We pick up where we left off with Anna-Lena Vigil, now a PhD candidate in the Crocker Group at EMBL, as her journey unfolds to moments beyond the bench that shaped her path. In part one, we traced how questions of metabolism, cell fate, and adaptation shaped her scientific path — from studying cancer cells in a dish to exploring how metabolic shifts guide development in flies. But Anna’s story is also one of personal growth, bold moves, and a deep curiosity which make her among the few who took the leap from US to pursue grad school in Europe.
Anna’s scientific appetite spans fields and organisms — from plants to hibernating mammals to cancer cells — always chasing the big questions. As a technician in Lucas Sullivan’s lab, she honed both her skills and her curiosity, studying how metabolism rewires under stress. That experience sparked her shift to Drosophila, where she now explores how metabolism shapes development and adaptation. Whether in cell lines or whole organisms, Anna bridges disciplines, using metabolism as a language to make sense of biology’s complexity. Check out all her work here.
In this next part of our conversation, she reflects on what kept her in the field, how she navigated her transition into graduate school, and what advice she has for others at the crossroads of research and curiosity. In both parts of her interview she credits the infectious scientific culture in the Sullivan Lab, the value of mentorship, the thrill of scientific independence, and how metabolism remains both a personal fascination and a powerful lens for asking big biological questions. Along the way, she shares lessons in resilience, advice for young scientists, and the joy of embracing life (and science) with curiosity and a sense of adventure.
What kept you interested in continuing to pursue the field of metabolism?
To me, the biochemical processes that make a cell, a cell and an organism, an organism, is the most fascinating lens to view biology. The best thing about metabolic research is that it is the foundation to almost anything that biology focuses on: any question you ask or area you pursue, there is a metabolic system/process that is intertwined with it. These biological systems are deeply connected, and metabolism provides a more complex layer into how systems operate. I think what propelled me to stay in this field was not only the amazing work that has been done thus far, but all the work there is still yet to do and all the questions that still need to be pursued.
Tell us about what experiences/results/training from your time in the Sullivan Lab at Fred Hutch motivated you to continue pushing forward in grad school ?
As a research technician, I think the main motivation that drove me to pursue grad school was the unexplored opportunities for growth that come associated with scientific research. There is always room for improvement in science, so for me, pushing through to receive further training was something I wanted to experience, and I felt like I wanted to keep pushing myself further to reach my higher academic potential. As a technician, I learned a lot of independence with techniques and experiments, but I always felt as though I could go further intellectually and push myself to ask the important questions and decide on the right next steps. I find myself growing as a scientist everyday, and this is the exact reason I wanted to continue onto grad school.
Could you briefly share your experience transitioning from Fred Hutch in the U.S. to EMBL in Heidelberg? How did you apply, and what was the process of getting into graduate school?
Apart from wanting to try out life in Europe for a while, I was really interested in diversifying my research experiences and learning how research questions were approached in different institutes and countries. I knew that I absolutely want to be a biologist/scientist, so the goal of obtaining a PhD was something that I found necessary for my future plans of pursuing a scientific career. Also, learning of the metamorphosis that occurs as young scientists go through graduate school was something that I wanted to experience for my own personal development; even with all the ups and downs that come associated with it.
As the requirements for entering a PhD program were a bit different in the U.S. versus in Europe, I wanted to find a place where I could go directly into a PhD program without needing a Master’s degree (in the U.S. you can enter directly into a graduate program from your Bachelor’s studies, provided you have sufficient research knowledge and experience to help make the transition easier; in Europe, a Master’s degree is usually required). I felt my time as a technician helped reinforce my research independence, so I felt like obtaining a Master’s degree was an unnecessary step for me. So when I found the PhD program at EMBL I immediately became interested in pursuing an application. Not only was EMBL a leader in scientific research in Europe, it also had really cool research groups (like Justin’s group!) which made me very interested in pursuing a PhD in Heidelberg, Germany.
What were the cultural changes, how did you adapt? what were the biggest challenges/setbacks – how did you overcome them?
I have always been used to German culture growing up, as my mom is German by birth. That being said, there were definitely cultural changes I had to adjust to. I grew up and studied in Las Vegas before moving to Seattle, so needless to say most Europeans are far different than anyone from Vegas. One challenge was navigating how serious people can be, especially in some research settings. However, I am a true believer in the notion that we should not take life, and definitely not science, so seriously and instead focus on the joy that it brings and the gratitude we should all feel in being scientists. I have done many jobs in my life, and this one by far is the most fun and rewarding (and of course challenging). So I try to bring this mindset to every interaction I have here, either in science or in society, and it usually leads to a smile, even from very serious people. And for me, that smile makes cultural barriers break down and reveal the shared humanity we all have, as happiness and joy is universal.
What were the positive changes/surprises?
Generally, I found the move to Germany from the U.S. to be very refreshing. The way of life in Europe is a little more relaxed, so it has been a pleasant experience so far. Scientifically, I think it was also helpful to have a variety of different research experiences before coming here to help me navigate the landscape of such a different scientific environment, and different ways of thinking and approaching scientific questions. That being said, I work with amazing people and they have been integral to adjusting to a new place, scientifically and culturally. Having good people around can make all the difference. Although it has been a tough challenge to be far away from family and friends back in the States, I found the move to be generally more positive and rewarding than anything.
What advice would you offer to undergrads/postbacs interested in exploring the intersections of nutrition/metabolism and cell fate decisions?
My advice would be to start in a lab that specializes in metabolism to learn more mechanistic details of metabolic systems and pathways, and then move into more broader-themed labs. I think the trajectory of starting small (molecular) and zooming out (to disease, development, or evolution) gives you a better understanding of the molecular underpinnings of cell fate decisions and evolutionary trajectories. Also, apply to as many summer internships as you can while in undergrad; they provide really amazing research experiences and will give you a head-start when it is time to continue on to graduate school. Lastly, another piece of advice is to never ever give up! You are capable of many great things and you owe it to yourself to find out what you can accomplish in this world. Follow your curiosities and enjoy the journey, wherever it takes you!
Were there any pivotal moments that shaped your career path?
The most life-changing event that happened for me academically was a summer internship that I participated in hosted by the University of Washington doing single-cell genomics in Arabidopsis. It not only gave me hands-on experience working in a state-of-the-art lab, but also opened many doors for me professionally. I think I would not be where I am today if I didn’t get a chance to have that summer research experience and I am so grateful to have had that opportunity.
What role does curiosity play in your life, both within and outside of science? What motivates you to be a basic science researcher?
Curiosity is the fuel that keeps all scientists going! Curiosity is crucial to make it through difficult periods that are inherent to science, and I am grateful to have new curiosities to pursue everyday. I think basic science research is the diesel that fuels our societal understanding of the natural world around us. I find it a great honor and privilege to be able to contribute my career to help broaden our collective understanding of how nature works. Knowing that something I observe in the lab may be the first time it has ever been observed is literally the most exciting thing you could experience. To me, there is no better way to spend the day! I believe that basic science questions lay the foundation for the rest of the scientific community to build upon, and aid in advancing our collective health as a society. Without basic science research, the realm of health advances that we have achieved would eventually collapse. Understanding basic principles of how biology occurs in a general sense is the only way we can progress in the fields of health sciences as well, as many basic science and clinical questions overlap. Moreover, I am a huge plant lover and have many plants in my house. I find having plants in my home and seeing all the crazy developmental stages and cell-type changes that occur when you propagate them brings me back to why I find biology so cool!
How do you maintain a balance between your rigorous research activities and personal life? Are there hobbies or practices you find particularly rejuvenating?
I really enjoy gardening and animals. My goal in the future is to have my own (small scale) farm with a few crops and of course a nice collection of chickens for eggs, sheep for wool, goats for milk, and dogs because they are a human’s best friend. So in my spare time I like to learn about best gardening and farming practices for the future, while tending to my house plants in the meantime. Alternatively, I just enjoy being outside and find time in nature to be very rejuvenating.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
If I didn’t pursue biology as a career, I probably would have pursued a career in anthropology or sociology. I like people, communities, and the wisdom that different cultures can provide, so I was very interested in studying people and the customs they have built from generational traditions.
Previously we learnt about the role of metabolism in developmental patterning and embryogenesis. Check out – Metabolic Origins: Steering of early developmental fatefeaturing Kristina Stapornwongkul. Krisitina will be starting her lab at her own lab at IMBA, Vienna and will be hiring soon. Check out her lab page here!
Today marks the end of the chapter for my time at the Node and Development. What a ride it has been!
I don’t think you can find many jobs like this one: on the same day I could be writing a Research Highlight about a Development paper on morphogenesis, editing drafts written by researchers, interviewing someone about their career path and organising an online workshop with Reinier and Helen, the amazing team of community managers at preLights and FocalPlane.
