preLights, our sister community site here at The Company of Biologists, just published an interview with developmental biologist and zebrafish aficionado Christian Mosimann. It covers his research history, career trajectory, and, as in this quote, his views on preprints in science:
“I only preprint papers that we send out for peer review at a journal and not to stake a claim on something, and I’ve found this to be very helpful for job searches and for grants….I think the more we normalise preprinting our work we are confident in, the more we can show everyone that nobody is going to scoop you the very next morning if you put your research on a preprint server.”
Oligodendrocyte precursor cells prune axons in the mouse neocortex JoAnn Buchanan, Leila Elabbady, Forrest Collman, Nikolas L. Jorstad, Trygve E. Bakken, Carolyn Ott, Jenna Glatzer, Adam A. Bleckert, Agnes L. Bodor, Derrick Brittan, Daniel J. Bumbarger, Gayathri Mahalingam, Sharmishtaa Seshamani, Casey Schneider-Mizell, Marc M. Takeno, Russel Torres, Wenjing Yin, Rebecca D. Hodge, Manuel Castro, Sven Dorkenwald, Dodam Ih, Chris S. Jordan, Nico Kemnitz, Kisuk Lee, Ran Lu, Thomas Macrina, Shang Mu, Sergiy Popovych, William M. Silversmith, Ignacio Tartavull, Nicholas L. Turner, Alyssa M. Wilson, William Wong, Jingpeng Wu, Aleksandar Zlateski, Jonathan Zung, Jennifer Lippincott-Schwartz, Ed S. Lein, H. Sebastian Seung, Dwight E. Bergles, R. Clay Reid, Nuno Maçarico da Costa
Size-dependent protein segregation creates a spatial switch for Notch signaling and function Minsuk Kwak, Kaden M. Southard, Woon Ryoung Kim, Nam Hyeong Kim, Ramu Gopalappa, Minji An, Hyun Jung Lee, Min K. Kang, Seo Hyun Choi, Justin Farlow, Anastasios Georgakopoulos, Nikolaos K. Robakis, Matthew L. Kutys, Daeha Seo, Hyeong Bum Kim, Yong Ho Kim, Jinwoo Cheon, Zev J. Gartner, Young-wook Jun
Effects of gestational age at birth on perinatal structural brain development in healthy term-born babies Oliver Gale-Grant, Sunniva Fenn-Moltu, Lucas França, Ralica Dimitrova, Daan Christaens, Lucilio Cordero-Grande, Andrew Chew, Shona Falconer, Nicholas Harper, Anthony N Price, Jana Hutter, Emer Hughes, Jonathan O’Muircheartaigh, Mary Rutherford, Serena J Counsell, Daniel Rueckert, Chiara Nosarti, Joseph V Hajnal, Grainne McAlonan, Tomoki Arichi, A David Edwards, Dafnis Batalle
A single short reprogramming early in life improves fitness and increases lifespan in old age Quentin Alle, Enora Le Borgne, Paul Bensadoun, Camille Lemey, Nelly Béchir, Mélissa Gabanou, Fanny Estermann, Christelle Bertrand-Gaday, Laurence Pessemesse, Karine Toupet, Jérôme Vialaret, Christophe Hirtz, Danièle Noël, Christian Jorgensen, François Casas, Ollivier Milhavet, Jean-Marc Lemaitre
Molecular diversity and lineage commitment of human interneuron progenitors Dmitry Velmeshev, Manideep Chavali, Tomasz J. Nowakowski, Mohini Bhade, Simone Mayer, Nitasha Goyal, Beatriz Alvarado, Walter Mancia, Shaohui Wang, Matthew Speir, Maximilian Haeussler, David Rowitch, Arturo Alvarez-Buylla, Eric J. Huang, Mercedes Paredes, Arnold Kriegstein
Novel epigenetic clock for fetal brain development predicts prenatal age for cellular stem cell models and derived neurons Leonard C. Steg, Gemma L. Shireby, Jennifer Imm, Jonathan P. Davies, Alice Franklin, Robert Flynn, Seema C. Namboori, Akshay Bhinge, Aaron R. Jeffries, Joe Burrage, Grant W. A. Neilson, Emma M. Walker, Leo W. Perfect, Jack Price, Grainne McAlonan, Deepak P. Srivastava, Nicholas J. Bray, Emma L. Cope, Kimberly M. Jones, Nicholas D. Allen, Ehsan Pishva, Emma L. Dempster, Katie Lunnon, Jonathan Mill, Eilis Hannon
Longitudinal dynamics of clonal hematopoiesis identifies gene-specific fitness effects Neil A. Robertson, Eric Latorre-Crespo, Maria Terradas-Terradas, Alison C. Purcell, Benjamin J Livesey, Joseph A. Marsh, Lee Murphy, Angie Fawkes, Louise MacGillivray, Mhairi Copland, Riccardo E. Marioni, Sarah E. Harris, Simon R. Cox, Ian J. Deary, Linus J. Schumacher, Kristina Kirschner, Tamir Chandra
In vitro models of the human esophagus reveal ancestrally diverse response to injury Daysha Ferrer-Torres, Joshua H. Wu, Charles J. Zhang, Max A. Hammer, Michael Dame, Angeline Wu, Emily M. Holloway, Kateryna Karpoff, Caroline L. McCarthy, Margaret S Bohm, Sha Huang, Yu-Hwai Tsai, Simon P. Hogan, Danielle Kim Turgeon, Jules Lin, Peter D.R. Higgins, Jonathan Sexton, Jason R. Spence
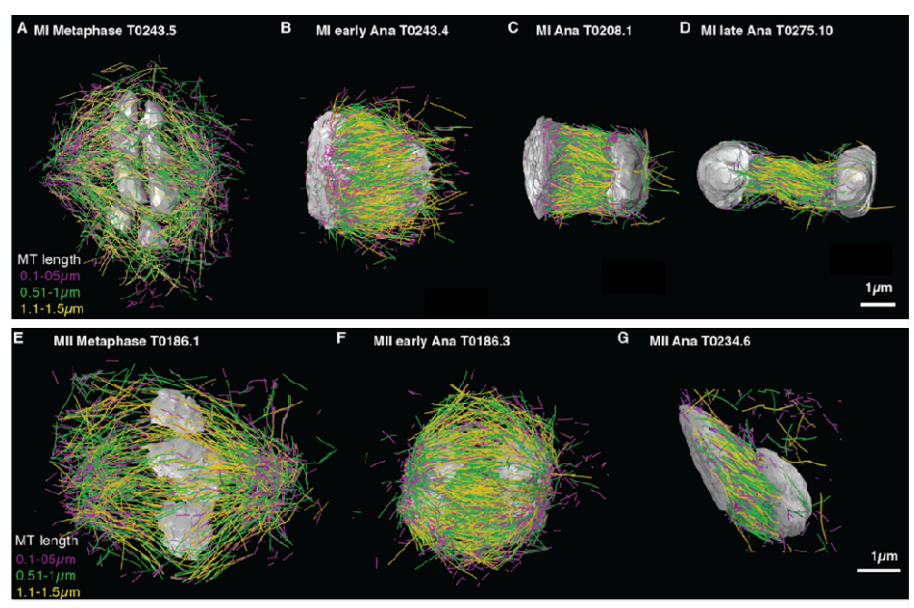
Microtubule re-organization during female meiosis in C. elegans Ina Lantzsch, Che-Hang Yu, Yu-Zen Chen, Vitaly Zimyanin, Hossein Yazdkhasti, Norbert Lindow, Erik Szentgyörgyi, Ariel Pani, Steffen Prohaska, Martin Srayko, Sebastian Fürthauer, Stefanie Redemann
Scaling of cellular proteome with ploidy Galal Yahya, Paul Menges, Devi Anggraini Ngandiri, Daniel Schulz, Andreas Wallek, Nils Kulak, Matthias Mann, Patrick Cramer, Van Savage, Markus Raeschle, Zuzana Storchova
Minian: An open-source miniscope analysis pipeline Zhe Dong, William Mau, Yu (Susie) Feng, Zachary T. Pennington, Lingxuan Chen, Yosif Zaki, Kanaka Rajan, Tristan Shuman, Daniel Aharoni, Denise J. Cai
