We are seeking to recruit an outstanding group leader who aims to explore bold new areas of biological inquiry and carry out interdisciplinary research to investigate multicellular development at all scales. How organisms respond and adapt to their environment, during development and throughout their lifetime, is of central interest.
The Developmental Biology Unit seeks to understand the general principles and mechanisms underlying the development of multicellular organisms. Researchers in the unit combine the power of genetic model organisms with quantitative imaging and -omics technologies, synthetic biology, reduced (in vitro) systems and theoretical modelling, to create a cross-cutting approach to modern developmental biology.
Research in the Developmental Biology Unit is firmly embedded within the overall EMBL environment, with extensive in-house collaborations, access to outstanding graduate students and postdoctoral fellows, and support from cutting-edge facilities, including genomics, transgenesis, metabolomics, mass-spectrometry, and microscopy.
Your role
You will lead a research group to pursue highly ambitious and original research at the frontier of developmental biology. In general, EMBL appoints group leaders early in their career and provides them with a very supportive, collaborative environment and generous work package for their first independent position. Significant core funding and limited teaching responsibilities allow you undertake a farsighted research program.
You have
The successful candidate will present a highly original and ambitious research plan that concisely describes the background and status of the questions that will be addressed, the experimental strategies and methods that will be employed, and the ultimate goals. A PhD degree in the Natural Sciences is expected. Candidates with a background in physics and modelling of developmental processes are also encouraged to apply.
Why join us
EMBL is an inclusive, equal opportunity employer offering attractive conditions and benefits appropriate to an international research organization with a very collegial and family friendly working environment. EMBL is committed to achieving gender balance and strongly encourages applications from women. Appointment will be based on merit alone. The remuneration package comprises a competitive salary, a comprehensive pension scheme, medical, educational and other social benefits, as well as financial support for relocation and installation, including your family, and the availability of an excellent child care facility on campus.
What else you need to know
We are Europe’s flagship research laboratory for the life sciences – an intergovernmental organisation performing scientific research in disciplines including molecular biology, physics, chemistry and computer science. We are an international, innovative and interdisciplinary laboratory with more than 1700 employees from many nations, operating across six sites, in Heidelberg (HQ), Barcelona, Hinxton near Cambridge, Hamburg, Grenoble and Rome.
Our mission is to offer vital services in training scientists, students and visitors at all levels; to develop new instruments and methods in the life sciences and actively engage in technology transfer activities, and to integrate European life science research. The working language of the institute is English.
In your online application, you will be asked to include a cover letter, your CV, the names and contact details of 3 referees and a concise description of research interests & future research plans, typically not exceeding five pages.
Further information about the position can be obtained from the Head of Unit, Anne Ephrussi (anne.ephrussi@embl.de).
Information on Group Leader appointments can be found here http://www.embl.org/gl_faq.
Interviews are planned for 18, 19 and 20 December 2019.
An initial contract of 5 years will be offered to the successful candidate. This is foreseen to be extended to a maximum of 9 years, subject to an external review.
Rapid turn over of sex determination mechanisms provides biologists with an elegant study system connecting sexual selection to molecular evolution. Striking examples of this turnover are found in African cichlids, where multiple sex determination signals exist not only within the same genera, but sometimes within the same species [1]; in the common house fly, where the primary sex determiner varies along a longitudinal gradient [2], and in the Japanese frog Rana rugosa, a species in which sex chromosomes segregate geographically but whose signals can be overridden by environmental (hormonal) intervention [3].
Downstream of the primary sex determination signal is a developmental pathway that also presents an intriguing puzzle for evolutionary biologists. While the sexual differentiation pathways of mice and humans are fundamentally similar, and while the downstream elements of this pathway are conserved between bees and flies, there are no shared genes in the sex determination pathways of mammals, arthropods, and nematodes with the striking exception of the Doublesex Mab-3 Related Transcription factor (DMRT) family. Furthermore, the fundamental biochemistry of sexual differentiation in these taxa is distinct. In mammals, signaling molecules establish feedback loops that promote one sexual identity and repress the opposite one [4]. In insects, sex-specific RNA splicing determines sexual identity [5]. And in nematodes, a cast of genes including a phosphatase and protease control the sexual fate of a cell [6]. Thus the puzzle: how does a pathway which appears stable on the order of hundreds of millions of years diverge so distinctly between different animal taxa? The Kopp lab was interested in how it is that the sexual differentiation pathway changes on a macro-evolutionary time scale.