I sometimes still can’t believe that I could be paid to do all these fun things. I also get to work alongside a group of brilliant people at The Company of Biologists, who had to endure my endless questions, my slight obsession with lists and spreadsheets, and my annoying complaints about the state of social media. I’ll really miss reading about all-things development biology (and beyond), working with everyone from the Company and interacting with the wider research community.
Here’s non-exhaustive list of my time at the Node and Development:
Research Highlights written: 59
Interviews conducted with people across the world at different career stages, with all kinds of career paths: 33
Hours spent on social media: too many…
Edits and feedback on drafts provided: I started counting but quickly lost track. My guess is it’s >120
Online webinars/workshops delivered: 10
Conferences attended: 8
Strava segment efforts on my cycle commute (specifically, the ‘Cambs north to Histon’ segment): 276
This axolotl was already waiting for me on the desk on my first day. I’ve named it Axie. No one knows how it got here and how old it is…
A highlight for me while at the Node is that I got to be around when The Company of Biologists turns 100 (and the Node turns 15). The Biologists @ 100 conference was hectic, but so much fun. I also enjoyed looking back at the origin story of the Node, and chatting to all the previous Community Managers.
Another highlight was reviving the Node’s career series. More than a decade ago, the Node featured a series of posts written by scientists who had gone on to do non-academic things, i.e. ‘alternative careers’, a term I want the Node to move away from. I decided to find people who have unconventional career paths – hence the name of the interview series: No such thing as a standard career path. I had the pleasure of talking to a group of people who have very different journeys to get to where they are now – all they have in common is that they are working, or have once worked, in development biology or related fields. It was a bit nerve-wrecking cold emailing people I’ve found on the internet. But every single conversation turned out to be really fun and thought-provoking. I hope the series has managed to demonstrate that many valid career paths exist.
The Node could only have survived for 15 years all thanks to every one of you in the community — whether you’re a quiet reader or an active writer for the Node — thank you for your support for the Node and The Company of Biologists! I hope to see the Company continue to support and inspire biologists across the world for years to come. I’d like to encourage you to:
Submit your manuscripts to Development (or Journal of Cell Science, Disease Models & Mechanisms, Journal of Experimental Biology and Biology Open if appropriate)
And of course, please consider posting on the Node – we welcome all kinds of posts, from a job advert to an event listing, from a behind the paper story to an ‘honest conversation’ about a topic that you’re passionate about but feel like it’s not well-discussed. Remember, you’re free to post without requiring approval. But the Node team is always happy to help at any point of the writing process.
I’m signing off from the Node now (although I’ve scheduled a few more posts for the quieter summer months, so you’ll see the ‘ghost’ of me posting). I’m off to continue my own ‘squiggly’ career journey – my next adventure is doing science communication in the biotech industry, an unfamiliar, but exciting world for me to explore. If you want to follow along, you can find me on LinkedIn.
The Node is 15 years old this year! We thought it’d be fun to get in touch with prolific authors of the Node over the past 15 years, revisit the posts they wrote for us and see what they’re up to now.
In this post, we caught up with Joachim Goedhart, whose “Data Visualization with Flying Colors” post remains the most viewed on the Node. You can check out all his other posts, mostly about data visualization and statistics.
What were you doing when you first started writing for the Node?
I wrote my first blog in 2017 and at that time I was an assistant professor at Molecular Cytology (part of the Swammerdam Institute for Life Sciences at the University of Amsterdam, The Netherlands).
I was – and I still am – developing fluorescent protein-based tools to study cellular processes. We are engineering fluorescent proteins and new biosensors based on fluorescent proteins. Since we have a strong focus on quantitative imaging technologies, the image processing, data analysis, and data visualization are important parts of our activities. Therefore, I also have a strong interest in data visualization and statistical analyses.
What motivated you to write for the Node?
Around 2017 I got interested in using R and ggplot2 for data analysis and making graphs and plots. This was totally new for me, so I was learning how to use this software and exploring its potential for data visualization. So, I thought that it would perhaps be helpful for others to share what I learned about R/ggplot2. I was looking for an online platform to post blogs and it turned out that the Node has a very nice infrastructure where it is super easy to write and post pieces. In addition, the audience of the Node aligns well with the audience that (I think) could be interested in the stuff that I write about. At first it was a bit scary that there is no moderation. At the same time this also provides freedom and gives the writer full control over the process which is cool. It took me a couple of posts to become comfortable with this situation, and I really appreciate that the Node provides this platform.
Choose a favourite/most memorable piece you’ve written for us and tell us why you’ve picked it.
It’s difficult to choose one, so I will choose two, a favourite and a memorable piece. The most memorable is a piece about “p-value parroting” as I called it. It’s the practice of mindless repeating what others do, and examples for statistical analysis – especially calculating p-values – are abundantly present in the scientific literature. To share my annoyance, I wrote a small correspondence that was published in Nature, but in the publication process the title was redacted. So, the piece on the Node was also meant to be able to share the original title that I had in mind.
My favourite piece is on “Data Visualization with Flying Colors” which discusses the use of colorblind friendly colors. This is something close to my heart as I’m red-colorblind, and it also seems to resonate with the community as it is often re-shared on social media.
Where are you now? What are you currently working on?
I’m still where I was in 2017 and I’m still roughly working on the same topics, although there’s a stronger focus on biosensors for quantitative imaging with fluorescence lifetime imaging microscopy. In general, it’s becoming easier to acquire multidimensional data (multiple timepoints, colors, stage-positions) with microscopes by automating the acquisition. Therefore, the datasets have become larger and there’s a lot of effort needed for processing, analysis and visualization of the data. It’s exciting to see that also the newest generation of students that we educate have an interest in data analysis and are motivated to learn coding and spending time on this important aspect of research.
Do you have any writing advice?
Write about something that you care about. I think that even if a blog is read by only a few people, it can be valuable (and it will actually be incredibly hard to write something that no one cares about). When I started posting pieces, I was also active on twitter, which helped a lot to find out what other people are interested in, and it was also a great platform to advertise the blogs and get reactions. Currently I’m using Bluesky to interact with people and share blogs, but I guess that other social media platforms can also be used for that.
What have you been reading/listening to lately? Any book or podcast recommendations?
I only recently discovered the Night Science podcast by Itai Yanai and Martin Lercher and I really enjoyed it. There are currently over 70 episodes and I only listened to a couple so there is still plenty to explore. As for books, I recently read the book “I can’t stop thinking about VAR” by Daisy Christodoulou. It discusses the introduction of the video assisted referee as a technology to support referees in soccer with their decisions. It is a nice example where a technology can solve some problems, but at the cost of creating new problems. In addition, it explains how difficult it is to make a binary decision (yellow card, offside), for a continuum of situations. These issues are also relevant for the scientific process and therefore I enjoyed the book a lot.
This week we’ll meet Anna-Lena Vigil, who is a PhD candidate in the Crocker Group, EMBL. From her postbac days in Seattle to her graduate research in Heidelberg, metabolism has remained her throughline: a dynamic system that powers cells, guides their fate, and adapts across contexts, from cancer to development. What drives her is the vastness of unanswered questions — the sense that even well-mapped pathways hold surprises when viewed through the lens of adaptation, evolution, or cell identity. Her first spark came in a college biochemistry class, where she realized metabolism wasn’t just “organic chemistry with a purpose,” but a living system at the heart of biology. That curiosity led her from studying gene regulation in plants to investigating how metabolism drives cancer cell proliferation at the Fred Hutchinson Cancer Center, Seattle. Along the way, she discovered how mentorship and the freedom to explore questions could transform a research career. Now, as a graduate student at EMBL in Heidelberg, Anna explores how metabolic signals guide cell fate decisions during development — and how these processes can adapt and evolve. In this first part of our conversation, Anna reflects on her scientific journey, the questions that drew her in, and why metabolism remains her lens for exploring life’s complexity using Drosophila as a model. In this first part of our conversation, Anna reflects on her scientific journey, the questions that drew her in, and why metabolism remains her lens for exploring life’s complexity. Check out all her work here.
Anna’s interview is a two part conversation, while you’re currently reading part 1 where she discusses her scientific journey, do check out the part 2 Between Molecules and Milestones, where she describes in detail – her journey to grad school, her continued interests in the field of metabolism, why she values curiosity and mentorship, and what keeps her motivated to continue moving forward.
What was your first introduction to the field of metabolism – what’s your first memory?
Outside of learning that the mitochondria is the “pOwErHoUsE” of the cell in high school biology class, my first introduction to metabolism started in my first biochemistry class at the University of Nevada, Las Vegas during my bachelor studies. My professor jokingly described the subject material of his class one day as: “organic chemistry but with a purpose.” During my time in this class, I was fascinated to learn how biological systems have figured out these extremely intricate ways of sustaining life through various different metabolic processes. It was after this class that I knew I wanted to learn more about how biochemical processes that make up metabolism can be the driving force of life on Earth.
