The lab is located in the Cochin Institute (14th arrondissement of Paris), in between la Butte aux Cailles and Montparnasse.
2. Research summary
Antoine: We use the cranial neural crest as a model to understand how cell fate decision and cell plasticity are regulated during embryogenesis. This population presents a remarkable differentiation potential as it gives rise to the entire peripheral nervous system and to most of the craniofacial skeleton. We are also testing whether cranial neural crest plasticity could be harnessed to enhance postnatal bone regeneration.
The Zalc Lab
3. Can you give us a lab roll call, with a sentence including what each person works on and career stage
Jean-Christophe: Engineer – trust in sphere, spend money the PI collect, love, and hate expectation
Saverio: Master2 student – trying to understand neural crest regionalization: cares for his many little spheres, gets happy when they behave nicely, starts shouting in Italian when they do random stuff just to annoy him.
Martina: Master2 student – trying to not get crazy when she obtains just 5 ng/ul of RNA from 150 spheres, in love with multichannel pipettes
Laura: Postdoc – juggles with differentiating spheres, sees the light at the scope, pipets hard at the bench, wants to understand the tuning of Oct4 for cell plasticity, is driven by curiosity
Antoine: PI – does the paperwork, have multiple meetings every day, jumps and/or dance when new data arrive
4. Favourite technique, and why?
Antoine: Depends on the question we are asking.
If I had to choose it will be genetic lineage tracing. The embryo being the best teacher, this technic provides a clear understanding of the decision made by the cells, and it generates great images.
Then RNA in situ hybridization, works beautifully on whole embryo or sections. Allows to the questions what are cells asking before, during and after they made a choice.
5. Apart from your own research, what are you most excited about in developmental and stem cell biology
Antoine: We are reaching a stage where we are done naming things. We are now able to understand development with equations. The advances of single cell omics will soon allow us to reconstruct an entire embryo in sillico. With this, it will be possible to predict how an environmental or genetic perturbation affects development. This will also allow us to generate several scenarios to rescue the defect, which can be much later than when the problem occurs.
I’m also really excited by the rapid progress made with organoïds. I was skeptical at first, but this is becoming an excellent tool to study basic cell behavior and logic.
6. How do you approach managing your group and all the different tasks required in your job?
Antoine: This is a learning process… I’m a strong believer that science must be fun! We spend so much time and effort doing it we should at least enjoy the process. For it to be fun, this implies scientific rigor, work ethics, good technical skills and simply being humane.
I’m still learning about the different task required by the job! Using the one thing I learnt during my PhD and postdoc which is learning how to learn. Thankfully, I have greats colleagues in the Institute that can help with this!
7. What is the best thing about where you work?
Jean-Christophe: Multiplicity of help you can hope and offer.
Saverio: Everything in the lab is brand new so every little step feels like a big achievement!
Martina: There is always chocolate!
Laura: The Institute is included in the Université Paris-city network which gives access to all exciting seminars occurring in the network.
Antoine: The Cochin Institute is multidisciplinary which means I learn a lot about various topics which stimulates creativity and open the way to great collaborations on various topics
8. What’s there to do outside of the lab?
Jean-Christophe: Why do you want to do things outside of the lab? Are you not happy here?
Saverio: If you work in research, try climbing. You will meet so many colleagues that you will feel like you never left the lab. Bring your friends!
Martina: Many sunsets along the Seine with a beer.
Laura: We are in Paris, a lot of culture, art, good coffee, much more.
Antoine: Paris is outside the lab, there’s plenty to do! I take care of my kids, which is a second job on itself, and spend time with my family. When I have time, I try to keep doing photography, keeping an eye and the dynamic Parisian Street Art scene.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
In our latest SciArt profile we hear from Ivana Henry, a science communicator and illustrator with a background in developmental genetics. Ivana enjoys creating 3D visualisations of scientific concepts in molecular biology and genetics.
Where are you originally from and what do you work on now?
I was born in beautiful southern Czech Republic and studied genetics at the University of South Bohemia. After completing a PhD in developmental genetics in Bayreuth (Germany), I undertook a postdoc at the Max Planck Institute of Molecular Cell Biology and Genetics in Dresden (Germany). I studied the role of planar cell polarity in tissue/organ development using the Drosophila ovary as a model. Later on, I studied how force generation moves tissues. I also developed light-sheet microscopy imaging approaches to study subcellular processes in curved epithelia. At this time, I found my love of scientific illustration and 3D visualisation and started to offer scientific illustration services. I currently work as a laboratory manager at the MRC Laboratory of Molecular Biology in Cambridge in the UK.
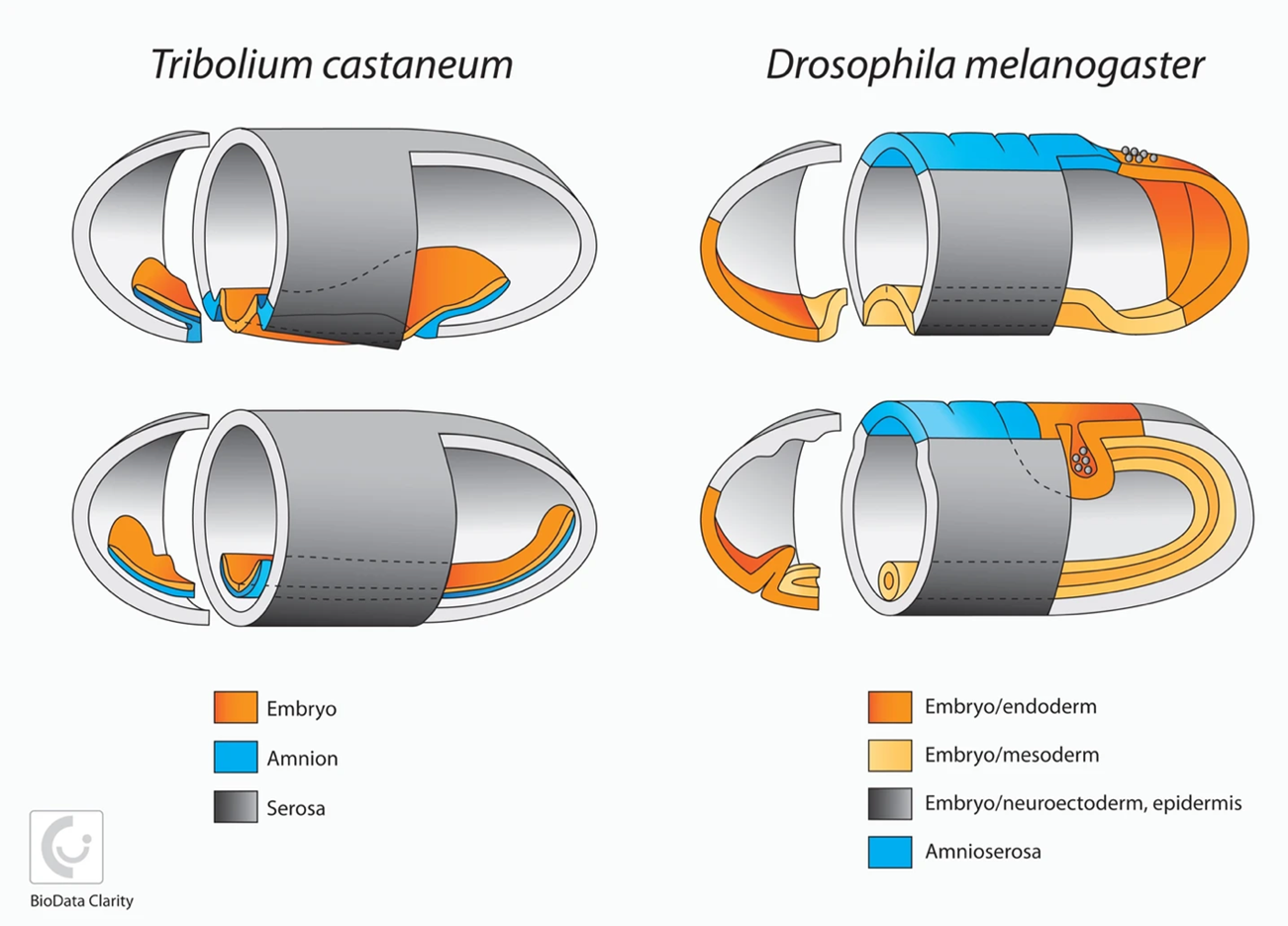
2D illustration showing a comparison of embryo development (Tribolium vs Drosophila)
Were you always going to be a scientist?
