The Node is a community site for developmental biologists, and is driven by content written by the community – once registered, you are free to post without requiring authorisation or approval. Today I want to share some ideas for how you could contribute to the Node, during the current COVID-19 pandemic and beyond (I recently circulated these ideas in our ‘Occasional Writing Ideas’ newsletter, which you can sign up for if you register for a Node account). Blogging might be the last thing on your mind at the moment, but the chance to write and connect with other researchers across the world could provide some welcome respite.
Here are some ideas for ways you can contribute to the Node:
Tales from isolation
How are you coping being away from the lab? Do you have any tips for other researchers? This could relate to science (e.g. online training resources) or personal wellbeing/mental health.
We would love to hear voices from across the world – pandemic diaries from Delhi to Delaware.
If you prefer speaking to writing, another option is a series of short video interviews with scientists in lock-down. Get in touch if you’d like to be involved.
Virtual meetings/symposia/seminar series
Organisers – which technologies and formats worked, which didn’t?
Attendees – how did it compare to being there in person? Is this the future of scientific conferencing?
Have you been part of one (as examiner or examinee)? What was the experience like? Any tips for those preparing for one?
Lablife
Which aspects of lab life have/haven’t you been able to replicate online?
Remote lab meetings – dos and don’ts.
PIs – any tips for maintaining lab spirit remotely?
Model organisms – how are they coping without you, and how are you coping without them?
Behind the paper stories
Have you recently published a paper or preprint? Tell us the story behind it – this could feature the personal side of scientific research or go deeper into the technical details than a paper allows
Catching up with the literature
Write a ‘Research Highlight’ on a paper that recently blew you away. A good chance to try out less formal scientific writing, and help promote the science you love.
Our Forgotten Classics series explored unjustly overlooked papers in the history of developmental biology – we’d love to hear about your Forgotten Classic.
Resources
Our Resources page contains content on advocacy, outreach, education, audiovisuals and research – have we missed anything useful, particularly for researchers at home? Get in touch.
Networking
If you’re missing out on networking in person, why not use and join the Node Network, our global directory of developmental biology and stem cell researchers. It’s designed to help those organising conferences, looking for referees and so on to identify individuals who might not otherwise come to mind. Maybe you’ll find your future collaborators there.
If you’re interested in writing anything for the Node, I’m happy to help at any point of the process, from sketching out an outline to editing drafts to helping with the final posting of the piece. I’d also love to hear ideas for different kinds of content not explored above – just email thenode@biologists.com.
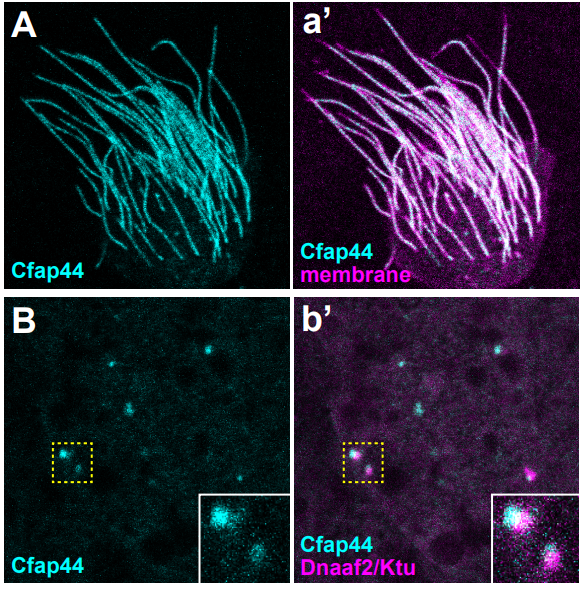
Our lab investigates the role of hydrostatic pressure as a morphogenetic force using zebrafish as a model system. This work was originally focused on tube formation and had revealed a crucial role for fluid secretion in single lumen formation (Navis and Bagnat, 2015). When I joined, the lab was conducting a forward genetic screen looking for phenotypes involving fluid regulation. We found many mutants with defects in the anterior-posterior (AP) axis and this brought our attention to the notochord. The notochord is the central structural element in the AP axis of all chordates and in vertebrates it serves as scaffold for the spine. In the developing vertebrate embryo, the notochord acts as a hydrostatic skeleton (Adams et al., 1990) and is important in the patterning of several tissues including the somites, blood vessels and pancreas. While the exact morphology of the notochord can differ between species, one key feature is highly conserved: the presence of large, fluid-filled vacuolated cells, also known as chordocytes (Grotmol et al., 2003). These cells are found in all vertebrate embryos and they persist beyond early development, being found in the nucleus pulposus of the intervertebral disks in mice, pigs, and humans (Lawson and Harfe, 2015). Their most salient feature is that the vast majority of their cell volume is occupied by a giant (~40µm) fluid-filled vacuole (Fig 1).
Fig 1. DIC and fluorescent image of a dissociated vacuolated cell from a 3 dpf zebrafish embryo. The cytoplasm is labeled with GFP surrounding a large central vacuole. Scale bar=50 µm
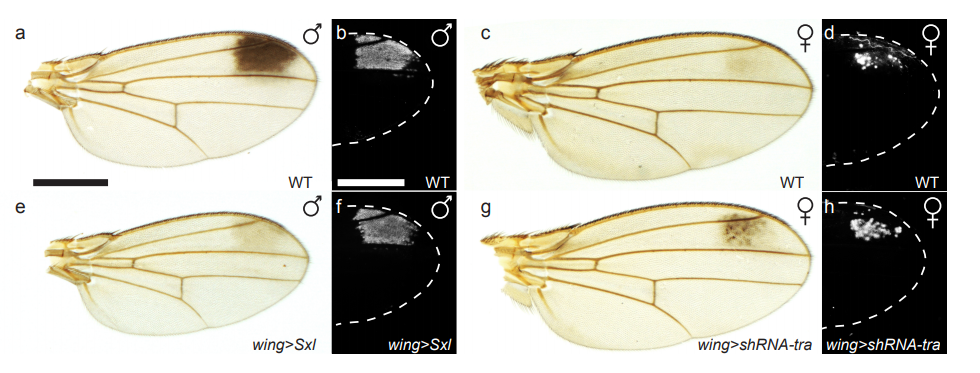
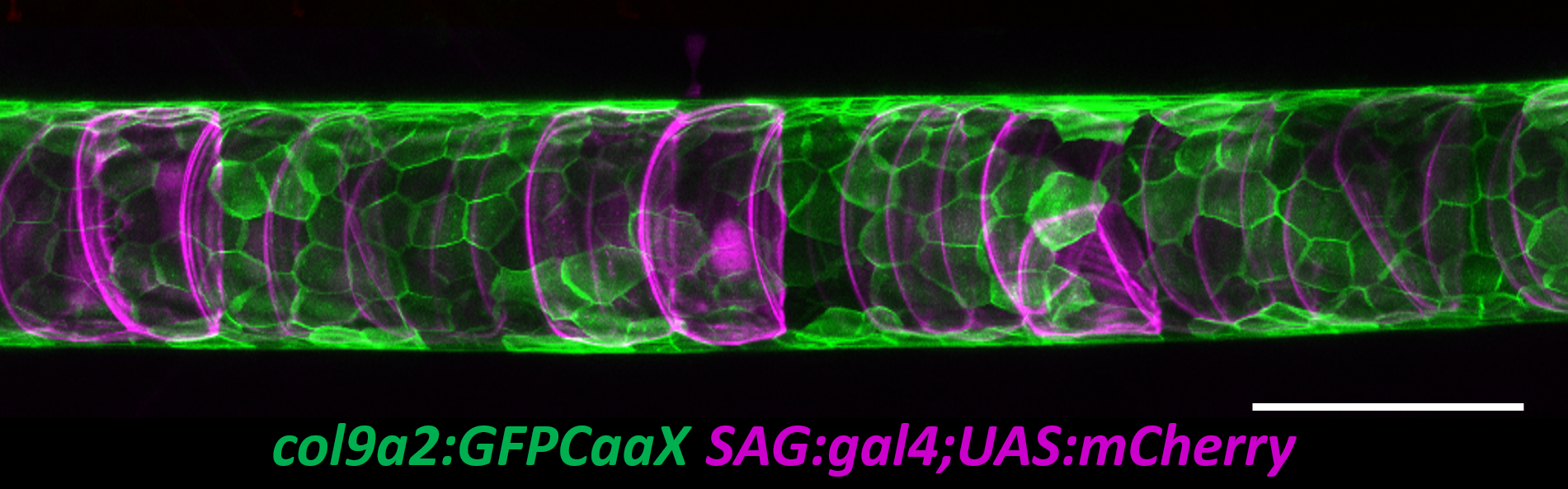
The zebrafish notochord is comprised of three parts: a core of large vacuolated cells; a layer of epithelial-like sheath cells that surrounds the core; and a thick extracellular matrix encasing the sheath cells. (Ellis et al., 2013b) (Fig 2.). When we started working on the notochord, we first wanted to define the identity of its unusual vacuoles. Combining a genetic and cell biological approach, we showed that notochord vacuoles are indeed lysosome related organelles which form via post-golgi biosynthetic trafficking and that they are crucial for embryonic axis elongation (Ellis et al., 2013a). Surprisingly, we also found that when vacuoles are disrupted during early development spine formation is also affected, causing a phenotype reminiscent of scoliosis in humans (Ellis et al., 2013a). Scoliosis is the most common developmental disorder of the spine and results in kinks and curvatures of the spines axis. Seeing spine malformations was a very exciting result for us and led us down a new road to understand how the notochord functions during spine morphogenesis.