Could you share your journey into studying metabolism and what inspired you to specialize in metabolic studies using two incredibly unique systems – mammals/cell lines and flies?
After my undergraduate studies, I was really interested in diversifying my research experiences in the early stages of my career. After studying gene regulation and stress tolerance in plants, and hibernation in a peculiar hibernator during my undergraduate research journey, I was really interested in understanding how metabolism can shape phenotypes, and how it may provide insight to help broaden our knowledge of disease mechanisms. So, with this in mind, and of course a stroke of good luck, I joined Lucas Sullivan’s lab as a research technician at the Fred Hutch Cancer Center in Seattle, where the lab’s main area of focus was understanding metabolic determinants of cancer cell proliferation. It was there where I learned of mechanistic details of how metabolism occurs in real systems, and how it can go wrong to lead to disease phenotypes, such as cancer. I think my drive for understanding metabolic systems really flourished in Lucas’s lab, as he was a great mentor and allowed me the resources and flexibility to pursue my own interests and curiosity. His passion for metabolism was infectious and he had a special way of promoting this same passion within his own lab members. Having a good mentor can really determine the rest of your research career! And after gaining a more informed mechanistic understanding about cancer metabolism, I wanted to learn more about physiological metabolic programs throughout development, so naturally using Drosophila as a model to learn more about this was a good way to transition from cancer metabolism to developmental metabolism.
Tell us about your undergrad/postbac work – particularly about the role of mitochondrial redox adaptations in regulating cellular fitness in the context of both normal and tumor cells.
The work on mitochondrial redox adaptations was an amazing project that was led by Dr. Madeleine Hart, a very talented graduate student in the Sullivan Lab at the time. She was primarily interested in understanding how certain subtypes of cancers, in particular, succinate dehydrogenase (SDH)-deficient cancers, were able to sustain intracellular aspartate when a major mitochondrial protein responsible for generating aspartate, was defective. SDH is also known for its role as complex II in the electron transport chain (ETC) in the mitochondria, and is responsible for the oxidation of succinate to fumarate. Fumarate is then later converted into malate and finally oxaloacetate, which is used to generate aspartate, a key amino acid that is required for cell proliferation. So, understanding how cancers with abnormal ETC activity sustained intracellular aspartate levels gave us an opportunity to learn more about the basic biology driving these types of cancers. In a glimpse into Madeleine’s work, she discovered that SDH-deficient cells were able to adapt to limiting environments by also adapting to downregulate the activity of complex I in the ETC. This downregulation resulted in restoring the NAD+/NADH balance to support further cancer cell proliferation. While we did not investigate this phenomenon in normal cells, the aspects we learned about how cell proliferation can be sustained in these scenarios helped provide insights into the broader theme of cellular adaptation to varying environments, in both normal and disease contexts.
The project I primarily worked on during my time as a technician included discovering novel metabolic fates in NRF2-activated cancers. As we all know that key metabolites and the components of metabolic pathways of cells have been known for decades, there are likely still metabolites, and entire metabolic pathways that are yet to be discovered. This is especially important when we think about how alterations in metabolism can lead to various human diseases. So to tackle the goal of trying to identify novel metabolites, we were interested in finding a system in which unknown metabolites likely existed, and so we decided to turn our attention to NRF2-activated cancers. NRF2 is a main transcription factor that regulates the production of various different antioxidants and detoxification programs. In the process of upregulating these detox mechanisms, one key target gene of NRF2 includes the xCT/SLC7a11 antiporter system, that is responsible for the uptake of extracellular cystine, and the excretion of glutamate. So with this in mind, and with the help of a clever isotope tracing method, we were able to uncover and validate ~9 novel metabolic fates, some of which can be detected in tumors. Also in this work, we learned very interesting aspects of intracellular cystine/cysteine that may help broaden our knowledge of cell proliferation mechanisms and metabolic phenotypes in cancer. I was very grateful to have been able to work on this project with such an amazing team, and hopefully you can read all about it soon!
Tell us how you got interested in cancer/immune metabolism for your post-bac work and how did you transition into Drosophila to study metabolic control of cell fate decisions?
For me, many aspects and fields of scientific research are extremely fascinating, but I decided to pursue metabolic research because I felt as though it viewed biology at the most molecular level in order to understand how life occurs. Many foundational concepts of metabolic programs that we know today were discovered using cancer model systems and in the context of cancer biology, so I felt as though joining a lab that studied cancer metabolism was a great way to enter the field. Although I greatly enjoyed my time working on how metabolism can be rewired to drive diseases, I wanted to learn about metabolic systems more from a normal physiological perspective, for example, in the context of developmental programs. In other words, I wanted to change from understanding how metabolism can go wrong (for a patient) and lead to disease, to learning about all the ways that metabolism can go right and facilitate the development of a whole organism from one single cell.
Metabolism is generally viewed as the sum of biochemical reactions that occur within cells and organisms to provide energy in the form of ATP, with the production of anabolic precursors and maintenance of NAD(P)+/NAD(P)H co-factor pools. A less appreciated view of metabolism is that metabolites themselves can act as signaling molecules to facilitate the up- or downregulation of other cellular processes. As certain cell types become more differentiated, so do their biological roles and needs, which at the root is driven by the utilization of alternative metabolic pathways. As someone who appreciates just how nuanced and complex biology can be, it was reassuring to learn that metabolism is also something that is extremely multifaceted, with new metabolites, roles of metabolites, and metabolic systems that are emerging everyday.
Tell us about your current work and how are you using flies as a model to study physiological consequences of metabolic signaling and its impacts cellular status and development?
My current work is about understanding how mutations in the genome can lead to altered metabolic states, and how these altered metabolic states can be inherited through generations. Mutations are thought to be the driving force of evolution; as mutations in coding regions are thought to affect mature RNAs or protein, noncoding mutations, or cis-regulatory mutations, are thought to affect the levels of transcription, acting as a knob to fine-tune expression programs throughout development. For my PhD work, I am interested in understanding how these mutations can lead to altered metabolic phenotypes, and in particular, how metabolism can be rewired to accommodate adaptive phenotypes, and how these adaptations can persist and lead to evolutionary novelties. These broad questions are especially important when considering natural Drosophila populations that are exposed to various different agrochemicals that are used in modern-day agricultural practices. I am interested in understanding how mutations in regions that are associated with detoxification programs affect adaptive phenotypes in the form of agrochemical resistance. These types of questions can hopefully assist us with understanding resistance mechanisms in natural populations, and how we can potentially use this knowledge to better design more targeted approaches when trying to combat agricultural pest species. Although this specific aim is not super relevant to human disease, learning about resistance mechanisms in general can provide insights into how certain aspects of this mechanism can potentially be conserved to humans and may help provide orthogonal evidence to understanding these mechanisms in general.
Your work intersects metabolism, development and cell biology. How do these fields overlap and how do you integrate these disciplines in your research, and what unique insights have emerged from this approach?
In my view, the field of metabolism is so deeply connected with every aspect of biology that when you think of any phenomenon in a biological system, there is a metabolic contribution to it. This is especially interesting when considering a developing organism, and how it has figured out methods to perfectly coordinate its metabolic needs in order to facilitate the existence of multiple cell types at once. As every cell type emerges throughout a given developmental program, the metabolic needs of each one of those cell types changes to then carry out more specialized functions. For example, mutations that affect metabolic systems which arise in the germline of a fly may or may not have an immediate impact on certain processes early in development, but then those changes are more apparent when certain cell types become more differentiated and specialized. As my work focuses on how mutations can impact metabolic phenotypes, understanding how these impacts manifest in different developmental stages will provide a more complete picture of the physiological mechanisms at play.
You have worked with both in-vivo and in-vitro systems. Tell us about their roles and how important it is to study both in the context of both normal development and diseases?
In vitro and in vivo systems both have their pros and cons. In my experience, working with in vitro cell culture was an easy way to get a generalized picture of how basic cell biology occurs and what aspects of metabolism are altered in the face of perturbation in the most basic biological level. Also, cell culture is actually very easy, as you just have to split your cells before confluence, switch out media, and freeze the cells when you don’t need them. In vitro systems do allow you to interrogate a disease space without the ethical constraints associated with testing ideas in actual patients, so it is still a very powerful method to test hypotheses about disease mechanisms.
In vivo systems, such as Drosophila, are a bit more difficult to maintain, but also more interesting in the context of multicellularity and development. Flies have longer generation times than cells in a dish, so it could be a few months before you have your transgenic line needed for your experiments. However, Drosophila do provide a very testable platform to study inheritance patterns, the developmental context of specific phenotypes, and metabolic aspects in the context of a whole organism.
Tell us about the experimental approach/techniques you are using for your project.