I don’t think so. As a child I liked all school subjects, so I think I probably wanted to be a teacher first. I liked art, languages, history, literature, and my interest in biology and chemistry came later. It was at secondary school/gymnasium that I discovered Louis Pasteur, Darwin’s theory and then molecular biology and genetics. I took part in biology Olympiads and became curious about insects and plants. The idea of becoming a scientist may have started to take shape in grammar school, but I didn’t seriously consider it until I was at university.
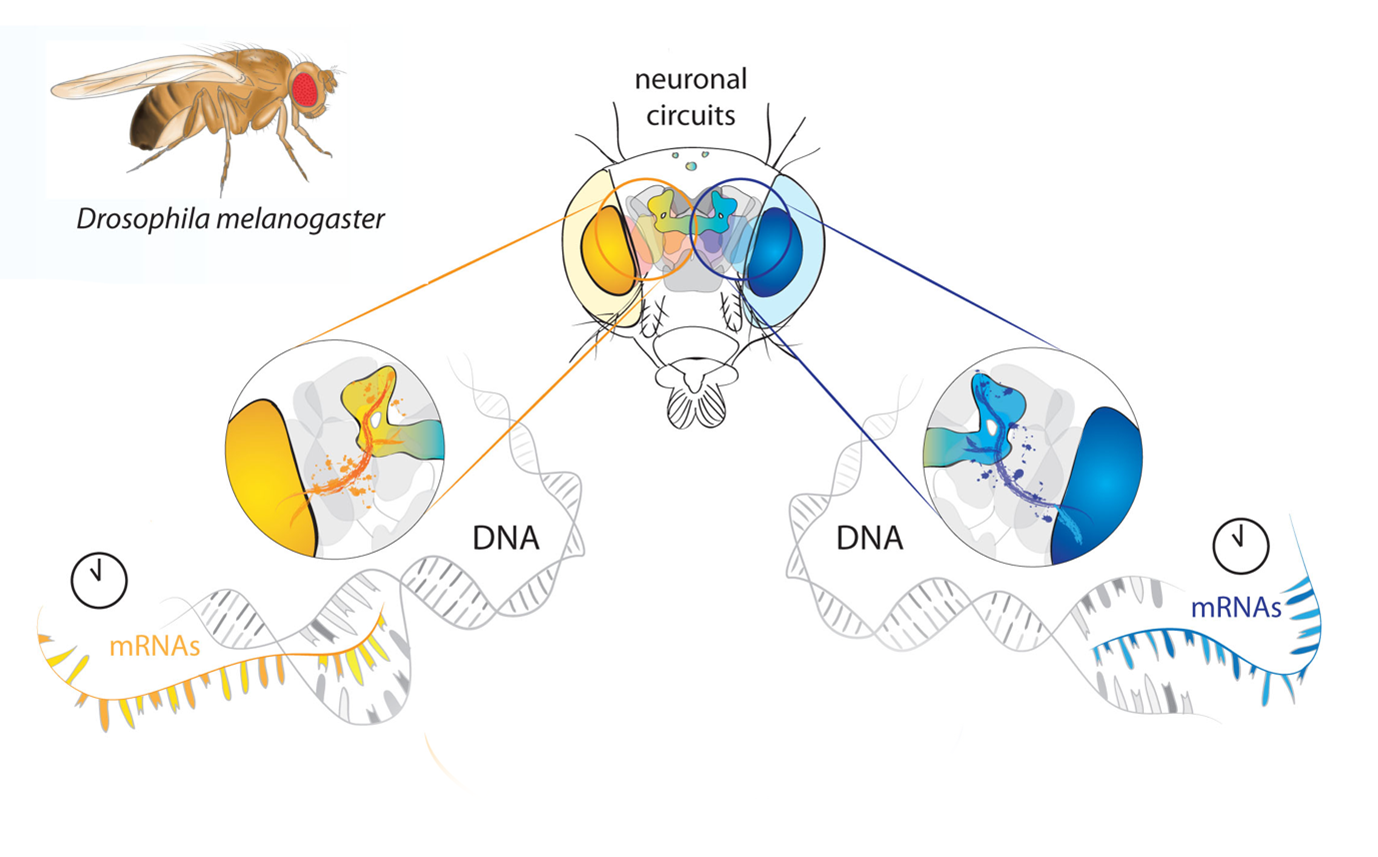
A detail of a graphical summary indicating a difference between the expression of the circadian clock genes in the summer and in the winter. Drosophila adult is digitally painted.
And what about art- have you always enjoyed it?
Yes, I always enjoyed art. As much as I loved going to natural history museums, I also loved going to art museums. Later on, I couldn’t resist visiting art museums in various destinations when I was at scientific conferences. Art simply has been always part of my life. It started with drawing portraits of my family and led to drawing portraits of my whole gymnasium class. But I never thought I was an artist or that I’d like to become one. I was drawn to the world of molecular biology and genetics. I wanted to understand how things work. But art comes through in different forms in my scientific journey. When I was at university, I used to draw an overview of the study material for each subject I was studying. This helped me to commit newly acquired knowledge into long-term memory. I think it is important to visualise scientific knowledge and I find it easier to remember information in the form of a picture rather than as long scientific text.
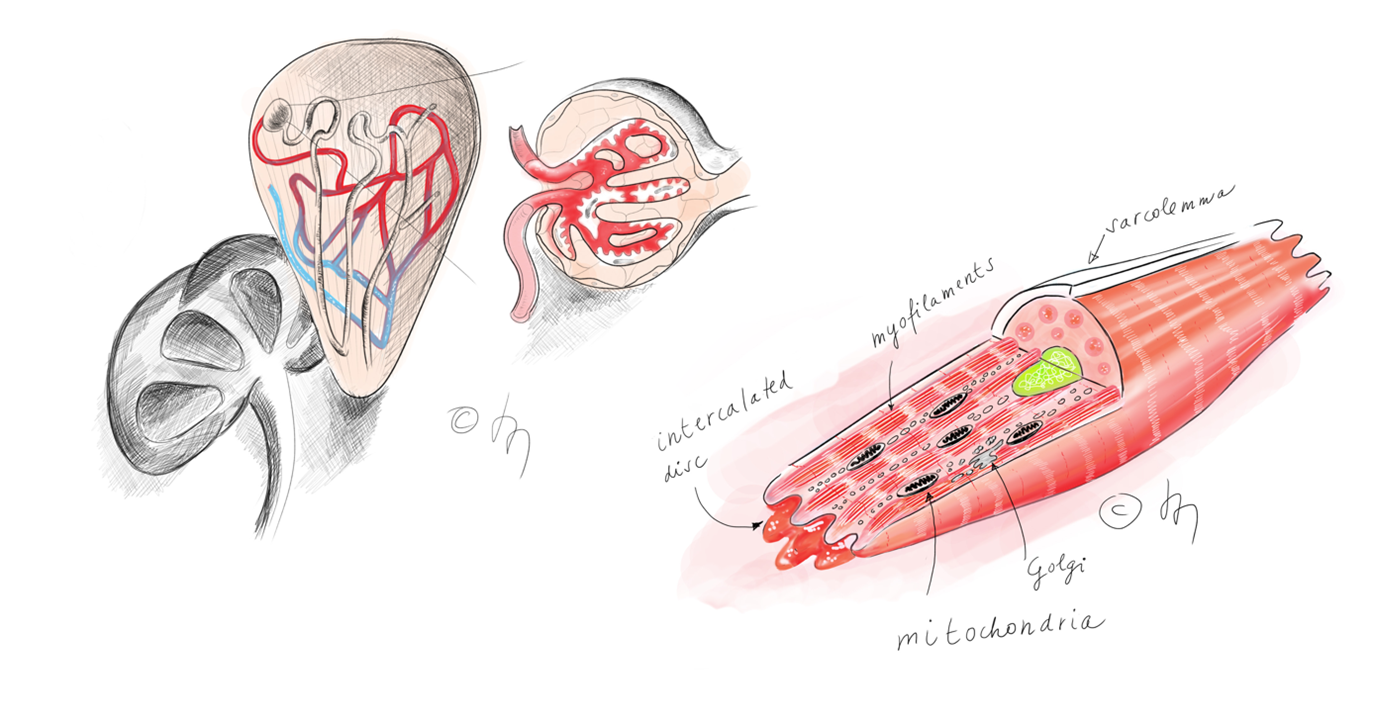
Digital drawings of a kidney showing nephron and glomerulus (on the left) and a cardiomyocyte (on the right).