Fig 2. Fluorescent maximum intensity projection of a live 5 dpf zebrafish notochord. Notochord sheath cells (green) are surrounded by vacuolated cells (magenta). Scale bar=100µm
To tackle this question, I started with a live imaging approach. We spent a lot of time generating transgenic lines to visualize the different cell types of the notochord and spine and learning how to image fish during late larval stages. While imaging the early embryo with confocal microscopy is relatively easy, imaging the notochord during spine development proved to be much more difficult. The tissue is thicker, external pigment develops, and it is much harder to keep the fish alive while on the scope. However, after a lot of trial and error we developed an imaging protocol that allowed us to image spine formation until about 6 weeks post fertilization. We also developed quantitative methods to determine the exact shape and position of every vacuolated cell in the notochord (Norman et al., 2018).
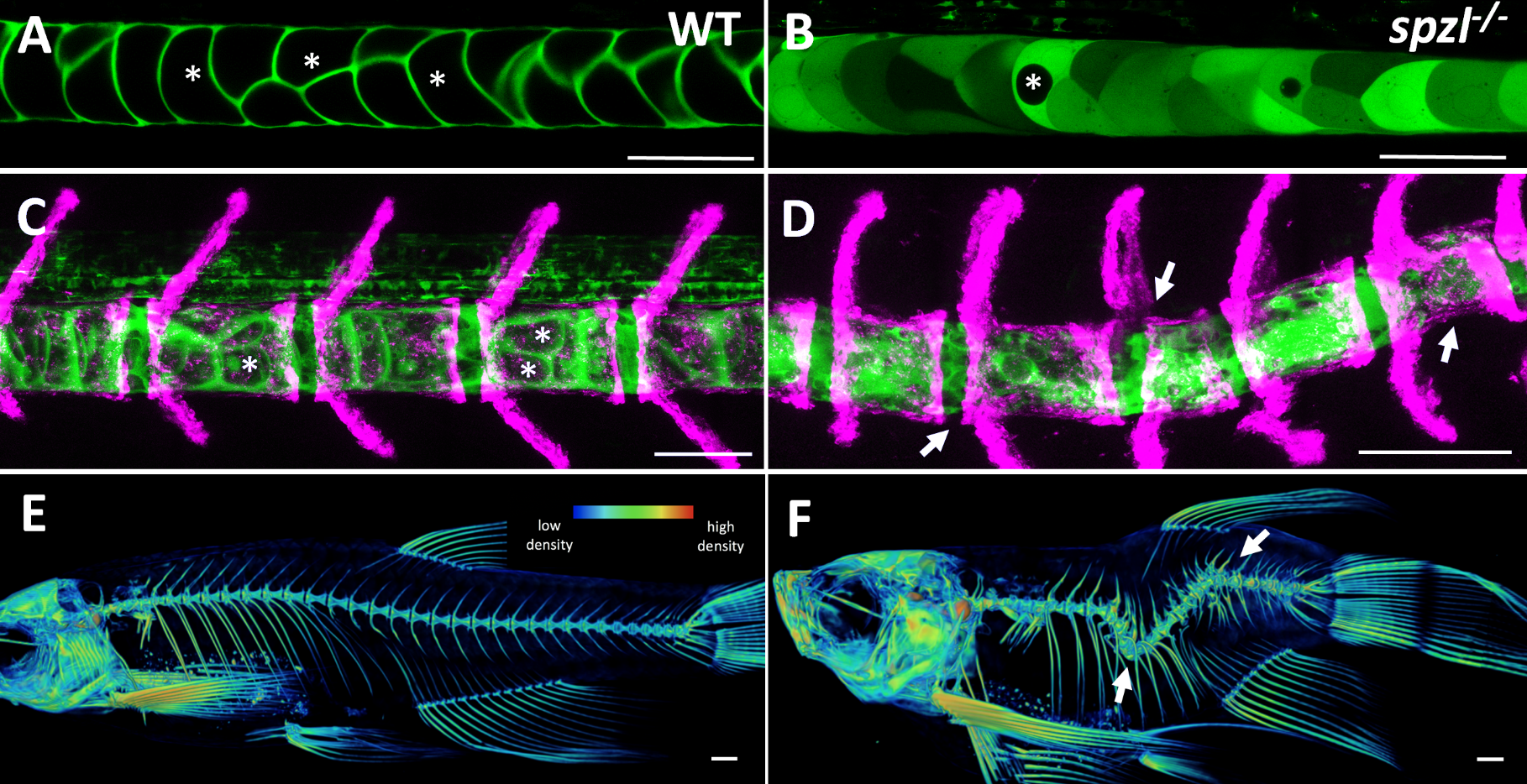
It soon became clear that vacuolated cells and their vacuoles persist through spine morphogenesis. Interestingly, we found that the morphology and arrangement of vacuolated cells changes during development. Vacuolated cells that remain under the growing vertebra change in shape from a rounded morphology to an elongated, pancake-like morphology; whereas vacuolated cells that end up in IVD (intervertebral disk) areas become tightly packed and their vacuoles fragment (Bagwell et al., 2020). It appeared as though the vacuolated cells are squished under the concentrically growing vertebrae, but are more rounded and clumped together in the IVD domains. This led us to hypothesize that the vacuoles are important in providing a scaffold that is able to initially resist the pressure of the growing vertebrae, symmetrically and consistently, allowing for the development of a straight spine (Fig. 3 A,C,E).
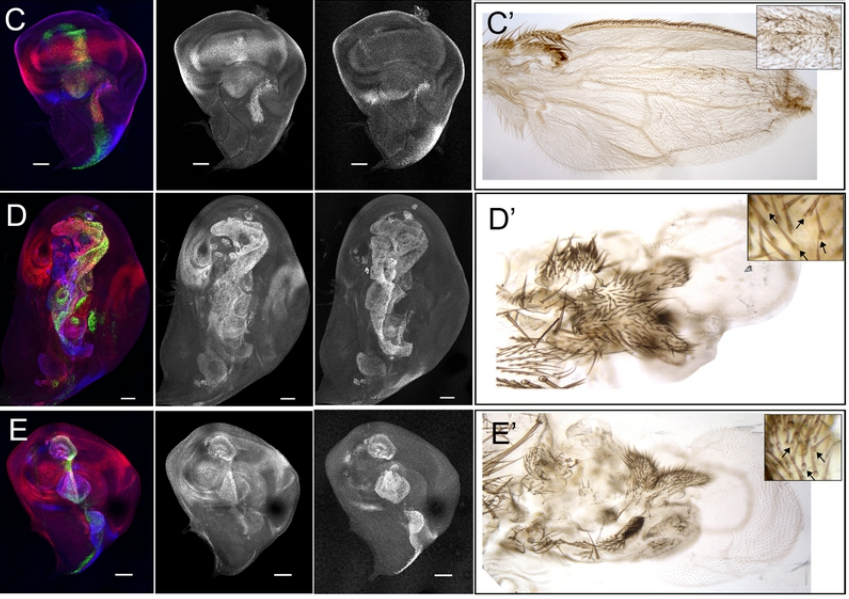
Fig 3. (A-B) Live confocal images of the zebrafish notochord prior to bone formation in WT (left) and spzl-/- (right). The cytoplasm of the vacuolated cells are labeled with SAG:gal4;UAS:GFP. Vacuoles are marked with an asterisk. (C-D) Live confocal image of the zebrafish notochord and spine at juvenile stages. The cytoplasm of the vacuolated cells is labeled with SAG:gal4;UAS:GFP and osteoblasts are labeled with osx:mcherry-NTR. Asterisks mark inflated vacuoles. Arrows indicate areas of spine kinks. Scale bars for A-D =100 µm. (E-F) Micro-CT heat map depicting relative bone density for 6 wpf WT (left) and spzl-/- (right). Arrows indicate spine kinks. Scale bar=1mm
At this point in the project, we were incredibly fortunate to collaborate with Didier Stanier, whose lab had carried out a forward genetic screen to identify mutants with defects in endodermal organs. They had found, by chance, a viable mutant with a shortened embryonic axis and what appeared to be a severe scoliosis phenotype. Because Didier knew we were working on understanding spine morphogenesis and the origins of scoliosis, he sent the mutant to our lab. This little mutant was so twisted and short, we affectionately named it spaetzle (spzl) after the German pasta (incidentally, Didier’s lab moved to Germany shortly after that). Right away we saw that the vacuoles in spzl were smaller and fragmented, and live imaging revealed that the vacuoles continued to deteriorate throughout development, leaving intact cells with really small or no vacuoles at all. We were surprised to find that the notochord and the AP axis were initially straight in spzl mutants. However, as vertebra formed the notochord was unevenly deformed, and severe kinking ensued as the bone grew (Bagwell et al., 2020). It was then clear that this mutant was a great genetic model to investigate the role of notochord vacuoles in spine formation.