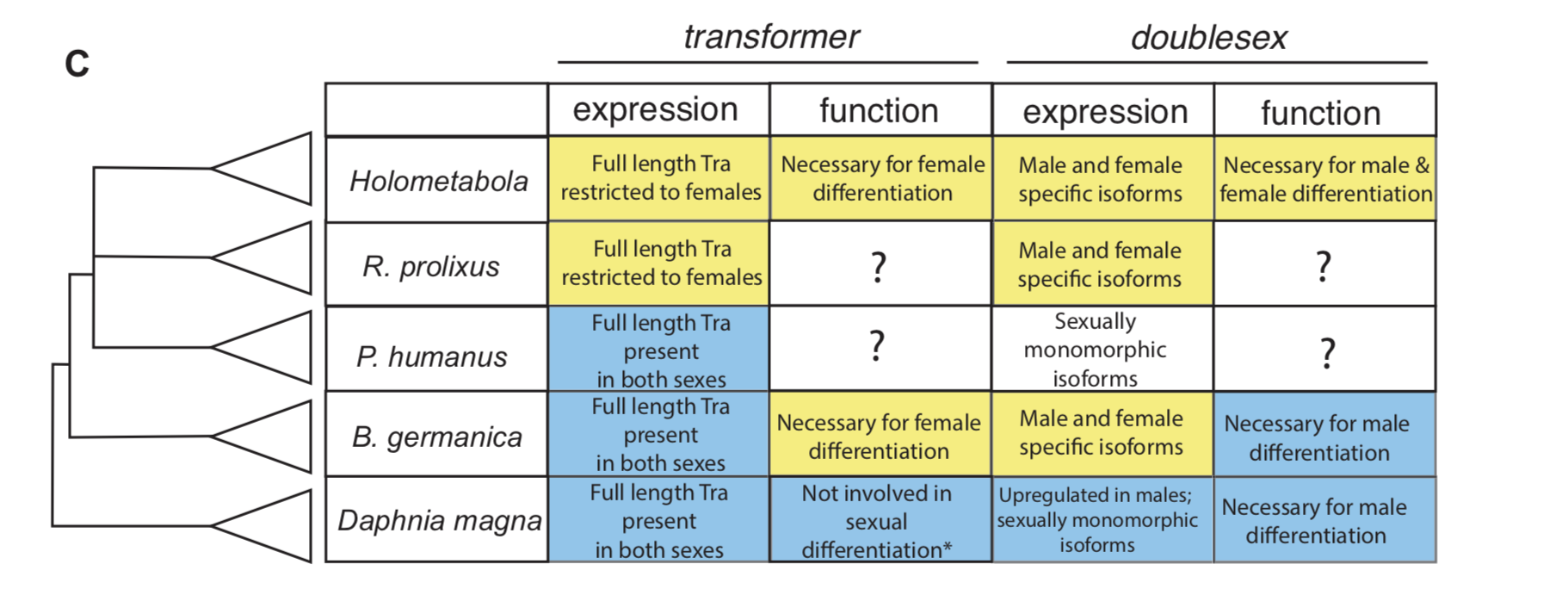
In insect developmental biology Drosophila is the point of reference for all other species. Sex differentiation is no different, where two genes at the bottom of this pathway — transformer (tra) and doublesex (dsx) — were first described in the fruit fly. doublesex (a member of the eponymous Doublesex Mab-3 Related Transcription factor family) is a transcription factor with male- and female-specific isoforms. Both isoforms contain the same DNA binding domain, while alternative, sex-specific splicing at the 3’ end of dsx transcripts changes the effect the male and female dsx isoforms have on their targets [7]. In Drosophila, the male isoform of dsx is produced by default. Active intervention by Tra changes the pattern of dsx splicing from a male to a female isoform. tra itself is also sex specifically spliced, such that a premature stop codon appears in the male, but not the female, transcript [8]. The outgroup to insects with the best studied developmental model is the class Branchiopoda, containing the model crustacean Daphnia magna. In Daphnia, dsx is upregulated in males and lacks male and female isoforms. It is required for male, but not female differentiation [9]. tra is not sex-specifically spliced in this species [10].
To investigate how this insect-specific pathway of sexual differentiation based on sex-specific RNA splicing evolved from an ancestral state without male and female isoforms, we had to look beyond the Kopp lab’s traditional Drosophila domain. Way beyond it. Drosophila-like splicing and function have been documented in Coleopterans and in Hymenopterans, suggesting that the tra–dsx axis of sexual differentiation was present in the common ancestor of Holometabola [11,12,13,14]. The phylogenetic interval between crustaceans and Holometabola is filled in large part by the paraphyletic group of hemimetabolous insects — or those that go through a partial metamorphosis. And because insect pests are more likely to have sequenced genomes, we settled on the following delightful trio of hemimetabolous insects: the kissing bug Rhodnius prolixus (order: Hemiptera), the louse Pediculus humanus (order: Pthiraptera), and the German cockroach Blattella germanica (order: Blattodea). Because R. prolixus and P. humanus are both obligate blood feeders, they cannot be cultured in a lab without strict regulations. Thus, we were reliant on the kindness of Ian Orchard’s lab in Toronto for R. prolixus tissue shipped in RNALater. For P. humanus, our tissue collection was somewhat more exciting — we struck up a relationship with a local louse collector, who is hired to comb nits and lice from the heads of infested humans. In contrast, my experience getting to know German roaches was much more civilized. I was lucky to receive an NSF EDEN grant to study in Barcelona for two months with Xavier Belles, where I learned how to knock down genes in B. germanica. The Belles lab has done extensive work developing B. germanica into an evo-devo model.
Double stranded RNAi targeting transformer masculinizes females. From Wexler et al, eLife 2019;8:e47490.
For all three species, we investigated the isoforms of tra and dsx present in males and females. In the German cockroach, we knocked down both genes to probe their functions. The phylogenetic breadth of our samples allowed us to observe the evolutionary trajectory of dsx and tra across a large evolutionary interval. We discovered that the holometabolous pattern of tra splicing, with a male-specific premature stop codon, evolved after tra evolved a functional role as a regulator of female development. Our results from R. prolixus, where we isolated tra with a male specific stop codon, showed that the “canonical” pattern of sex-specific splicing of this gene did evolve before the common ancestor of Holometabola. In contrast, it appears that the familiar holometabolous pattern of male- and female-specific dsx isoforms differing at their 3’ end evolved before dsx became a key regulator of female differentiation. So for each gene, we observed a different story: with dsx, the “holometabolous” splicing pattern evolved before the “holometabolous” functional role; for tra, this pattern was reversed.
Summary of tra and dsx expression and function in the three hemimetabolous insect orders in comparison to the canonical holometabolous mechanism and to Daphnia. From Wexler et al, eLife 2019;8:e47490.