“How do we do this at a distance?!” A descriptive study of remote undergraduate research programs during COVID-19 Olivia A. Erickson, Rebecca B. Cole, Jared M. Isaacs, Silvia Alvarez-Clare, Jonathan Arnold, Allison Augustus-Wallace, Joseph C. Ayoob, Alan Berkowitz, Janet Branchaw, Kevin R. Burgio, Charles H. Cannon, Ruben Michael Ceballos, C. Sarah Cohen, Hilary Coller, Jane Disney, Van A. Doze, Margaret J. Eggers, Stacy Farina, Edwin L. Ferguson, Jeffrey J. Gray, Jean T. Greenberg, Alexander Hoffman, Danielle Jensen-Ryan, Robert M. Kao, Alex C. Keene, Johanna E. Kowalko, Steven A. Lopez, Camille Mathis, Mona Minkara, Courtney J. Murren, Mary Jo Ondrechen, Patricia Ordoñez, Anne Osano, Elizabeth Padilla-Crespo, Soubantika Palchoudhury, Hong Qin, Juan Ramírez-Lugo, Jennifer Reithel, Colin A. Shaw, Amber Smith, Rosemary Smith, Adam P. Summers, Fern Tsien, Erin L. Dolan
We are happy to announce that the UK Chick Developmental Biology Meeting 2021 will be held online on Friday 10th September.
This one-day virtual meeting brings together researchers from Universities and Research Institutes across the United Kingdom to promote and further our in-depth strength in using the chick embryo as a model organism to study a variety of topics in developmental biology. The meeting itself is a mix of talks from junior and senior developmental biologists and aims to stimulate productive interactions between research groups and individuals from different subject areas to exchange knowledge and expertise. This meeting will also provide a platform of support for early career researchers to engage with the community during a difficult time.
We are delighted to have Prof Marianne Bronner (Caltech, USA) as our international keynote speaker to join other invited speakers from the UK such as Prof Kate Storey (Dundee), Dr Matt Towers (Sheffield), Dr Siobhan Loughna (Nottingham), Dr Mike McGrew (Roslin) and Dr Fengzhu Xiong (Cambridge).
In addition we will have talks from early career researchers (i.e. those just starting their labs, post docs and graduate students) – so please do let us know if you or someone in your lab has a nice story that they could present please contract and email Dr Gi Fay Mok (g.mok@uea.ac.uk) or Dr Eirini Maniou (e.maniou@ucl.ac.uk) to discuss and submit a short abstract. The deadline for abstract enquiries and submissions is Friday 15th July.
In this episode of the Genetics Unzipped podcast we’re taking to the night skies with a closer look at the genetics of bats. Usually the stuff of horror films and Hallowe’en, these fascinating mammals have many important genetic secrets to share with us about evolution, longevity, immunity and more.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
This post is to let you know about an initiative to form a European Drosophila Society (EDS), which aims to represent our research community, support common initiatives and infrastructure, as well as oversee the organisation of the biennial European Drosophila Research Conference. We have set up a web site (http://europeandrosophilasociety.org/) where you can register as an EDS member (free!). You can also follow us on Twitter @Fly_EDS. You will find information about Drosophila community news and job postings, as well as the EDRC conferences.
The EDS will be overseen by the European Drosophila Board (EDB), which will reflect the geographic and thematic diversity of our community. To get the EDS under way, we have formed an initial EDB, and plan to hold elections in 2023 to renew EDB membership. Our aim is to be inclusive: we wish to represent and support the whole breadth of European Drosophila research, as well as establish strong ties with other invertebrate/vertebrate/model organism research communities. The EDS exists to promote your research and can only work with your support, therefore we urge to join the EDS by visiting the web site and signing up. You can also contact board members to propose initiatives or offer help to our working groups.
All registered members of the community can post news, events and jobs on the web site, as well as search for Drosophila researchers based on expertise or research interests using our search engine. Group leaders can add their lab to the interactive map of European Drosophila labs. We welcome suggestions of other services we could offer to the Drosophila community.
Sincerely yours,
The European Drosophila Board Michael Boutros, Nick Brown, Virginie Courtier, Eileen Furlong, Alex Gould, Ilona Grunwald Kadow, Gabor Juhasz, Estee Kurant, Bruno Lemaitre, Marco Milan, Lisa Meadows, Ruth Palmer, Frank Schnorrer, Nic Tapon, Luis Teixeira
On Wednesday 12 May Development welcomed three more researchers with interests in developmental neurobiology to our seventh Development presents… webinar.
Below you’ll find each of the talks, plus a Q&A chaired by Development Editor Paola Arlotta. The next #DevPres webinar will be held on 14 April 2021, and chaired by Steve Wilson – subscribe to our mailing list for updates.
Tian Tian (University of Edinburgh) – ‘Pax6 regulates the morphological and electrophysiological development of mouse prethalamic neurons’
For our ninth webinar in the Development presents… series, Development Editor Maria-Elena Torres-Padilla (Institute of Epigenetics and Stem Cells) has invited three talks on the topic of chromatin regulation and epigenetics.