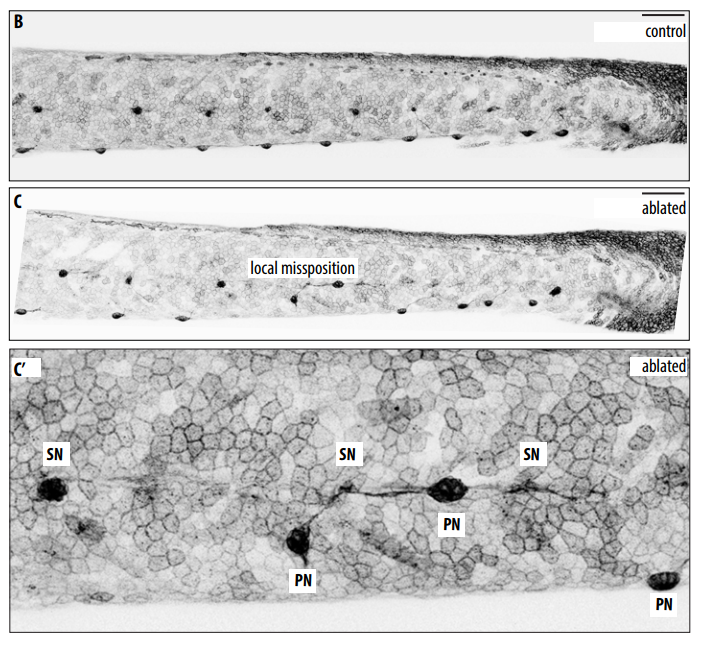
The first step for experimentally testing our ideas was to locally disrupt vacuoles to see if spine curvatures or kinks would occur at those specific locations. Simple right? There are many tools ablate cells including nitroreductase lines, photo ablation and physical injury, just to name a few. We had nitroreductase transgene perfect for this experiment and we could express it mosaically, kill a few cells and then see what happens with the spine. So I tried it successfully killing the cells, raised them to juvenile stages, and imaged their spines, but found no scoliosis. I was puzzled: how could we have such a strong phenotype in our spzl mutant, but be unable to reproduce the phenotype experimentally? Around this time Jamie Garcia, a graduate student, joined the lab and “Team Notochord”. She was looking at the role of cup shaped invaginations called caveolae, which the Parton lab had shown to be incredibly abundant in the notochord (Nixon et al., 2007). In parallel studies, Jamie and I on one hand (Garcia et al., 2017), and the Parton lab on their own (Lim et al., 2017), showed that upon locomotion, vacuolated cells lacking caveolae collapse, causing lesions in the notochord. We then found that as the vacuoles burst they release their contents, particularly ATP and UTP, which induce notochord sheath cells to locally invade the notochord core, transdifferentiate, and vacuolate, thus restoring the integrity of the notochord and allowing for a straight spine (Garcia et al., 2017). This finally explained why mutants lacking caveolae did not have a spine phenotype and why all the ablation assays I tried and tried did not work.
This ability of the notochord to regenerate vacuolated cells upon physical or mechanical injury reinvigorated our hypothesis that vacuolated cells are critical for spine development. I thought that if the notochord has the ability to fix itself, it must be important for later development. While vacuoles are lost in the spzl mutant, vacuolated cells remain intact and do not induce any sheath cell invasion or vacuolated cell regeneration. Through light sheet microscopy, I showed that spzl mutants are unable to undergo vesicle fusion to the vacuole membrane and are therefore unable to make and maintain a fully inflated vacuole, leaving a notochord that is lacking an even distribution of fully inflated vacuoles. Using nuclear deformation assay that James Norman and I developed, we also found that loss of vacuole integrity reduces the internal pressure of the notochord. Then, as vertebrae begin growing and squeezing the notochord, the structure deforms irregularly and is unable to absorb compression evenly. As a result, asymmetric deformations develop along the notochord, ultimately leading to severe kinking of the spine axis (Bagwell et al., 2020) (Fig 3 B,D,F). We were also able to recapitulate the mechanism with an experimental manipulation by expressing a dominant-negative form of the GTPase Rab32a. We showed that mosaic expression of DN-GFPRab32a causes vacuole fragmentation and local spine deformation at juvenile stages, further confirming our hypothesis. With the help of Sarah McMenamin’s lab we were also able to show using high-resolution micro CT imaging that loss of notochord vacuoles also leads to increased vertebral bone mineralization, suggesting notochord stiffness and vertebral bone formation are cross-regulated (Bagwell et al., 2020).
Our notochord journey started about 10 years ago. Our path began by asking two simple questions: what are notochord vacuoles and what are they good for? Like many questions in science this has led us down a road with lots of forks, turns and roadblocks. The forks in our path have led us to many new and exciting stories about the notochord such as its role in spine segmentation (Wopat et al., 2018), and there are many more to come. We have learned so much about this simple yet dynamic structure, but what is clear is that there is still so much more to learn. In the words of Yogi Berra
There is an open PhD position available in my lab. The project deals with the role of glial cells in post-ingestive nutrient sensing in Drosophila. Please see the advertisement for further information: Stellenausschreibung_Schirmeier.
Research into genetic conditions relies on information from patients and their families, whether that’s detailed health records or genomic data. As the tools and techniques for DNA and data analysis become cheaper and more organisations get in on this fast-growing field, it’s vital to make sure that the most valuable research resource – human lives – doesn’t get overlooked in the rush.
Fiona Copeland is the chair of a support group for UK families affected by primary ciliary dyskinesia or PCD – a rare genetic condition affecting the lungs – and is the mother of two adult sons with the condition. She explains what her role as a patient representative involves and shares her advice for how researchers can engage and involve patient groups more effectively. Her top tip? Don’t make children cry!
We also chat to Patrick Short,CEO of Sano Genetics – a Cambridge-based startup that aims to connect researchers with patients who want to take part in genomic research.
Finally, we hear from Shelley Simmonds, a disability rights campaigner and rare disease advocate whose son Fraser was initially given a diagnosis of Duchenne Muscular Dystrophy as a baby. When Fraser didn’t seem to be progressing as might be expected for a child with the disease, she and her family got involved in Genomics England’s 100,000 Genomes Project in search of clarity – but things turned out not to be quite so simple. Shelley talks what happens when the question “What’s wrong with my child?” has no answer.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Last week, I gave a talk (online, of course) about ‘Writing review articles’. It was aimed at graduate students who, as part of their training, had to identify a topic in the field of developmental biology and write a mini-review on that particular topic. However, my talk contained some general advice about writing review-type articles, as well as some general writing tips, so I thought I’d share a summary of it here.
Types of Review articles
I guess the first thing to point out is that review-type articles come in lots of different ‘flavours’. They all vary with regard to length, scope, style and overall purpose, and are given different names by different journals. But they all aim to summarise and distill research findings. This makes them very different to primary research articles, which aim to present data, although they are handled in similar way, i.e. they are submitted to a journal and peer-reviewed by 2-3 experts in the field.
The many names that journals use to label review-based articles
What’s the purpose of a (good) Review article?
A good review article might aim to:
summarise key research findings
highlight ‘must-read’ articles in the field
act as educational material
However, an excellent review article will also:
be timely
provide critique of studies
highlight areas of agreement as well as controversies and debates
point out gaps in knowledge and unanswered questions
highlight current technologies that are helping/can help the field
suggest directions for future research
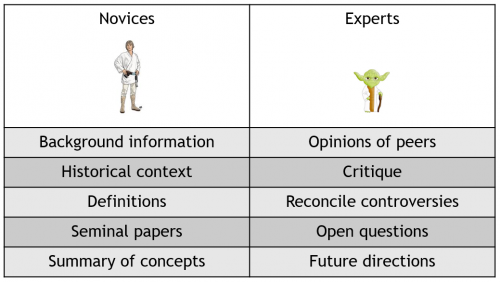
But remember that readers are usually a mix of experts and non-experts who will be looking for very different things so a good review will cater for both of these audiences. For example, a graduate student might turn to a review article when they start in a new lab to find out more about the history of a field, or to get a summary of key findings. By contrast, an experienced post-doc or PI might want to read a review written by one of their peers to find out what the current state of thinking in a field is. Ideally, a good review should therefore aim to provide a combination of balanced summaries and critique whilst being authoritative, forward-looking and inspirational. However, note that the exact ‘flavour’ or format of the review will also dictate its purpose, e.g. a ‘Perspective’ article in Journal X might aim to summarise a handful of recent studies, whereas an ‘Essay’ in Journal Y might aim to provide a more comprehensive analysis of the last decade of research.
The things that different types of readers look for in review articles
Where to begin?