The knockdown experiments lead to some very fun collaborations as we attempted to probe the resulting phenotypes. After I returned from Spain and set up my own B. germanica colony at UC Davis, I was reliant on the generosity of Coby Schal, who shared a few roaches from his colony, allowing me to set up my own. The Schal lab specializes, among other things, in the study of insect behavior. When I attended the conference of the International Congress of Entomology with Coby a year later, I excitedly sat down to show him images of the masculinized females we observed upon knocking down tra. Coby then offered to quantify the degree to which these roaches’ behavior was masculinized. The experiments he did with his lab technician Ayako Wada-Katsumata revealed that like in Drosophila, sex-specific behavior of the German roach is under the control of tra. Separately, I began chatting with my teammate from the UC Davis cycling team about the work. This teammate and friend, Matt Amicucci, also happened to be a graduate student in the chemistry department, who then offered to help investigate the oligosaccharides produced by masculinized females in their tergal glands. Thanks to Matt’s expertise, we learned that tra also controls the chemical physiology of German roaches.
The breadth of the questions motivating this project and the number of questions that came out of it; the diversity of skills among the paper’s authors, and the generous support of everyone involved really made this work a wonderful and memorable PhD.
Above: Wild type males respond to the stimulus of a wild type female antenna by raising their wings and presenting their tergal gland.
Above: Females treated with dstra respond as wild type males do to the stimulus of a wild type female antenna. These treated females raise their wings in a stereotypical male courtship display.
The research group headed by Werend Boesmans and Veerle Melotte is looking for a highly motivated and talented PhD student to investigate the role of the intrinsic innervation of the gut in cancer. The enteric nervous system, also known as the second brain, is a mostly ignored member of the tumor microenvironment but is linked to the development and progression of colorectal cancer. In the current project, the involvement of specific neural cell types in colorectal tumorigenesis will be investigated in both in vivo and in vitro models, and combined with advanced optical microscopy, genetic lineage tracing, marker gene expression analysis and single cell transcriptomics.
This PhD project takes place within the School for Life Sciences (SLS) of the Transnational University of Limburg (tUL), a unique collaboration between Hasselt University (UHasselt, Belgium) and Maastricht University (UM, The Netherlands). The student will perform research at the Biomedical Research Institute (BIOMED, https://www.uhasselt.be/biomed) and the School for Oncology and Developmental Biology (GROW, https://www.maastrichtuniversity.nl/research/school-oncology-and-developmental-biology) embedded in the Department of Pathology of the Maastricht University Medical Center (MUMC).
You can only apply online.
The selection procedure consists of a preselection based on the application file and an interview.
Please provide the contact details of two referents in your application.
Story behind our recent paper in Current Biology “Cadherin-Mediated Cell Coupling Coordinates Chemokine Sensing across Collectively Migrating Cells” (Tugba Colak-Champollion, Ling Lan, Alisha R. Jadhav, Naoya Yamaguchi, Gayatri Venkiteswaran, Heta Patel, Michael Cammer, Martin Meier-Schellersheim, Holger Knaut)
Guided cell migration is a crucial event in many biological and mechanical processes. During development, orchestrated movements of cells pattern tissues and organs. Wounds in our bodies close by the migration of cell sheets. Cell migration is one of the weapons of the immune system, which sends leukocytes to the site of infection to fight against pathogens. But cell migration also contributes to several pathological conditions, such as the dissemination of cancer cells to the sites of metastasis in the body. Thus, detailed studies of mechanisms of cell migration are likely to improve our understanding of animal development, homeostasis, and disease progression.
Cell migration occurs in two major modes: single cell migration and collective cell migration. Single cell migration can be defined as a cell migrating on its own upon extracting directional information and polarizing and moving towards the target site. While collectively migrating cells also extract directional information and polarize toward the target site, they also have the additional task of coordinating with each other to move in the same direction and at the same speed.
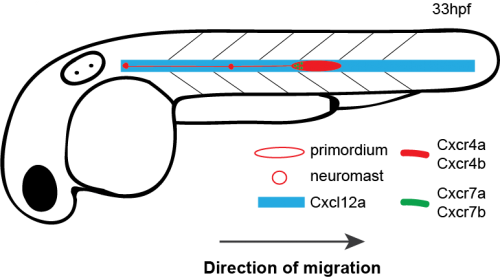
How do cells move as a coherent collective following guidance cues? In the latest study from our lab [1], my coauthors and I investigated how directional sensing in collectively migrating cells is organized. We used the Zebrafish lateral line primordium (primordium) as a model for our study. The primordium is a collective of tightly adhering ~100 cells, which originates behind the ear of the fish at around 18 hours post fertilization (hpf) and migrates to the tip of the tail by 42 hpf.
The primordium migration is guided by chemokine signaling. For proper migration, the primordium requires the chemokine Cxcl12a and its receptors Cxcr4b and Cxcr7b. Importantly, the signaling receptor Cxcr4b is expressed in every primordium cell, while the Cxcr7b expression is restricted to the rear of the collective, where it acts as a sink receptor and generates a Cxcl12a gradient across the tissue [2-5].
Schematic of primordium migration and chemokine signaling system in primordium
Primordium migration in wild-type embryo
In collective cell migration, a prevailing model suggests that there is a division of labor between two groups of cells: leaders and followers. These two groups of cells have both topological and functional attributes. For example, leaders are located in front of the collective, whereas followers are located in the rear. In terms of functionality, the model suggests that leader cells explore their surroundings extracting directional information and relaying this information to the follower cells.
It was proposed that the primordium migration also follows a leader-follower model. An elegant study by Haas and Gilmour generated supporting evidence for this model using chimeric analysis [6]. They placed wild-type cells, which can see the chemokine signal into cxcr4b mutant primordia whose migration stalls prematurely. When at least a few wild-type cells ended up in the front region, the migration was restored – albeit at a slower speed than the speed of wild-type primordia.
However, when I started my Ph.D. project in the Knaut lab, two studies that recently were published made my advisor Holger Knaut and me rethink the leader-follower model [4, 5]. These two studies demonstrated that there is a linear chemokine signaling gradient across the tissue that is essential for the directionality of the primordium. This gradient is available to both leader and follower cells, not just to the leader cells.