What and who are your most important artistic influences?
One of my favourite artists is our Czech Art Nouveau artist, Alphonse Mucha. I like his detailed graphic and illustrative style and admire his dreamy paintings of The Slavic Epic on such a large scale. I’m not sure this influence is visible in my art, but his works often serve as my inspiration. I also very much like Impressionism, especially Claude Monet and his later paintings, including the Water Lilies series as they are almost abstract art. Bright and complementary colours used by impressionists usually reflect in my colour code.
In addition, I’m often inspired by new digital approaches from contemporary 3D artists and several renowned biomedical studios creating visuals for industry / the medical field.
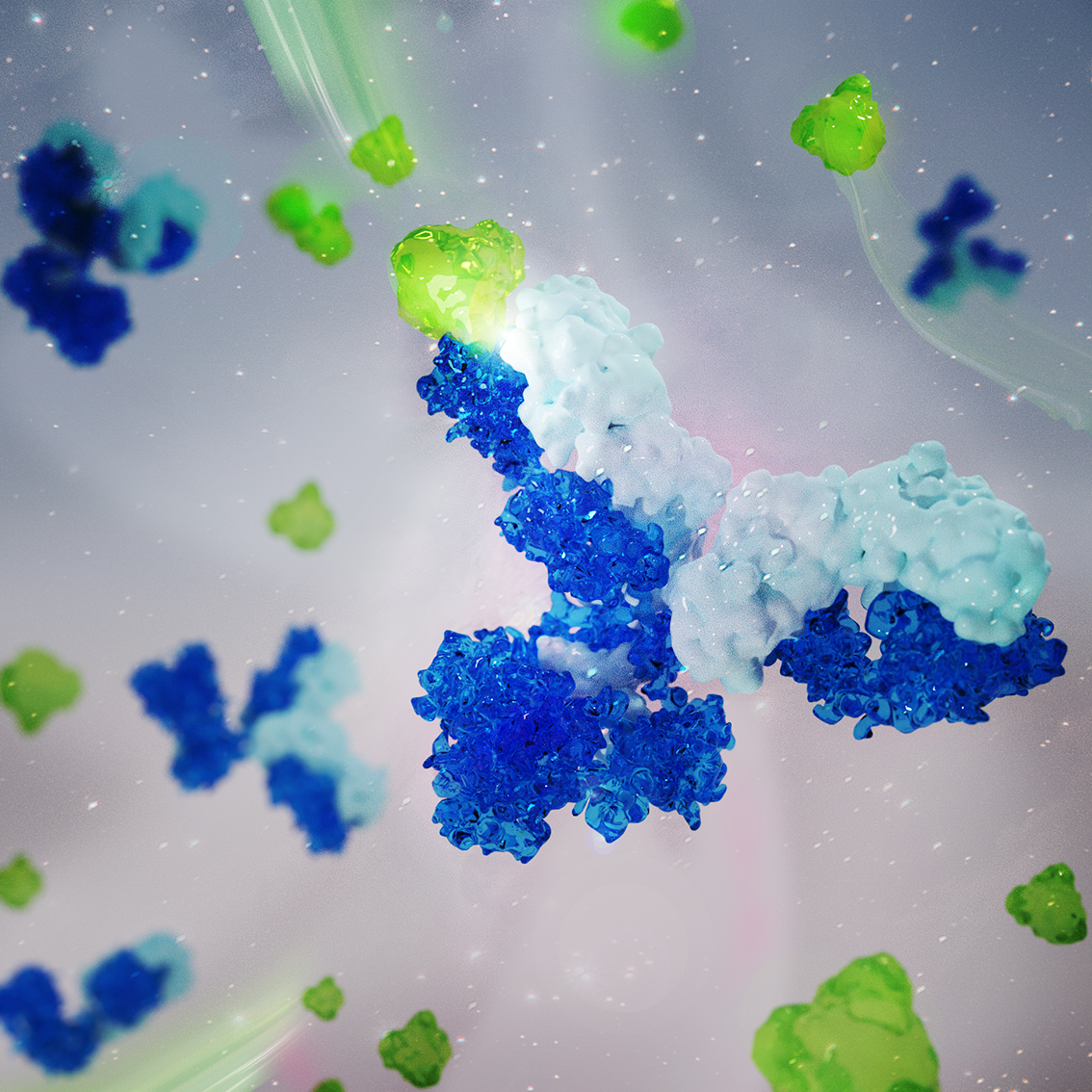
A metaphorical digital drawing of the physical forces that play a crucial role in shaping animal tissues and organs. This illustration shows the actomyosin filament (pink thread) that drives tissue fluidization during epithelial gap closure in embryogenesis or wound healing. Similarly, holes in old socks can be sewn together.3D visualisation of antibody molecules (heavy chains in dark blue and light chains in light blue) detecting antigens (green)
How do you make your art?
I usually draw an initial concept as a pencil sketch, creating different versions. I present these to my client and then convert the preferred version into digital form. The final digital version depends very much on the subject and what it will be used for. I usually create vector graphic illustrations for graphical summaries/abstracts, adding final digital/painting touches. I also create 3D visuals using 3D software, which are best suited for presentations, websites and cover art where you want to engage your audience.
Digital drawing of Thomas H. Morgan
Does your art influence your science at all, or are they separate worlds?
When I was doing my own research, it was probably a direct influence. I don’t know if you could call it art, but much like at university, I used drawings to summarise my findings and it helped me to put them into a bigger picture/see what direction to take next. It usually forced me to simplify and focus on the main points. Now it is more my relaxation/hobby time. I do enjoy the process of translating a key scientific message into a visual form. It is not classical art per se, but it does require certain ability to come up with an original idea that leads to a visually pleasing and self-explanatory design that attracts your audience.
3D visualization of a ‘springing’ neuron
What are you thinking of working on next?
I’m looking forward to the art and craft show at our institute, where I plan to show some of my art made with traditional painting materials. It is a hobby that forces me to leave my computer screen and gives me new ideas for digital art. I also hope to have more time to update my website and to launch more products incorporating my scientific designs.
Artworks were created using the Fluid Art technique to evoke the abstract representation of plant tissue. Both 40x40cm on deep edged canvas. These were created for the 2022 exhibition.
Dr Taiichi Tsuyama, Professor Tadashi Uemura and colleagues from Kyoto University recently published a paper in Development entitled ‘Dynamic de novo adipose tissue development during metamorphosis in Drosophila melanogaster‘, identifying the precursor cells that give rise to the adult fat body in Drosophila. We caught up with the authors to learn more about the story behind this work.
What were known about the origin and developmental processes of adult adipose tissue in fruit flies before your work? At the start of this project, many fly people knew that the adult fat body (AFB) exists in adult flies immediately after eclosion; however, only several studies tried to reveal the developmental aspects of the AFB in detail. Using transplantation techniques, Lawrence and Johnston (1986) reported that the AFB is mesodermal in embryonic origin. Hoshizaki et al. (1995) tackled the origin of the AFB using histochemical techniques with state-of-the-art genetic reporter lines in those days. The Hoshizaki paper has been one of the best references for the development of the AFB for about 30 years. However, it had been cited only ~30 times by 2021 despite many studies employing the mature AFB to study fat metabolism in adult flies. No previous study had identified precursor cells of AFB and characterized their cellular dynamics underlying AFB formation.
Why is this such a challenge to unravel? We think a major obstacle was the lack of genetic tools that specifically control gene expression in the AFB but not in the larval fat body (LFB). The larval fat body cells, which are generated in the embryo, persist during metamorphosis and locate near the AFB in young adult flies. Thus, genetic tools specific to the AFB are required to unravel the developmental progress of the AFB. Our interest in the adult fat body might be kind of serendipitous. One central theme in our laboratory has been how neuronal dendritic arbors achieve their complex and diverse morphological patterns and how they undergo remodeling during metamorphosis (for example, Shimono et al. 2014; Tsuyama et al. 2017). When we had attempted to study how systemic communications affect the metamorphic remodeling of dendritic trees in flies, we noticed that there were no good tools to control gene expression in larval and adult fat cells individually during metamorphosis. It prompted us to establish new genetic tools, which enabled us to visualize the developmental progression of the AFB in metamorphic flies.