The first step is to choose the topic you want to write on and come up with a rough idea of the scope of your article. You may already have this in mind but it’s important, before you begin writing, to really nail the exact purpose of your article. To help you do this, I‘d suggest the following:
Identify the particular theme/topic/idea that you want to focus on. In most cases, this will be something that’s closely related to the topic you work on, e.g. you might be working on something, or reading up on a particular area, and feel that a review would be helpful. If you need inspiration (i.e. if you want to write but aren’t sure what to write about), read, speak to people, and think about talks you’ve been to. What’s exciting in your field right now? Are there papers that change the way we think about something? Have you seen/read papers that converge on a similar theme/idea?
Check that there aren’t already reviews on this topic, i.e. something that’s been published within the past year or so. This is important; no-one wants to read a review that doesn’t offer anything new.
Decide if there is enough recent material to include (or too much). At this point, you may need to go back to the drawing board to either expand on or refine the scope of your article. It’s also helpful to read a few reviews (mini-reviews vs longer reviews) to get a feel for how much material a review can cover.
Identify and write down the main aim/purpose of your article. What’s the key message you want to get across? Why is this important and timely? Why would people want to read your article?
Note that lots of reviews are commissioned, i.e. the author is invited to write by a journal/editor. So, if you know you want to write a review on a particular topic and have a pretty clear idea of what your review will cover, a good place to start is by contacting a journal to see if they’d consider it. This also then means that you’ll (hopefully) be working alongside an editor from the outset to develop and refine the scope of your article. You’ll also have your target audience, article format and word limit in mind while you’re writing so can tailor the review accordingly.
Before you begin writing
Plan, plan and plan some more! Having worked with authors on review-type articles for years now, I can’t stress this enough.
Think about the sections/sub-sections you might use. What material would you cover in each of these? What’s the message of each section? How can you link the sections?
Think about the key concepts/words/specialist terms that you need to introduce and define. Where, when and how should you introduce these? (e.g. in Intro, in a figure, in a text box). What needs to be introduced first? What’s the best order in which to discuss these?
Think about the display items (figures, text boxes, tables) that might be helpful. How/when should they be used? What material would they contain?
When you start writing
Once you have a plan, you can start writing. I’d suggest that you start with the Title, Abstract and Introduction – these are the first parts that the reader sees of the article so they need careful thought. By starting off with these, you’ll also have the scope/purpose of the article clear in your own mind. You can then work on the main text of the article (the ‘meaty’ bit) and the Conclusions with this scope/purpose in mind, although you’ll need to return to the Title, Abstract and Introduction for a tidy up once you’ve written the main text.
Things to think about:
Title, Abstract and Introduction: These should be short and self-contained, and should complement each other. Each one in turn should provide more detail, aiming to draw the reader in. Remember: lots of readers will only read the title and abstract (e.g. when they search for articles in Pubmed) so these basically act as a ‘hook’ to grab their attention. They also need to be ‘discoverable’ on the Web, i.e. database friendly and containing the relevant keywords.
Choosing a title: Choose something that is short, clear and self-explanatory; try to avoid puns/idioms and colloquial phrases or references. Try to convey the key message but also provide context.
Abstract: The abstract should then aim to highlight the most important parts of the article. The answers to the following 5 questions provide a good starting point: What is the main topic you’re going to focus on? What do we know so far? What is new/why is this now an interesting time for this field? What are the broad implications of these newer findings? What does your review aim to do?
Introduction: The Introduction should then expand on the Abstract and set the scene. Provide context by first introducing the topic: why is this topic interesting/significant, what do we know about it so far, how has the field progressed, what has the new progress shown? Ideally, the Introduction should end with a clear description of the article’s scope, aims and structure, i.e. a walk-through of the main topics that will be discussed and the order in which these will be covered. This just lets the reader know what they can expect from the article. If possible, introduce or re-iterate the main ‘message’ of the article.
Conclusions: Emphasize the key message or theme of the article and, if needed, reiterate the data that support this message. Highlight the broader significance of this conclusion. Finally, if possible, bring your voice to the article: What do you think are the most compelling questions raised by these studies? What approach(es) could be taken to address these open questions? Are there technical hurdles that need to be overcome? What are the broader implications of this, i.e. why are further studies needed and what benefits might they offer?
Display items: Use figures to emphasize or illustrate key concepts/processes, or to introduce or summarize. Remember that figures should ideally act as stand-alone items; you should be able to follow them by eye and without referring to the main text, although each figure should have a clear title and a figure legend the walks the reader through the figure. In general, schematics are easier to follow than images reproduced from primary articles. Tables can be useful for summarizing lots of information, for comparing/contrasting things, or for highlighting advantages and disadvantages. Some journals encourage the use of text boxes, which can house additional or background information or material that is peripheral to the main theme of the text.
General things to think about while you’re writing (and to re-visit before you finish off!)
Structure
Try to group your discussion into sections/sub-sections. This just helps to break up long chunks of text (and helps to keep the reader interested). If you already have a plan (e.g. a list of headings/sub-headings) this structuring will be much easier.
Each section should begin with a small introduction.
Each sub-section (and/or even each paragraph) should then have a clear message/point to it, e.g. What question did particular sets/types of studies set out to address? What did these show (and here you can go into the detail)? What could be concluded from these?
It’s also helpful to add in a few lines to wrap up each section and ease transition into the next section.
Content
Make sure that all statements are adequately supported by a citation. Cite the source/primary article whenever possible (but note that it is okay to cite Reviews for established concepts or to refer to a large body of evidence).
Think about the word count and how much can be covered/how much detail you can go in to; you may find that it’s easier to write lots first then trim at a later stage.
Avoid regurgitating the conclusions drawn in the papers you cite without giving them some thought.
Don’t shy away from discussing findings that contradict each other. It’s better to highlight what can/cannot be reconciled and the possible cause of any discrepancies. Also use this as an opportunity to draw out the questions that remain and discuss how these questions could be addressed.
Similarly, remain balanced – make sure you discuss the findings from the field as a whole (and not just the data from a few select labs).
Make it clear when you are stating results versus providing speculation or alternative interpretations.
Provide critique if you can…but keep it polite and constructive.
Accessibility
Remember your audience: the article needs to accessible to expert and non-expert readers alike.
Introduce/define/explain specialist terms, cell types, tissues, phrases on first mention.
Consider using display items to house any material that a non-expert reader might find useful.
Don’t assume the reader knows what you’re thinking and how things link together; you might feel like you’re sometimes stating the obvious but it’s better to do this than to leave readers feeling lost.
Style
Stick to using clear and simple sentences…but try to vary the pace of your writing, e.g. by using a mixture of long and short sentences.
A general rule is to write as you would speak, using active rather than passive tense/sentence construction.
Be thrifty with your words: completely eliminate any that aren’t needed.
Avoid vague sentences. For example, say ‘Factor A causes an increase/decrease in Factor B’, rather than ‘Factor A modulates Factor B’.
Importantly, be patient and don’t get frustrated! A good writing style needs to be developed over time and comes with practice. Of all the things highlighted above (structure, content, accessibility and style), I’d say that style is the hardest to really nail. Getting a good and consistent writing style is also challenging if you have multiple authors working on the same article. In this case, I’d recommend that you nominate one author to do a final comb-through to iron out any inconsistencies, although hopefully you’ll have an editor who’ll also assist with this! On this note, I should point out that the amount of input you receive from an editor will vary from journal to journal, e.g. some journals have dedicated editors who spend a significant amount of time, working alongside the authors, to edit and improve a review.
Developing your writing style
Finally, some tips from fellow editors!
We have a bunch of experienced editors here at the Company of Biologists so I asked them all for their key pieces of advice. Here are just some of the things they suggested:
Plan, plan, plan – make sure you have a good idea of the overall structure before you think about details
Get feedback. Before you submit your review, send it to someone whose opinion you trust and ask them for their honest thoughts. Don’t be discouraged if they give lots of feedback – this is exactly what you want!
A review shouldn’t just be a list of facts, e.g. X showed this, Y showed this, Z showed this. A narrative thread or argument that connects is much more engaging.
Take time to pull back and look at the overall structure. Does it make sense? Can you see how the ideas join together and flow from beginning to end?
Remember that readers aren’t psychic. Explain why you’ve chosen the scope you have, why you’ve chosen to discuss particular examples, why you’re moving on to the next topic. Also make sure you clearly link up relevant observations and state conclusions rather than expecting the reader to make connections.
Don’t assume that the reader can link two statements that you might be able to link in your mind; you have to explain the link.
Think about the graphics at an early stage – figures can often feel like a bit of an afterthought but good figures can really help to get the message across far more concisely than text.
Break the article up into sections so that people can easily find the particular piece of information they might be looking for; recognize that not everyone is going to read from start to finish.
Remember that your readers will know far less about the topic than you do. So before you dive into the new and exciting findings in the field, make sure you’ve given a clear overview of the system you’re writing about. Imagine that you’re writing for a new PhD student who’s never worked in this particular field.