Jessica Zuin (Postdoc in Luca Giorgetti‘s lab at the Friedrich Miescher Institute for Biomedical Research) ‘Nonlinear control of transcription through enhancer-promoter interactions’
The webinar will be held in Remo, our browser-based conferencing platform. After the talks you’ll have the chance to meet the speakers and other participants at virtual conference tables. If you can’t make it on the day, talks will be available to watch after the event on the Node. You can also sign up to our mailing list for email alerts.
By Bernat Corominas-Murtra and Nicoletta I. Petridou
Embryo morphogenesis is a play whose outcome is the result of a complex and delicate plot, made of balances and agreements among many actors: the execution of the genetic program, biochemical communication among cells, mechanical forces, energy consumption, geometry, cell autonomous and collective behaviours… all of them interlinked in a precise way in space and time. This process is by no means always smooth, and in some developmental stages, dramatic, sudden shifts in the properties or structure of the embryo occur.
It was precisely one of these sudden shifts that started puzzling us about three years ago during a scientific retreat in the beautiful town of Sitges, in the Catalan coast, between the groups of Edouard Hannezo and Carl-Philipp (CP) Heisenberg from IST Austria. At that time, I (Nicoletta) was a postdoc in CP’s group, and one of the targets of our research was to establish how tissue material properties change in space and time within the zebrafish embryo at the onset of morphogenesis. At this stage, the embryo is composed of a few thousand cells (the blastoderm) sitting on top of a yolk cell. The blastoderm starts spreading and engulfing the yolk, defining the onset of gastrulation. By measuring blastoderm viscosity in vivo we knew that the viscosity of the embryo tissue drops by more than an order of magnitude at the exact moment when morphogenesis starts, in a highly reproducible way. That is, in a matter of minutes, the embryo jumps from a “solid” non-deformable state to a “fluid” highly-deformable one. Right after this abrupt shift, the tissue starts recovering its viscous properties by undergoing “thickening” (in rheological terms, like a honey that thickens and loses deformability). These spatiotemporal changes in tissue viscosity are essential for correct tissue spreading at the onset of morphogenesis. As a funny coincidence, exactly on the very first day of the retreat we had just found out that our manuscript on these findings was accepted for publication (Petridou et al., 2019). Besides the celebratory mood, we were still excited and puzzled by the abrupt nature of this shift, in which the intensity and speed of the changes reminded us of what in physics is called a “phase transition”.
But, what kind of phase transition? The theory of phase transitions is a broad field in physics. It is maybe true that most of the results of the theory of phase transitions lie far away from biology; but it is also true that a lot of this theory is becoming more and more relevant in understanding biological phenomena. This was an intriguing a topical problem with several theoretical models of material phase transitions emerging (Petridou and Heisenberg, 2019; Lawson-Keister and Manning, 2021), but rather disappointingly we couldn’t experimentally match these models to our in vivo system. During the poster session of the retreat, I (Bernat, who had just joined Edouard’s group as his first postdoc) was discussing these issues with Nicoletta. Soon after, we came up with the idea that this “phase transition”-like behaviour could be addressed as