The Department of Biology within the College of Natural and Health Sciences at The University of Tampa invites applications for a full-time tenure-track position in developmental biology beginning in August 2019.
The University of Tampa is a medium-sized, comprehensive, residentially based private institution of 9,200 undergraduate and graduate students. The University is ideally situated on a beautiful 110-acre campus next to the Hillsborough River, adjacent to Tampa’s dynamic central business district, which is a growing, vibrant, diverse metropolitan area. UT reflects this vibrancy; with 22 consecutive years of enrollment growth UT boasts 260 student organizations, a multicultural student body from 50 states and 140 countries and “Top Tier” ranking in U.S. News & World Report.
Teaching responsibilities will include introductory biology for majors, an upper division course in developmental biology, and other courses as needed. The department is interested in attracting a broadly trained biologist with expertise in developmental biology. The candidate is expected to engage in research activities that involve undergraduates and yield peer-reviewed publications. Limited start-up packages and modest research space are available for tenure-track positions.
PhD required, ABD candidates considered, with prior teaching and research experience with undergraduates desirable.
Review of applications will begin October 10, 2018, and continue until the position is filled.
For further details and to apply, please visit our website at www.ut.edu/jobs
The University of Tampa is an equal opportunity/affirmative action employer dedicated to excellence through diversity and does not discriminate on the basis of age, race, sex, disability, sexual orientation, national origin, religion, marital status, gender identity, veteran status or any other non-job related criteria. The University of Tampa recognizes the importance of a multicultural community of students, faculty, and staff who seek to advance our commitment to diversity. The University invites applications from underrepresented groups and those who have academic experiences with diverse populations.
We are looking for ambitious and motivated postgraduate candidates to join the new computational biology group, led by Dr. Linus Schumacher, at the MRC Centre for Regenerative Medicine in Edinburgh. The group is using expertise in mathematical modelling of cellular processes in development and collective behaviour in biological systems, with a recent focus on stem cells and regeneration.
There is one fully-funded position (UK/EU/overseas) focussing on mathematical & computational modelling of stem cell populations in regeneration and experimentally relevant predictions. Find out more here: https://www.findaphd.com/search/ProjectDetails.aspx?PJID=101556
Further opportunities are available through the University of Edinburgh’s Wellcome Trust funded Tissue Repair postgraduate training programme. Students in this programme will have the opportunity to do research rotations and PhDs with our group (in joint supervision with experimental labs).
This is an opportunity to conduct your PhD research on mathematical and computational biology, embedded in a world-leading centre for multidisciplinary research in mammalian stem cell biology and regenerative medicine. Find a list of all PhD opportunities here: http://crm.ed.ac.uk/join-us/phd-training/phd-opportunities
The MRC Weatherall Institute of Molecular Medicine (WIMM) has fully funded 4-year Prize PhD (DPhil) Studentships available to start in October 2019. These Studentships are open to outstanding students of any nationality who wish to train in experimental and/or computational biology.
The Institute is a world leading molecular and cell biology centre that focuses on research with application to human disease. It includes the recently opened MRC WIMM Centre for Computational Biology and houses over 500 research and support staff in 50 research groups working on a range of fields in Haematology, Gene Regulation & Epigenetics, Stem Cell Biology, Computational Biology, Cancer Biology, Human Genetics, Infection & Immunity. The Institute is committed to training the next generation of scientists in these fields through its Prize PhD Studentship Programme.
The fully funded studentships include a stipend of £18,000 per annum and cover University and College fees.
Further information on the studentships, how to apply, and the projects available can be found at:
Studying limb regeneration in model organisms is important for the advancement of regenerative medicine in humans. We set out to study regeneration in the hind limbs of the African Clawed Frog Xenopus laevis – this animal is able to regenerate its hind limbs very early in development, but it loses this ability during metamorphosis. Additionally, there is a large and still growing body of evidence suggesting that charged particles called ions (for example, sodium, potassium, and chloride) are important for regulating pathways that control growth and regeneration. Though a relatively small number of biologists’ work is centered around this field, called bioelectricity, its implications thus far in the fields of developmental biology and regenerative medicine have been compelling. As an undergraduate, I had the privilege to be trained and to perform research in a lab that is at the forefront of this field, which had many unanswered and yet unresearched questions.
