In our recent paper published in Nature, we unravel a new mechanism of an extracellular matrix protein secreted by muscle satellite (stem) cells, thereby playing the unusual role of acting as a signaling molecule to maintain the stem cell population. Here, I share the story behind this discovery and discuss the questions related to niche regulation.
In a ChIP-sequencing screen in myogenic cells performed by the Tajbakhsh and Stunnenberg labs (Castel et al., 2013) to identify direct targets of nuclear NOTCH (NICD) and its downstream effector RBPJ, we found an enrichment of extracellular matrix proteins, among which were specific collagens genes including Collagen 5 (Col5a1 and Col5a3) and Collagen 6 (Col6a1 and Col6a2). I was in the first year of my PhD when Dr. Philippos Mourikis, an ex post-doc in the Tajbakhsh lab, asked me to help with some collagen immunostainings on embryonic muscles over-expressing NICD (E17.5 Myf5Cre; R26NICD) (Mourikis et al., 2012). We nicely showed that both COLV and COLVI secretion were upregulated upon Notch induction.
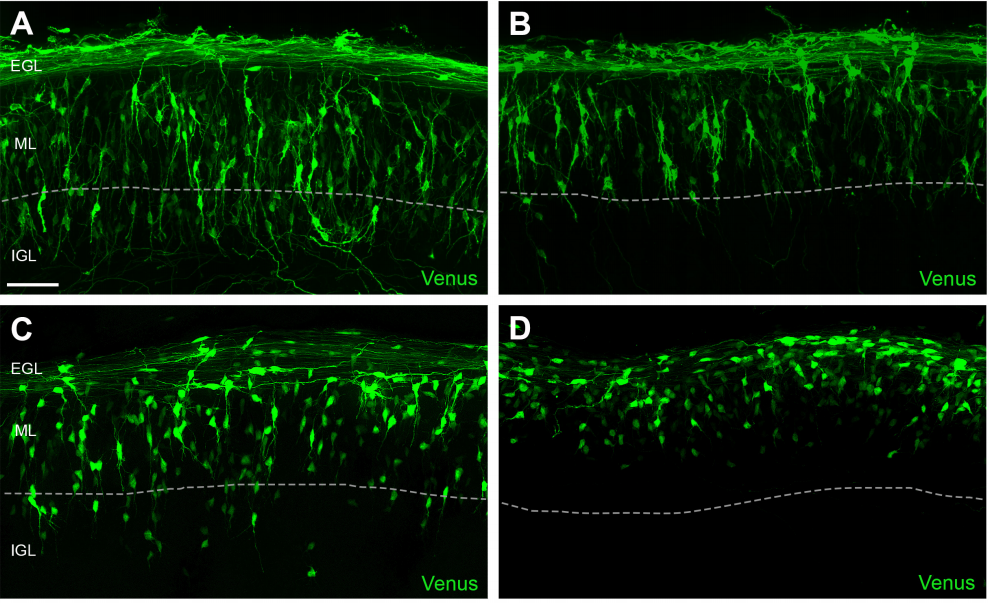
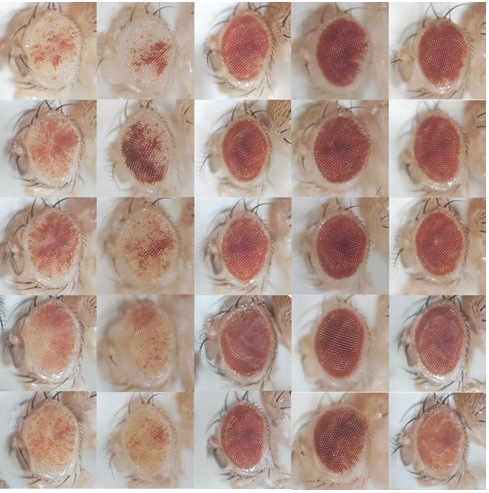
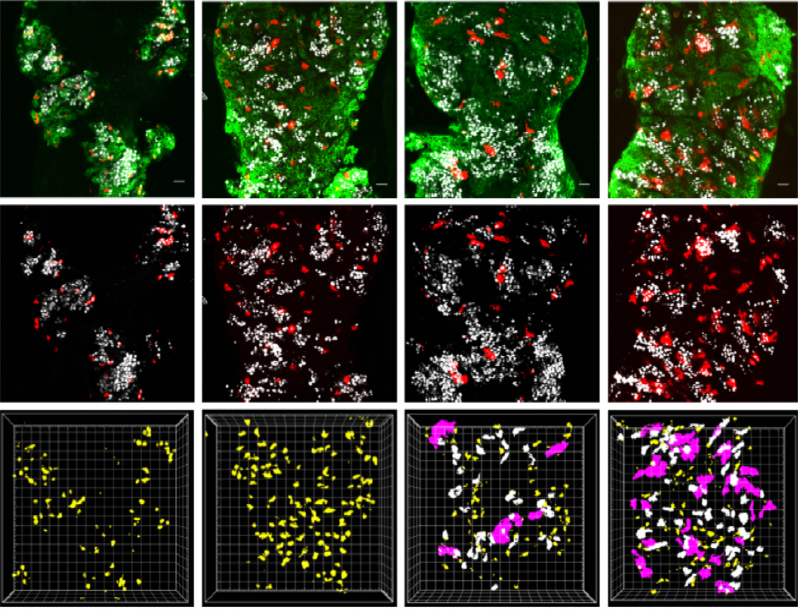
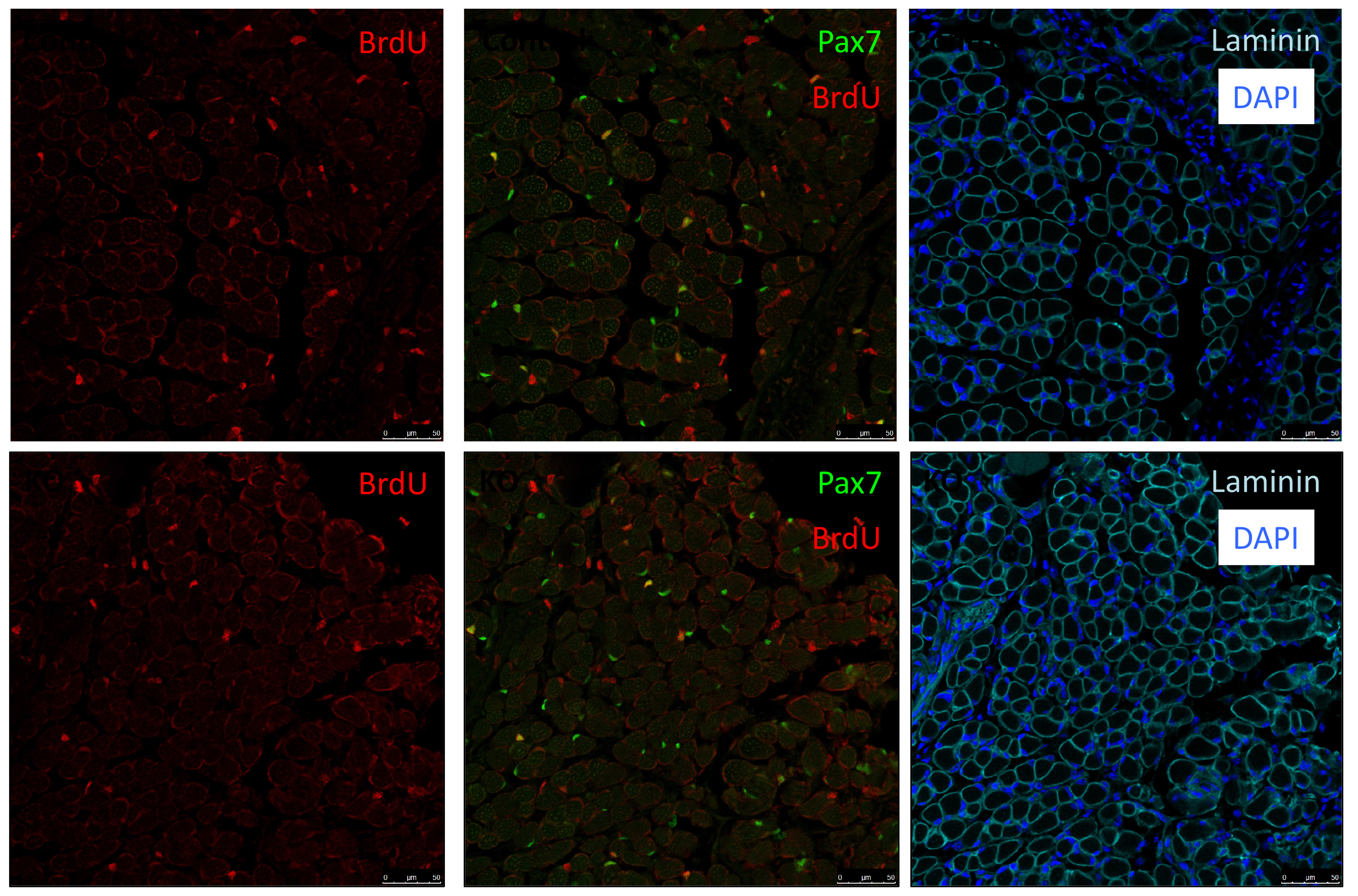
As Col5a3 gene was the most responsive gene to Notch activity, both in vitro and in vivo, we thought that Col5a3 KO mice might present a muscle phenotype (Huang et al., 2011). In collaboration with Dr Guorui Huang from the Greenspan lab, we analyzed the number of satellite cells (PAX7+) in adult and postnatal P8 pups muscle sections. Unfortunately, we could not observe any alterations in stem cell number nor proliferation capacity (BrdU pulse).
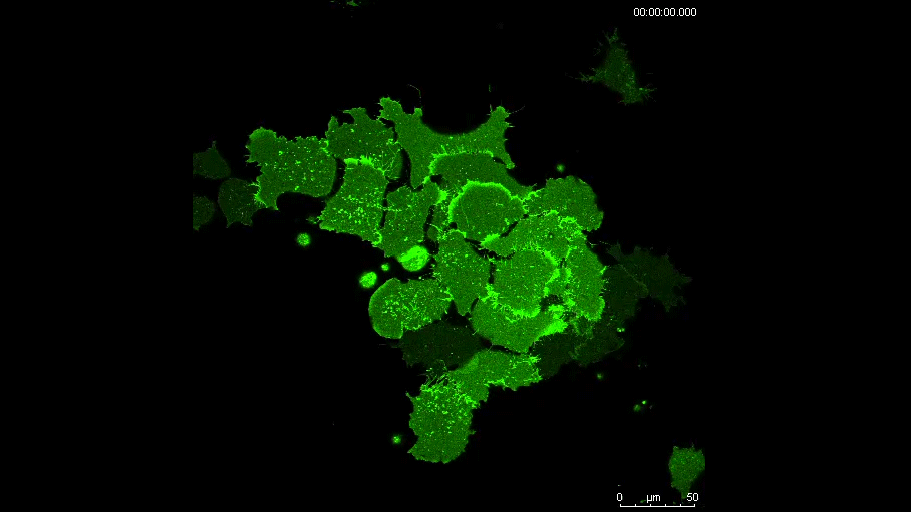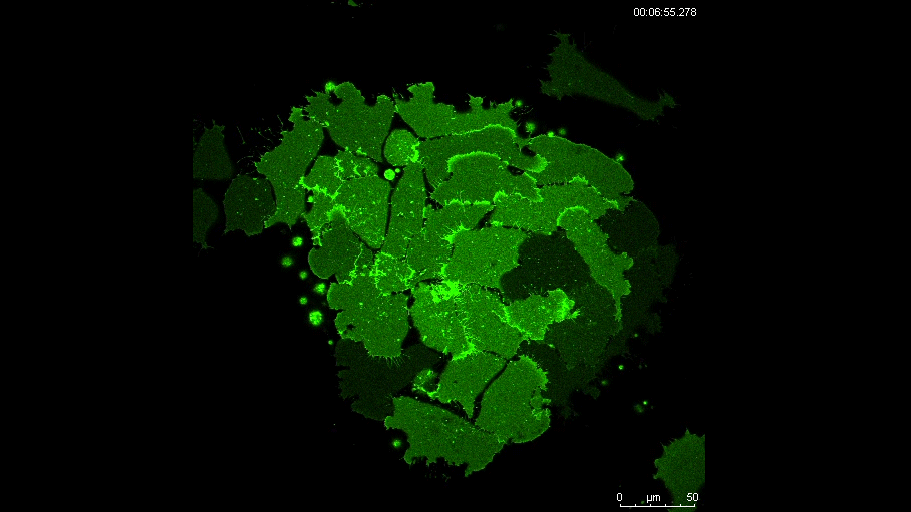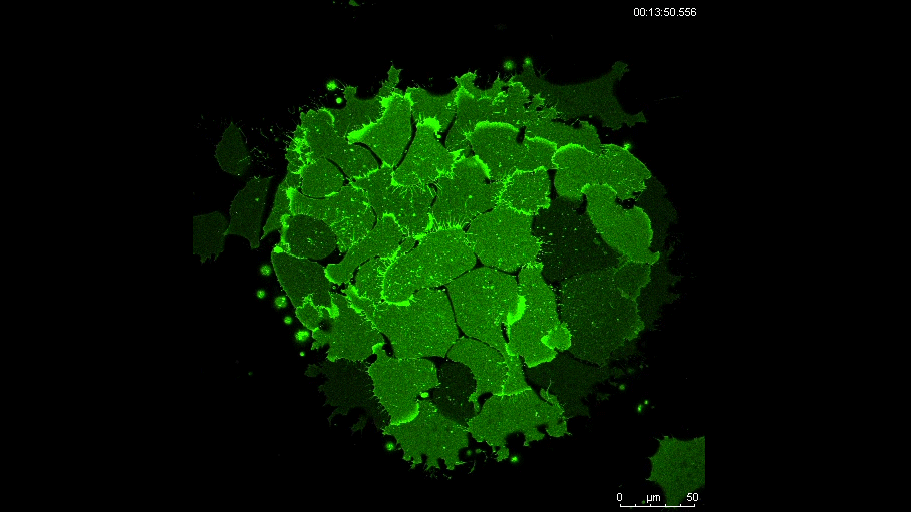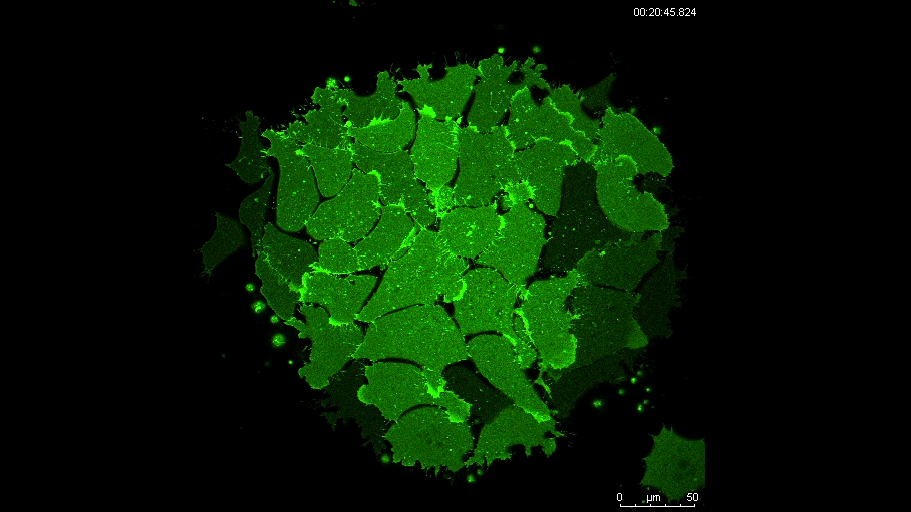
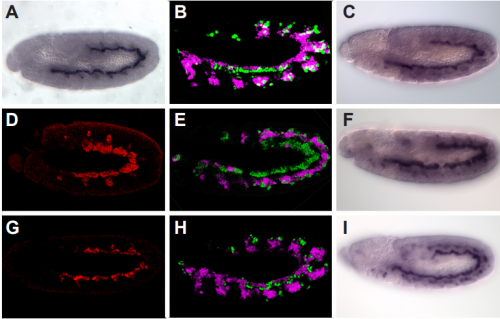
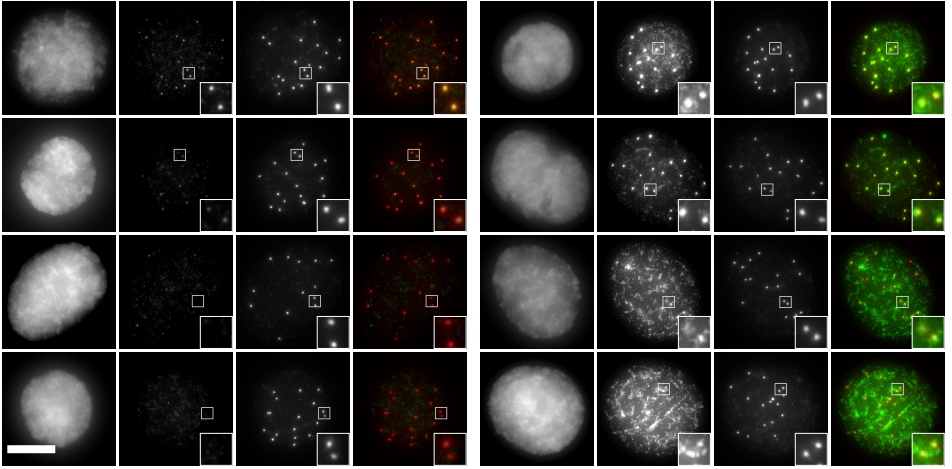
PAX7 (satellite cells), BrdU (proliferation) and Laminin immunostaining on transverse sections of Tibialis Anterior muscle of Control (WT) and Col5a3 KO P8 (8 days after birth) juvenile mice. No difference in satellite cells number and proliferation capacity between control and mutant pups.
As my initial PhD project was on hold due to mice breeding issues, we decided that I could perform a simple culture experiment to close up the collagen story. We performed two sets of experiments: in the first, we added different collagens in soluble form on cells plated on gelatin-coated dishes; in the second, cells were cultured directly on collagen-coated dishes. Surprisingly, only COLV in its soluble form retained cells in a more stem-like state delaying their proliferation and differentiation. With these unexpected results, we realized that COLV might be acting as a signaling molecule rather than a biomechanical cue. As collagen molecules are known to bind to specific receptors such as Integrins or Discoidin Receptors (Leitinger, 2011), we tested whether their inhibition could abrogate COLV anti-myogenic effect. To our disappointment, COLV was still able to delay myoblast differentiation even when these receptors were inhibited.