One final point: there’s no ‘winning formula’. This is just my advice based on the articles I’ve handled and the authors I’ve dealt with, so you may find that some of it doesn’t work for you or that someone else’s advice differs. Ultimately, you should aim to develop a writing approach, technique and style that works for you.
PhD position in leukemic stem cells and metabolism
A PhD position is available in the laboratory of ‘’Stem Cells and Leukemia’’ at the University Clinic of Hematology & Central Hematology, Department of Biomedical Research (DBMR), Inselspital Bern, Switzerland. We are looking for a highly motivated and creative scientist with a master’s or MD degree to contribute to research projects aiming to explore leukemic stem cell dependencies and vulnerabilities using transgenic mouse models of myeloid malignancies and primary patient samples from leukemia patients. The employment starting date is 01.08.2020 (or negotiable).
Your profile
The applicant should have a master’s/diploma or MD degree. The potential candidate must be experienced in working with transgenic mouse models, and multicolor flow cytometry. Expertise in working with stem cell cultures, CRISPR/Cas9 gene manipulation techniques, and willingness to work with the primary patient samples are preferred. Demonstrated competence to independently conduct experiments and analyses, and willingness to collaborate internally and externally is mandatory. The applicant should have an excellent command of spoken and written English.
We offer you
The post-holder will be involved in well-funded interdisciplinary research projects in a young and dynamic environment. Our team works closely with clinicians and pharma industries in Switzerland and abroad. The Department of Biomedical Research (DBMR) at the University of Bern and Inselspital Bern provides a stimulating environment and offers all the possibilities to be creative and highly productive. Doctoral thesis associated with an excellent qualification program. The PhD student will be enrolled in the interfaculty Graduate School for Cellular and Biomedical Sciences at the University of Bern (www.gcb.unibe.ch). The position is limited to 3 years with a possibility of extension. The salary will be according to the guidelines of the University of Bern.
Application / Contact
To apply for this position please submit your application in a single Pdf file including 1-page motivation letter specifying your research experience and interests and the desired start date, CV, summary of master thesis (max. one page), copies of master’s/diploma degree or MD degree, and the names and contact details of 2-3 referees to: tata.nageswararao@dbmr.unibe.ch
PhD position in leukemic stem cells and metabolism
A PhD position is available in the laboratory of ‘’Stem Cells and Leukemia’’ at the University Clinic of Hematology & Central Hematology, Department of Biomedical Research (DBMR), Inselspital Bern, Switzerland. We are looking for a highly motivated and creative scientist with a master’s or MD degree to contribute to research projects aiming to explore leukemic stem cell dependencies and vulnerabilities using transgenic mouse models of myeloid malignancies and primary patient samples from leukemia patients. The employment starting date is 01.08.2020 (or negotiable).
Your profile
The applicant should have a master’s/diploma or MD degree. The potential candidate must be experienced in working with transgenic mouse models, and multicolor flow cytometry. Expertise in working with stem cell cultures, CRISPR/Cas9 gene manipulation techniques, and willingness to work with the primary patient samples are preferred. Demonstrated competence to independently conduct experiments and analyses, and willingness to collaborate internally and externally is mandatory. Applicant should have an excellent command of spoken and written English.
We offer you
The post-holder will be supported by a well-funded interdisciplinary research project in a young and dynamic environment. Our team works closely with clinicians and pharma industries in Switzerland and abroad. Department of Biomedical Research (DBMR) at the University of Bern and Inselspital Bern provides a stimulating environment and offers all the possibilities to be creative and highly productive. Doctoral thesis associated with an excellent qualification program. The PhD student will be enrolled in the interfaculty Graduate School for Cellular and Biomedical Sciences at the University of Bern (www.gcb.unibe.ch). The position is limited to 3 years with a possibility of extension. The salary will be according to the guidelines of the University of Bern.
Application / Contact
To apply for this position please submit your application in a single Pdf file including 1-page motivation letter specifying your research experience and interests, CV, summary of master thesis (max.one page), copies of master’s/diploma degree or MD degree, desired start date, and the names and contact details of 2-3 referees to: tata.nageswararao@dbmr.unibe.ch
Meeting report: 2nd EMBO workshop “Awakening of the genome: the maternal-to-zygotic transition”
Amanda Amodeo1*, Johanna Gassler2* and Ksenia Kuznetsova3*, Philip Zegerman4
*These authors contributed equally
Lewis-Sigler Institute for Integrative Genomics. Carl Icahn Laboratory Washington Rd., Princeton University. Princeton, NJ 08544
Institute of Molecular Biotechnology of the Austrian Academy of Sciences (IMBA), Vienna Biocenter (VBC), Dr. Bohr-Gasse 3, 1030 Vienna, Austria
Max Planck Institute of Molecular Cell Biology and Genetics Pfotenhauerstrasse 108, 01307 Dresden, Germany
Wellcome Trust/Cancer Research UK Gurdon Institute and Department of Biochemistry, The Henry Wellcome Building of Cancer and Developmental Biology, University of Cambridge CB2 1QN, UK
Abstract
The fusion of gametes to form a zygote triggers the complex journey of embryogenesis. The first zygotic divisions are driven largely by maternal gene products, pre-loaded in the oocyte. One of the first steps in embryogenesis is the transition from the expression of the maternal genome to the expression of the new genome of the zygote – the maternal to zygotic transition (MZT). This transition is essential for the correct patterning and differentiation of the embryo and encompasses dramatic changes in genome organisation, chromatin state, cell cycle length and RNA stability. The MZT therefore offers a unique window to understand how fundamental aspects of cell and developmental biology are coordinated. Here we report highlights of the 2019 EMBO workshop “Awakening of the genome: the maternal-to-zygotic transition”, a unique synergy of insights from multiple disciplines and model systems into early metazoan development.
In the beginning……
The transition from the oocyte, a highly differentiated and arrested cell type, to a fertilised, developmentally active, totipotent cell requires dramatic changes in almost all aspects of cell physiology. Across species, an influx of calcium ions is required for egg activation and preparation of the oocyte for embryogenesis. One role of calcium ions is to trigger the activation of the phosphatase calcineurin, but how calcinuerin triggers multiple events during oocyte maturation is poorly understood. Using Drosophila oocyte activation as a model, Mariana Wolfner (Cornell University, US) presented a proteomic analysis of the changes in the phospho-proteome that occur in a calcineurin-dependent manner. The hundreds of proteins regulated by calcinuerin 1 provide a unique dataset to understand how the many changes that must occur during oocyte maturation are coordinated. One example of this is the critical translation regulator GNU, which, as described by Emir Avilés Pagán (Orr-Weaver lab, MIT, US), is phosphorylated and inhibited in arrested Drosophila oocytes by CDK, but gets dephosphorylated by calcineurin leading to activation of the GNU-binding kinase PNG. PNG is critical to trigger mRNA translation in the embryo by phosphorylating and inhibiting translational repressors, such as TRAL (Trailer Hitch), PUM (Pumilio) and BICC (Bicaudal-C) 2. This phosphorylation/dephosphorylation switch at the oocyte to embryo transition is a common feature among metazoa. Indeed Swathi Arur (MD Anderson Cancer Center, University of Texas, US) showed that in C. elegans the miRNA processing enzyme Dicer is inhibited by phosphorylation in the oocyte, but gets dephosphorylated after egg activation. This regulation is important to impede degradation of maternal RNA in the oocyte. It is clear that switching the phosphorylation state of many key proteins at the oocyte to embryo stage permits the rapid and wholesale changes in cellular physiology that must occur at this transition.
So long mother, and thanks for all the mRNA.
Oocytes are loaded with mRNA encoding between one third (mouse, C.elegans) to three quarters (Drosophila, sea urchins, fish) of all protein coding genes 3. Following egg activation, subsets of the maternal mRNA stores are degraded, first by maternally contributed factors (early mRNA decay) and then by zygotically expressed factors (late decay). The timing and order of these degradation events is critical for the onset of zygotic transcription and proper embryogenesis.
Several processes control mRNA stability including RNA binding proteins (RBPs), polyA tail length, miRNAs and RNA modification. In mice, mRNA decay begins already in the oocyte. Heng Yu Fan (Zhejiang University, China) demonstrated that the mRNA deadenylation machinery comprising of the CCR4-NOT complex together with the adaptor protein BTG4 are important for degradation of mRNA in the mouse oocyte. An additional pathway involving terminal uridylation of mRNAs by the TUT4/TUT7 enzymes also promotes maternal mRNA clearance both in the oocyte and in the embryo 4.
In Drosophila, RNA binding proteins play key roles in the timing of mRNA degradation and translation. Large scale analysis of ribonucleoprotein complexes in Drosophila presented by Craig Smibert (University of Toronto, Canada) suggested that a high affinity binding site of an mRNA towards a repressor, such as Pumilio would induce early mRNA decay, whereas degradation in later stage embryos required Pumilio-mRNA stabilisation by other factors, such as zygotically expressed miRNAs. These mechanisms could explain the temporal regulation of different transcripts in spite of the presence of Pumilio throughout early embryogenesis. Such a temporal order is also enforced by post-translational regulation of the RNA binding proteins themselves. Olivia Rissland (University of Colorado, US) described a screen in Drosophila for factors that degrade a key global mRNA regulatory complex, ME31B/Cup/TRAL during the MZT. These data provided evidence that differential degradation of RBPs might influence the timing and mechanisms by which transcripts are inhibited during embryogenesis.