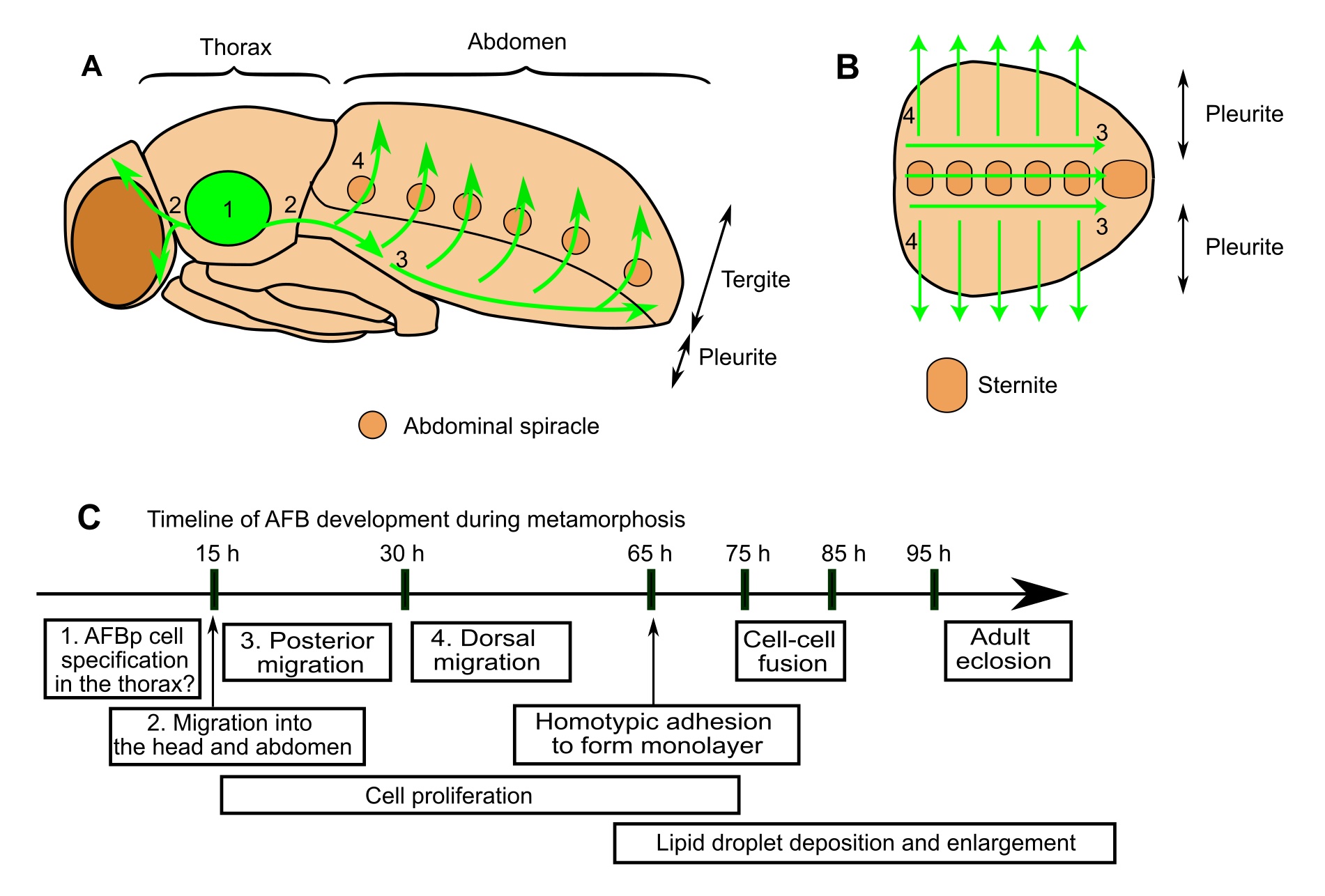
Can you summarise your key findings? We identified precursor cells that give rise to the AFB and delineated their dynamic cellular behaviors at the single-cell resolution (Figure 1; Tsuyama et al. 2023). These precursor cells emigrate from the thorax with polarized cell shapes and oriented motility, and undergo a long journey to disperse to the abdomen and head. After this spatiotemporal large-scale migration, these cells adhere to each other, assembling into the AFB with a sheet-like architecture. Cell proliferation takes place continuously during and after the migration to make up one of the largest tissues in the abdomen of adult flies. Another intriguing behavior is homotypic cell fusion after the sheet formation, resulting in the formation of multinucleated adult fat cells. We also tested the roles of candidate genes and found that Ecdysone Receptor (EcR), a steroid hormone receptor critical for the metamorphic progression of insects, and the GATA-factor transcription factor Serpent support AFB organogenesis.
Adult fat body precursor cells undergo a long journey to disperse across the whole body. Schematic illustrations of AFBp migration pathways in the fly (A,B). While our data support the notion that AFBp cells originate from thoracic segments, the detailed site of origin is still unclear. (C) Timeline of fly AFB development during metamorphosis (h: hours after puparium formation). Numbers in text boxes refer to numbers in A and B.
Are you surprised that the adult fat body precursor cells have to migrate over a long-distance from the thorax to disperse across the body? Yes. The migration-based distribution strategy makes a striking contrast with those of other mesodermal organs. Larval fat cells and muscle precursors differentiate in a segmentally-repeated manner in the embryo. Adult muscles, another class of mesodermal tissue that undergo metamorphic remodeling, are also locally generated during metamorphosis. Thus, such on-site differentiation is likely to be the canonical mechanism for the broad distribution of mesodermal organs in the fly; in contrast, the long journey of the AFB precursors appears to be exceptional.
When doing the research, did you have any particular result or eureka moment that has stuck with you? We experienced at least two eureka moments in our searches for Gal4 driver stocks that can induce gene expression in the AFB. After establishing new Gal80 lines, which block Gal4 activity in larval fat cells, we tested various known fat-body Gal4 drivers with our Gal80 lines and found that the c833-Gal4 driver could visualize migrating adult fat precursor cells. Then, in our additional search for Gal4 lines related to mesodermal genes, two svp Gal4 strains exhibited persistent Gal4 expression from early in the AFB lineage onward when used with lineage tracing tools. The moment when we saw the precursor cells migrating from the thorax into the abdomen was memorable indeed (Movie 2 in Tsuyama et al. 2023).
And what about the flipside: any moments of frustration or despair? With the new Gal4-based tools, we started lots of imaging with fluorescent protein markers and got a rough picture of the developmental progression of the AFB. Then, we examined whether wildtype flies without protein markers start to deposit lipid droplets at the same stage using Nile Red (a lipid stain); however, we were puzzled to find that various wildtype flies exhibited reduction or absence of Nile Red-positive lipid droplets with variable degrees of penetrance late in metamorphosis. We suspected that fluorescent markers might affect the developmental timings of the AFB but finally found that various fly stocks, including a widely used wildtype strain Canton-S, showed disorganized or lost AFB tissues with approximately 30% (!) penetrance even in the mature adult stage. We first could not come up with such an idea that a major tissue is occasionally lost in wildtype flies. This finding prompted us to examine defects in AFB development in a collection of inbred lines, Drosophila Genetic Reference Panel, performed by Yusaku Hayashi. Our results strongly suggest that the aberrant AFB development observed in those wildtype genetic backgrounds might be due to genetic variations, and we are attempting to characterize novel candidates of genes controlling AFB development as our ongoing study.
Where will this story take the lab? Tadashi Uemura: So far as a separate project, I have been studying long-term effects of nutritional environments during larval stages on the reproduction and lifespan of adult flies. The background of that project and a potential link with our study on AFB development are the following: massive and rapid growth of juveniles is heavily influenced by the quality and quantity of nutrients consumed. The impact of the nutritional environment in the early life (the nutrition history) is not restricted to that stage; the nutrition history exerts long-term effects on adult health in the later life. I have been investigating on underlying mechanisms including identification of the key cell where the history is stored, and the AFB precursor could be one of such candidate cells.