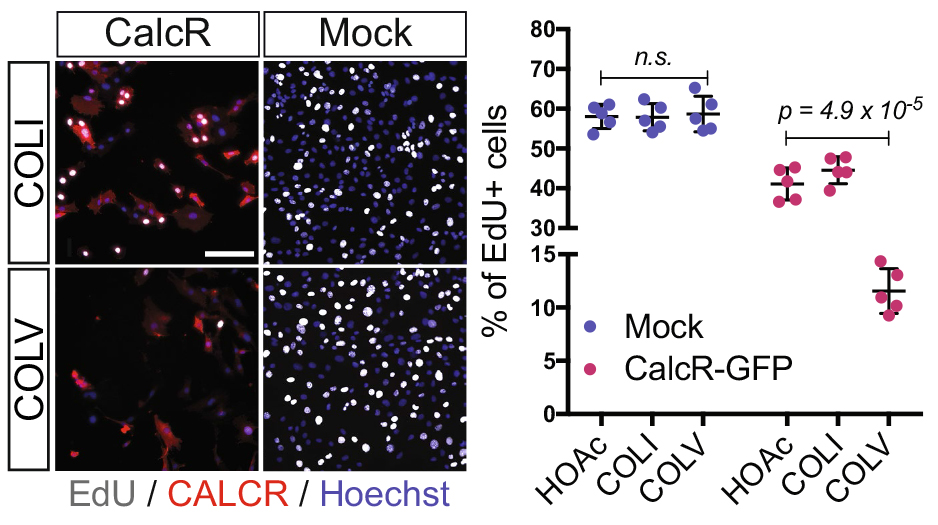
After looking at the results from several angles, we came to the conclusion that we lacked a mechanism and we should probably wrap up the story with what we had, so that I could go back to my initial PhD project. With a sense of unfinished business, we started gathering the data into figure formats. We were then struck by an old RT-qPCR assay on satellite cells that were treated with the different collagens, where Calcitonin receptor (CalcR), a known satellite cell quiescence marker, was upregulated more than 20-fold. We remembered that CALCR is a G-protein couple receptor (GPCR) and found in vitro evidence that some collagens could indeed bind to GPCRs (Luo et al., 2011; Paavola et al., 2014). This was a major turning point in the project, and the path was wide open for detailed mechanistic studies.
In parallel on the other side of the world, our collaborator Dr. So-Ichiro Fukada in Osaka had just a few weeks earlier published a very nice study showing that specific ablation of Calcr in muscle stem cells led to their spontaneous exit from the quiescent niche (Yamaguchi et al., 2015). Using their retroviral tools to overexpress CALCR and Calcr null muscle stem cells, we were able to demonstrate that CALCR was necessary to respond to COLV. This was too exciting, my other project had to wait!
EdU (2h pulse) and CALCR staining of GFP+ C2C12 cells isolated by FACS and transduced with Calcr-GFP or mock-GFP retrovirus and cultured for 24 h with COLI (top) or COLV (bottom). Quantification of EdU+ Calcr-transduced C2C12 cells or mock-GFP cells treated for 24h with COLV or with the controls, COLI and HOAc. There was no significant difference between HOAc and COLI treated samples.
On the other hand, these results were puzzling…CalcR already has its ligand: Calcitonin hormone produced by the parathyroid gland, suggesting that satellite cell quiescence is under systemic control. Why would the regulation of muscle stem cells that are dispersed throughout muscle masses in the body – and that would be solicited with different kinetics – be under systemic control? Can COLV act as the local ligand of CalcR for muscle stem cells?
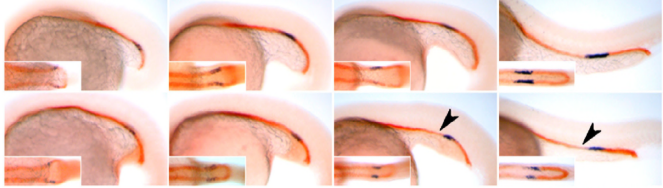
Meanwhile, our first priority was to assess whether the loss of function of COLV specifically in satellite cells using Tamoxifen-inducible Tg:Pax7-CreERT2; Col5a1flox mice could affect their behavior. Another unexpected surprise was the observation that only 18 days following recombination, satellite cells escaped the quiescent niche, differentiated and fused to the pre-existing fibres.
Another consideration is the source of collagen in skeletal muscle – being predominantly from interstitial myofibroblasts, which deposit collagen between myofibres (Zou et al., 2008). We also found expression of Col5a-1, -2, and -3 in other cell types present in the muscle; namely Fibro-Adipogenic Progenitor (FAPs) and resident macrophages.
So why does this source of COLV not compensate for the loss of satellite cells-produced COLV?
Possible explanations include the basal membrane (BM) forming a physical diffusion barrier between the satellite cells and the interstitial space – rendering them blind this source of COLV. Although the myofibre that is in intimate contact with satellite cells under the BM could be a provider, we showed that myofibres do not express detectable COLV to sustain satellite cell quiescence.
Another possibility is that COLV produced by satellite cells has a specific configuration. In our siRNAs experiments on isolated myofibers, we observed that targeting of the Col5a3 isoform phenocopied siRNA against Col5a1, strongly suggesting that the active triple helix is the a3-COLV isoform composed of both a1 and a3 chains as an [α1(V)α2(V)α3(V)] heterotrimer. Although the presence of soluble COLV is difficult to demonstrate in vivo, we favoured the possibility that COLV produced by satellite cells rapidly binds to CALCR upon secretion, rather than being integrated in the collagen network. This would then explain why loss of COLV function exhibited a phenotypic loss of satellite cells within such a short period.