In addition to mRNA stability, mRNA translation is also a critical regulatory step for gene expression at the MZT. Mike Sheets (University of Wisconsin, US) showed that Bicc1 (Bicaudal-C) in Xenopus laevis inhibits mRNA translation through direct binding to the mRNA 5. As spatial gradients of Bicaudal-C are established during oogenesis, this likely explains the maternal role of Bicc1 in determining anterior-posterior patterning. Despite this, not all Bicc1 target mRNAs are inhibited to the same extent, suggesting that other properties of the mRNA, such as translation efficiency may also be important. In line with this, Ariel Bazzini (Stowers Institute, US) showed that the best predictor of the half-life of an mRNA in human cells 6 as well as in zebrafish embryos 7 is its codon usage.
The plethora of pathways that exist to control mRNA stability and expression in the early embryo, likely reflects the importance of the exact timing and amount of protein production during development. However, using quantitative proteomics in mouse and Xenopus embryos respectively, Michele Boiani (MPI Munster, Germany) and Martin Wühr (Princeton University, US) showed that proteins remain relatively constant until the morula stage in mice 8 or the MBT (mid-blastula transition) in frogs. By separating nuclear from cytoplasmic protein, Martin Wühr showed in Xenopus that the nuclear fraction of the proteome changes during early development, which may explain some of the ordering of nuclear events during embryogenesis such as PolII versus PolIII transcription. Together with the multitude of pathways that regulate maternal mRNA, this suggests that post-translational regulation of maternal protein, including protein localization and modifications, must also be critical during the early embryonic cycles.
OK Child, over to you…. Setting up the transcriptional state of the zygote
The transition from maternal to zygotic control of the developing embryo requires not only the erasure of maternal RNA and proteins, but the activation of zygotic transcription. The reprogramming of parental DNA, combined with the correct timing of gene expression from the zygotic DNA requires major chromatin and transcriptional remodelling.
Nuclear organization of early pre-ZGA embryos is characterized by the absence of heterochromatin and the de novo acquisition of histone modifications and DNA methylation. Despite this, many chromatin-modifying enzymes, such as H3K9 methyltransferases, are expressed before the ZGA, raising the possibility that there are maternally-deposited inhibitors of heterochromatinisation. Mary Goll (University of Georgia, USA) provided evidence of one such inhibitor, the chromatin remodeller Smarca2 in zebrafish 9. Removal of Smarca2 is critical for global heterochromatin formation and interestingly Smarca2 clearance is driven by zygotic transcription of the miRNA miR-430, which links chromatin formation to the ZGA itself. Analysis of gene specific chromatin marks in Xenopus however, (Gert Jan Veenstra, Radboud University, Netherlands) demonstrated that several modifications, such as H3 K4 and H3 K27 trimethylation, arise in the absence of transcription in the early embryo. Indeed in Drosophila, Nicola Iovino (MPI Freiburg, Germany) showed that maternally inherited H3 K27 methylation is critical for inhibiting premature transcription before the ZGA 10. A similar role was proposed by Shifeng Xue (A Star, Singapore) for the chromosomal protein Smchd1 in zebrafish, whereby maternally supplied Smchd1 regulates DNA methylation and repression of embryonic genes. Therefore, both maternal and zygotic factors are important for determining the establishment of chromatin modifications and zygotic transcription.
Although the chromatin modification and DNA methylation landscape are remodelled in the early embryo, a key question is how this is directed to ensure that the right genes are turned on at the right time. Brad Cairns (University of Utah, USA) presented work in Zebrafish showing that key nucleosomes established before ZGA act as “placeholders” to determine the hypomethylated regions of the genome 11. At the ZGA these “placeholder” nucleosomes gain bivalency (the ability to switch between active and repressed epigenetic states) and are key determinants for turning housekeeping genes on, but developmental genes off. A critical feature of the placeholder nucleosomes is that they contain a histone variant (H2AZ). Genevieve Almouzni (Institut Curie, France) and Amanda Amodeo (Princeton University, USA) investigated the role of a different histone variant H3.3 during the ZGA in Xenopus and Drosophila. Amodeo demonstrated the large scale replacement of H3 by H3.3 on chromatin 12, although the functional significance of variant histone incorporation is poorly understood in Drosophila. However in Xenopus, Almouzni showed that the S31 residue which is specific to the H3.3 variant is uniquely essential for normal development through gastrulation 13. Together these results show an important role for histone variants in marking sites of future transcription and allowing normal developmental progression.
All hail the pioneers
The dramatic changes in chromatin during the ZGA are also accompanied by changes in transcription factor (TF) interactions with DNA. Whether chromatin modifications are a cause or a consequence of TF recruitment remains an important question. Of particular significance are the ‘pioneer’ factors, which are a unique class of TFs that bind to chromatin 14 and establish regions of accessibility and gene activation. A key pioneer factor in Drosophila is the transcriptional activator Zelda, which Melissa Harrison (University of Wisconsin, USA) demonstrated is needed continuously throughout early development for both the minor wave of zygotic transcription and the later major wave of transcription 15. How Zelda achieves transcriptional activation is poorly understood, but Chris Rushlow (NYU, USA) showed that Zelda modifies the probability, timing and rate of transcription of key downstream patterning genes 16. In contrast to Zelda, which is a global activator, Angela Stathopoulos (Caltech, USA) examined the role of broadly expressed repressors in the timing of ZGA in Drosophila. She found that both Runt and Su(H) act as position independent repressors in the pre-MZT Drosophila embryo 17. These findings suggest the possibility that competition between activators and repressors may be a general feature in the timing and spatial activation of different genes during ZGA.
To understand the timing of chromatin changes relative to TF factor binding Shelby Blythe (Northwestern, USA) used ATACseq in Drosophila embryos to show that cell fate specific chromatin states result from the binding of maternally deposited transcription factors on an initial ground state chromatin, pre-MBT 18. In zebrafish, Liyun Miao (Antonio Giraldez lab, Yale, USA) reported that key TFs of early transcription during zebrafish ZGA, Nanog/Oct4/SoxB1, are required to open the chromatin prior to histone acetylation. In addition, Daria Onichtchouk (University of Freiburg, Germany) used MNase seq to show that Nanog/Oct4/SoxB1 are initially recruited to high nucleosome affinity regions (HNARs) within the genome, while subsequently establishing open chromatin domains during ZGA 19. Consistently in Xenopus, Ken Cho (University of California, USA) showed that maternally deposited pioneer transcription factors such as Foxh1 pre-mark enhancers before transcription and before the generation of histone modifications 20. Together, these findings suggest that pioneer transcription factors may be upstream of transcription and chromatin changes during the ZGA across species. While the order of events is important, it is clear that we do not yet know all the factors involved in establishing transcriptional activation at the ZGA as Celia Alda-Catalinas (Wolf Reik lab, Babraham Institute, UK) presented a CRISPR screen in mouse ESCs, identifying new candidates that positively regulate genome activation 21.
ZGA in 3D
The emergence of totipotency and the subsequent specification of zygotic transcription at the ZGA results in major rearrangements in the spatial positioning and the 3D architecture of chromatin. Maria Elena Torres-Padilla (Helmholtz Institute, Munich, Germany) used Dam-ID to show that lamina associated domains (LADs) of chromosomes are established shortly after fertilization in a parental specific manner 22. Paternal LAD formation may be dependent on de novo establishment of H3K4 methylation providing new insights into the temporal order of chromatin architecture establishment in early embryos.
Topologically associated chromatin domains (TADs) are established during the MZT in Drosophila. Gabriel Cavalheiro (Eileen Furlong lab, EMBL, Germany) investigated which insulator proteins and TFs are required for TAD formation and demonstrated that distinct loci establish chromatin organization by different mechanisms. Interestingly using hybrid Drosophila embryos to distinguish between homologous chromosomes, Jelena Erceg, (Ting Wu lab, Harvard Medical School, USA) demonstrated the existence of trans-homolog TADs, which correlate with gene expression and Zelda binding 23.
To understand how pioneer factors physically change chromatin Thomas Quail (Jan Brugues lab, MPI Dresden, Germany) used reconstituted Xenopus egg extracts and fluorescent dCas9 to show that pioneer factor FoxA1 leads to increased rates of chromatin diffusion. While studies in zebrafish embryos presented by Nadine Vastenhouw (MPI Dresden, Germany) suggested that transcribed RNA together with RNA binding proteins are required for euchromatic domain establishment. Such organization might be explained by microphase separation of euchromatin, enabling interactions inside specific regions of the nucleus 24. Further evidence of such compartmentalisation was demonstrated by Ferenc Mueller (University of Birmingham, UK). Using live imaging of native transcription he showed that first transcription in zebrafish (prior to canonical ZGA), such as at the miR-430 locus, occurs in defined nuclear compartments 25, which may be a mechanism to safe-guard early transcription even before the embryo becomes permissive for bulk zygotic gene expression.