Taiichi, what brought to you join Prof. Uemura’s lab? And what is next for you after this paper? When I was a senior undergraduate student, I studied molecular mechanisms underlying plant-microbe symbiotic interactions using Lotus japonicus (supervised by Dr. Shingo Hata; I never imagined that studying gut-microbe and plant-microbe interactions is so trending now!). Reading papers with whole plants brought up my interest on molecular genetics. I was also interested in morphogenesis of cells. At that time, Uemura-san’s lab had been studying the molecular basis of dendritic morphogenesis using fly molecular genetics. So, I thought it might be the best place to join as a graduate student. After some studies on how aberrant ATP metabolism due to mitochondrial dysfunction causes the loss of dendritic trees of sensory neurons as my doctoral project (Tsuyama et al., 2017), we started this study on the development of the AFB. Even though these projects took a whole lot of time, Uemura-san generously and continuously supported my thoughts and plans. Recently, I have been focusing on ATP metabolism by developing genetic tools to manipulate energetic metabolism and structural analyses of proteins related to ATP metabolism in Prof. Ken Yokoyama’s laboratory at Kyoto Sangyo University.
References Lawrence, P. A. and Johnston, P. (1986). Observations on cell lineage of internal organs of Drosophila. Journal of embryology and experimental morphology 91, 251–66.
Hoshizaki, D. K., Lunz, R., Johnson, W. and Ghosh, M. (1995). Identification of fat-cell enhancer activity in Drosophila melanogaster using P-element enhancer traps. Genome 38, 497–506.
Shimono, K., Fujishima, K., Nomura, T., Ohashi, M., Usui, T., Kengaku, M., Toyoda, A. and Uemura, T. (2014). An evolutionarily conserved protein CHORD regulates scaling of dendritic arbors with body size. Scientific Reports 4, 4415.
Tsuyama, T., Tsubouchi, A., Usui, T., Imamura, H. and Uemura, T. (2017). Mitochondrial dysfunction induces dendritic loss via eIF2α phosphorylation. Journal of Cell Biology 216 (3), 815–834, jcb.201604065.
Tsuyama, T., Hayashi, Y., Komai, H., Shimono, K. and Uemura, T. (2023). Dynamic de novo adipose tissue development during metamorphosis in Drosophila melanogaster. Development 150 (10): dev200815.
Our June webinar will be chaired by Development’s Editor-in-Chief, James Briscoe (The Crick), and features three early-career researchers investigating growth and morphogenesis. The webinar will be held using Zoom with a Q&A session after each talk.
Wednesday 21 June 2023 – 16:00 BST
Patricio Pérez-Henríquez (University of California, Riverside) ‘PIN2-mediated self-organizing transient auxin flow contributes to auxin maxima at the tip of Arabidopsis cotyledons’
Stefania Tavano (MedUni Vienna) ‘You shall not pass! How ectoderm patterning modulates lateral mesendoderm migration in the early zebrafish gastrula’
Stefan Harmansa (IBDM and Turing Center for Living Systems) ‘Morphogenesis in a growing shell’
Doing great science depends on teamwork, whether this is within the lab or in collaboration with other labs. However, sometimes the resources that support our work can be overlooked. Our ‘Featured resource’ series aims to shine a light on these unsung heroes of the science world. In our latest article, we hear from Denis Bienroth, who describes the work of VR-Omics.
What is VR-Omics?
VR-Omics is a computational framework that analyses, visualises, explores, and interprets spatially resolved transcriptomics (SRT) data. It supports SRT data from various technologies, including sequencing-based and imaging-based platforms. Notably, VR-Omics is the first tool to analyse and visualise SRT data in both 2D desktop and virtual reality (VR) environments. It incorporates an automated workflow for data preprocessing and spatial mining. Additionally, VR-Omics is available at our VR-Omics Website. It is an open-source software, ensuring accessibility for all users. This platform is highly valuable for researchers as it also facilitates cross-platform comparisons, particularly when deploying different SRT technologies.
The VR-Omics Introduction Video showcases a comprehensive array of SRT Methods, providing an overview of all the options available.
What inspired VR-Omics’ development and its target challenges?
The development of VR-Omics was inspired by the realisation that existing tools for spatially resolved transcriptomics (SRT) data analysis required a high level of computational knowledge, making them inaccessible to many biologists. We aimed to create a solution that empowers researchers by providing a user-friendly graphical user interface (GUI) and eliminating the need for extensive computational expertise. VR-Omics puts the power back in the hands of biologists, allowing them to easily work with their own data and explore the fascinating world of gene expression landscapes.
Which SRT methods are currently supported?
VR-Omics currently supports a variety of spatially resolved transcriptomics (SRT) methods, including both sequencing-based and imaging-based technologies. The supported sequencing-based SRT methods include Visium from 10X Genomics, Tomo-seq, and STOmics from BGI. In terms of imaging-based SRT methods, VR-Omics supports Xenium from 10X Genomics and MERFISH from Vizgen. Additionally, VR-Omics allows for the analysis of custom SRT data, providing flexibility for users who are using their own spatial transcriptomics data not suitable with any of the aforementioned vendors. It’s worth mentioning that CosMx by Nanostring will also be available in VR-Omics shortly, further expanding the range of supported SRT methods. The wide range of supported SRT methods in VR-Omics ensures its compatibility with diverse experimental setups, enabling researchers to analyse and visualise their spatial data effectively.
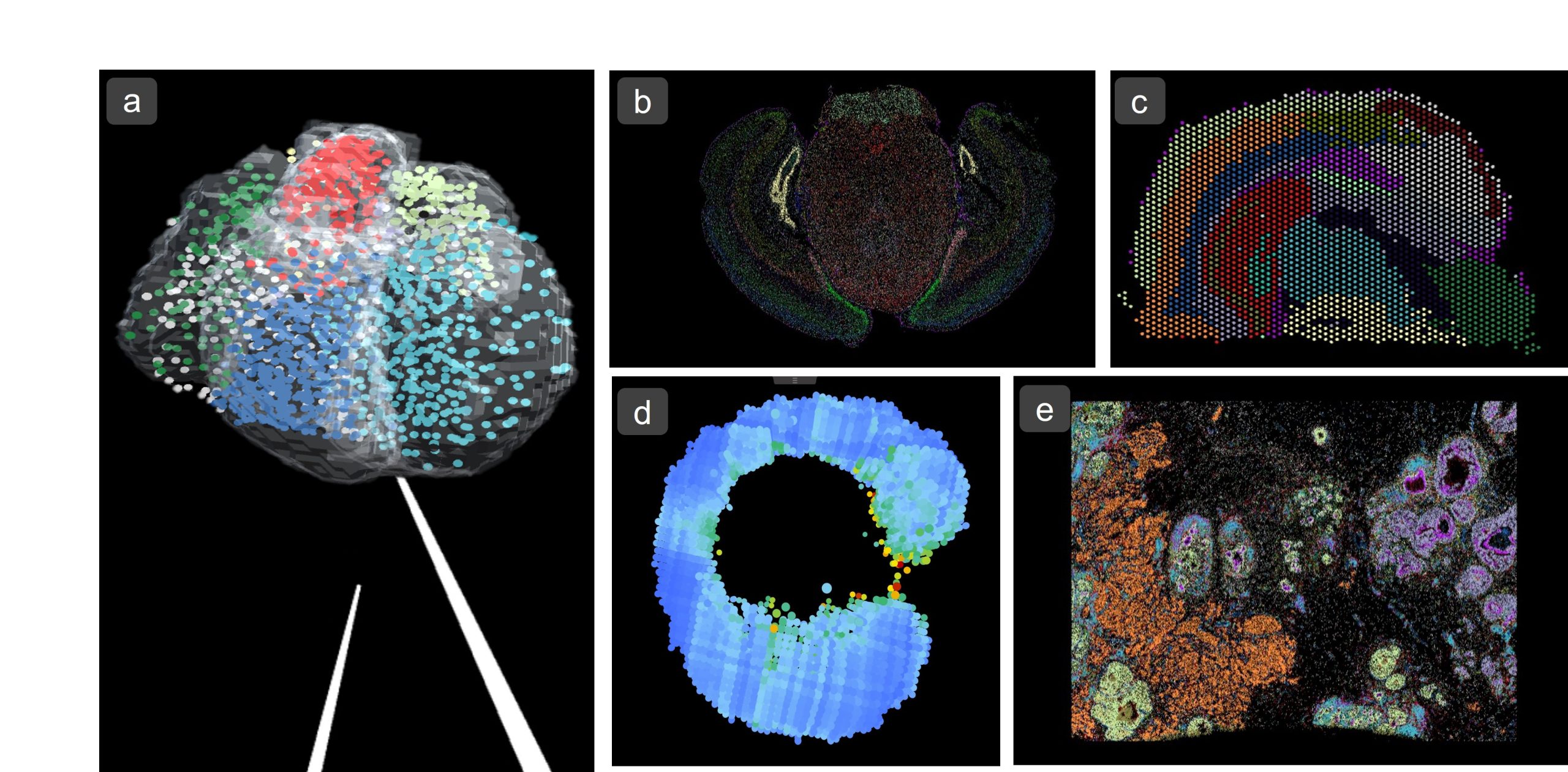
Figure 1: Visualisation of different Spatial Transcriptomics data sets using VR-Omics. a) 3D Dataset of Human Developing Heart1 (Visium – 10X Genomics) as VR environment. b) Mouse Brain Receptor Map2 (MERFISH – Vizgen) c) Mouse Brain Coronal Section3 (Visium – 10X Genomics) d) Zebrafish Embryo SS154 (Tomo-Seq) e) Breast Cancer Tumour Section5 (Xenium – 10X Genomics)
How can biologists’ access and utilise VR-Omics for their research?