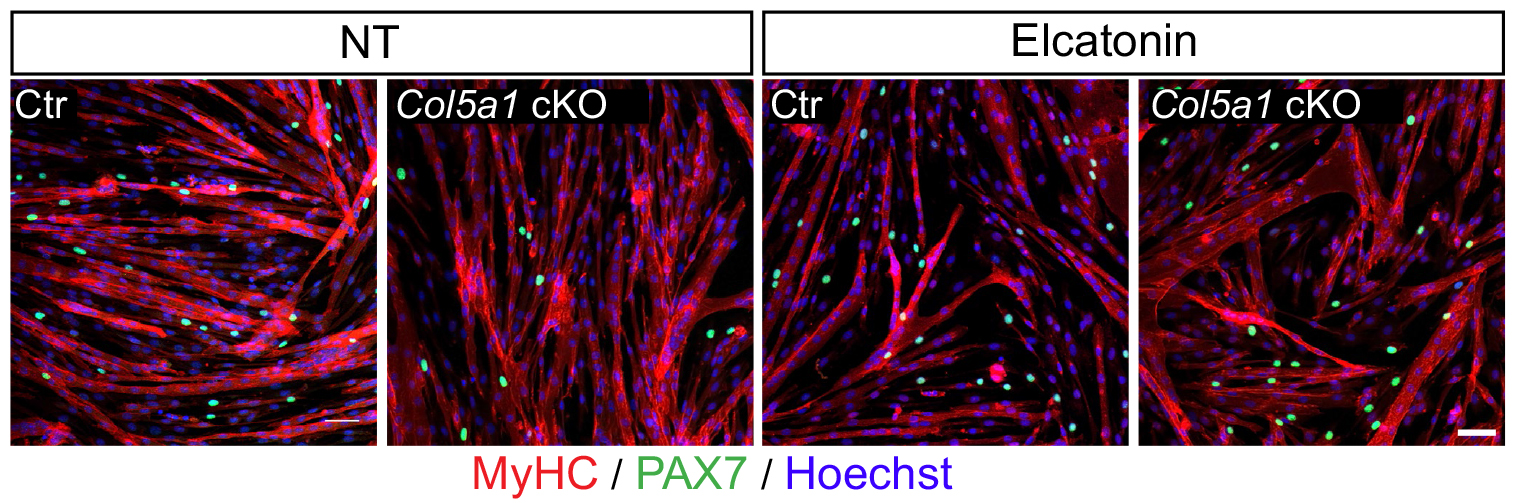
We then needed to demonstrate that the newly discovered Notch/COLV/CALCR axis is functional for the maintenance of quiescent cells in vivo. To do so, we injected Col5a1 cKO (Tg:Pax7-CT2; Col5a1flox) mice subcutaneously with a derivative of the known CALCR ligand calcitonin (Elcatonin). Interestingly, exogenous Elcatonin administration compensated for the lack of COLV and prevented their exit from quiescence, thereby validating our results linking COLV and CALCR. It also told us that endogenous calcitonin was not bioavailable in the muscle to rescue the loss of COLV. Interestingly, thyroidectomy in adult individuals does not show any striking overall pathological consequences (Russell et al., 2014); it would be interesting to study satellite cell dynamics in mice suffering from hyper/hypothyroidism.
An interesting outcome of the rescue experiments was that Elcatonin administration in control mice also showed an improvement in self-renewal capacity, suggesting that this could be a good therapeutical candidate for myopathies.
Rescue of loss of COLV by Elcatonin in an ex vivo self-renewal reserve-cell model, where PAX7+ non-proliferative cells return to quiescence. MyHC and PAX7 staining of Control (Ctr: Tg:Pax7-CT2; Col5a1+/+; R26mTmG) and Col5a1 null (Tg:Pax7-CT2; Col5a1flox/flox; R26mTmG) non-treated (NT) or treated with Elcatonin (Elcat). No GFP+/EdU+ cell (12h pulse) could be detected in any of the conditions indicating GFP+ cells are quiescent (data not shown).
In addition, it has been shown that the satellite cell population is heterogenous according to the muscle anatomical location and even within a single muscle (Rocheteau et al., 2012; Sambasivan and Tajbakhsh, 2015). Therefore, we cannot exclude the possibility that some satellites cells do indeed respond to calcitonin stimulation. Our results showing that Elcatonin administration induces a deeper quiescent state could support this notion. In addition, CALCR is expressed in about 80% of satellite cells, suggesting that 20% of them are most likely not under direct Notch/COLV/CALCR control.
While exploring the quiescence in muscle stem cells, we learned that when studying the role of collagens one should consider not only the traditional mechanotransduction role of these molecules, but also their potential role as signaling agents. Given that neural and intestinal, and perhaps other stem cells, are maintained by Notch signaling, it is possible that this model operates in stem cells located in other tissues and organs.
The Echeverri lab at the MBL seeks a highly motivated individual to join the Eugene Bell Center for Regenerative Biology and Tissue Engineering as a Postdoctoral Researcher. The successful candidate will work on the molecular mechanisms of scar free skin regeneration in axolotls.
The specific goal of the project is to examine the role of different cell types in responding to the injury cue and in later remodeling collagen.
Basic Qualifications:
Applicants should have a Ph.D. in a biology related field. Must have prior experience working in the field of cell and developmental biology, as well as experience with molecular biology. Must be independent, enthusiastic, self-motivated, productive, and enjoy working in a highly collaborative environment.
Preferred Qualifications:
The ideal candidate will have direct experience with working in vivo in an animal model. Previous experience with cell culture, molecular biology and imaging would be a plus.
Required documents:
Cover letter explaining specifically why you are interested in joining our lab to work on this project and what positive qualities you would bring to our team.
Curriculum vitae.
List of 3 references (Please do not have letters sent at this time. Letter writers will be contacted directly by the PI)
Please e-mail your application to Dr. Echeverri: echev020@umn.edu
The Echeverri lab at the MBL seeks a highly motivated individual to join the Eugene Bell Center for Regenerative Biology and Tissue Engineering as a Postdoctoral Researcher. The successful candidate will work on the comparative evolution of molecular regeneration in various aquatic research organisms. The specific goal of the project is to examine how pathways that are essential for regeneration have evolved in different species with different regenerative capacity.
Basic Qualifications:
Applicants should have a Ph.D. in a biology related field. Must have prior experience working in the field of cell and developmental biology, as well as experience with molecular biology. Must be independent, enthusiastic, self-motivated, productive, and enjoy working in a highly collaborative environment.
Preferred Qualifications:
The ideal candidate will have direct experience with working in vivo in an animal model. Previous experience with cell culture, molecular biology and imaging would be a plus.
Required documents:
Cover letter explaining specifically why you are interested in joining our lab to work on this project and what positive qualities you would bring to our team.
Curriculum vitae.