a network phase transition. A network phase transition is a sudden change of the network properties that is usually associated to smooth changes in the connectivity of the nodes. For example, if one removes connections of a network gradually, we will observe that the net commonly remains more or less stable until it abruptly breaks into many small fragments. In this particular example, the exact point of the connectivity that triggers the fragmentation is a critical point of a phase transition between unconnected and connected network regimes, called “percolation”. Network phase transitions play a relevant role in understanding ecosystems, epidemic spreading or material properties, among many other systems. In the past I had been working on several problems involving network phase transitions, and the idea of explaining phenomena in embryo development with this framework was as much exciting as unexpected, and marked the start of a beautiful collaboration between biologists and physicists. We combined approaches originating from both biology and physics to address a key problem of embryo development, in a truly two-way collaboration trying to overcome the artificial borders between fields of knowledge.
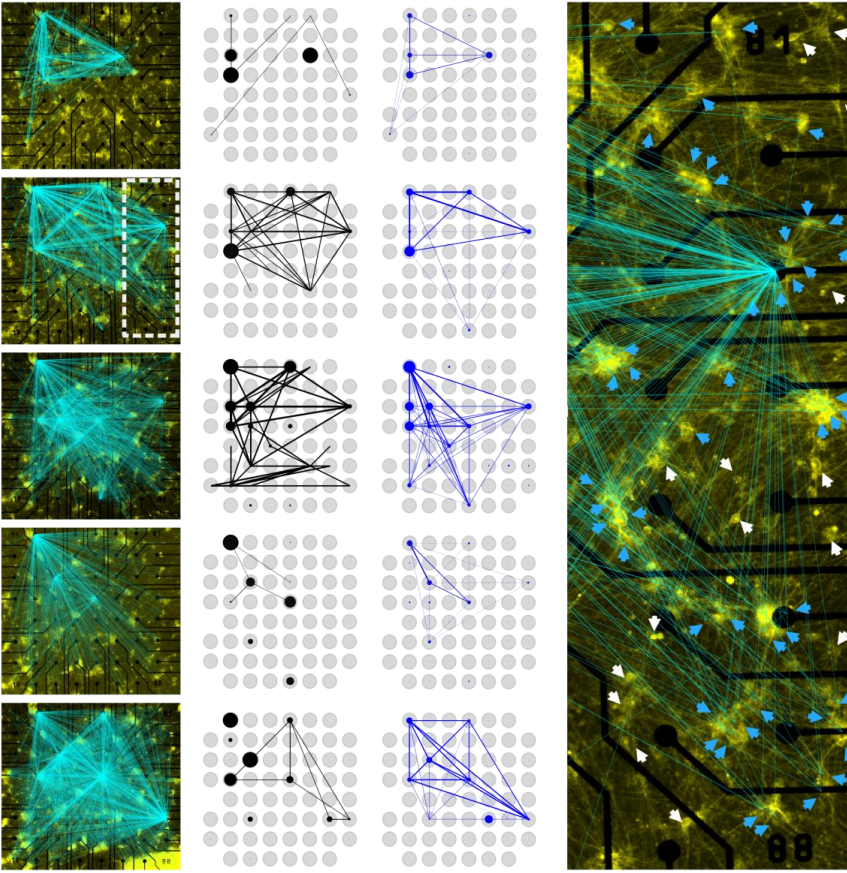
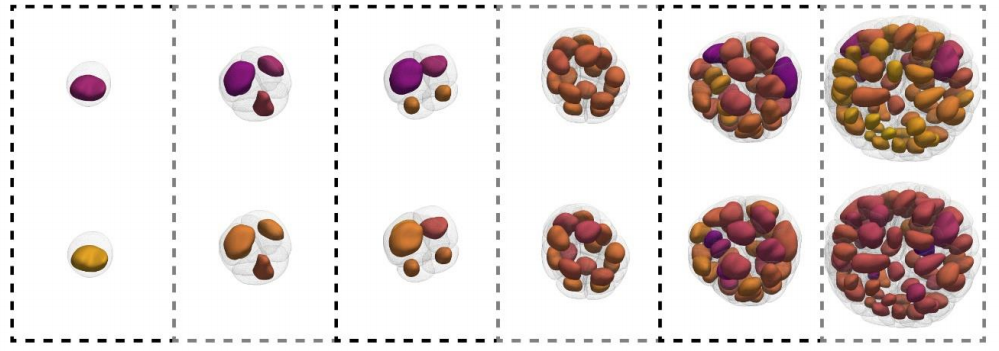
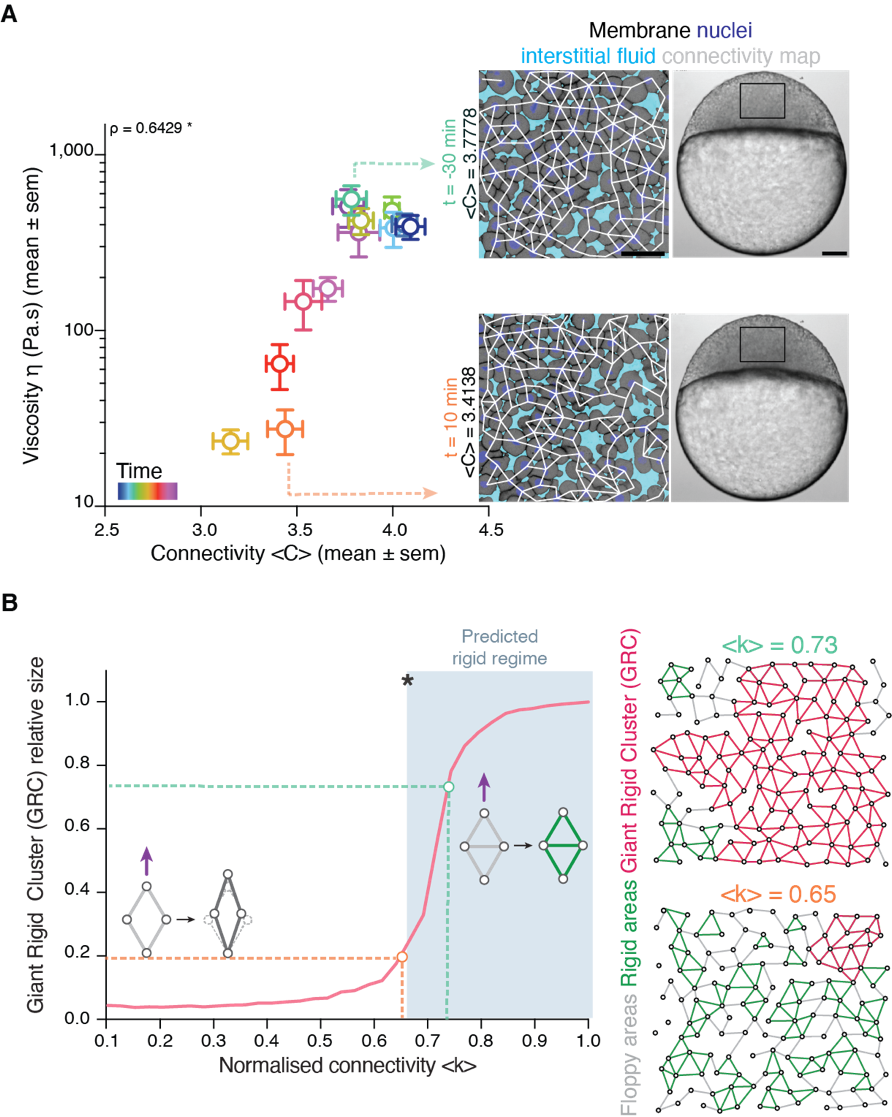
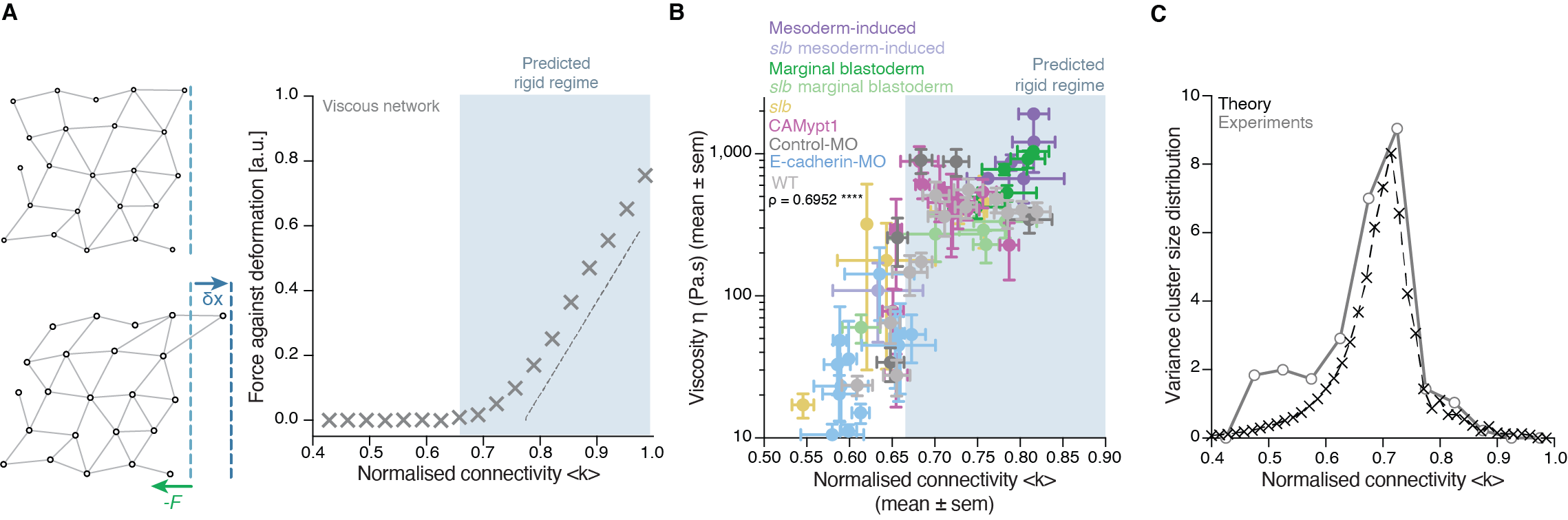
Back to IST, we started frantically analyzing embryo tissues as if they were a network made of cells (nodes) and links (the cell-cell contacts) between nodes (Fig. 1A). At a first sight, the results were not really encouraging: The networks of cell-cell contacts were not that different before and after the fluidization event. What could be reported was just a small, smooth drop in the average number of contacts per cell coinciding with the fluidization point (Fig. 1A). However, when dealing with phase transitions, one has to be careful not to neglect the relevance of small changes in some parameters of the system: close to the critical point (the point where the system changes its phase), any tiny fluctuation can result in an abrupt, macroscopic change. For example: if we leave a glass of water in the fridge at 0.1oC, it will remain forever liquid; if we drop the temperature an almost imperceptible 0.2oC, therefore putting the fridge at -0.1oC, the water in the glass will sooner or later freeze, which is a radically different state than the one we had when we were at 0.1oC. And somehow this was similar to what was happening in the embryo: the tiny decay in the number of contacts per cell was precisely crossing a critical point: the critical point of rigidity. That is, in a network with a lower number of average contacts per cell, the system as a whole remains floppy, and one can deform it with almost no effort. Beyond the rigidity threshold, no independent movements of the cells are possible without applying a force over the system. In particular, a rigid cluster of nodes (called Giant Rigid Cluster – GRC) abruptly emerges, almost spanning the whole network, and thereby opposing resistance to any deformation (Fig. 1B). The critical point in rigidity (also called the “rigidity percolation threshold” or “isostatic point”) is found when the number of average contacts per cell is 4 (which corresponds to the 2/3 of the maximum potential connectivity of a network, that is, 6, if one considers a complete packing made of a large number of hexagonal-like cells) (Fig. 1B). Strikingly, this is one of the older, if not the oldest, prediction of a critical point, as the result goes down to Maxwell (Maxwell, 1870), who solved the problem in the middle of the XIX century.