VR-Omics empowers biologists to improve their research and make significant findings by providing an immersive and intuitive platform for spatially resolved transcriptomics (SRT) data analysis. Its visualisation capabilities enable a deeper understanding of gene expression landscapes, while cross-platform compatibility facilitates collaboration and data comparison. By streamlining workflows and eliminating computational complexities, VR-Omics allows researchers to focus on interpreting their data and uncovering valuable insights, ultimately accelerating scientific progress in diverse fields. VR-Omics is available at our VR-Omics Website.
Can I use VR-Omics with my own data?
Absolutely! VR-Omics is specifically designed to empower you to use your own data. It seamlessly supports spatially resolved transcriptomics (SRT) data generated through sequencing-based or imaging-based technologies, ensuring compatibility with multiple SRT platforms. Once loaded, VR-Omics streamlines the analysis process with its integrated workflow, automating tasks such as clustering, filtering, and spatial variable gene analysis. This means you can effortlessly explore and gain valuable insights from your data.
Can I export my findings to share them with my colleagues?
VR-Omics provides convenient export options for sharing your findings. You can export the output of the automated workflow, including plots, CSV files, and HDF files for downstream analysis. Additionally, the software allows you to export data directly from the Visualiser, such as gene lists, selected Regions of Interest (ROIs), screenshots, and videos. These versatile export capabilities enable you to easily share your results and collaborate effectively with your colleagues.
How can I explore my spatial data with VR-Omics?
With VR-Omics’ Visualiser feature, you can explore your spatial data on your desktop or in an immersive virtual reality (VR) environment. It supports side-by-side gene comparisons, merging gene expression patterns in one slide, and overlaying 3D objects for improved orientation. The Figure Viewer allows real-time interaction with output plots. VR-Omics offers a range of powerful tools to enhance your exploration and analysis of spatial data.
Creating 3D dataset from sequential Visium slides and exploration of the 3D data using VR-Omics.
What if I don’t know how to process my data? Do I need computational knowledge to run VR-Omics?
Not at all! VR-Omics is designed to be user-friendly and accessible, even for those without extensive computational knowledge. Its integrated workflow automates essential processing steps, allowing you to analyse your data without requiring deep computational expertise. With VR-Omics, you can easily load your data and navigate the intuitive graphical user interface (GUI) for seamless data processing and analysis. It empowers biologists to explore and interpret their spatial transcriptomics data effortlessly, regardless of their computational background.
Can VR-Omics facilitate the creation and exploration of 3D spatial data?
Certainly! VR-Omics is specifically designed to facilitate the creation and exploration of 3D spatial data. While many platforms are limited to visualising individual sections from a single spatially resolved transcriptomics (SRT) experiment, VR-Omics takes it a step further. It enables you to seamlessly navigate through multiple sections, such as serial sections of the same tissue, for a more comprehensive understanding of spatial gene expression patterns in three-dimensional space. With the immersive capabilities of virtual reality (VR), VR-Omics provides an enhanced environment to study complex biological systems and unravel intricate spatial relationships. It empowers researchers to delve into the three-dimensional landscape of their SRT data, gaining deeper insights into the biology of the system under study.
VR-Omics Features of the Visualiser to explore and mining of spatial data. Gene search (Heatmap and Binary); Side-By-Side comparison and merge function; ROI selection; Cluster visualisation and environment customisation on desktop or VR.
What are the next plans and features for VR-Omics?
The future of VR-Omics is driven by our commitment to constant improvement and expanding its capabilities. Our plans include incorporating new spatially resolved transcriptomics (SRT) methods, such as Nanostring (CosMx), to broaden the range of compatible technologies. We are dedicated to staying up to date with the latest algorithms and packages used in the literature, ensuring that the Automated Workflow remains robust and relevant. Additionally, we are excited to introduce new features, to enhance the user experience and analysis possibilities. Our goal is to continuously evolve VR-Omics, incorporating user feedback and advancements in the field, to provide a cutting-edge software solution for spatial data analysis.
What are the benefits of using VR for spatial data work, and can I use VR-Omics without VR?
VR-Omics offers the flexibility to work with or without VR hardware, allowing users to choose their preferred mode of interaction. While VR provides an immersive experience for spatial data exploration, the benefits go beyond visualisation. In a virtual reality environment, users can effectively navigate and analyse complex 3D spatial structures, revealing hidden patterns, identifying spatial co-expression relationships, and gaining new insights into biological phenomena. However, even without VR, VR-Omics provides powerful analysis and visualisation tools, ensuring that users can effectively analyse and interpret spatial data. Whether in VR or non-VR mode, VR-Omics empowers researchers to unlock the full potential of their spatial data.
What if a necessary feature is missing in VR-Omics Visualiser or the Automated Workflow that would benefit my project?
We highly value user feedback and are dedicated to continuously improving VR-Omics based on the needs of our users, particularly biologists working with spatial data. If you find that there is a missing feature or a step in the Automated Workflow that would benefit your project, we are always eager to hear your suggestions. We actively seek input from users and encourage collaboration to enhance the software’s functionality. By engaging with the community, we aim to incorporate new features and improvements that address the specific requirements of our users. Your input is invaluable, and we are committed to making VR-Omics a powerful and user-friendly tool for spatial data analysis.
What if I have any questions, is VR-Omics documented and supported?
If you have any questions or need assistance while using VR-Omics, you can rely on its Documentation and support resources. VR-Omics is well-documented, providing detailed guides, tutorials, and documentation that explain its features and functionality. Additionally, the VR-Omics team is available to help address any queries or issues you may encounter. They are committed to providing support and ensuring a smooth user experience with the software. Shot us an email (denis.bienroth@mcri.edu.au) if you have any questions or inquiries.
References
1: Asp, M. et al. A Spatiotemporal Organ-Wide Gene Expression and Cell Atlas of the Developing Human Heart. Cell179, (2019).
4: Junker, J. P. et al. Genome-wide RNA Tomography in the Zebrafish Embryo. Cell, 662–675 (2014).
5: High resolution mapping of the breast cancer tumour microenvironment using integrated single cell, spatial and in situ analysis of FFPE tissue, https://www.10xgenomics.com/products/xenium-in-situ/preview-dataset-human-breast
We are happy to announce the 2nd edition of the EMBO workshop “The Evolution of Animal Genomes”.
The event will take place in Seville (Spain), from 18-21 September 2023 (Registration and abstract deadline: 12th July 2023)
Genome evolution represents the basis of species adaptation to changing environments and habitats. Recent breakthroughs in sequencing technologies resulted in the acquisition of complete genome information for an increasing number of animal species, propelling the field of evolutionary genomics into a new era of discovery. Yet, our limited capacity to interpret genome variation hinders our understanding on how phenotypical changes drive adaptation. This urges the development of novel strategies to reconcile genomic sequence and function, for which a proper integration of cell-specific gene programs, non-coding regulation and 3D chromatin organization becomes essential. Further, establishing causal relationships between genome mutations and phenotypes still remains a major challenge in the field. Within this context, novel synthetic biology approaches are emerging as a means to understand developmental processes in the context of evolution. The aim of this EMBO workshop is to bring together international scientists with distinct, but complementary expertises on interpreting genome variation, on mechanisms of gene regulation and on in vivo synthetic biology approaches. This allows a comprehensive overview that goes from fundamental principles encoded in genomes to their ultimate biological significance on the formation of living, evolving organisms.
An exciting line-up of speakers (keynote lectures by Edith Heard, Mike Levine and Neil Shubin) will cover the following topics:
– Genomics of ecological adaptation – Evolution of cell types – Mechanisms of regulatory variation – 3D genome organization and structure – Synthetic biology approaches to model evolution
There will be short talks selected from abstracts, as well as ample time for networking.