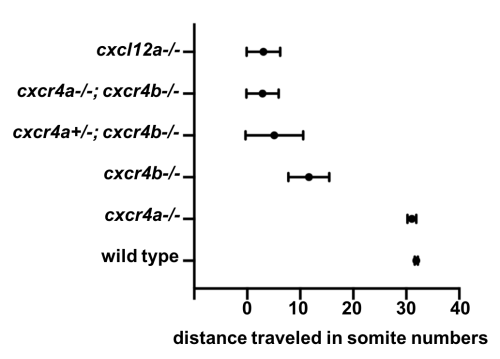
Meanwhile, Gayatri Venkiteswaran, a post-doc in our lab, made an interesting observation when she was scoring the primordium migration phenotype in chemokine receptor mutants. Previously, it was known that loss of Cxcr4b leads to primordium stalling. However, loss of the other Cxcr4 receptor, Cxcr4a, that is also expressed in the primordium cells [7] does not result in a primordium migration defect. Gayatri found that taking away one copy of cxcr4a from cxcr4b-/-primordia makes the cxcr4b-/- phenotype worse and taking away both copies of both receptors is much worse than the cxcr4b-/- or cxcr4a+/-; cxcr4b-/-phenotypes. This observation suggested that Cxcr4a also contributes to migration and a primordium that is cxcr4b-/- can still see a little bit of the chemokine signal. This meant the earlier study which suggested that the existence of a few chemokine-sensitive cells could restore the migration phenotype of chemokine-blind (cxcr4b-/-) cells was conducted using primordia that could still see some of the directional signal because they retained functional Cxcr4a.
Quantification of primordium migration in 48 hpf embryos of indicated genotypes
Primordium migration in cxcr4b-/- embryo
Primordium migration in cxcr4a-/-;cxcr4b-/- embryo
In the light of these two pieces of new information—existence of a linear gradient across tissue and Cxcr4a’s contribution to directional migration—we decided to take a closer look at the leader-follower model and the contribution of follower cells to directionality and speed of the primordium.
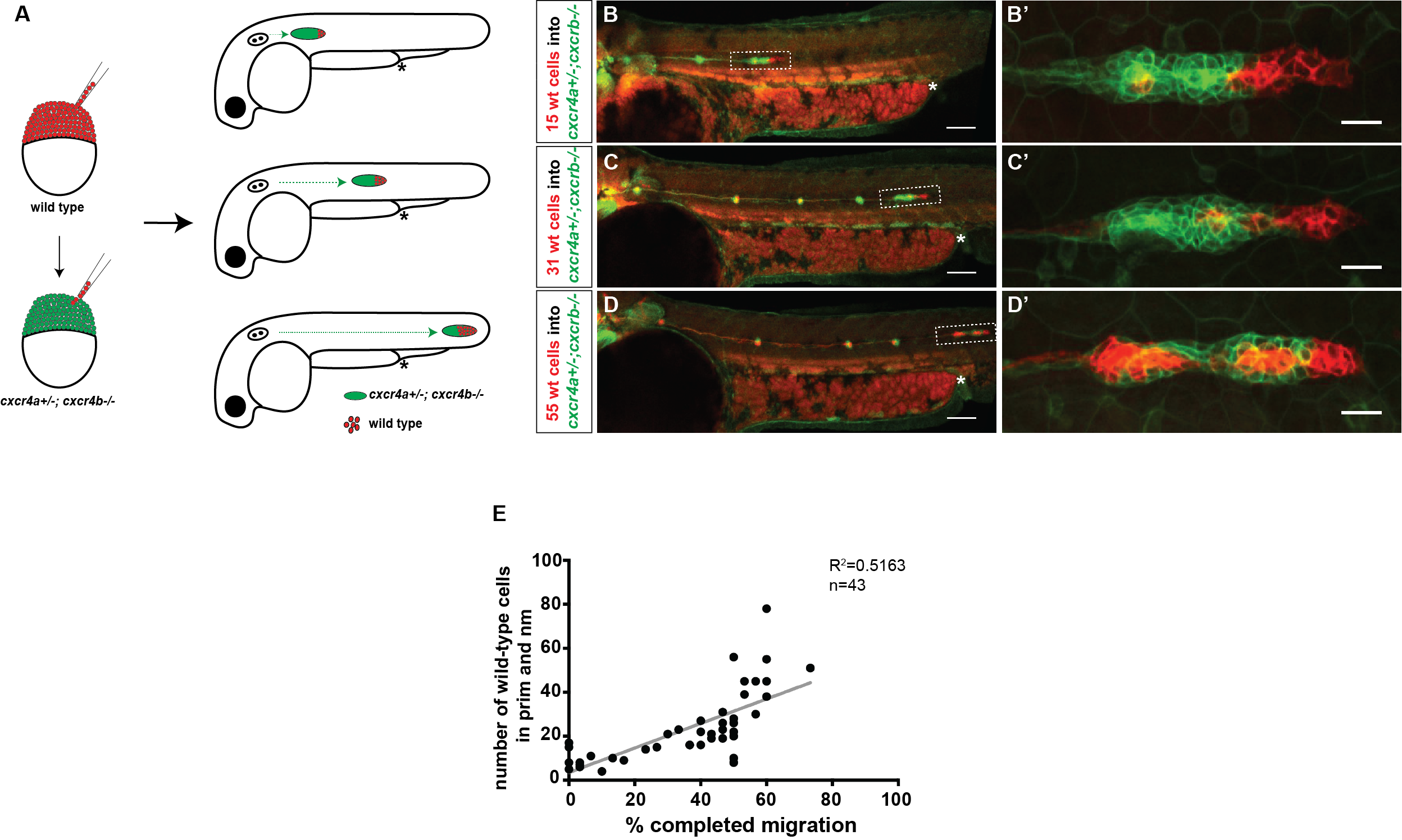
Our first question was whether a few chemokine-sensitive (wild-type) cells could restore the migration phenotype of completely blind (cxcr4a-/-; cxcr4b-/-) primordia. To answer this question, we used a classical developmental biology technique named chimeric analysis. The idea was putting cells from wild-type donor embryos into cxcr4a-/-; cxcr4b-/- host embryos at the blastula stage. The donor primordium cells are labeled with a red transgenic marker and the host primordium cells are labeled with a green transgenic marker. After transplantation, the embryos are grown until 30 hpf and the transplanted host embryos are screened for the presence of red donor cells in the primordium. Although chimeric analysis provides very clean and reliable data, it is an inefficient technique. Unfortunately, not every single embryo transplanted gets the donor cells in the primordium. Additionally, it is a difficult technique and takes a while to perfect it. Even when you become very good at it, you can still have some days when the host mortality is high for unknown reasons.