Transposon activation, the cost of pluripotency?
Up to 50% of the mammalian genome is made of integrated transposable elements (TEs). The majority of TEs (95%) are retroelements (ERVs, LINES, SINES etc). Reprogramming during early embryogenesis creates a dangerous window where TEs could be de-repressed. Small RNAs called piRNAs expressed in the embryo and germline repress LINE elements through transcriptional and post-transcriptional silencing. Donal O’Carroll (CRM, UK) presented a novel factor in mice called Spocd1 that represses LINE expression through DNA methylation. Despite this, young TEs show a peak of activation during early embryonic development and Didier Trono (EPFL, Switzerland) demonstrated that these young elements have a positive role in increasing chromatin accessibility and enabling human embryonic genome activation, but specialised repressors of the KZFP family (Krüppel-associated box (KRAB)-containing zinc finger proteins) are induced to silence these TEs in a negative feedback loop 26. Juanma Vaquerizas (MPI for Molecular Biomedicine, Muenster) presented a low input chromatin conformation capture technique (low-C) to study the link between TE activation and chromatin architecture and its influence on embryonic transcription 27. He demonstrated that the Murine Endogenous Retroviral Element (MuERV-L/MERVL) family of transposable elements drive the reorganisation of TAD boundaries and activation of a subset of genes in early 2-cell mouse embryo–like cells. Together these studies point towards roles for TE-based regulatory sequences as chromatin control elements, which are co-opted during embryonic development.
Cell cycle regulation in the early embryo
The oocyte to embryo transition is not only associated with transcriptional rewiring but is also associated with dramatic changes in cell cycle control across species. Marc Kirschner (Harvard Medical School, USA) described how the lengthening of the cell cycle is a primary event at the MBT in Xenopus, before the onset of bulk zygotic transcription and cell motility 28. The importance of the rapid early embryonic cell divisions was underscored by K. Neugebauer (Yale, USA) who showed that rapid cell-cycle signalling generates a significant energetic cost in the zebrafish embryo 29. Despite this, the fundamental functions of cell cycle control in coordinating events of the MBT remain poorly understood.
The lengthening of the cell cycle at the MBT is characterised by changes in replication timing during S-phase. Christopher Sansam (OMRF, USA) showed that in zebrafish, replication timing changes during the first 10 hours post fertilization with the most dramatic changes occurring on chromosome 4 30. Unlike Drosophila however 31, the global regulator of chromatin and replication timing Rif1 is not a major contributor to cell cycle length in zebrafish. A significant contributor to cell cycle lengthening is the DNA damage checkpoint, which in Drosophila is activated by transcription-replication conflicts 32. Anahi Molla-Herman (Huynh lab, Collège de France, France) showed that RNA-DNA hybrids formed at highly transcribed tRNA genes might be responsible for checkpoint activation and proper cell cycle timing at the MBT in Drosophila.
Outlook and conclusions
Understanding the coordination of dramatic changes in gene expression, chromatin remodelling, reprogramming, cell cycle as well as a host of other processes which characterise the oocyte to embryo transition requires the coming together of scientists from a multitude of disciplines. This topic benefits from the universality and evolutionary conservation of these control mechanisms as it brings together information from a host of model systems from planaria (Jochen Rink, MPI Dresden) to the marsupial fat-tailed dunnart (Stephen Frankenberg, University of Melbourne, Australia) and importantly to human embryos (Sanna Vuoristo, University of Helsinki, Finland). As quoted by Marc Kirschner “underlying the extreme complexity we may discover a simplicity which now escapes us” (FG Hopkins 1913). The 2019 EMBO workshop “Awakening of the genome: the maternal-to-zygotic transition” provided a unique opportunity to understand the simplicity behind how gametes are rewired to make embryos.
References
Zhang, X. et al. Quantitative Mass Spectrometry to Interrogate Proteomic Heterogeneity in Metastatic Lung Adenocarcinoma and Validate a Novel Somatic Mutation CDK12-G879V. Mol. Cell. Proteomics 18, 622–641 (2019).
Hara, M. et al. Identification of PNG kinase substrates uncovers interactions with the translational repressor TRAL in the oocyte-to-embryo transition. Elife 7, (2018).
Vastenhouw, N. L., Cao, W. X. & Lipshitz, H. D. The maternal-to-zygotic transition revisited. Development 146, (2019).
Sha, Q.-Q. et al. Characterization of zygotic genome activation-dependent maternal mRNA clearance in mouse. Nucleic Acids Res. 48, 879–894 (2020).
Dowdle, M. E. et al. A single KH domain in Bicaudal-C links mRNA binding and translational repression functions to maternal development. Development 146, (2019).
Wu, Q. et al. Translation affects mRNA stability in a codon-dependent manner in human cells. Elife 8, (2019).
Bazzini, A. A. et al. Codon identity regulates mRNA stability and translation efficiency during the maternal‐to‐zygotic transition. EMBO J. 35, 2087–2103 (2016).
Israel, S., Casser, E., Drexler, H. C. A., Fuellen, G. & Boiani, M. A framework for TRIM21-mediated protein depletion in early mouse embryos: recapitulation of Tead4 null phenotype over three days. BMC Genomics 20, 755 (2019).
Laue, K., Rajshekar, S., Courtney, A. J., Lewis, Z. A. & Goll, M. G. The maternal to zygotic transition regulates genome-wide heterochromatin establishment in the zebrafish embryo. Nat. Commun. 10, 1551 (2019).
Zenk, F. et al. Germ line–inherited H3K27me3 restricts enhancer function during maternal-to-zygotic transition. Science (80-. ). 357, 212–216 (2017).
Murphy, P. J., Wu, S. F., James, C. R., Wike, C. L. & Cairns, B. R. Placeholder Nucleosomes Underlie Germline-to-Embryo DNA Methylation Reprogramming. Cell 172, 993-1006.e13 (2018).
Shindo, Y. & Amodeo, A. A. Dynamics of Free and Chromatin-Bound Histone H3 during Early Embryogenesis. Curr. Biol. 29, 359-366.e4 (2019).
Sitbon, D., Boyarchuk, E. & Almouzni, G. Regardless of the deposition pathway, aminoacid 31 in histone variant H3 is essential at gastrulation in Xenopus. bioRxiv 612515 (2019). doi:10.1101/612515
Zaret, K. S. & Carroll, J. S. Pioneer transcription factors: establishing competence for gene expression. Genes Dev. 25, 2227–2241 (2011).
McDaniel, S. L. et al. Continued Activity of the Pioneer Factor Zelda Is Required to Drive Zygotic Genome Activation. Mol. Cell 74, 185-195.e4 (2019).
Yamada, S. et al. The Drosophila Pioneer Factor Zelda Modulates the Nuclear Microenvironment of a Dorsal Target Enhancer to Potentiate Transcriptional Output. Curr. Biol. 29, 1387-1393.e5 (2019).
Koromila, T. & Stathopoulos, A. Distinct Roles of Broadly Expressed Repressors Support Dynamic Enhancer Action and Change in Time. Cell Rep. 28, 855-863.e5 (2019).
Soluri, I. V., Zumerling, L. M., Parra, O. A. P., Clark, E. G. & Blythe, S. A. Zygotic pioneer factor activity of Odd-paired/Zic is necessary for establishing the Drosophila Segmentation Network. bioRxiv 852707 (2019). doi:10.1101/852707
Veil, M., Yampolsky, L. Y., Grüning, B. & Onichtchouk, D. Pou5f3, SoxB1, and Nanog remodel chromatin on high nucleosome affinity regions at zygotic genome activation. Genome Res. 29, 383–395 (2019).
Paraiso, K. D. et al. Endodermal Maternal Transcription Factors Establish Super-Enhancers during Zygotic Genome Activation. Cell Rep. 27, 2962-2977.e5 (2019).
Alda-Catalinas, C. et al. A single-cell transcriptomics CRISPR-activation screen identifies new epigenetic regulators of zygotic genome activation. bioRxiv 741371 (2019). doi:10.1101/741371
Borsos, M. et al. Genome–lamina interactions are established de novo in the early mouse embryo. Nature 569, 729–733 (2019).
Erceg, J. et al. The genome-wide multi-layered architecture of chromosome pairing in early Drosophila embryos. Nat. Commun. 10, 4486 (2019).
Hilbert, L. et al. Transcription organizes euchromatin similar to an active microemulsion. bioRxiv 234112 (2018). doi:10.1101/234112
Hadzhiev, Y. et al. A cell cycle-coordinated Polymerase II transcription compartment encompasses gene expression before global genome activation. Nat. Commun. 10, 691 (2019).
Pontis, J. et al. Hominoid-Specific Transposable Elements and KZFPs Facilitate Human Embryonic Genome Activation and Control Transcription in Naive Human ESCs. Cell Stem Cell 24, 724-735.e5 (2019).