Luckily for me, Dr. Xueying Li, a previous postdoc in the lab, developed a method which involves fusing transcription factors with a cytosine deaminase domain that induces mutations in proximal regions of specific transcription factor binding sites. This method, termed TF-HighEvolutionary, can be used to induce targeted mutations within networks of interest and can potentially lead to new phenotypic outcomes. My approach involves using this tool in combination with a lab evolution setup, so the hard part will be having to wait a while for cool results to emerge!
Tell us about how you see the future of metabolism evolve with the new upcoming tools.
In my work, I have extensively used various isotope tracing techniques measured by targeted and untargeted liquid chromatography-mass spectrometry (LC-MS), but I think newer techniques that allow you to visualize different metabolites, such as metabolic biosensors or combined spatial assays with cell- or tissue-level resolution, are going to be very useful for the future of metabolic studies.
What are your upcoming plans? What metabolic pathways or signals you aim to investigate further to understand their role in cell fate/cancer progression?
As I worked on the NRF2/antioxidant pathway during my time at the Hutch, I am really interested in investigating this orthologous pathway in Drosophila as well. I am excited to see what physiological aspects of this pathway are conserved, and how it can operate in a non-oncogenic developmental context.
What changes have you seen in the scientific community in regard to studying unique aspects of metabolic signaling in flies?
I think there are many great groups studying many fascinating aspects of metabolic signaling. I really appreciate how the field is moving toward investigating not only specific pathways, but how these pathways are integrated as a system rather than considering them in isolation. I do believe we are moving toward a more nuanced understanding when we consider them on a systems-level and I can’t wait to see what the future holds for the field.
What role does curiosity play in your life, both within and outside of science? What motivates you to be a basic science researcher?
Curiosity is the fuel that keeps all scientists going! Curiosity is crucial to make it through difficult periods that are inherent to science, and I am grateful to have new curiosities to pursue everyday. I think basic science research is the diesel that fuels our societal understanding of the natural world around us. I find it a great honor and privilege to be able to contribute my career to help broaden our collective understanding of how nature works. Knowing that something I observe in the lab may be the first time it has ever been observed is literally the most exciting thing you could experience. To me, there is no better way to spend the day! I believe that basic science questions lay the foundation for the rest of the scientific community to build upon, and aid in advancing our collective health as a society. Without basic science research, the realm of health advances that we have achieved would eventually collapse. Understanding basic principles of how biology occurs in a general sense is the only way we can progress in the fields of health sciences as well, as many basic science and clinical questions overlap. Moreover, I am a huge plant lover and have many plants in my house. I find having plants in my home and seeing all the crazy developmental stages and cell-type changes that occur when you propagate them brings me back to why I find biology so cool!
In the second part of our conversation – Between Molecules and Milestones, Anna reflects on how her training shaped her scientific independence, shares her journey from research in the U.S. to graduate studies in Germany, and discusses how she transitioned — all while holding on to her fascination with the many roles metabolism plays across biology.
Previously we learnt about the role of metabolism in developmental patterning and embryogenesis. Check out – Metabolic Origins: Steering of early developmental fatefeaturing Kristina Stapornwongkul. Krisitina will be starting her lab at her own lab at IMBA, Vienna and will be hiring soon. Check out her lab page here!
This week we’ll meet Dr Kristina Stapornwongkul, a new incoming faculty at IMBA, Vienna where her lab will focus on how metabolism influences the dynamic process of embryonic development. Kristina’s journey into the world of biology began with a simple school experiment involving potatoes, iodine, and saliva—an early lesson in the unseen chemical choreography that drives life. Today, she is at the forefront of a rapidly evolving field that explores how metabolism shapes embryonic development. With a background in developmental biology and a growing toolkit of synthetic and molecular approaches, Kristina investigates how cellular metabolism influences stem cell fate decisions during the earliest stages of life. Her recent work using gastruloids – a stem cell-based model of early embryos, reveals how metabolic pathways like glycolysis do more than supply energy; they act as key regulators of signaling and pattern formation. She often refers to metabolites and metabolic enzymes as “moonlighting” agents, highlighting their unexpected and influential roles in directing cellular behaviour. As she prepares to launch her own lab at IMBA in Vienna, Kristina is driven by a deep curiosity about how cells make decisions under changing nutritional conditions, and how robust development is maintained despite metabolic challenges. Through her interdisciplinary lens, she brings fresh insights into how environmental and cellular metabolism shape the blueprint of life. Check out her Lab page here and give her a follow over Twitter and Bluesky. She will be hiring soon at all levels so please reach out to her if you’re interested !
What was your first introduction to the field of metabolism, what’s your first memory?
It was actually the first experiment I ever did in school: an iodine starch test with potatoes. We took a potato slice and applied saliva to one half before adding the iodine solution, which normally turns black in the presence of starch. The half without saliva turned black as expected, while the other half didn’t—showing that something in the saliva had already broken down the starch into simpler sugars. That clear, visual result was such a striking demonstration of how our bodies are built to break down food, and I think that’s why it made such a lasting impression on me.
Tell us how you got interested in the field of nutritional and metabolic aspects of animal development from a cell cycle/cell fate perspective?
I did my Master thesis in the Aulehla lab which did some pioneering work in the field of developmental metabolism at that time. It was a completely new and fascinating concept for me. So even though I didn’t work on a metabolism-related project myself at that time, it really got me interested in that topic.
Your work intersects metabolism, development and cell biology. How do you integrate these disciplines in your research, and what unique insights have emerged from this approach?
To understand how metabolism shapes development, I believe we need to uncover molecular mechanisms at the cellular level and understand how they influence tissue-level behaviour and function. So far, my work has been mainly based on developmental and synthetic biology approaches. Looking ahead, I would like to incorporate mass spec-based readouts and develop new tools to manipulate metabolism in a targeted manner.
Introduce us to the field of embryonic development and how does cellular metabolism influence stem cell behavior and fate decisions during embryonic development. Tell us about your recent work on the relationship between glucose metabolism and signaling pathways during cell fate determination during embryonic development?
In the last decade, it has become increasingly clear that metabolic pathways do more than meet the bioenergetic needs of cells—they also play an active role in regulating differentiation. The underlying mechanisms include metabolite-driven post-translational modifications, metabolite-protein interactions, and moonlighting functions of metabolic enzymes, which can influence the epigenetic and signalling state of cells. Based on this, I set out to investigate whether the metabolic state can significantly impact cell fate decisions during the exit from pluripotency.
Using an in vitro model for gastrulation based on mouse embryonic stem cells (gastruloids), we found that inhibiting glycolysis promotes ectodermal differentiation at the expense of mesoderm and endoderm lineages. This effect is dose-dependent, indicating that germ layer proportions can be modulated by adjusting exogenous glucose levels. We further showed that glycolysis acts upstream of key developmental signalling pathways, including Nodal and Wnt, and that its influence on cell fate can be separated from its effects on growth. DOI: 10.1016/j.stem.2025.03.011.
What evidence supports the idea that glycolytic activity acts as a signaling regulator rather than merely an energy source during gastruloid development and what are the broader implications of glycolysis functioning as an activator of morphogen signaling pathways in early development?
The inhibition of glycolysis resulted in the clear downregulation of Nodal and Wnt signalling targets, which are absolutely required for mesoderm and endoderm specification. This suggested that glycolytic activity might be upstream of morphogen signalling. To test this we tried to rescue the phenotype by activating Nodal or Wnt signalling while inhibiting glycolysis. To my surprise, this restored normal germ layer patterning, even though glycolytic activity and overall growth were not recovered. That indicates that glycolysis is not merely fueling signalling but rather functions as an upstream activator!
Tell us about gastruloids as a model. In your view, what advantages do gastruloids offer for understanding early development and metabolism?
The original work establishing gastruloids as a model is here – https://doi.org/10.1242/dev.113001. For me, stem cell-based model systems are an exciting and versatile tool for studying specific processes during development. Pluripotent stem cells are easy to genetically engineer, which opens the door to powerful synthetic and (opto)genetic tools for controlling metabolism in space and time. Their accessibility makes it possible to observe metabolic and signalling dynamics in real time, and the controlled culture conditions allow us to explore how different nutritional environments influence cell behaviour.
How challenging were the experiments in the paper—both in terms of building or standardizing the model, and in the day-to-day logistics?
Mouse gastruloids are a well-established and robust model system, and they were already up and running in the Trivedi Lab when I joined. Thanks to that, it was quite straightforward for me to start working with them. But I did get great help from others in the lab, especially during revisions. So, it was really a team effort.
What are your upcoming plans – what questions are you excited to pursue in future?
I am currently trying to put together an enthusiastic team and tackle some of the questions I am really excited about: How does metabolism influence cell fate decisions? What is the energetic cost of morphogenesis, and do cells adapt their metabolism to overcome energetic constraints? How robust are developmental processes, such as patterning and morphogenesis, to changes in the nutritional environment? We’ll definitely keep an eye on glycolysis, but I’m also really keen to explore other metabolic pathways and see what else we can discover.