Schematic of blastomere transplantation
Despite the difficulty and inefficiency of chimeric analysis, we still chose to do it because it was the best way to answer our questions. There was an additional layer of difficulty which turned out to be a good thing later: to generate cxcr4a-/-; cxcr4b-/-embryos, we had to in-cross cxcr4a+/-; cxcr4b-/- fish because cxcr4a-/-; cxcr4b-/- adults are not viable. This meant only a quarter of our host embryos would have the desired genotype. Half would be cxcr4a+/-; cxcr4b-/- and a quarter would be cxcr4b-/-. We had no way of knowing the genotype of the chimeric host embryos until going through a genotyping protocol after imaging these embryos. Nevertheless, we got what we wanted and more.
We found that a few chemokine sensitive (wild-type) cells do not restore the migration phenotype of completely blind (cxcr4a-/-; cxcr4b-/-) primordia, an observation that is inconsistent with the classical leader-follower model. However, we had one embryo in which about half of the primordium consisted of wild-type cells, and the other half consisted of cxcr4a-/-; cxcr4b-/-cells, and this chimeric primordium migrated about half of its path. This made us consider the possibility that cells pull on each other during migration. When there are many cells which can see the chemokine, they might generate sufficient pulling forces in the right direction to drag their chemokine blind neighbors along—in the case of this chimeric embryo, half of the way.
We decided to follow up on this observation using a quantitative approach. We predicted that as we increase the number of chemokine-sensing cells in an otherwise chemokine-blind primordium, the distance migrated by the chimeric primordium should increase. To quantify this behavior, we needed a lot of samples. Using chemokine-blind primordia (cxcr4a-/-; cxcr4b-/-) as hosts posed two challenges. First, it is difficult to get them (remember that only one-quarter of the embryos are of this genotype). Second, it might require a lot of chemokine-sensing cells to pull their completely chemokine-blind neighbors along. To overcome these challenges, we considered using cxcr4a+/-; cxcr4b-/- primordia as hosts. These almost chemokine-insensitive primordia have only one copy of cxcr4a left. Gayatri’s quantification of the migration behavior showed that the migration of these cxcr4a+/-; cxcr4b-/- primordia are worse than cxcr4b-/-primordia and a little bit better than cxcr4a-/-; cxcr4b-/- primordia. Having just a little bit of Cxcr4a activity would help us resolve the relationship between migrated distance and chemokine-sensing cell number, we hoped.
A) Schematic of experimental design and predictions. B-D’) Examples of chimeric primordia with low, medium, and high donor cell contribution. E) Graph of the total number of wild-type cells in the chimeric primordia (total primordial wild-type cells include interneuromast, neuromast (nm) and primordium (prim) cells) versus the migration distance of cxcr4-deficient primordia. 48 hpf.
Luckily, we already imaged a lot of chimeric primordia that consisted of chemokine-sensitive (wild-type) and chemokine-insensitive (cxcr4a+/-; cxcr4b-/-) cells. So we went back to this data set and counted the number of wild-type cells and measured how far these chimeric primordia migrated. As we predicted, the chimeric primordia migrated further as the number of chemokine-sensitive cells increased. This observation was consistent with the idea of cells “pulling” on each other. Next, we asked what could facilitate this “pulling”. We turned to the obvious candidates: cell-cell adhesion molecules. And there are a few of them expressed by the primordium cells including E-cadherin (Cadherin 1, Cdh1) and N-cadherin (Cadherin 2, Cdh2) [8-10].
We first decided to focus on Cdh1 and Cdh2. Our first question was how cdh1-/- and cdh2-/- cells behave during migration. The simplest method of answering this question would be observing the migration behavior of cdh1 or cdh2 mutant primordia under a microscope. However, this was not an option for either of these two genotypes. The problem with cdh1-/- embryos is that they die during gastrulation before the primordium is formed. As for cdh2-/- embryos, they have problems with somite development that affect the proper expression of Cxcl12a (the directional cue). Thus, we went back to our favorite technique, chimeric analysis by blastomere transplantation.
Live imaging of cdh1-/- or cdh2-/- cells in an otherwise wild-type primordium showed that lacking either of the cadherins does not affect migration. These mutant cells co-migrated normally with their wild-type neighbors. This raised the next question: Are these cadherins acting redundantly? To answer this question, we needed to generate cdh1-/-; cdh2-/- embryos to be used as donors. However, this is a real challenge, as the cadherin mutants are not adult viable. To obtain the desired genotype, we had to in-cross cdh1+/-; cdh2+/- fish. The chance of finding a double cadherin mutant embryo is one in sixteen! Despite the odds, we did this experiment—repeatedly. We never found a chimeric embryo whose donor embryo was cdh1-/-; cdh2-/-. But we obtained a good sample size that made us confident to think that either such double cadherin mutant cells do not become primordium cells or they dissociate from the primordium very early on. Despite not being able to find what we set out to find, we encountered some interesting genetic scenarios. For example, cdh1-/-; cdh2+/- cells dissociated from the migrating primordium when they were placed at the tip of the primordium, whereas cdh1+/-; cdh2-/- cells fell off the primordium when they were located at the rear of the primordium. These findings were puzzling until the next observation.