Figure 1: Abrupt temporal changes in blastoderm viscosity resembling a network rigidity phase transition. (A) (left) Plot of experimentally measured blastoderm viscosity as a function of cell connectivity over time (color-coded) and (right) two exemplary confocal images of the zebrafish blastoderm before (t=-30min) and just after (t=10min) fluidization displaying very similar connectivity but very different tissue viscosity. (B) (left) Plot of the size of the Giant Rigid Cluster (GRC) as a function of connectivity and (right) two simulated networks with connectivity slightly above (up) and below (bottom) the isostatic/critical point of Maxwell rigidity transition exhibiting a big difference in the GRC size. From Petridou, Corominas-Murtra, Heisenberg and Hannezo, 2021.
It turned out that, indeed, fluidization occurred at the moment where the average number of contacts per cell went below the critical point of 2/3 of the maximum cell connectivity of the tissue (Fig.2). But there was more: after that, the number of average contacts per cell increased very slowly, but crossing the critical point again to start “thickening” and recovering its viscosity/rigidity (Fig.2). The incredible correspondence of experiments to the theory of network rigidity percolation and viscosity by solely one simple criterion, cell connectivity, triggered one of these paradoxical moments in science where the excitement of the discovery is immediately followed by an uncomfortable feeling of uncertainty due to the overwhelming number of new questions that the discovery implies.
Figure 2: Rigidity phase transition during embryo morphogenesis. (A) Exemplary blastoderm confocal sections of the central blastoderm with overlaid connectivity maps (top) and their rigidity profile (bottom) at consecutive time points during the fluidization/thickening process (color-coded) showing the abrupt reduction of the size of the GRC and its gradual recovery. (B) Plot of blastoderm viscosity as a function of normalized connectivity <k> and integrated plot illustrating the time-trajectory (color-coded) of the central blastoderm material phase state (relative size of GC) as a function of its connectivity (critical connectivity point, kc). From Petridou, Corominas-Murtra, Heisenberg and Hannezo, 2021.
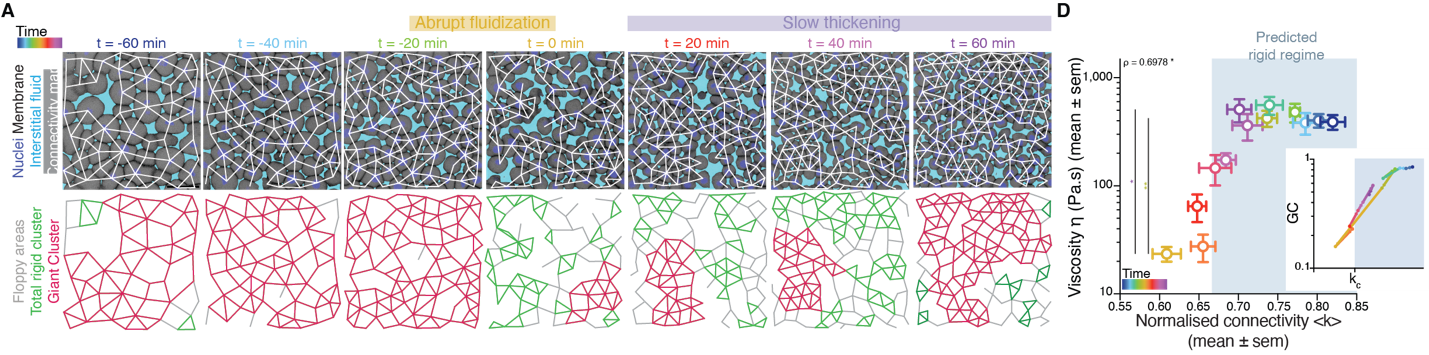
There was, first, a theoretical problem: We were measuring tissue viscosity, but the theory is about topological properties of the networks. Intuitively, they seem related, but we wanted a rigorous connection, as it is the key tie between theory —and its predictions— and experiments —with its observations. This turned out to be an interesting problem of bridging description scales: on the one side, we have a microscopic property, namely, the number of connections per cell, and on the other side, we had a macroscopic property, namely, the viscosity, hypothetically emerging from the microscopic structure. With the help of computer simulations and theoretical arguments, we could prove that, indeed, the critical point in connectivity for rigidity transitions triggers a qualitative shift in the behaviour of viscous networks. From almost negligible force response values in the region where the network is floppy, a sudden, steady increase is observed when passing the rigidity percolation point (Fig. 3A). In addition, we wanted to test the generality of our approach. How reliable is connectivity as a control parameter for predicting the different viscosity regimes of the tissues? We experimentally measured cell connectivity, tissue rigidity (GRC size) and tissue viscosity in numerous experimental conditions. For example, we examined this in embryonic tissues with differences in cell-cell adhesion, contractility, cell fate, geometry, genetic backgrounds: remarkably, using only the connectivity value, we could reproducibly predict the rigidity regime of the tissues (Fig. 3B).