Fee waiver, travel and childcare grants available.
A recent paper in Nature Cell Biology entitled ‘Co-option of epidermal cells enables touch sensing’ reports a new type of specialised epidermal cells involved in touch sensing in Drosophila. We caught up with Dr Federica Mangione, the first author of this paper, to find out more about the story behind the paper.
How did you get started on this project?
I joined the Tapon lab for my postdoc, aiming to study how cell differentiation impacts the structure and function of the somatosensory system, using my beloved Drosophila as an in vivo model. I initially wanted to understand how the tactile bristles, hair follicle-like structures decorating the adult epidermis, develop to allow the adult fruit fly to sense tactile stimuli. Given that the terminal differentiation of these touch-sensitive organs was largely unexplored, what I did first was to visualize this developmental stage, combining bristle-specific genetic labelling with live imaging. While analysing the cellular dynamics underlying the differentiation of the four lineage-related cell types that make up each bristle within the epidermis, I identified the epidermal F-Cell as a novel cell type in the assembly of the mature tactile organ. The temporal and spatial precision underlying the acquisition of the F-Cell fate led us to pursue a detailed study on the structure and function of the tactile bristle and its association with the F-Cell.
What was known about the role of epidermal cells in the function of touch-sensitive organs before your work?
The epidermis is the outermost layer of animals’ body, and it integrates various specialized cells and cellular structures that associate with sensory neurons to shape the sense of touch. In mammals, specialized cells of epidermal origin include the Merkel cells and a subset of the cells making up the hair follicles. While both Merkel cells and hair follicles associate with specific subsets of sensory neurons for touch sensing, the specific role of the epidermal cells of the hair follicles in touch sensation is not well understood. Our work shows that the tactile bristles, hair follicle-like structure in the Drosophila epidermis, associate with specialized epidermal cells, the F-Cell, to sense touch and reveal that the insect epidermis also contains specialized epidermal cells involved in sensory detection.
Can you summarise your key findings?
One key finding is that F-Cell fate specification occurs post-mitotically: selective elimination of the F-Cell through laser microsurgery induced de novo specification of the F-Cell fate in the remaining epidermal cell adjoining each tactile bristle. The precise dynamic of this event led us to perform a series of genetic and optical experiments which, together, indicated that the shaft cell of the bristle is orchestrating a short-range signalling to select F-Cell fate within the epidermis. Another key finding is that this short-range signalling is dependent on the conserved epidermal growth factor receptor (EGFR) signalling. While corroborating these findings, we asked what happens after F-Cell fate acquisition. Through temporal volume electron microscopy (vEM) and light microscopy during the terminal differentiation of the epidermis, we established that the F-Cell is the only epidermal cell that changes shape to wrap around the tactile bristle. The close association between the F-Cell and the tactile bristle suggested a functional requirement for this cell in touch sensing. Through in vivo electrophysiological recordings and behavioural assays, we found that the F-Cell is indeed essential for touch sensing. Altogether, our work established that the F-Cell is a specialized epidermal cell which shapes the functional assembly of the tactile bristles.
Morphological differentiation of the F-Cell. See Mangione et al., Nat Cell Biol, 2023
This interdisciplinary work involved diverse expertise, can you talk a bit about your experience collaborating with people in other fields?
Collaborating with people with different known-how and points of view has been incredibly beneficial, both professionally and personally. By working closely with Catherine Maclachlan in the team of Lucy Collinson (The Francis Crick Institute, London, UK), I have learned the many steps behind the beauty of an EM image and the power of vEM in gaining information on cellular morphologies in 3D. I am looking forward to discovering more about the cells of the tactile bristles through EM as I carry on working together with this great team of experts! Working with Joshua Titlow in the team of Ilan Davis (University of Oxford, UK) has been a fantastic experience too! He guided me through the complex process of generating a successful recording of neuronal activity and I am so grateful for his dedication and patience during my many visits to Oxford. This collaboration definitely stoked my passion for neuroscience, which will stay with me for the rest of my career. I feel fortunate to have met Michel Gho (Sorbonne University, Paris, FR) while I was characterizing the genetic basis of F-Cell fate specification. Collaborating with a leading expert in bristle development and genetics such as him has been an absolute honour and of great help to focus on EGFR as a key signal for F-Cell fate specification. All the support from our collaborators have been crucial to shape this paper the way it is!
Did you have any eureka moment that has stuck with you?
One in particular, indeed! While performing live imaging, I observed that, at some point during their differentiation, the cells expressing a bristle-specific cell marker changed from four to five. I could not detect any cell division within the bristle lineage or outside in the epidermis at that time, so how did this switch from four to five occurs? And why? This observation had me puzzled for a while! Three decades of studies established that each bristle is composed of four cell types, all derived from a single precursor cell, so why was I counting 5 instead? How is it that 2+2=5? The eureka moment arrived within the first 2 years of studies, when I found that specific manipulations of bristle cells were affecting the appearance of the F(ifth)-Cell. This told me that the F-Cell is co-opted by the tactile bristle form the neighbouring epidermis, and this makes the impossible possible: 2+2 really equals 5! The 2 pairs of sister cells in the bristle progenitor lineage require “a fifth element” for the assembly of a functional tactile organ. Eureka!
The Five Cells of the Differentiating Tactile Bristle. sh: sheath cell, n: neuron, Sf: Shaft cell, So: Socket cell, and the F-Cell. See Mangione et al., Nat Cell Biol, 2023
And the flipside: were there any moments of frustration or despair?
Finding a new cell type is very exciting! A flipside, however, is that gaining insights into what makes an uncharacterized cell type unique is quite challenging experimentally. For example, after first observing the F-Cell, I didn’t have any genetic tools to manipulate gene expression in a restricted manner for quite a long time. Also, many of the genetic manipulations I was initially testing affected the fate of bristle cells too, preventing me from cleanly disentangling bristle and F-Cell specification. These obstacles were successfully overcome by combining laser microsurgery with live imaging to target individual cells and in a temporally controlled manner (PMID: 36685184). Performing light and electron microscopy imaging was also technically challenging, especially at later stages of development, when the epidermis become stiffer to form the adult exoskeleton of the fruit fly. We dedicated a lot of effort in optimizing sample preparations and imaging set ups, which definitely worthwhile as we were able to gain access to these developmental stages too! The limited/intermittent access to the lab during the pandemic was also very frustrating of course. Support from the lab helped me in keeping a positive attitude and I saved ‘positive energy stores’ for optimizing experimental designs and close gaps in the project once I was back in the lab.
Why do you think the role of the F-Cell was not characterized before?
I have been asked this question many times, and yet I am not sure that there is a ‘right’ answer. The beauty of Drosophila bristles has attracted scientistic for three decades, as they are a powerful cellular context to address fundamental questions about cell fate determination and asymmetric cell divisions. Surprisingly, however, their differentiation dynamics (how the mature tactile organ is built from its constituent cells) was understudied. The reason behind this lack of knowledge remained mysterious to me until I started performing live imaging during bristle differentiation, which revealed various technical challenges. Thus, I would say that the F-Cell remained uncharacterized as accessing to late developmental stages is not straightforward. My love for microscopy and support from the lab and collaborators helped me in overcoming some of these limitations and gaining accessibility to these developmental stages.
What is next for you after this paper?
I discovered the F-Cell shortly after starting my postdoc in the Tapon lab and collecting the data for this paper has been a great journey. This journey is still going on! There are many aspects of the development of this cell and its association with the tactile bristle that I wish to explore further. I am now completing my postdoctoral training and I am very excited about the prospect of leading a research team in the future and gain more insight into the cell biology of touch!
If you are interested in learning more about the volume electron microscopy (vEM) technique used in this paper, check out the FocalPlane post ‘Inputs and Outputs of vEM in a Sensory System‘.
In October 2020, we started to observe haemorrhages in the human fetal brain tissue that we received from the HDBR. We were previously using the tissue from HDBR to investigate features of typical cortical development, but we quickly realised that we would need to change track and investigate why these tissue samples contained these haemorrhages, and if this was linked to the COVID-19 pandemic we were in the middle of.
Two and a half years after receiving the first haemorrhagic sample arrived in the lab, we published the paper in Brain (https://doi.org/10.1093/brain/awac372). Here, we (first author Marco Massimo and second author Carlotta Barelli) share the story behind the paper.
How did you get started on this project?