List of 3 references (Please do not have letters sent at this time. Letter writers will be contacted directly by the PI)
Please e-mail your application to Dr. Echeverri: echev020@umn.edu
Contact inhibition of locomotion is a widespread phenomenon in migrating cells. However, cells often migrate collectively as a sheet, raising the question of how contact inhibition is overcome in these scenarios. A new paper in Development addresses this problem by studying the signals that regulate collective migration in Xenopus leading edge mesendoderm (LEM) cells. We caught up with the paper’s two authors Martina Nagel and Rudolf Winklbauer, Professor in the Department of Cell and Systems Biology at the University of Toronto, to find out more about the story.
Rudolf and Martina
Rudolf, can you give us your scientific biography and the questions your lab is trying to answer?
RW I did my PhD work at the MPI for Virus Research in Tübingen, now MPI for Developmental Biology, in the lab of Peter Hausen. I was the first graduate student in his lab who used Xenopus to study embryonic development, and I remained with the frog embryo ever since. As a postdoc in Ray Keller’s lab at Berkeley, I settled on my present research agenda: cell migration in the gastrula. Back in Germany, I led a small research group in Hausen’s Department at the MPI in Tübingen. From 1993 to 1999, I held an Assistant Professor-like position at the University of Cologne. It was non-tenure-track, and to keep me off the street afterwards, Peter Hausen offered me a Senior Researcher position at his Department. Since 2001, I am at home at the Cell and Systems Biology Department at the University of Toronto.
As it turned out, almost every gastrulation movement in the frog embryo can be considered a cell migration process. The basic question is: how do cells migrate across and between other cells all the while they adhere firmly to each other to keep the embryo together? Naturally, with this question in mind, one becomes also interested in problems of cell adhesion and tissue mechanics.
Martina, PubMed tells me you first published with Rudolf in 1991 – how did you initially come to work together, and what questions have driven your research since then?
I was Rudi’s first graduate student, and my task was to find out how prospective head mesoderm cells find their target region as they migrate through the frog gastrula. Subsequently I studied also other aspects of mesoderm cell motility, but this primordial question is still my main interest.
In that 1991 paper, you described how LEM aggregates formed oriented lamellipodia during their migration. What initially drew you to this phenomenon?
MN That lamellipodia all pointed in the direction of overall mesoderm movement had been observed in the intact embryo. It had been the first indication that the cells could somehow sense the direction of their target at the animal pole of the embryo. We reproduced this orientation process in vitro to study its mechanism.
RW Tina politely disregarded my enthusiastic attempts to supervise my first graduate student and, against my nicely laid-out plans, performed an elegant experiment which demonstrated that mesoderm cells are guided by cues in the extracellular matrix of the ectoderm.
MN A major step in our understanding of the cell guidance mechanism was the identification of PDGF-A signalling as an essential factor in the process. We did this in collaboration with Karen Symes, Boston University. Our subsequent attempts to understand how PDGF-A actually functioned as a guidance cue led us to the current work.
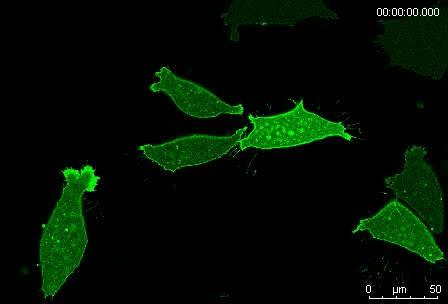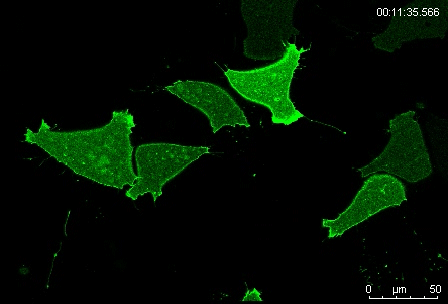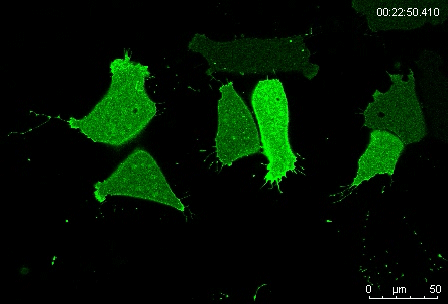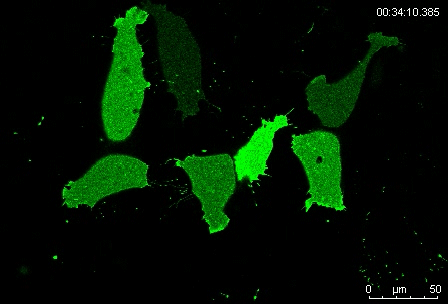
Protrusion formation and behavior of single mesoderm cells on fibronectin. Movie 1 from the paper
Can you give us the key results of the paper in a paragraph?
MN Outside of the embryo, mesoderm cells show contact inhibition of migration: if the lamellipodium of a cell contacts another mesoderm cell, it collapses, and movement ceases. On the endogenous substratum, however, PDGF-A bound to the extracellular matrix suppresses contact inhibition, which allows cells to form long-lasting lamellipodia. In a way that we do not yet understand, this suppression of lamellipodia collapse is spatially biased, leading to the preferential survival of protrusions that point into the predetermined direction of migration. This potentially generates the uniform orientation of lamellipodia that was the starting point of our investigation.
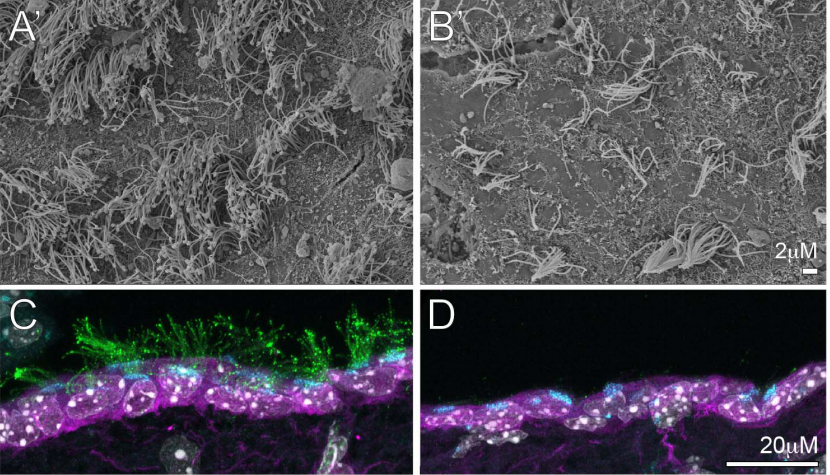
SEM images of the substratum-facing surface of the LEM, from Fig. 8 in the paper
So you have pathway involving a series of inhibitory interactions linking Pak1 to ephrin: do you have any idea of the molecular basis of any of these interactions?