Once we had established a theoretical and experimental connection between connectivity rigidity and tissue viscosity, we wanted to know if indeed what we observed in the embryos corresponded to a genuine phase transition. This was a great challenge for us, and when we started sharing our results with the scientific community, many sceptical questions followed. What class of a PT is this? How can it be a true PT in such a small living system? Such criticism is indeed necessary when merging fields especially when there are no similar examples or guidelines on the right way to do it! To prove a phase transition in physics for example, it requires to find the traces of a singularity or divergence in a macroscopic observable in terms of a control parameter that, in turn, increases smoothly. In addition, in a high order phase transition —as it is the case of rigidity percolation—, one expects to observe characteristic phenomena, like power-law statistical patterns of the diverging observable. Having a large amount of experimental data allowed us to perform reliable statistics and unambiguously detect such patterns. In particular, in rigidity phase transitions a singularity/divergence is expected to occur at the critical point of the phase transition which corresponds to the variance in the size of the rigid clusters other than the GRC. We observed a clear diverging trend in the experimental data and well-defined traces of a power-law distribution in the size of rigid clusters (Fig. 3C), with exponents compatible with the theory. These findings suggested that the embryo is operating very close to a surprisingly well-defined critical point of phase transition or, in other words, that the embryo is at criticality.
Figure 3: Rigidity percolation transition as a proxy of tissue viscosity and in vivo hallmarks of criticality. (A) (left) Schematic diagram of the force response for a deformation induced by a small displacement of the edge layer of viscous 2D networks and (right) Plot of the force response as a function of normalized connectivity <k>, showing that the critical point of rigidity percolation transitions also defines deformability in viscous networks. (B) Plot of blastoderm viscosity as a function of normalized connectivity <k> for several experimental conditions showing that the connectivity critical point is a reliable control parameter to predict the rigidity regimes of embryonic tissues. (C) Plot of the variance of the distribution of rigid cluster sizes other than the GRC, as a function of their normalized connectivity <k>, in simulated networks of the same size as the average size of experimental networks (black) and in the experimental networks shown in (B), showing a diverging trend at the critical point, with good, parameter-free agreement between theory and experiment. From Petridou, Corominas-Murtra, Heisenberg and Hannezo, 2021.
Many puzzling questions were constantly opening up: Can we engineer the system to fine-tune its material properties based on its network representation? Can we transform this simple but “static” representation of rigidity into a dynamic model taking into consideration the time-scales of cell and tissue viscoelasticity? The finding that the tissue is poised at criticality also gave a twist to our research, because we realised that we not only have a framework to predict the tissue material state but we can start exploring questions regarding robustness and stability of the system. To address this question within the proper environment, we met many times enjoying the bitter and intense taste of the expresso at Coffee Pirates —our favourite coffee shop in Vienna. And we always came to the same, fundamental question: What is the benefit of a living system being poised at criticality? A system at criticality can change its state with almost no energy investment, thus criticality could be an effective strategy in this developmental stage. However, this comes with a cost: a system at criticality is highly unstable, and this is not what we experimentally observe. For example, the rigidity phase transition occurs only once in the embryo at the onset of its morphogenesis. How is the embryo precisely operating between rigid and floppy regimes when positioned so close to the critical point? Our results so far suggest that synchrony in the cell division may play a fundamental role. Cell connectivity in the early embryo is dominantly regulated by cell division, where cell-cell contacts between neighbouring cells disassemble during mitosis and reassemble during interphase. We also found evidences of a counterintuitive but interesting relation between spatio-temporal randomness in cell division patterns and temporally precise and spatially uniform changes in tissue structural and material properties (Petridou et al., 2021).
Well, this was the moment when we knew that we could not answer all the questions –at least, not within our postdoc period! We never thought that this interdisciplinary scientific retreat could lead to such a fruitful collaboration, especially since it took us a long time to find a common communication language between physics and biology. We spent hours for example discussing what rigidity means in physics or what adhesion means in biology, going back to basic textbooks! We are very grateful to both CP and Edouard for supporting this challenging project, and, now, equipped with this full package of unanswered questions, we are both looking forward to address experimentally and theoretically how tissue rigidity and criticality orchestrate embryo development in our very recently starting groups at EMBL Heidelberg (Nicoletta) and University of Graz (Bernat), respectively.
There are dozens more ‘behind the paper’ stories featuring the highs and lows and serendipities of research on our dedicated page. Does your paper have a story behind it? We’d love to hear it.
By Alejandro Aguilera Castrejon and Jacob H. Hanna
Understanding the developmental processes leading to the formation of tissues and organs represents one of the most fundamental questions in developmental and stem cell biology. In mammals, most of this process takes place after the embryo implants inside the maternal uterus. After implantation, mammalian embryos initiate the process of gastrulation, in which stem cells differentiate into the three germ-layers, and subsequently commence organ formation, transiting from a symmetrical ball of stem cells into an advanced embryo with defined head, tail, organs and limbs. The intrauterine confinement of developing embryos has limited the study of post-implantation embryogenesis, due to the inability to observe and manipulate living embryos at these stages. Further, the very small size of the early post-implanted embryo makes it extremely difficult to observe inside the uterus using techniques applied for monitoring development of older fetuses. Thus, for over a century, scientists have attempted to devise ex utero culture systems capable of supporting mammalian post-implantation embryo growth, and had very limited success.
The motivation for our group to initiate this project was brought by the lack of methods for continuous and robust culture of early post-implanted embryos until advanced organogenesis, while at the same time, we have been able to routinely culture mouse embryos in vitro through all stages of pre-implantation development for decades. It is also not feasible to transfer explanted post-implantation embryos back to the uterus as commonly done for pre-implantation embryos. More specifically, we embarked on this project following our success to get low contribution of human derived iPSC cells into cross-species chimeric mouse embryos for the first time (Gafni et al., 2013), as we wanted to follow the integration of human iPSC derived cells in post-implantation mouse embryos, and also try to improve it (Bayerl et al., 2020). Further, devising continuous in vitro culture systems for mammalian embryos during the period from pre-gastrulation until advanced organ formation would facilitate direct experimental investigation of key processes during post-implantation embryogenesis.
To the best of our knowledge, the earliest attempts to culture post-implanted rodent embryos started in the thirties by using rat plasma combined with embryo extract (Nicholas and Rudnick, 1934). Several research groups improved this technique over the years, by modifying different culturing parameters such as supplementing the embryo environment with different types of media and rotating the cultured embryos (New and Coppola, 1970; New, Coppola and Terry, 1973; P. P. Tam and Snow, 1980; Sadler and New, 1981; Rivera-Pérez, Jones and Tam, 2010; Piliszek, Kwon and Hadjantonakis, 2011). However, normal embryonic development in previous studies was inefficient, and limited to only brief periods of time after the post-gastrulation embryos were dissected out of the uterus (24-48 hours) (Sadler, 1979). Such short-term culture techniques do not allow to study embryogenesis comprehensively, regardless the age of the embryo isolated from the uterus, because most of the organs develop and mature over several days.