We – being myself and my team of mentors, Dr. Patrick McMillen and Professor Michael Levin, who were both Tufts undergraduates ten and twenty years before me, respectively – married the concept of bioelectrics and Xenopus hind limb regeneration to ask the question: what bioelectric changes do younger, regenerating froglets exhibit in response to amputation that older, non-regenerative tadpoles do not? This is part of the lab’s mission to discover regenerative therapies, based on manipulating bioelectric signaling, for various biomedical applications. We amputated the limbs of both regenerating tadpoles and non-regenerating tadpoles, and soaked them in a solution containing a molecule that glows in response to changing ion concentrations across cell membranes, also called membrane potential. By the time we were ready to analyze the results of this work, the product of observation and the scientific method led us to a bigger, more perplexing question to answer.
In performing these initial experiments, I did what I believed was my due diligence as a researcher: I needed controls! So I used the contralateral, uncut limbs of the froglets as controls, a common method used across many fields, to make sure the dye wasn’t being randomly soaked up by cells and to establish a baseline background for what the fluorescence would look like in un-injured, intact tissues. In fact, I quickly found that the dye was being taken up by cells in the contralateral limb, but only in the intact limbs of frogs that had the other limb amputated. Un-amputated froglets did not exhibit staining from the dye in either limb. I took this information to my mentors, sure that they would already have an explanation prepared, as professors and teachers before them always had when a question arose in lecture. For the first time in my academic career, nobody had an answer for me. This phenomenon had never been observed before, therefore nobody had asked the question; there was truly no one with an answer.
It is at this point in many young scientists’ careers that the potentially interesting project is swooped out from under them, their mentors realizing the value of the new findings, hungry for the credit. For others, their mentors do support intellectual curiosity and give them the freedom to pursue their own projects, but their projects do not see the light of day because there simply are not enough resources. For these reasons, the Tufts biology department, and more specifically, the Levin lab, are extremely unique. I could not have been more fortunate, because I was given both the freedom to head my own project and the resources to do so.
Ultimately, we found that the contralateral limbs of regenerating tadpoles glowed (in the presence of that molecule I mentioned earlier) in a region on the contralateral limb that very closely mirrors the plane of amputation. Our data revealed that the un-injured limb somehow knows the relative location (and even type) of injury, within about 30 seconds (Busse et al.Development 2018; doi:10.1242/dev.164210). This phenomenon has interesting implications for regenerative medicine. What we found is a distal region that not only recognizes, but also encodes information about an injury incurred by the body. If similar signaling phenomena can be found in mammalian systems, there is potential for the development of surrogate site diagnostics – looking at one site to decipher information about the health of another. This information is also evidence that contralateral limbs are not an appropriate control for experiments, and hopefully encourages everyone to think twice before using them as controls!
There are many more questions that we intend to answer in the future; for example, how does one part of the body sense that another part of the body has been injured? In the meantime, the context of this research is extremely important for anyone in any field of research to understand. I asked a question that nobody at the time could answer, and was given the opportunity to find an answer. The result was that I learned more collaborating with Dr. Patrick McMillen, Professor Michael Levin, and many others in the Levin lab than any biology course could have taught me. Many students are prompted to spend time answering questions throughout the duration of their degree, but I was encouraged to ask them, and it has fundamentally changed the way I think about learning. I had teachers who were not eager to prove what they knew, but rather who were eager to teach how they knew it.
The Turing Centre for Living Systems is seeking a highly motivated research engineer to fill the post of ‘leading data scientist/data analyst’, within CENTURI’s technology transfer platform. This position will be a central point of data analysis, data sharing and code sharing of our research community.
Three-year contract from the Aix-Marseille Université.
Deadline for Application: November 23
Applications should include:
a CV (including a list of publications)
a cover letter (describing past experience with data analysis)
Applications are open for the Wellcome Trust funded four year PhD programme in Developmental Mechanisms at the University of Cambridge. We are looking for talented, motivated graduates or final year undergraduates, and are keen to attract outstanding applicants in the biological sciences, who are committed to doing a PhD. We are able to fund both EU and *non-EU students.
Closing date: Thursday 3 January 2019 (by 12:00pm midday UK time)
For more details about the application process and the programme please see the website:
A postdoctoral position is available in the laboratory of Dr. Sophie Astrof at Rutgers University to study roles of cell-extracellular matrix (ECM) interactions in cardiovascular development and congenital heart disease. We have recently discovered that progenitors within the second heart field (SHF) give rise to endothelial cells composing pharyngeal arch arteries (Dev Biol 421:102–111, 2017). Projects in the lab focus on the role of ECM in regulating the development of SHF-derived progenitors into endothelial cells and their morphogenesis into blood vessels. The successful candidate will combine genetic manipulation, embryology, cell biology, and confocal imaging to study molecular mechanisms by which cell-ECM interactions and tissue microenvironment regulate cardiovascular development. Additional projects focus on the investigation of cell type-specific and cell-autonomous functions of fibronectin in development and signaling (Development 143:88-100, 2016). Interested candidates should send their CV and the names of three references to Sophie.astrof@rutgers.edu
Jeff Rasmussen tells the story behind his recent paper from the Sagasti Lab in Dev Cell.