To observe Cdh1 and Cdh2 expression in the primordium, we made BAC transgenic lines that expressed Cdh1-GFP and Cdh2-mCherry. A closer look at the primordia in these transgenic fish revealed that Cdh1 was expressed more in the front region of the primordium and Cdh2 was expressed more in the rear. This could be a possible explanation for the observation mentioned above, that cdh1-/-; cdh2+/- cells fell off from the tip positions and cdh1+/-; cdh2-/- cells fell off from the rear positions. Perhaps Cdh1 was needed more in the front and Cdh2 more in the rear?
The observation of cadherin deficient cells falling off from the primordium also suggested that cell-cell adhesion through cadherins couples the migrating cells. However, is such cell coupling necessary for cells to pull on each other? To answer this question, we transplanted cdh1-/- or cdh2-/- cells into cxcr4b-/-primordia. While cxcr4b-/- primordia have a migration defect due to not being able to see the chemokine signal properly, cadherin mutant cells still have the chemokine receptors and should migrate directionally. To our surprise, cdh1-/- cells located in the front region split away from the cxcr4b-/- cells. But cdh2-/- cells located in the front region pulled the cxcr4b-/- cells along. This finding was consistent with the differential expression pattern of Cdh1 (more in front) and Cdh2 (more in rear) that we had observed earlier. Additionally, a little bit of literature digging revealed that in vitro studies suggested that Cdh1 could withhold more force than Cdh2 [for example 11]. It is plausible that there is increased tension between the donor population, which moves persistently in a specific direction, and the host population, which moves in random directions. A strong physical attachment might be necessary to keep these two groups together.
These findings suggested that the physical coupling of primordium cells is important for the group’s directional migration. But how does cell-cell adhesion affect individual cell directionality within a group? Up on discussions of our data with Martin Meier-Schellersheim, who is a physicist at the NIH and a co-author, we decided to take a quantitative approach using nuclear movement as a proxy for cell movement.
Next, we utilized our favorite technique, blastomere transplantation, once again and placed donor cells (wild type, cdh1-/-, cdh2-/-, or cxcr4b-/-) labeled with H2A-GFP into wild-type host primordia whose cell nuclei were labeled with H2A-mCherry.
Using commercial image software, we tracked the individual donor and host nuclei in different primordium locations at high spatial and temporal resolution. To analyze the tracking data, we used custom scripts to assess directional sensing using three measures: neighbor-neighbor distance, directionality index, and directionality angle. The nuclear tracking analysis showed that directional sensing of cadherin and cxcr4b deficient cells was impaired based on all three categories, lack of Cdh1 having the most severe effect. Together these data showed that in addition to directional cue sensing, efficient migration requires cadherin-mediated cell coupling.
Our observations suggested that cadherin-mediated cell-cell adhesion is important for coordinating cell movements in the migrating primordium. To test this further, we decided to use a gene trap line that expresses functional alpha E-catenin tagged with Citrine (Ctnna1-Citrine) from the endogenous promoter that we recently obtained from Scott Fraser’s lab [12]. Ctnna connects Cadherins on the plasma membrane to the actin cytoskeleton; therefore, lack of Ctnna should impair cadherin-mediated cell-cell adhesion. Chimeric analysis using blastomere transplantation would be the way to approach this experiment since the ctnna-/- embryos die during somitogenesis, before the primordium develops. But luckily, our lab recently developed a protein degradation system named zGrad that could be an efficient alternative to chimeric analysis [13]. As expected, the time-lapse analysis showed that depletion of Ctnna1 resulted in primordia that migrate less directionally. Interestingly, cells separated from each other during migration forming large irregular gaps consistent with the idea that cadherin-catenin complexes mediate the adhesion among the cells in the primordium.
In summary, these results suggest that all the cells in the primordium interpret the directional information and are physically coupled to each other to achieve robust migration. This behavior is not unlike some Turkish folk dances, which are characterized by groups of dancers who hold hands tightly as they dance to a tune in a synchronized fashion. Just as each dancer needs to listen to the music, each cell needs to sense the directional signal in order to coordinate their movements. Through their tight connections, dancers and cells alike synchronize their individual motions, thereby moving in unison.
References
Colak-Champollion, T., et al., Cadherin-Mediated Cell Coupling Coordinates Chemokine Sensing across Collectively Migrating Cells.Curr Biol, 2019. 29(15): p. 2570-2579 e7.
Dambly-Chaudiere, C., N. Cubedo, and A. Ghysen, Control of cell migration in the development of the posterior lateral line: antagonistic interactions between the chemokine receptors CXCR4 and CXCR7/RDC1.BMC Dev Biol, 2007. 7: p. 23.
Valentin, G., P. Haas, and D. Gilmour, The chemokine SDF1a coordinates tissue migration through the spatially restricted activation of Cxcr7 and Cxcr4b.Curr Biol, 2007. 17(12): p. 1026-31.
Dona, E., et al., Directional tissue migration through a self-generated chemokine gradient.Nature, 2013. 503(7475): p. 285-9.
Venkiteswaran, G., et al., Generation and dynamics of an endogenous, self-generated signaling gradient across a migrating tissue.Cell, 2013. 155(3): p. 674-87.
Haas, P. and D. Gilmour, Chemokine signaling mediates self-organizing tissue migration in the zebrafish lateral line.Dev Cell, 2006. 10(5): p. 673-80.
Siekmann, A.F., et al., Chemokine signaling guides regional patterning of the first embryonic artery.Genes Dev, 2009. 23(19): p. 2272-7.