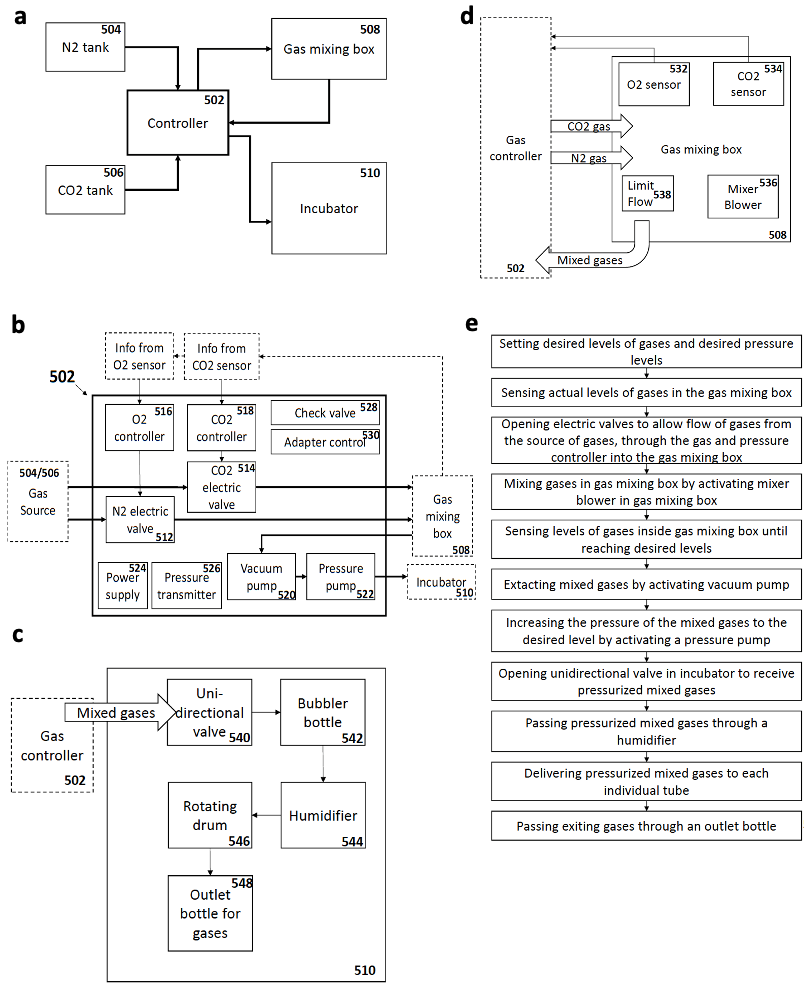
As an attempt to overcome these limitations, my supervisor Jacob Hanna thought about starting a project to revisit and re-evaluate the previously established short-term culture protocols and try to understand why they failed to obtain continuous embryogenesis ex utero. We reasoned that sensitive regulation of O2 and CO2 gas concentration is likely to be a critical parameter for embryogenesis. We also thought that controlling and increasing gas pressure could improve culture outcome and efficiency by increasing oxygen diffusion into the embryonic bloodstream, as routinely applied by lung ventilation machines used in hospitals (that also control gas pressure to enhance oxygen diffusion). However, there was no available system to control gas concentration and pressure precisely, so we engineered an electronic devise specially fitted for this purpose, which could be connected to the roller culture incubator (Fig. 1), and hired an external engineer to assemble it (Fig. 2a).
Fig. 1. a-e. Graphic schemes for the ex utero controller system developed and general design of different components.
Once the gas regulation module was designed, and after I joined the lab as a master student, we started testing for different combinations media, oxygen concentrations, and gas pressures that allowed embryo growth with the highest survival rate and for the longest period of time. We tested different combinations of sera from different species, synthetic sera, nutrients and other supplements which have been used for culture of stem cells and embryos. By these means, we established a platform that supports growth of late gastrulating embryos at E7.5 until the hindlimb formation stage E11 by culturing the embryos in rotating bottles at hyperbaric pressure (Fig. 2b).
Fig. 2. a. Electronic gas and pressure regulation module connected to the roller culture incubator system. b. Images of cultured embryos inside the bottles at the beginning of the roller culture (E7.5) and after 3 days (E10.5).
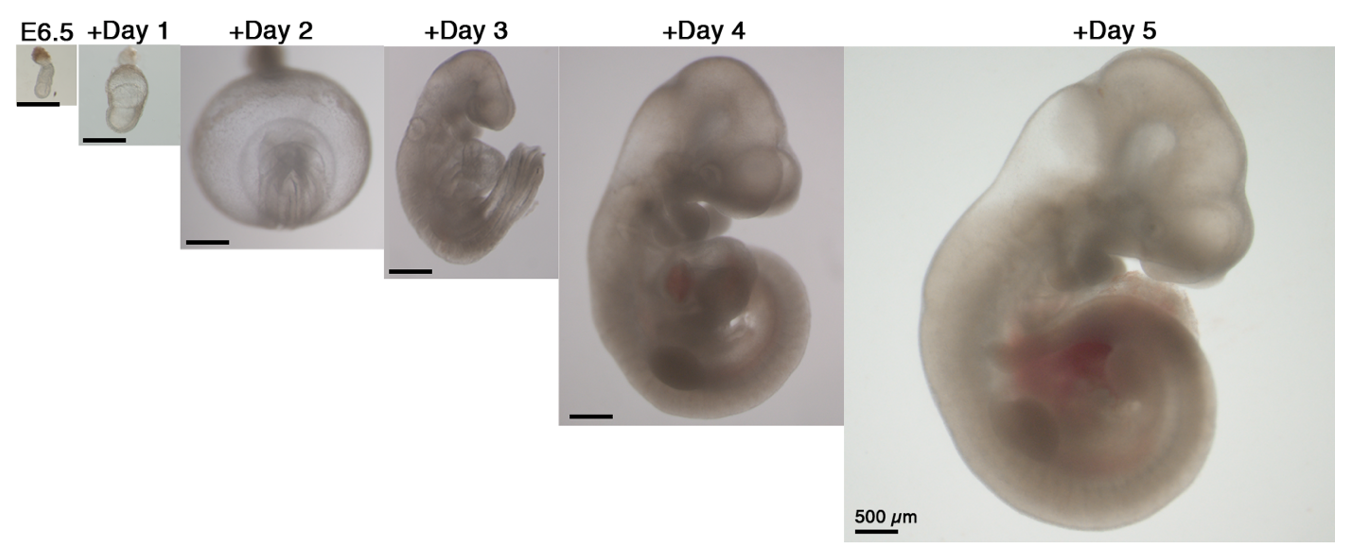
Our next step was to expand the ex utero culture platform one more day, aiming for starting with early gastrulating embryos at E6.5 until advanced organogenesis. However, no matter which conditions we tested, E6.5 embryos were not able to grow beyond two days using rotating bottle settings, possibly because E6.5 seemed very fragile when placed in rotating bottle conditions. At this stage we turned to devise cultures in static plates for early-gastrulation stage embryos, as we realized that there was not a well-established protocol for static culture of embryos at E6.5, since the culture conditions used per lab were variable, together with a disparity on efficiency and quality of embryo survival (P. P. L. Tam and Snow, 1980). We then decided to seek for conditions allowing robust development of embryos in static cultures from E6.5 until E8.5, since static culture does not support development beyond the early somite stage at E8.5. After testing diverse oxygen concentrations, gas pressures, types of sera, extracellular matrices and supplements we generated a protocol that allowed proper development of most explanted embryos. Remarkably, transfer of embryos cultured using our established static protocol to the roller culture allowed continuous growth from E6.5 to E11 (Fig. 3). Later, we realized that this static protocol is also suitable for growing embryos dissected at the earliest day of post-implantation development E5.5, and combined with the rotating bottles culture we were able to reach up to 6 days of mouse development ex utero.