Marco Massimo: I always say that this project ‘happened’ to us, as it was totally unexpected. In our lab we work with human fetal brain tissue provided by the Human Developmental Biology Resource HDBR (https://www.hdbr.org). It was October 2020 when HDBR sent us the first haemorrhagic sample. Although haemorrhages in fetal cortex have been observed, they are extremely rare, and initially we were all surprised to see such an injured sample. Over the following months we kept receiving more and more samples presenting haemorrhages and it got to the point that we couldn’t do our experiments investigating typical cortex development anymore! As you all will remember, in October 2020, in the UK, we were in the middle of the SARS-CoV-2 pandemic. Covid cases per day were remarkably high and a vaccine was yet to be offered to the public. We had never received this kind of injured samples before the pandemic, and it is highly unlikely to observe such a high number of haemorrhagic samples in such a short amount of time. That’s why we thought that the highly unusual number of haemorrhagic samples might have something to do with the ongoing Covid19 pandemic. So, this is how we began this project, we hypothesised that those haemorrhages could be associated with SARS-CoV-2, and, with the support of HDBR, together with our collaborators in Trieste and Edinburgh, we started investigating this option.
Carlotta Barelli: I had already been working in the lab for a couple of months as an undergraduate internship student when we started receiving an abnormally high number of human fetal brain tissue samples displaying cortical haemorrhages. These samples could not be used to study normal brain development and in December 2020 we had received enough of these haemorrhagic samples to start analysing them. Due to the timing, we began investigating whether SARS-CoV-2 could be involved in the injuries observed in these fetal brain tissue samples.
Can you summarise your findings?
MM and CB: In our study we report SARS-CoV-2 infection in human fetal brain in association with haemorrhage, disrupted endothelial integrity and infiltration of immune cells in the developing cortex.
Cortical haemorrhages were linked to a reduction in blood vessel integrity and an increase in immune cell infiltration into the foetal brain. Our findings indicate that SARS-CoV-2 infection may affect the foetal brain during early gestation and highlight the need for further study of its impact on subsequent neurological development.
Video abstract below:
When doing the research, did you have any surprising results?
MM: The whole project was a surprise. I guess the biggest shock was when we first detected the SARS-CoV-2 spike protein in the choroid plexus and in the cortex of haemorrhagic samples. That was the proof that there was an association between SARS-CoV-2 infection and the haemorrhages that we had observed.
CB: It was interesting to see that the majority of the haemorrhagic samples were between 12-14 post conception weeks (pcw). The haemorrhages found in these younger samples (12-14 pcw) were more recent than the haemorrhages found in older samples (19-21 pcw). This suggests a critical window of development where viral infection could have more serious consequences on fetal brain health. Specifically, the integrity of the vasculature could be more severely affected at these younger stages (12-14 pcw) when the blood-brain barrier is still forming, making the brain more susceptible to neurovascular damage.
What were the challenges you faced when working on this project?
MM: To me the biggest challenge was keeping all the data organised. We processed 26 different samples (plus a similar number of aged-matched controls) which came with a unique 5-digit HDBR number that we had to replace with other codes in order to make our analysis blind. Hundreds of immunofluorescence stainings, for tens of biological markers, had to be imaged generating a huge amount of data that had to be properly saved, analysed and ultimately had to be linked back to all the samples examined.
Another challenge, given the nature of this research, was that before sharing our results we had to be completely sure that our data was correct. We had to be extra careful in analysing and evaluating our staining, using isotype control antibodies, and making sure our results were consistent with our collaborators’.
CB: Due to the limited knowledge on such a new topic, it was hard to investigate the link between SARS-CoV-2 infection and the high incidence of cortical haemorrhages. When we got started on the project, the only studies on the impact of SARS-CoV-2 infection on the brain had been carried out in cortical organoids, which lack vasculature as well as immune cells. We asked various labs with expertise in brain development and maternal infection, but nobody had ever seen such injuries in human fetal brain tissue samples. This was how we started collaborating with the Giacca, Miron and Williams labs whose contribution was key to progress in this project.
What impact will/should your results have on public health advice?
MM: This is an important finding, given that the Covid19 pandemic is still ongoing, and it could have an impact on public health advice. At the time it wasn’t clear if SARS-CoV-2 could be passed from mother to foetus and what the consequences of SARS-CoV-2 infection on foetal brain could be. Although, we still don’t know if these haemorrhages are the indirect result of an immune reaction from the mother or are a direct effect of the viral infection, we thought it was important to share our findings with both the scientific community and the general public, so that everyone was aware.
KL (Katie Long): As the mechanisms leading to the haemorrhages are not yet understood, we aren’t currently in a position to give public health advice, however our colleague Professor Lucilla Poston CBE, Professor of Maternal & Fetal Health at King’s College London, recommended “We know that severe viral infection may influence the fetal brain, but this important study is the first to suggest that this may occur in pregnancies affected by COVID infection. Whatever the cause, a direct effect of the virus or an indirect consequence of maternal infection, this study highlights the need for pregnant women to be vaccinated against COVID-19, thus avoiding complications for both mother and baby.”
CB: Our results should raise awareness on maternal viral infection during pregnancy and encourage pregnant women to take vaccination against SARS-CoV-2. Our study also highlights the need to better understand the risks associated with maternal SARS-CoV-2 infection and its impact on later neurological development.
Where will this story take thelab, and more broadly research in this area?
KL: We are still receiving fetal brain samples with haemorrhages, although the incidence of this has slowed significantly after pregnant women were offered the COVID-19 vaccines. We will continue to try to understand why these haemorrhages occur and what impact they might have on the developing brain.
What next for you after this paper?
MM: This was my very first paper and I am glad to have contributed to this important finding. I learnt a lot from this, and I am proud of the work we all did. As a 2nd year PhD student my main focus now is to investigate neuronal migration disorders in the human fetal brain which is what my project was originally about. Even though I won’t have time to continue this research, as I have to prioritise my PhD project, I will be happy to help and support whoever will keep working on this.
CB: I have now left the lab and started a PhD focusing on glioblastoma and neural stem cells. Despite the shift in research topics, working on this project has taught me a lot about academic research. Specifically, it showed me how scientific research is often nonlinear, results are sometimes unexpected, publishing can be quite complex but, ultimately, the whole process is extremely rewarding.
The fifth episode of Made the Same Way, a podcast produced by the Wellcome-funded Human Developmental Biology Initiative, features Lucía Cabriales Torrijos discussing human lung development with performer Beth (aka BABYFLXKCO).
At the end of the episode, the pair collaborate on an original piece of music inspired by their conversation.
It’s amazing how there are options if someone is born premature.
-Beth
About the participants
Lucía is a biophysicist who loves to understand how the cells that make up our bodies perceive and respond to mechanical forces. For the last couple of years, her research has focussed on mimicking foetal breathing movements to understand how these affect cell functions (differentiation) in the formation of the lung. Apart from science, Lucía is interested in art and loves going to museums, the theatre and the cinema.
Beth, aka Babyflxcko, is a 23 year old female singer, songwriter and producer creating soulful jazz, R&B instrumentals and soulful melodies. Brought up in a musical family, she always had a passion for music, writing her first song at just 12 years old. But it wasn’t until she hit 19, living alone in London that she decided to pursue and create her own music full-time.
She released her first EP, Emotions, in 2021 after working alongside other musicians developing musical skills on a project with Reform Radio called Soundcamp 2021. Her musical influences include Jhene Aiko, Billie Eilish and Amy Whinehouse, just to name a few. She is known for doing shows in Manchester, Sheffield and local areas.
Beth has also recently released a new single, Inhalando Y Exhalando (the original music that she created as part of this podcast) on all streaming platforms. You can listen and download here.
Please subscribe and listen to Made the Same Way on Apple podcasts, Spotify, or wherever you get your podcasts. If you enjoy the podcast, please rate and review us on Apple podcasts to help others find us!
Calling all developmental biology postdocs and graduate students!
Interested in attending an exciting meeting? Want to expand your dev bio network? Want to present your work to a supportive audience?
Come to the 2023 Developmental Biology Gordon Research Conference and Seminar
Date: June 25 – 30
Venue: Mount Holyoke College, Mass, USA
Abstract deadline May 21, 2023
The organisers have raised significant funds to support registration fees and travel grants for postdocs and graduate students. If you need financial assistance please contact the organisers.

 (3 votes)
(3 votes)





 (No Ratings Yet)
(No Ratings Yet)