What role does curiosity play in your life, both within and outside of science? How important it is for you to answer basic science questions?
I would say that being curious is one of my most important character trait, and I really cherish it. It’s what drives me to explore new people, cultures, places, and ideas. When it comes to basic science questions, I think curiosity is absolutely essential, since you can’t always rely on other motivations, such as direct applications to human health. For me, basic science questions are usually the most exciting ones, and I wouldn’t want to work on anything that doesn’t truly fascinate me. I guess it comes from the longing to understand how life works. How can that not be exciting J?
How can insights from understanding basic science aspects of early development help us understand the impact of maternal nutrition and metabolic microenvironment on embryonic health and the risk of congenital/metabolic disorders in humans?
I think understanding basic science aspects of early development is absolutely crucial to understand the impact of the nutritional environment on embryonic development on a molecular level. We know since a long time that the maternal nutrition impacts even early stages of embryonic development. What we often don’t understand are the phenotypes and their underlying mechanisms. So, it’s important to support basic science on early development to better understand what goes wrong in suboptimal nutritional environments or during metabolic disorders.
Tell us about how you see the future of developmental metabolism and cell fate evolve with the new upcoming tools.
Development happens in time and space, so I believe that visualizing metabolic dynamics is essential for better understanding the role of metabolism during development. Techniques like spatial metabolomics and the use of biosensors will be incredibly valuable for this purpose.
I’m also really excited about the development of new tools that allow us to manipulate metabolic pathways in a spatiotemporal manner. In my recent work, I developed a genetic tool to restrict glucose availability by leveraging a sucrose-cleaving enzyme from yeast, and I’m eager to further refine and expand this approach in the future.
Were there any pivotal moments that shaped your career path? What’s an unexpected place you’ve found inspiration for your work? What advice would you offer to students and early-career scientists interested in exploring the intersections of metabolism and cell fate regulation?
One pivotal moment was seeing a zebrafish embryo develop during an undergraduate course (thank you, @Gerrit Begemann!). It was so beautiful and fascinating that I immediately wanted to understand how something like that works.
Not sure, whether there is an unexpected place but I like to think about things I don’t understand (including science) when I am moving between places, especially while cycling. Maybe it’s something about being in motion.
For students early-career scientists and actually everyone interested in the intersection of metabolism and cell fate regulation, my advice is to seek as much feedback as possible on your ideas and work. This is a complex and rapidly evolving field, and most of us were trained primarily in either developmental biology or metabolism, but rarely both. Engaging with experts from different backgrounds can really broaden your perspective and strengthen your research.
How do you maintain a balance between your rigorous research activities and personal life?
I really like to do outdoor sports, such as rock climbing and beach volleyball. It helps me to clear my head.
If you hadn’t embarked on a career in biological research, what other profession might you have pursued, and why?