Matsuda, M. and A.B. Chitnis, Atoh1a expression must be restricted by Notch signaling for effective morphogenesis of the posterior lateral line primordium in zebrafish.Development, 2010. 137(20): p. 3477-87.
Revenu, C., et al., Quantitative cell polarity imaging defines leader-to-follower transitions during collective migration and the key role of microtubule-dependent adherens junction formation.Development, 2014. 141(6): p. 1282-91.
Kozlovskaja-Gumbriene, A., et al., Proliferation-independent regulation of organ size by Fgf/Notch signaling.Elife, 2017. 6.
Panorchan, P., et al., Single-molecule analysis of cadherin-mediated cell-cell adhesion.J Cell Sci, 2006. 119(Pt 1): p. 66-74.
Trinh le, A., et al., A versatile gene trap to visualize and interrogate the function of the vertebrate proteome.Genes Dev, 2011. 25(21): p. 2306-20.
Yamaguchi, N., T. Colak-Champollion, and H. Knaut, zGrad is a nanobody-based degron system that inactivates proteins in zebrafish.Elife, 2019. 8.
This project will generate transgenic G. mellonella lines with fluorescent haemocytes, to allow visualisation of the immune response in this non-mammalian model organism. The larvae will be injected with the human fungal pathogen Candida albicans and the haemocytes tracked via time-lapse photography. The aim is to build an evidence base to support the wider adoption of G. mellonella larvae for studying host/pathogen interactions, providing an alternative to the use of mice. We’re looking for an experienced and dynamic Postdoc (Grade E or F) and a 0.5FTE Technician to join the team, alongside an existing PhD student.
For further details of the PDRA/F post see: https://jobs.exeter.ac.uk/hrpr_webrecruitment/wrd/run/ETREC107GF.open?VACANCY_ID%3d066891Q4Cg%1BUSESSION=68D96D4363BB4B7192452B3DEB6F3F37&WVID=3817591jNg&LANG=USA
For further details of the 0.5FTE Tech D post see:
The Tunicate embryology Research Team and the MRI core imaging facility in Montpellier, France, have an opening for a computer scientist in charge of the analysis of 3D + time fluorescence imaging data of living embryos of a class of marine invertebrates, the ascidians. The contract is for up to 4 years.
The selected candidate will use and further develop a tool for the automatic segmentation of fluorescence imaging data obtained using a light sheet microscope, ASTEC (Automated Segmentation and Tracking of Embryonic Cells) developed by the team and its collaborators (for an example output of ASTEC, watch the Video showing a segmented Phallusia embryo).
Missions:
– Use ASTEC to segment and track the imaging data collected by the team’s biologists. Perform statistical analyses of embryonic morphology (reproducibility between individuals and between species, phenotyping,…).
– Develop ASTEC’s ergonomics and man-machine interface to make the system as user-friendly as possible for biologist users. Participate in its deployment on the MRI imaging facility.
– Train biologist users in the use of the tool and set up and maintain a website presenting the system and its documentation to the scientific community.
– Improve ASTEC’s performance after finding its limits and the most important imaging parameters for the successful segmentation and tracking of cells.
Context:
The work will be carried out mainly within the tunicate development research team in Montpellier, Southern France. Frequent interactions will take place with the core facility’s engineers and with the computer scientists developing the core of ASTEC in the Morpheme, ICAR and MOSAIC teams.
Prerequisites:
– Undergraduate studies level in computer science, bioinformatics, applied mathematics, image analysis or physics
– Good knowledge of Python
– Knowledge in statistics
– Interest in biological image analysis
– Some training in biology would be a plus, but is not required
– A sufficient level in written / spoken English to interact daily with non-French speaking scientists. French knowledge not required
Contact:
Apply through the CNRS employment website. An informal contact can be established beforehand with the scientist in charge of the project, Patrick Lemaire, CRBM, patrick.lemaire@crbm.cnrs.fr
The laboratory of Dr. Angelo Iulianella is seeking a graduate student and postdoctoral research scientist to study the establishment and maintenance of cell types identities during neural development. The ideal candidates will have experience in molecular biology, cell culture, microscopy, transcriptional profiling, and neural development. The positions are available from early 2020, although start time is flexible. For postdoctoral candidates, funding is available for 3 years and recent graduates are encouraged to apply.
About us: Dalhousie University is one of Canada’s leading research-intensive universities. Our lab is part of the multidisciplinary Brain Repair Centre (http://www.brainrepair.ca) and Atlantic Mobility Action Project (www.amap.ca), which seeks to understand the development and repair of the neural circuitry of movement. We are situated in a highly collaborative environment with access to confocal and super-resolution microscopy, cytometry, and proteomic facilities.
About Halifax, Nova Scotia: With a population of 400,000 people, Halifax is the capital city of Nova Scotia and the educational, cultural and economic hub of Atlantic Canada. Discover rugged shorelines, sandy beaches, and hiking trails all within reach of the urban centre. Achieve your ideal work-life balance in a beautiful part of the world, while doing amazing science!
Please forward your CV, a statement of your research interest, and reference information by e-mail to:
Cell-cell communication plays a central role in the coordination of morphogenesis and fate specification. Most components of the major signalling pathways have been identified. We however lack a quantitative understanding, in time and space, of the dynamics of signal transduction from the membrane to the nucleus. The CRBM Tunicate embryology Research Team uses molecular and 3D + time live imaging approaches to study this process during ascidian embryogenesis.
The PhD project:
One of our major projects combines experimental and mathematical modelling approaches to produce a quantitative model of the information flow between membrane and nucleus for two major signal transduction pathways.