Fig. 3. Proportional change in size of embryos developing ex utero from early gastrulation (E6.5) to the hindlimb formation stage (E11). Scale bars, 500 µm.
Notwithstanding, a limitation of the protocol was the dependence on freshly isolated human umbilical cord blood serum, which availability can be limiting. Thereby, we intended to find another type of sera which could replace it. We tested first using commercial human blood sera, but the results were not encouraging, possibly because often commercial serum productions is not done rapidly after blood collection, which leads to increased hemolysis by-products that can be toxic to embryos. Thus, we turned to isolating serum from fresh human adult blood in-house, which is relatively easier to obtain. Indeed, we were very happy to find that freshly isolated adult human serum was able to support mouse embryo growth, which makes the protocol much more available to the community.
A fundamental step was to perform a thorough examination of the embryos in order to verify that those developing ex utero mimic their counterparts growing inside the uterus. By means of morphological, histological, immunofluorescence, and single cell transcriptomic analyses, we validated that the embryosdeveloping ex utero are comparable to embryos growing inside the maternal uterus at the level of tissue architecture and cell composition. In particular, for the single cell RNA-seq analysis we chose to examine embryos cultured in the extended static and roller culture protocol, since embryos cultured from E7.5 in rotating bottles are cultured for a shorter time than those starting at E6.5. It was very important for us to conduct single cell RNA-seq based comparisons, since the latter can be viewed as a very stringent and unbiased way of comparing embryos and their organs.
Furthermore, one of the key advantages of our ex utero culture is the amenability of the embryos to physical, chemical and genetic manipulations, that can be followed for over up to six days of development. For this purpose, we looked at all those experimental techniques that have been available widely for pre-implantation embryos, and that we have always fantasized to apply for studying post-implantation development, for instance, gene targeting, high resolution imaging, or generation of chimeric embryos by cell transplantation. First, we wanted to demonstrate the ability to introduce genetic perturbations in the developing embryos, for which we employed two strategies: on the one hand we used whole-embryo electroporation, which is usually transient and localized to a specific region; on the other hand, we carried out lentiviral transduction to introduce DNA in a stable and widespread manner all over the embryonic and extraembryonic tissues. For live cell imaging, we aimed to show the ability to image in high resolution using confocal microscopy. We chose to image two of the most intricate processes of embryonic development: gastrulation and neural tube closure. The first process is perfectly suitable for imaging in static culture, and for the second one we wanted to show the possibility to image embryos that were initially grown using the roller culture system and then moved to static conditions for imaging under the microscope while they continue developing. Finally, we have always been fascinated by the ability of cells to integrate into developing embryos to generate chimeric animals; procedure frequently done in our lab by grafting mouse or human pluripotent stem cells into mouse blastocysts, which can further develop by transfer to a surrogate mother. Nevertheless, such an assay was not available to evaluate the integration of cells into post-implanted mammalian embryos. In this regard, by analyzing the integration of mouse and human cells into gastrulating embryos, we demonstrated that our platform offers a novel opportunity to assay long-term integration of cells into mouse embryos at post-implantation stages. Overall, we proved that we can perform a variety of embryo manipulations without affecting embryo development or culture efficiency.
In summary, by establishing platforms for continuously growing pre-gastrulation mouse embryos outside of the uterus from the embryonic day 5 to 11, our recently published study (Aguilera-Castrejon et al., 2021) provides a proof-of-concept for the ability to continuously capture mammalian gastrulation and advanced organogenesis in an artificial environment, and underscores the self-organizing ability of the embryo. This culture system may help uncover mechanisms of cell fate specification and organ formation in a mammalian model, as it alleviates the uterine barrier to allow dynamic experimentation in living mouse embryos. We hope that our ex utero culture will be adopted by many labs around the world, and that culturing mouse embryos in a tube for extended periods of time will become a commonly used technique as culturing cells or embryos from non-mammalian species. Another interesting possibility for this system would be to use it in combination with synthetic stem cell-derived embryo models (Harrison et al., 2017), for which this platform could provide an ultimate test to evaluate their resemblance to natural embryos.
Leaving aside the ethical implications that the ex utero culture may have for human embryos, by creating these platforms for culturing mouse embryos we intend to investigate the intricate processes shaping mammalian organs during embryogenesis, which in the future, may have a broad range of implications for human health, from understanding congenital disorders and cancer, to stem cell biology and tissue engineering.
Aguilera-Castrejon, A., Oldak, B., Shani, T. et al. Ex utero mouse embryogenesis from pre-gastrulation to late organogenesis. Nature (2021). https://doi.org/10.1038/s41586-021-03416-3
There are dozens more ‘behind the paper’ stories featuring the highs and lows and serendipities of research on our dedicated page. Does your paper have a story behind it? We’d love to hear it.
In the latest episode of Genetics Unzipped, Kat Arney takes a look at the genetics of giants and the science of small. Why do some species grow so large? What’s the genetic legacy behind Charles Byrne and the Giants of Ireland? And what was it about life on a Mediterranean island that miniaturised a mammoth?
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
 (No Ratings Yet)
(No Ratings Yet)









.jpg){kind=link}