That’s a tough question—I really love what I do! But if I hadn’t gone into biological research, I think I’d still want a career where I’m surrounded by smart, creative people and constantly learning new things. Whether it was in education, technology, or even the arts, the most important thing for me would be working in an environment that challenges me intellectually and encourages curiosity.
Anything you’d want to highlight for the future !
Yes, I actually will be starting my lab at IMBA Vienna in September! We’ll be looking at environmental and metabolic regulators of embryonic development. There is more info on our website (https://www.oeaw.ac.at/imba/groups/kristina-stapornwongkul). So please reach out if you feel enthusiastic to join the team!
Previously we learnt about the role of metabolism in seasonal adaptations and phenotypic plasticity using two unique insect models – butterflies and budworms. Check out – The season’s script: Tales of Metabolic adaptation (Karin Van Der Burg).
Spotted a preprint in this list that you love? If you’re keen to gain some science writing experience and be part of a friendly, diverse and international community, consider joining preLights and writing a preprint highlight article.
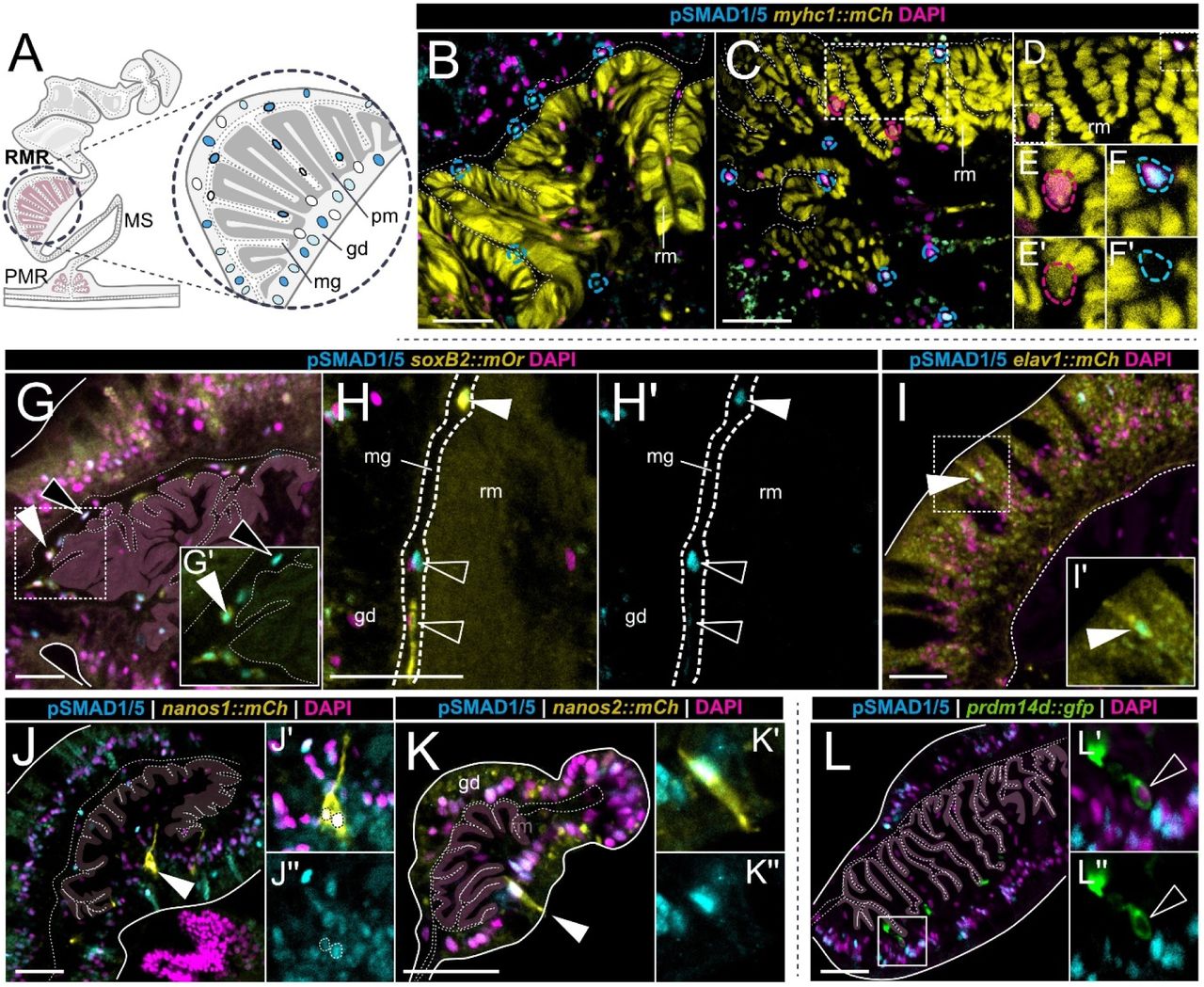
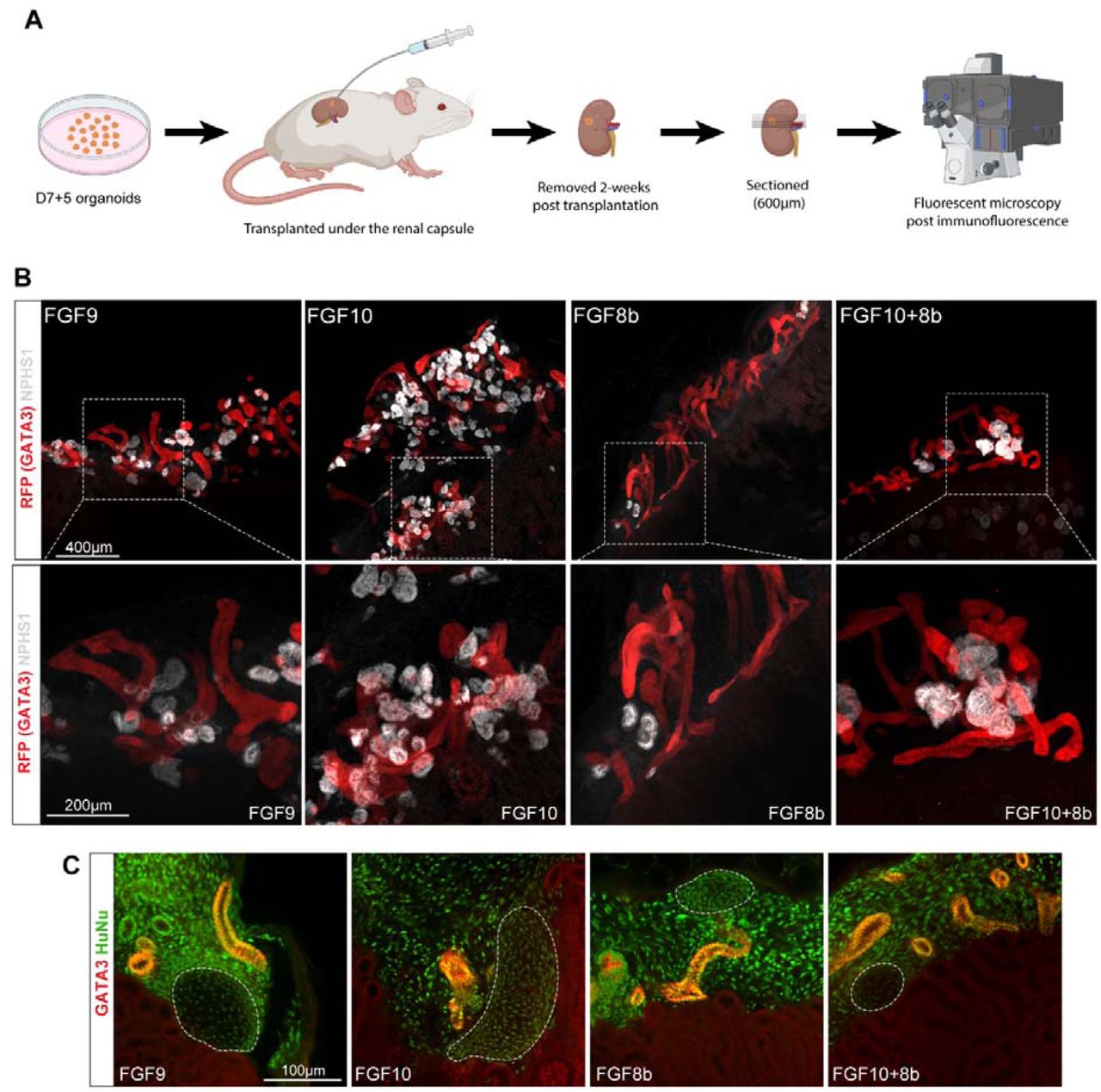
Ravindra B. Chalamalasetty, Haley Tran, Ryan Kelly, Samuel Kuo,Mark W. Kennedy,Moonsup Lee, Sara Thomas,Nikolaos Mandalos,Vishal Koparde,Francisco Pereira Lobo,Terry P. Yamaguchi
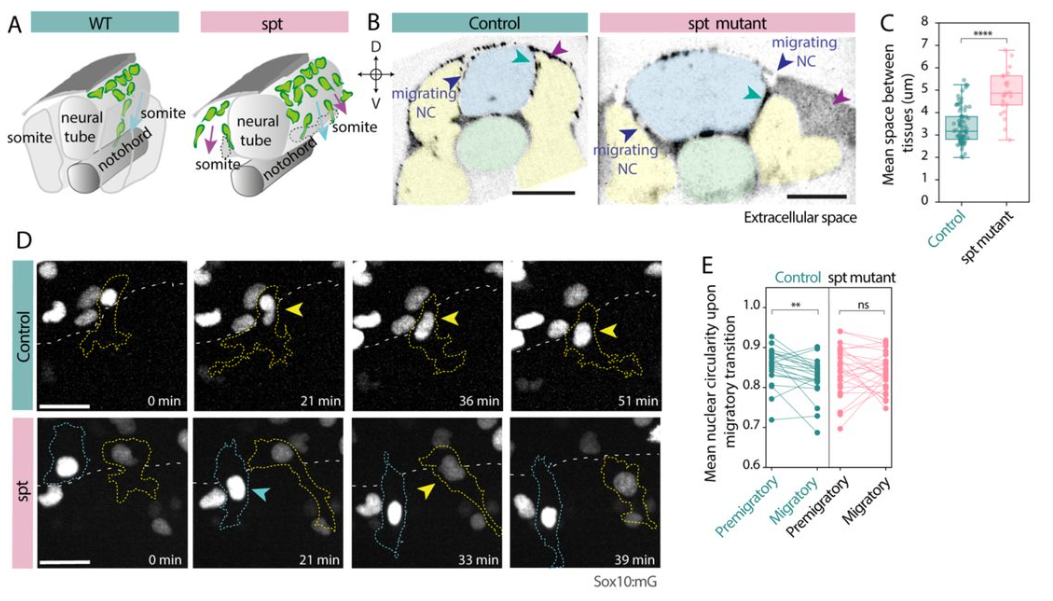
Auriana Arabpour, Jonathan Adam DiRusso, Qiu Ya Wu, Mark Larsen, Young Sun Hwang, Elsie Jacobson, Thi Xuan Ai Pham, Nicole Agranonik, Megan Sparrow, Vernon Leander Monteiro, Zenya Rebecca Bian, Nicolas Pelaez-Restrepo, Antuca Callejas-Marin,Vincent Pasque,Kathrin Plath, Amander T. Clark
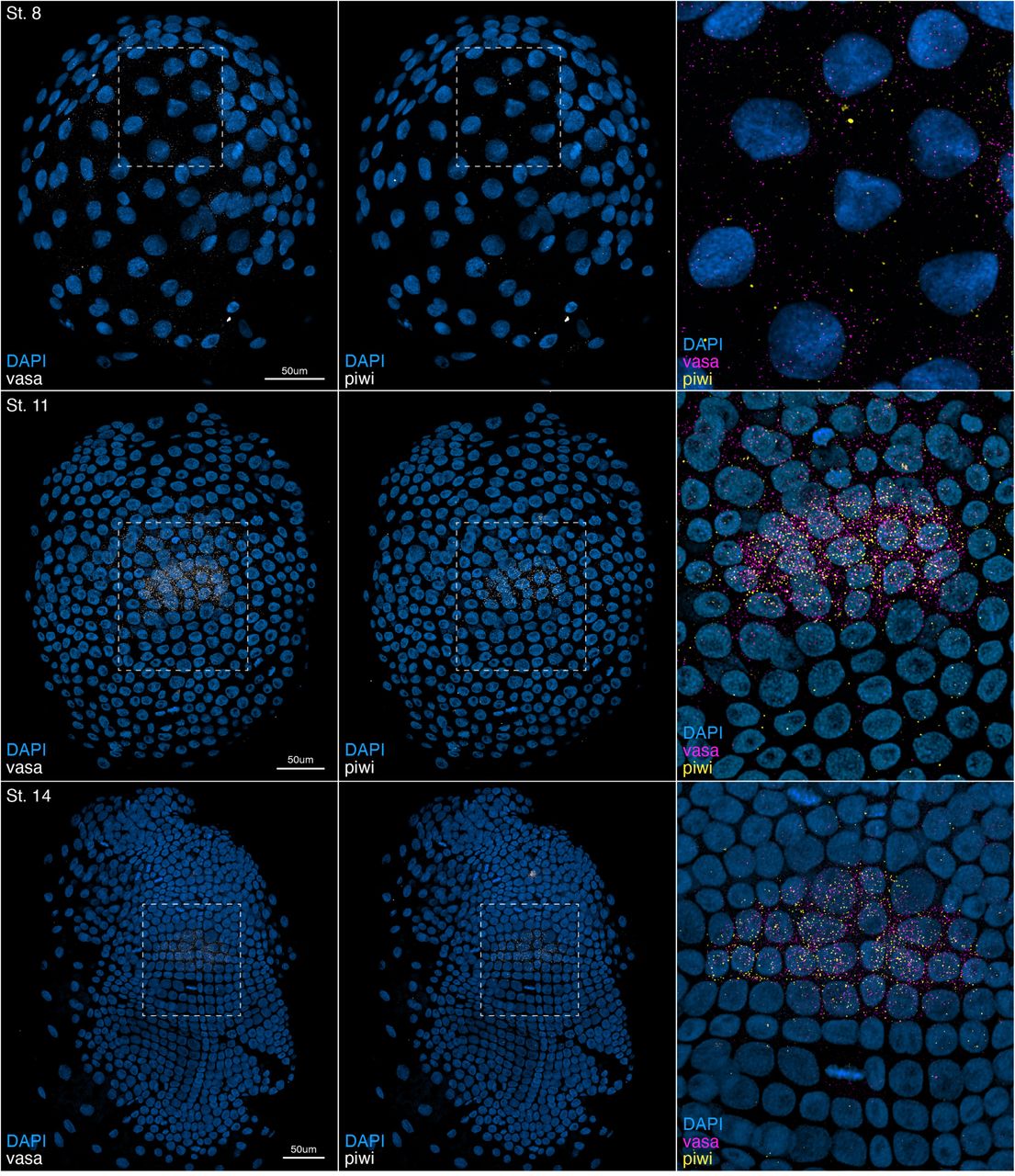
Elizabeth Abraham, Thomas Roule, Aidan Douglas, Emily Megill, Olivia M. Pericak, Jordan E. Howe, Carmen Choya-Foces, Joanne F. Garbincius, Henry M. Cohen, Paula Roig-Flórez, Mikel Zubillaga, Mark D. Andrake, Seonhee Kim, John W. Elrod, Naiara Akizu, Conchi Estaras
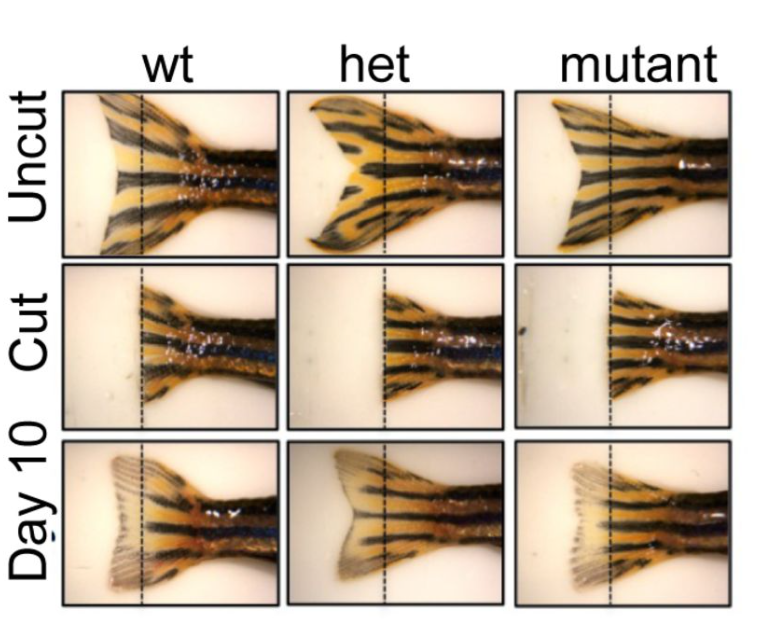
Christopher J. Panebianco, Yuming Huang,Nidal Khatib,Devin C. Gottlieb,Maha Essaidi,Saima Ahmed,Nathaniel A. Dyment,Rebecca A. Simmons,Joel D. Boerckel,Niamh C. Nowlan
Jeremy E Sandler, Ya-Yin Tsai, Shiyuan Chen, Logan Sabin, Mark E. Lush, Abhinav Sur, Elizabeth Ellis, Nhung TT Tran, Malcolm Cook, Allison R Scott, Jonathan S. Kniss,Jeffrey A. Farrell, Tatjana Piotrowski
Hyunwook Lee, Abigail Jaquish, Sharlene Fernandes, Barbara Zhao, Amber Elitz, Kathleen Cook, Sarah Trovillion,Natalia Bottasso-Arias, Simon J. Y. Han, Samantha Goodwin, Nicholas X. Russell,Amanda L. Zacharias, Samantha A. Brugmann,Jeffrey A. Whitsett,Debora Sinner, Xin Sun, Daniel T. Swarr,William J. Zacharias
Madeleine Fosslie, Erkut Ilaslan, Trine Skuland, Adeel Manaf, Mirra Louise Cicilie Soegaard, Marie Indahl, Maria Vera-Rodriguez, Rajikala Suganthan, Ingunn Jermstad, Shaista Khanam, Knut Tomas Dalen,Ragnhild Eskeland, Michel Choudalakis, Magnar Bjoras, Peter Zoltan Fedorcsak, Gareth David Greggains, Mika Zagrobelny,John Arne Dahl,Mads Lerdrup
Rocío B. Marquez,Estefanía Sánchez Vázquez,Andrés M. Alonso, Yanel E Bernardi,Emilio M. Santillan,Peter Lwigale,Luisa Cochella,Marianne E. Bronner,Pablo H. Strobl-Mazzulla
Pavel A. Komarov, Philipp Bammer,Ching-Yeu Liang,Hans-Rudolf Hotz,Grigorios Fanourgakis,Sunwoo Chun,Hubertus Kohler,Tim-Oliver Buchholz,Jean-Francois Spetz,Antoine H.F.M Peters
Joyce Esposito,Felipe de Souza Leite,Igor Neves Barbosa,Thaís Maria da Mata Martins, Giovanna Gonçalves de Oliveira Olberg,Ziad Al Tanoury,Kayque Alves Telles-Silva,Mayana Cristina da Silva Pardo,Tatiana Jazedje,Raul Hernandes Bortolin,Mario Hiroyuki Hirata,Olivier Pourquié,Mayana Zatz