MN Ideas, yes; but no data yet. We find it an attractive hypothesis that direct interaction between factors enables or prevents signalling between cells, depending on the size of these factors. For example, Eph receptor/ephrin signalling requires close cell-cell contact, but patches of syndecan 4 in the cell membrane would locally preclude such close contact due to the size of the syndecan molecules. Another possibility is the exchange of cytoplasmic signals between factors, for example between the PDGF receptor and integrin. But so many putative downstream signalling pathways exist for all the factors involved…
When doing the research, did you have any particular result or eureka moment that has stuck with you?
RW Many eureka moments; too many!
MN Ever so often, we thought we finally understood how the whole thing worked, until the next experiment gave us exactly the opposite result from what we had predicted.
RW Back to the drawing board. But the nice thing was: after a while…
MN … a long while…
RW … after a while, this pattern changed, we more and more often predicted correctly what would happen, and gradually most of our observations found their place in the puzzle.
A mesoderm aggregate on a fibronectin-PDGF substratum. Movie 9 in the paper
And what about the flipside: any moments of frustration or despair?
MN As just mentioned: whenever a really cool idea went to dust after an experiment.
RW Cell and developmental biologists have done an enormous job over the last decades, and so many facts and concepts are known now that are relevant to our work. This makes adding new results to the existing body of knowledge so complicated. Where we had to align two little known interacting factors for a paper thirty years ago, we now have to juggle a handful of them. And they all are known to do multiple things and to interact with multiple other factors.
And where will this work take you?
MN We hope that we will better understand how migration is guided by the substratum in systems where cells move as parts of a large, tightly coherent cell mass.
RW We also fancy that we might identify a core pathway regulating contact inhibition of migration. By determining the position in the pathway of other factors that we and others have found to be involved in the process. We wonder whether such a pathway exists, or whether lamellipodia collapse is easy to get and regulated in different ways in different contexts.
When you think about it more generally, cells being able to freely crawl around in the organism, to exchange places and to re-group is an essential part of what makes an animal an animal at the cell level. Green plants don’t do it, red or brown algae won’t, and so on. Fungi seem to try. And after all: it is animals who found out all this stuff about cell migration that we were just talking about.
Finally, let’s move outside the lab – what do you like to do in your spare time in Toronto?
RW Watching how our little backyard garden turns slowly into a tiny patch of forest in the middle of the city, where the squirrels put on a show, the sparrows ask for bread crumbs, and…
MN Half the year it is winter in Toronto!
RW Well, say a third of the year. And there are always books.
Iain Martyn & Tatiane Kanno share their experiences of the discovery of the human organizer
“It’s alive!” Iain’s first impressions
“Hybrid human chicken embryos: HALF HUMAN – HALF CHICKEN abomination created in US lab” was my favourite headline reporting on our work1. While the headline and accompanying article managed to miss the science completely, the author may have been surprised to know how close he or she came to capturing mood of that first, Frankenstein-like moment of discovery of the “abomination”.
For starters, it really was a dark and stormy night. The lab, high on the seventh floor of a sheer, grey, impenetrable tower, was deserted and silent. Only the intermittent odd hum and hiss of an incubator or the sound of rain and wind lashing on the windows broke the stillness and betrayed the presence of something living and growing in its confines. Far below, next to the seething, storm-overloaded river, a hunched figure made its way hurriedly across a narrow bridge towards the tower.
That would of course be me, making the dash from my apartment to the lab foolishly without a rain-jacket and trying not to get soaked. Almost exactly 24 hours previously I had grafted human embryonic stem cells into a developing chicken embryo, a long-shot search for the never-before-seen human organizer, and now it was time to check the result. What exactly was I going to see? What would it look like? A monster? Some sort of bird-human chimera? Despite the gothic atmosphere I thought to myself it was more likely that all I was going to see was a mess of dead or dying cells. This was after all my first attempt and I was a novice at chick embryology. The only reason I was here at this midnight hour in the first place was because I had run late the night before, taking over six hours to set up what any half-competent chick embryologist could do in two. Still, as I made my way into the darkened lab, took the grafts from the incubator, and loaded them onto the microscope, I could not help the apprehension rise within me.
In the first dish the cells were indeed dead or dying, torn apart by a clumsy error made during the grafting. In the second dish the graft was alive, but relatively unchanged from last night, undisturbed by the developing host chick and not disturbing it in turn. In the third dish…well three really is a lucky number: in the third dish was a little “abomination”. There, besides the normally developing host chick, the fluorescently tagged human cells had grown, expanded their area, and fused with the host tissue. More dramatically, they had also coalesced and grown into a long thin rod-like structure, emanating from the center of the graft and pointing like a dismembered finger towards the host. This is the point where lightning should have struck, thunder should have boomed, and I should have stood up and shouted “it’s alive!”, but I was more concerned with gathering evidence and recording what I saw. In fact, I think I only released the breath I’d been holding when I was sure I had taken two good pictures with the microscope’s camera and saw that they were each safely stored on the computer.
Good thing that I did as well, for none of the remaining grafts showed anything so remotely as dramatic. And when I returned to the successful graft the following morning to see if it had grown into anything even more remarkable I found only dead or dying cells. Those pictures and the memory of the previous night were all that remained, and if it were not for them, and not for that one successful graft, I might have given up and gone back to my co-PIs Ericand Alito report that it was a total failure. As it were, I became convinced that if it happened once it would happen again. The way forward to fully studying and proving the existence of the human organizer was still long and difficult, and it required teaming up with a bona fide chick embryologist, but after that night I was sure we could get there.
Iain and Tati conducting graft experiments
“…but what is it?” The striking moment for Tati
My story in the Brivanlou lab begins before I join the team as a postdoc, not so very long ago. At that time, I was a PhD student visiting the lab to learn and perform some experiments with embryonic stem cells. When I finished my internship, I returned to Brazil to defend my thesis. Few months went by and there I was, coming back to New York.