This project began as an extension of my earlier postdoc work in Alvaro Sagasti’s lab investigating removal of axon debris following skin injuries in the larval zebrafish [1] and led me into scientific territory that I never anticipated. It is a story that would not have happened without open-mindedness, encouragement and—most importantly—help from colleagues in the fish community.
Sensory axon endings profusely innervate the skin, and skin injuries trigger axon degeneration. We previously discovered that keratinocytes are the primary phagocyte for degenerating axons in larval skin. But what cells eat axons that degenerate in the more complex adult skin?
In order to answer this question, I first needed a way to visualize sensory axons in adults. Most work in zebrafish has focused on the larval system, so markers for adults have lagged behind. Luckily, I came across a transgenic line made by a previous graduate student in Alvaro’s lab [2] that showed bright and specific expression of sensory axons in adults. The pattern of adult skin innervation revealed by this line caused this project (and my career) to take a twist.
A Striking Axon Pattern
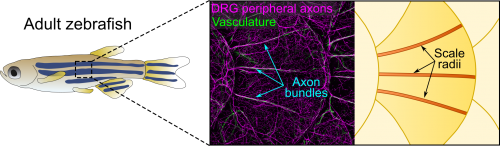
Axons of Rohon-Beard neurons innervate most of the larval fish skin. Rohon-Beard axons in the skin rarely bundle (fasciculate). By contrast, this transgenic line revealed that axons of dorsal root ganglion (DRG) neurons, which innervate adult skin, were frequently bundled. More surprisingly, these bundles were evenly spaced across the surface of scales, which underlie the adult epidermis (Figure 1). Alvaro whole-heartedly encouraged me to dig deeper into what created these evenly spaced bundles, despite his lab having no experience working with post-embryonic stages.
Figure 1. Anatomy of adult zebrafish skin showing axons, vasculature and scale radii.
Thi’s Developmental Studies
Thi Vo, an undergraduate researcher in Alvaro’s lab, took on the challenge of analyzing post-embryonic development. Because many Rohon-Beard neurons die after only a few days of development, we expected that DRG axons would innervate the epidermis shortly thereafter. However, by staining isolated scales to visualize axons, Thi found that the axon bundles only appeared during late juvenile stages. So Thi next analyzed mutants lacking DRG neurons and found that the bundles never formed, proving that they were indeed DRG axons.
Thi also stained scales with phalloidin to visualize cell morphology and found that a set of elongated cells apparently presaged the path of the axon bundles. What could these cells be?
Lindsey’s Schwann cells
We initially hypothesized that the elongated cells were Schwann cells, the glia of the peripheral nervous system. Schwann cells are neural-crest derived, and Shannon Fisher’s group recently made a neural crest lineage reporter [3], which we learned was growing in Gage Crump’s lab at USC. To determine if the line labeled Schwann cells in adults, I made a trip across town to visit Lindsey Barske, a postdoc in the Crump lab, who was working with the neural crest reporters. One of Lindsey’s lines labeled Schwann cells coating the axon bundles, showing that they are nerves and giving us a key reagent to visualize their development. But when Thi and I analyzed the timing of Schwann cell migration, they appeared too late to pioneer the path of the DRG bundles, ruling out this hypothesis.
Michael’s Vessels
After hearing about our work at a Southern California Zebrafish Meeting, Michael Harrison, a postdoc in Ellen Lien’s lab working on heart and vascular development, suggested we analyze several of his chemokine mutants. Thi and I took a bus trip down Sunset Blvd to CHLA to collect scales from these mutants. Although this effort was ultimately fruitless, as luck would have it, Michael’s mutants expressed multiple transgenic reporters. One of these transgenes was fli1a:EGFP (a vascular reporter made by Brant Weinstein’s group) [4], which revealed that the axon bundles tightly associated with blood vessels (Figure 1). This was unexpected because we had initially ruled out a vascular contribution based on analysis of a vessel reporter not as broadly expressed as fli1a.
