 (1 votes)
(1 votes)


 (No Ratings Yet)
(No Ratings Yet)