We are looking for a PhD student to develop an optogenetic control strategy for the FGF/SOS/Ras/ERK and Eph/RasGAP/Ras signaling pathways. This approach will open the way to a variety of questions including: how long does signal transduction take from the membrane to the nucleus? during which phase(s) of its cell cycle is the cell competent to respond to receptor activation? what is the minimum activation time of the receptor needed to produce a stable nuclear response? what is the function linking the activation level of the receptor and that of ERK? The experimental results will be integrated into a mathematical model, in collaboration with theoreticians, which will provide suggestions for further experiments.
Training:
This project is mostly experimental. It will give the selected student a solid expertise in embryology (microinjections, in vitro fertilization…), signal transduction and advanced light-sheet imaging. In addition, the PhD student will frequently interact with our computer science collaborators, the MOSAIC and ICAR teams. Participation in public outreach actions (Science festivals, My Thesis in 180 seconds, …) will be encouraged.
Necessary skills:
Master training in cell biology or development, with a strong interest for embryonic development
An interest in mathematical modelling (no specific mathematical knowledge needed).
A first experience in molecular cloning and confocal/light-sheet microscopy of live samples would be appreciated.
An experience in RNA or proteins microinjection into oocytes would be a plus but is not required.
No Knowledge of French required. Working knowledge in written / spoken English needed.
Application:
This project can be joined directly as a PhD student in fall/winter 2019, or as a Master intern in winter 2019, the PhD only starting in fall 2020. Funding is for 3 years.
The host research team is located at a major Cell Biology institute in Southern France, the CRBM (CNRS /U. Montpellier). All seminars and meetings are in English. The institute has a very well-equipped Imaging core facility, hosting a Luxendo MuViSPIM microscope on which lightsheet microscopy experiments will be carried out.
References linked to the project:
Leggio, B; Laussu J; Carlier, A; Godin, C; Lemaire, P and Faure, E (2019) MorphoNet: An interactive online morphological browser to explore complex multi-scale data. Nat Commun. 10(1):2812
Guignard*, U.-M.Fiuza*, B. Leggio, E. Faure, J.Laussu, L. Hufnagel, G. Malandain, C. Godin#, P.Lemaire# (2017) Contact-dependent cell communications drive morphological invariance during ascidian embryogenesis. bioRxiv 238741 https://www.biorxiv.org/content/early/2017/12/24/238741
U-M Fiuza, T. Negishi, A. Rouan, H. Yasuo, P. LemaireNodal and Eph signalling relay drives the transition between apical constriction and apico-basal shortening during ascidian endoderm invagination (2018) bioRxiv 418988 https://www.biorxiv.org/content/early/2018/09/15/418988
Lemaire P. (2011) Evolutionary crossroads in developmental biology: the tunicates, Development, 138(11):2143-52.
Tassy, O., Daian, F., Hudson, C., Bertrand, V., Lemaire, P. (2006) A quantitative approach to the study of cell shapes and interactions during early chordate embryogenesis. Current Biology 16:345-58
Postdoctoral Position in applying light sheet microscopy to understanding algal-spotted salamander endosymbiosis
A postdoctoral position is available in the laboratory of Dr. David Matus at Stony Brook University to investigate symbiotic and developmental processes with Selective Plane Illumination Microscopy. We have funding for a minimum of one year of postdoctoral support to develop protocols for extensive in vitro and in vivo imaging of cell invasion processes with a focus on the tissue and cellular entry of endosymbiotic algae as they enter their spotted salamander embryo hosts (Ambystoma maculatum). This work is funded by the Gordon and Betty Moore Foundation in collaboration with researchers from University of Arizona, the Bigelow Marine Laboratory, and Gettysburg College (see Shelf Life Episode 11). The project will combine molecular biology, embryology, cell biology, and extensive light sheet imaging. Preferred candidates will have backgrounds in light sheet and/or confocal microscopy, working with modern tissue clearing and staining methodologies and interests in advanced imaging methods at the intersections of cell and developmental biology with ecology and evolution. My laboratory is a part of a modern and well-equipped Department of Biochemistry and Cell Biology at Stony Brook University on Long Island, NY. For further information on our work, please see the following publications on techniques,cell invasion, and the symbiosis.
To apply, please send a letter of interest detailing your expertise, CV and names and contact information of three references to david.matus@stonybrook.edu and apply to the advertised position here.
The Developmental Biology Group (Prof. I. Lohmann) at the Centre for Organismal Studies (COS) in Heidelberg, Germany is looking for a
POST-DOC
to study the role of HOX transcription factors in controlling cellular plasticity and cell fate maintenance.
You will join the Lohmann lab (http://ilohmann-lab.org), which is located at the Centre for Organismal Studies (COS) at the University of Heidelberg in Germany, and studies the role of HOX transcription factors in defining cell type identities using Drosophilaas a model.
Cell fates are controlled by networks of transcription factors (TFs) that activate transcriptional programs realizing the distinct properties of cells of a given cell types.However, how TFs control different cell fates is still un unsolved question. HOX TFs represent an excellent model to address this fundamental problem, since they are broadly expressed yet perform highly specific functions within different cell types. We have previously shown that Hox TFs stabilize cell fate choices by suppressing the multipotency encoded in the genome via the interaction with the Polycomb complex. We now seek for enthusiastic new colleagues to analyse our hypothesis that elimination of the Hox code, which is maintained throughout the lifetime of an organism, results in “memory-less” naïve cells that are easy to reprogram.
The Heidelberg Molecular Life Science Community offers a vibrant molecular research community, as well as state-of-the-art core facilities.
Successful candidates should have experience in genomic approaches like RNA-seq, ChIP-seq, ATAC-seq, analysis of genomic data, confocal microscopy, advanced immunohistochemistry and possibly in Drosophila genetics and molecular biology. The ability to quickly integrate into an interdisciplinary team and work independently within an academic research environment is essential. The position is immediately available, the salary is according to TV-L regulations. Disabled persons with comparable skills will be preferentially considered.
 (No Ratings Yet)
(No Ratings Yet)
 (3 votes)
(3 votes)