Allara K. Zylberberg,Emma I. Scully,Pei Xuan Er, Hannah Baric, Michelle Scurr,Mian Xie,Thanushi Peiris,Sara E. Howden,Kynan T. Lawlor,Melissa H. Little
Angelica Varesi, Murtaza S. Nagree, Isabella Di Biasio, Andy G.X. Zeng, Sayyam Shah, Hyerin Kim, Michael Zhang, Alex Murison, John E. Dick,Stephanie Z. Xie
Daniel P. Moore, Heather B. Steele-Stallard, Luca Pinton, Valentina Maria Lionello, Lucia Rossi, Artadokht Aghaeipour, Salma Jalal, Cherry Tsz Yan Wong, Angela Clara-Hwang, Gisèle Bonne, Peter S. Zammit,Francesco Saverio Tedesco
Nicole Szeluga, Noor AlBader, Samantha Pelletier, Kylie Weis, Arielle Johnson, Noah Fahlgren,Mikhaela Neequaye, Gus Vogt, Ryan DelPercio,Patricia Baldrich, Kelsey J.R.P. Byers,Blake C. Meyers,Margaret H. Frank
Jim Renema,Svitlana Lukicheva, Isabelle Verwaerde,Stijn Aesaert,Griet Coussens,Jolien De Block,Carolin Grones,Thomas Eekhout,Bert De Rybel,Rhoda A.T. Brew-Appiah, Christopher A. Bagley,Lennart Hoengenaert,Klaas Vandepoele,Laurens Pauwels
Chancelor B. Clark,Denise Caldwell, Qiang Zhu, Dominic Provancal, Austin C. Edwards, Qijian Song, Charles V. Quigley,Anjali S. Iyer-Pascuzzi, Jianxin Ma
Allan M. Carrillo-Baltodano,Emmanuel Haillot,Steffanie Mutiara Meha, Imran Luqman, Artenis Pashaj, Yun-Ju Lee,Tsai-Ming Lu,David E. K. Ferrier,Stephan Q. Schneider,José M. Martín-Durán
Zaili Luo, Mingyang Xia, Feng Zhang, Dazhuan Xin, Rohit Rao, Karrie M. Kiang, Kalen Berry, Yu Xiong, Hongqi Liu, Yifeng Lin, Ming Hu, Mei Xin, Jie Ma, Hao Li, Michael D. Taylor, Wenhao Zhou, Q. Richard Lu
Chenhao Tong, Tao Lin, Han Wang, Carolina De La Torre, Hui Liu, Chen Shao, Seddik Hammad, Roman Liebe, Matthias P Ebert, Huiguo Ding, Steven Dooley, Hong-Lei Weng
E. I. Bakhmet,E. V. Potapenko,O. Y. Shuvalov,A. A. Lobov,E. A. Repkin,N. E. Vorobyeva, A. N. Korablev,A. S. Zinovyeva, A. A. Kuzmin,N. D. Aksenov,A. T. Kopylov,G. Wu,H. R. Schöler,A. N. Tomilin
Siddharth Acharya,Eric Hanssen,Veronica B. Botha, Tia M. Smith,Sahan Jayatissa, Zlatan Trifunovic,Lucy A. Bartho,John E. Schjenken,Tu’uhevaha J. Kaitu’u-Lino,Anthony V. Perkins,Joanna L. James,Kirsty G. Pringle,James C. Bouwer,Roger Smith,Joshua J. Fisher
Suma Garibova, Eva Stickler, Fatima Al Ali, Maha A Abdulla, Abbirami Sathappan,Sahar Da’as, Lilian Ghanem, M Nadhir Djekidel, Rick Portman,Matteo A Avella
Abhrajit Ganguly, Cynthia M Carter, Aristides Rivera Negron, Hua Zhong, Alvaro Moreira, Matthew S. Walters, Lynette K Rogers, Y S Prakash, Trent E Tipple, Arlan Richardson
Christina M. Kaszuba, Sonali Sharma, Benjamin J. Rodems, Cameron D. Baker, Palomi Schacht, Takashi Ito, Kyle P. Jerreld, Chen Yu, Edgardo I. Franco, Emily R. Quarato, Francisco A. Chaves, Jane L. Liesveld, Laura M. Calvi, Hani A. Awad, Roman A. Eliseev,Jeevisha Bajaj
Eloïse Bertiaux, Vincent Louvel, Caitlyn L. McCafferty, Hugo van den Hoek, Umut Batman, Souradip Mukherjee,Lorène Bournonville, Olivier Mercey, Isabelle Mean, Adrian Müller, Philippe Van der Stappen, Garrison Buss, Jean Daraspe,Christel Genoud, Tim Stearns,Benjamin D. Engel,Virginie Hamel,Paul Guichard
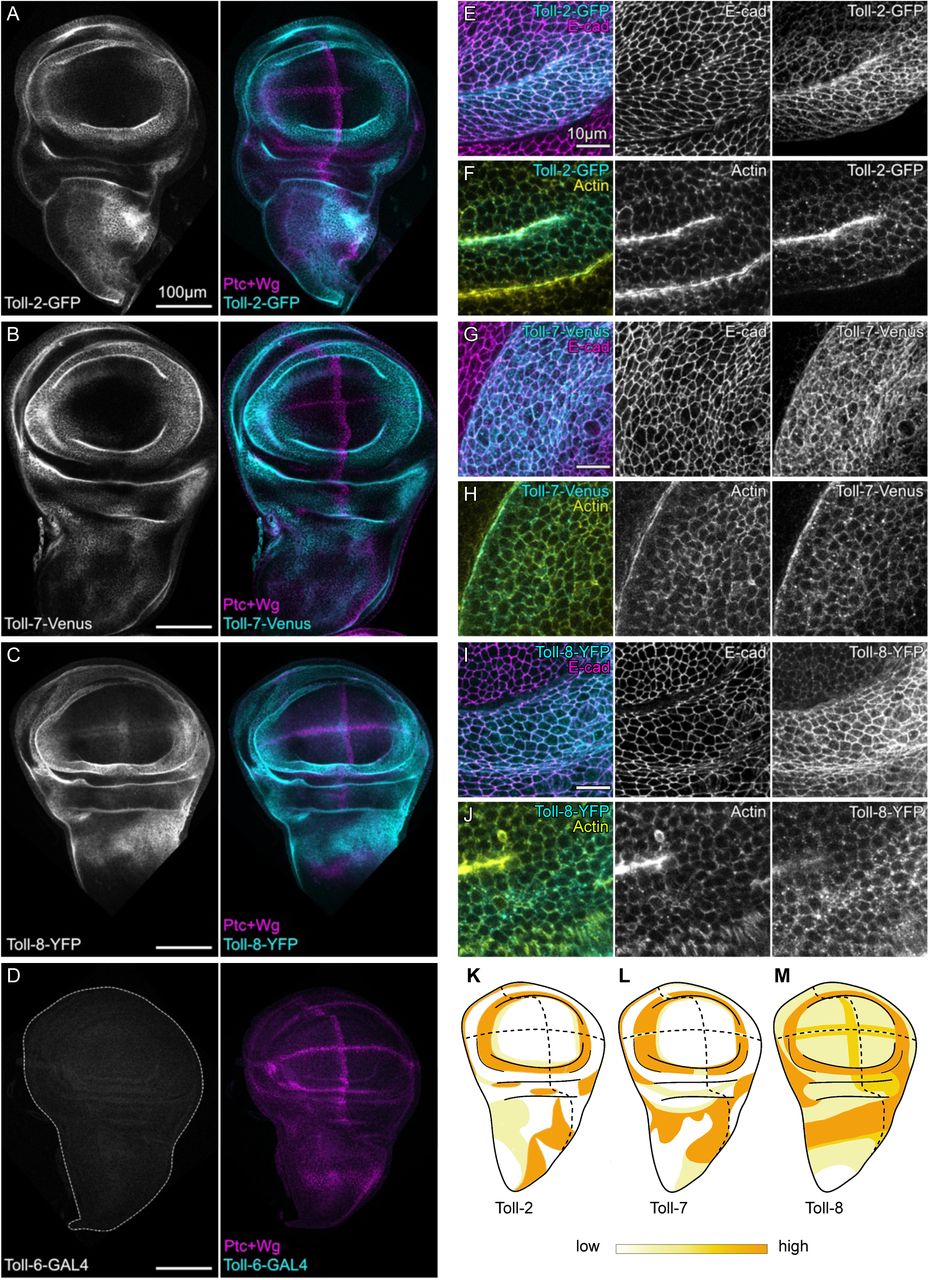
Rotem Leshem, Syed Murtuza Baker, Joshua Mallen, Lu Wang, John Dark, Andrew D Sharrocks, Karen Piper Hanley, Neil A Hanley, Magnus Rattray, Simon D Bamforth, Nicoletta Bobola
Stanley E. Strawbridge,Lawrence E. Bates, Connor Ross, Kenneth A. Jones,Takuya Azami,Tim Lohoff,Maike Paramor, Vicki Murray,Ana Luíza Cidral, James Clarke,Maria Rostovskaya, Ge Guo,Jennifer Nichols
I am excited to announce the release of our event carbon calculator, our latest resource to support the creation of sustainable events. Estimate your event’s carbon footprint in five minute or less. Our event carbon calculator provides insights into the main drivers of your meeting’s greenhouse gas emissions, offers reduction scenarios to test and provides resources to help you build a sustainable strategy for your event.