Kruse, K. et al. Transposable elements drive reorganisation of 3D chromatin during early embryogenesis. bioRxiv 523712 (2019). doi:10.1101/523712
Kimelman, D., Kirschner, M. & Scherson, T. The events of the midblastula transition in Xenopus are regulated by changes in the cell cycle. Cell 48, 399–407 (1987).
Rodenfels, J., Neugebauer, K. M. & Howard, J. Heat Oscillations Driven by the Embryonic Cell Cycle Reveal the Energetic Costs of Signaling. Dev. Cell 48, 646-658.e6 (2019).
Siefert, J. C., Georgescu, C., Wren, J. D., Koren, A. & Sansam, C. L. DNA replication timing during development anticipates transcriptional programs and parallels enhancer activation. Genome Res. 27, 1406–1416 (2017).
Seller, C. A. & O’Farrell, P. H. Rif1 prolongs the embryonic S phase at the Drosophila mid-blastula transition. PLOS Biol. 16, e2005687 (2018).
Blythe, S. A. & Wieschaus, E. F. Zygotic Genome Activation Triggers the DNA Replication Checkpoint at the Midblastula Transition. Cell 160, 1169–1181 (2015).
Acknowledgements
We apologize to all those whose work we have not been able to include due to space limitations and we thank EMBO for funding this meeting. J.G. is supported by the European Research Council (ERC-StG-336460ChromHeritance to Kikuë Tachibana) and the L’Oréal Austria Fellowship for Women in Science and is an associated student of the DK Chromosome Dynamics (W1238-B20) supported by the Austrian Science Fund (FWF). K.K. is supported by the Max Planck Society and Deutsche Forschungsgemeinschaft (DFG, SPP2191).
Welcome to our monthly trawl for developmental biology (and related) preprints.
In lock down you might find yourself with more time to read; reading some science might also be a welcome distraction from the pandemic. Lots of fascinating stuff in March, hosted on bioRxivand arXiv– let us know if we missed anything. Use these links to get to the section you want:
Structural and developmental principles of neuropil assembly in C. elegans
Mark W. Moyle, Kristopher M. Barnes, Manik Kuchroo, Alex Gonopolskiy, Leighton H. Duncan, Titas Sengupta, Lin Shao, Min Guo, Anthony Santella, Ryan Christensen, Abhishek Kumar, Yicong Wu, Kevin R. Moon, Guy Wolf, Smita Krishnaswamy, Zhirong Bao, Hari Shroff, William Mohler, Daniel A. Colón-Ramos
Dynamic Sex Chromosome Expression in Drosophila Male Germ Cells
Sharvani Mahadevaraju, Justin M. Fear, Miriam Akeju, Brian J. Galletta, Mara MLS. Pinheiro, Camila C. Avelino, Diogo C. Cabral-de-Mello, Katie Conlon, Stafania Dell’Orso, Zelalem Demere, Kush Mansuria, Carolina A. Mendonça, Octavio M. Palacios-Gimenez, Eli Ross, Max Savery, Kevin Yu, Harold E. Smith, Vittorio Sartorelli, Nasser M. Rusan, Maria D. Vibranovski, Erika Matunis, Brian Oliver
Defining totipotency using criteria of increasing stringency
Eszter Posfai, John Paul Schell, Adrian Janiszewski, Isidora Rovic, Alexander Murray, Brian Bradshaw, Tine Pardon, Mouna El Bakkali, Irene Talon, Natalie De Geest, Pankaj Kumar, San Kit To, Sophie Petropoulos, Andrea Jurisicova, Vincent Pasque, Fredrik Lanner, Janet Rossant
Timing of organ initiation is crucial for robust organ size
Mingyuan Zhu, Weiwei Chen, Vincent Mirabet, Lilan Hong, Simone Bovio, Soeren Strauss, Erich M. Schwarz, Satoru Tsugawa, Zhou Wang, Richard S. Smith, Chun-Biu Li, Olivier Hamant, Arezki Boudaoud, Adrienne H. K. Roeder
A new role for histone demethylases in the maintenance of plant genome integrity
Javier Antunez-Sanchez, Matthew Naish, Juan Sebastian Ramirez-Prado, Sho Ohno, Ying Huang, Alexander Dawson, Deborah Manza-Mianza, Federico Ariel, Cecile Raynaud, Anjar Wilbowo, Josquin Daron, Minako Ueda, David Latrasse, R. Keith Slotkin, Detlef Weigel, Moussa Benhamed, Jose Gutierrez-Marcos
Embryo-like features in developing Bacillus subtilis biofilms
Momir Futo, Luka Opašić, Sara Koska, Nina Čorak, Tin Široki, Vaishnavi Ravikumar, Annika Thorsell, Domagoj Kifer, Mirjana Domazet-Lošo, Kristian Vlahoviček, Ivan Mijaković, Tomislav Domazet-Lošo
Midbody remnant inheritance is regulated by the ESCRT subunit CHMP4C
Javier Casares-Arias, María Ujué Gonzalez, Alvaro San Paulo, Leandro N. Ventimiglia, Jessica B. A. Sadler, David G. Miguez, Leticia Labat-de-Hoz, Armando Rubio-Ramos, Laura Rangel, Miguel Bernabé-Rubio, Jaime Fernández-Barrera, Isabel Correas, Juan Martín-Serrano, Miguel A. Alonso
Conserved epigenetic regulatory logic infers genes governing cell identity
Woo Jun Shim, Enakshi Sinniah, Jun Xu, Burcu Vitrinel, Michael Alexanian, Gaia Andreoletti, Sophie Shen, Brad Balderson, Guangdun Peng, Naihe Jing, Yuliangzi Sun, Yuliang Wang, Patrick P L Tam, Aaron Smith, Michael Piper, Lionel Christiaen, Quan Nguyen, Mikael Bodén, Nathan J. Palpant
Content and performance of the MiniMUGA genotyping array, a new tool to improve rigor and reproducibility in mouse research
John Sebastian Sigmon, Matthew W Blanchard, Ralph S Baric, Timothy A Bell, Jennifer Brennan, Gudrun A Brockmann, A Wesley Burks, J Mauro Calabrese, Kathleen M Caron, Richard E Cheney, Dominic Ciavatta, Frank Conlon, David B Darr, James Faber, Craig Franklin, Timothy R Gershon, Lisa Gralinski, Bin Gu, Christiann H Gaines, Robert S Hagan, Ernest G Heimsath, Mark T Heise, Pablo Hock, Folami Ideraabdullah, J. Charles Jennette, Tal Kafri, Anwica Kashfeen, Samir Kelada, Mike Kulis, Vivek Kumar, Colton Linnertz, Alessandra Livraghi-Butrico, Kent Lloyd, Richard Loeser, Cathleen Lutz, Rachel M Lynch, Terry Magnuson, Glenn K Matsushima, Rachel McMullan, Darla Miller, Karen L Mohlke, Sheryl S Moy, Caroline Murphy, Maya Najarian, Lori O’Brien, Abraham A Palmer, Benjamin D Philpot, Scott Randell, Laura Reinholdt, Yuyu Ren, Steve Rockwood, Allison R Rogala, Avani Saraswatula, Christopher M Sasseti, Jonathan C Schisler, Sarah A Schoenrock, Ginger Shaw, John R Shorter, Clare M Smith, Celine L St. Pierre, Lisa M Tarantino, David W Threadgill, William Valdar, Barbara J Vilen, Keegan Wardwell, Jason K Whitmire, Lucy Williams, Mark Zylka, Martin T Ferris, Leonard McMillan, Fernando Pardo-Manuel de Villena
Comparing quality of reporting between preprints and peer-reviewed articles in the biomedical literature
Clarissa F. D. Carneiro, Victor G. S. Queiroz, Thiago C. Moulin, Carlos A. M. Carvalho, Clarissa B. Haas, Danielle Rayêe, David E. Henshall, Evandro A. De-Souza, Felippe E. Amorim, Flávia Z. Boos, Gerson D. Guercio, Igor R. Costa, Karina L. Hajdu, Lieve van Egmond, Martin Modrák, Pedro B. Tan, Richard J. Abdill, Steven J. Burgess, Sylvia F. S. Guerra, Vanessa T. Bortoluzzi, Olavo B. Amaral
The Centre for Integrative Biology of Toulouse (CBI) is looking to strengthen its strong research community in Cellular and Developmental Biology. Existing groups investigate numerous aspects from cell division, morphogenesis to progenitor/stem cell biology using model organisms. Candidates complementing or reinforcing this community are encouraged to apply. Particular attention will be given to interdisciplinary, experimental and/or theoretical projects encompassing biophysics, systems biology and/or computational approaches. Further information can be found on the CBI website (http://cbi-toulouse.fr/eng/accueil-nouvelles-equipes) or by contacting Patrick Blader directly (patrick.blader@univ-tlse3.fr).
 (3 votes)
(3 votes)
 (No Ratings Yet)
(No Ratings Yet)


 (39 votes)
(39 votes)