Intriguingly, axons and vessels also tightly associate in mammalian skin and axons promote vascular remodeling and arterial differentiation in mouse [5]. Were blood vessels the elongated cells that arose early in scale development? No, since we found that blood vessels only appeared along mature scales, once animals reached adulthood. What about the converse: did axons pattern the vessels? By again analyzing mutants lacking sensory neurons, we found that blood vessels appeared normal. Thus, in contrast to mammals, skin nerves and vasculature are independently patterned in fish. This was an important finding but, once again, the identity of the pioneering cells remained elusive.
Sandeep’s Osteoblasts
The scale surface is made from bone and contains a number of striking patterns. Remarkably, we noticed that the axon bundles and vessels aligned with scale radii, grooves in the bone that radiate from the scale center (Figure 1). Could osteoblasts, bone-forming cells, or osteoclasts, bone-degrading cells, guide axons and vessels? We first examined osteoclasts but found no evidence that they form the radii. Next, I made another crosstown trip to the Crump lab—this time with the help of Sandeep Paul—to look at osteoblast reporters. Sandeep’s lines showed that osteoblasts line mature radii, as suggested by pioneering ultrastructural studies [6, 7]. Imaging osteoblasts early in scale development revealed that they create the radial paths by polarized migration.
To test if osteoblasts promote skin innervation during regeneration, we used an inducible osteoblast ablation line made by Ken Poss’ group [8] and found that blocking scale regeneration by osteoblast ablation resulted in a reduction of axon density. To test if scale development similarly promoted innervation, we analyzed mutants that prevent scale development [9, 10]—provided as part of a “scale care package” by Matt Harris’ lab. These mutants had reduced skin innervation and vascularization, showing that scales are also required during ontogeny.
Figure 2. Intubation allows extended live-cell imaging of scale development and regeneration.
Full Scale Ahead
Although I never planned to work on scales as a model system, I am really excited about the potential for these mini-organs to reveal the cellular and molecular basis for cell type patterning during skin development and repair—questions that I will be pursuing in my newly formed research group at the University of Washington. Scales are evolutionarily related to other types of specialized skin appendages like feathers and hair. Thus, studies of fish scales may reveal general mechanisms for coupling organ maturation and growth to skin patterning. Scales may also yield insights into bone-nerve interactions that occur in diverse tissues, like antlers, teeth and long bones. An increasing number of genetic tools, together with advances in live-cell imaging of post-embryonic stages (Figure 2), suggest the future is bright for a resurgence of scales as a model system.
1 PhD Position : Characterization of Hedgehog morphogens in vitro and in vivo
Hedgehog (Hh) morphogens play important roles in development and cancer, but their mode of extracellular transport to target cells is only poorly understood. Thus, we aim at the characterization of various unusual posttranslational regulatory mechanisms in Hh biology, such as Hh multimerization on the surface of secreting cells via structural and biochemical analysis of Hh clusters, and the unusual mode of Hh transport and gradient formation.
We use a wide range of biochemical methods, such as recombinant protein production and chromatographic/functional characterization of proteins and Heparan sulfate-proteoglycans (HSPGs), determination of Shh/HSPG binding and in vivo testing of any obtained models of protein association and transport (In Drosophila melanogaster).
We invite applications from highly qualified and motivated students of any nationality. The applicant will hold a Masters degree (Biology, Chemistry, Biotechnology, Pharmacy or Biochemistry) and has gained first biochemical research experience. Experience in Drosophila experimentation is highly welcome, but not required. The successful applicant will find strong support within the excellent interdisciplinary environment of the SFB1348 of the University of Muenster.
CENTURI is recruiting up to 10 highly motivated postdoctoral fellows to work in an interdisciplinary life science environment. The available postdoctoral projects will be advertised online, with more projects being added to our website up until November 15.
CENTURI postdocs will have the opportunity to develop their project with more than one research group, bridging biology to other disciplines.
Candidates will have a background in any of the following fields: cell or developmental biology, immunology, neurobiology, biophysics, theoretical physics, computer science, bioinformatics, applied mathematics or engineering.
Candidates can either apply to one of the advertised CENTURI projects or submit their own project, providing that they meet the application criteria and that their application is supported by at least one host lab. These projects should be submitted to: postdocproject@centuri-livingsystems.org
Deadline for application and project submission: November 30, 2018
About CENTURI
Mainly located on the Luminy science campus (Marseille France), the Turing Centre for Living Systems is an interdisciplinary project federating a growing community of biologists, physicists, mathematicians, computer scientists and engineers in 15 research institutes.
The project focuses on 3 missions: Research, Education and Engineering
 (No Ratings Yet)
(No Ratings Yet)

 (3 votes)
(3 votes)