Our calculator is still in its beta stage, so please explore our tool and send us feedback on how to make it better suited for your needs. We hope you will enjoy using our event carbon calculator and that it will help to inspire more sustainable actions in your academic meetings.
At the end of each month, I pick the same month from a random year from the past 15 years of the Node, and take a look at what people were talking about back then.
Previously, I’ve been busy travelling back to February 2011, March 2013, April 2014 and May 2016 to have a look around the Node. It’s been really fun peeking around the archives. This June, let’s jump back to 2013…
We used to partner with the Woods Hole embryology course to run an image competition, with the winning image being featured on Development’s cover. We’re hoping to revive this tradition – watch this space!
The Fly Board is excited to announce the second cycle of FlyCROSS (2025–2026), following a successful launch in 2024. The goal of FlyCROSS is to empower early career Drosophila researchers by connecting them with mentors who can help them fly in their careers – whether in academia or beyond.
Through FlyCROSS, early career scientists in the Drosophila community will be paired with an experienced mentor who will help them navigate the world of fly research with ease. Designed to prioritize the needs of mentees by matching them with mentors of their preferences, the program offers seasoned researchers an opportunity to share their knowledge and pay it forward by supporting scientists who are seeking guidance and fresh perspectives. The program consists of three parts: the mentor survey, the mentee survey, and matching.
We are currently seeking mentors. Scientists with experience in Drosophila research, includingpostdoctoral researchers, faculty members, and those in equivalent roles across diverse scientific career paths are eligible to serve as mentors and invited to complete the Mentor survey by July 14. The mentor survey will gather important details about your areas of expertise and personal career journey. We’re looking for mentors from all career paths—academia, industry, biotech, science communication, policy, and more—who have experience with Drosophila research and are eager to support the next generation of fly scientists. (If you have earned a PhD and are currently involved in any scientific career path and have experience working with fruit flies as a model system at any stage of your career, we encourage you to volunteer to support the next generation scientists)
The prospective mentee survey will be open July 21 through September 1 and will collect in-depth information about the type of mentorship each mentee is seeking. Early-career scientists including graduate students, postdoctoral researchers working in fly labs or equivalent settings, and pre-tenure faculty leading Drosophila research groups are eligible to participate as mentees.
Mentees will review the list of available mentors and their responses, with the option to indicate their preferred mentors. After receiving mentee submissions, the FlyCROSS Committee will carefully review the preferences and finalize pairings, ensuring an optimal match between mentors and mentees. Matches will be announced by October. Incase of any queries reach out to dmelcross@gmail.com. FlyCROSS was adapted from a similar mentoring program running in the worm community, and we sincerely appreciate their support of our endeavor.
Thankyou Shefali FlyCROSS Co-chair Grad student rep, Fly Board PhD candidate, Tennessen Lab IU Bloomington Bluesky : @iamshef.bsky.social X : @iam_shef

 (6 votes)
(6 votes)

 (No Ratings Yet)
(No Ratings Yet)