It was my first day back as an official lab member when I first came across this project. I remember being in the conference room, feeling that mixture of excitement and anxiety for starting a new chapter in my career when I heard “Hey, welcome back! Can I show you something cool?” That was the moment when I was introduced to lucky embryo number 3. As an embryology enthusiast, I got thrilled with those pictures! Some ideas had already started to pop up in my mind. We teamed up to optimize the chick experiments and that was just the beginning of our long journey in search for the human organizer.
The first set of grafting took longer than I expected: even being very familiar with chick embryo manipulation, it was my first time trying to generate a chimera. It was late, I was exhausted and hungry crossing the narrow bridge back home, but I was also feeling an excitement and eagerness for the daybreak to see the results. As it turned out, our first grafted embryos looked more like a Picasso painting. I still think MoMa museum would love to exhibit our nightmarish sci-fi art. But a tweak here and there and we managed to keep the embryos alive and looking more… normal-ish! After that, it was a marathon. Besides the long hours in lab grafting, swayed by Brazilian forró songs and replenishing ATP with Iain’s hidden snacks, we also had to go through endless washing steps for in situ and long confocal imaging sessions.
And then, finally there it was! In the elongated structure emanating from the human cells we found expression of SOX2!! How awesome that could be?! To me, that was the mind-blowing moment, but of course I still had to hold my horses and wait for the in situ results of SOX3 probe to confirm our findings. SOX2 and SOX 3 were ectopically induced in chick cells that surrounded the human cells!! We had generated our very first chick-human chimera, our “Chuman”! Our results bring valuable insights into early human development.
This work was one of those “high risk, high reward” kinds of project. It could lead to an amazing discovery or could give us nothing. Gladly, with a wonderful teamwork, we got the reward!
Welcome to our monthly trawl for developmental biology (and other related) preprints!
This month we found a tranche of preprints getting deep into the mechanics of fly development, a clutch on organoids (retinal, cerebral, cortical!), an investigation into the role of gender in scientific collaboration, and a veritable zoo in our evo-devo section – from ladybirds to placozoans via pufferfish, hydra and choanoflagellates.
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Let us know if we missed anything, and use these links to get to the section you want:
Amot regulates neuronal dendritic tree through Yap1
Katarzyna O. Rojek, Joanna Krzemien, Hubert Dolezyczek, Pawel M. Boguszewski, Leszek Kaczmarek, Witold Konopka, Marcin Rylski, Jacek Jaworski, Lars Holmgren, Tomasz J. Proszynski
Morphogen-Lineage Selector Interactions During Surface Epithelial Commitment
Sandra P Melo, Jillian M Pattison, Samantha N Piekos, Jessica L Torkelson, Elizaveta Bashkirova, Maxwell R Mumbach, Charlotte Rajasingh, Hanson Hui Zhen, Lingjie Li, Eric Liaw, Daniel Alber, Adam J Rubin, Gautam Shankar, Howard Y Chang, Paul A Khavari, Anthony E Oro
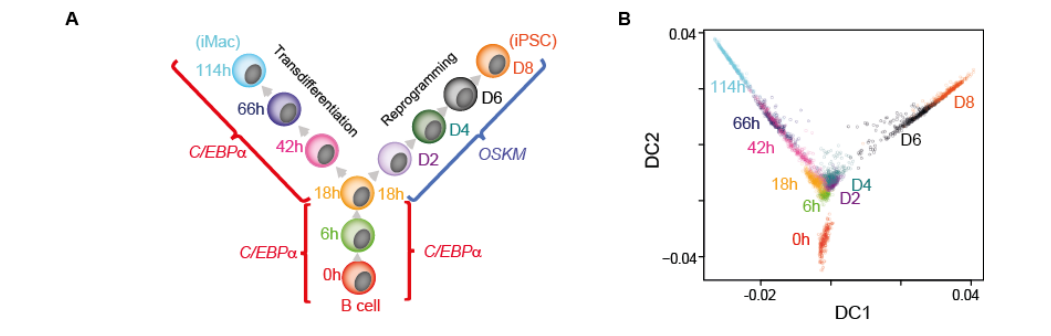
Francesconi, et al.’s transdifferentiation/reprogramming schema
Active fluctuations modulate gene expression in mouse oocytes
Maria Almonacid, Stephany El-Hayek, Alice Othmani, Isabelle Queguiner, Fanny Coulpier, Sophie Lemoine, Leïla Bastianelli, Christophe Klein, Tristan Piolot, Philippe Mailly, Raphaël Voituriez, Auguste Genovesio, Marie-Hélène Verlhac
Genome-scale oscillations in DNA methylation during exit from pluripotency
Steffen Rulands, Heather J Lee, Stephen J Clark, Christof Angermueller, Sebastien A Smallwood, Felix Krueger, Hisham Mohammed, Wendy Dean, Jennifer Nichols, Peter Rugg-Gunn, Gavin Kelsey, Oliver Stegle, Benjamin D Simons, Wolf Reik
How ladybirds get their spots, from Gautier, et al.’s preprint
The genomic basis of colour pattern polymorphism in the harlequin ladybird
Mathieu Gautier, Junichi Yamaguchi, Julien Foucaud, Anne Loiseau, Aurelien Ausset, Benoit Facon, Bernhard Gschloessl, Jacques Lagnel, Etienne Loire, Hugues Parrinello, Danny Severac, Celine Lopez-Roques, Cecile Donnadieu, Maxime Manno, Helene Berges, Karim Gharbi, Lori Lawson-Handley, Lian-Sheng Zang, Heiko Vogel, Arnaud Estoup, Benjamin Prud’homme
Transcriptomic atlas of mushroom development highlights an independent origin of complex multicellularity
Krisztina Krizsan, Eva Almasi, Zsolt Merenyi, Neha Sahu, Mate Viragh, Tamas Koszo, Stephen Mondo, Brigitta Kiss, Balazs Balint, Ursula Kues, Kerrie Barry, Judit Cseklye, Botond Hegedus, Bernard Henrissat, Jenifer Johnson, Anna Lipzen, Robin A. Ohm, Istvan Nagy, Jasmyn Pangilinan, Juying Yan, Yi Xiong, Igor V. Grigoriev, David S. Hibbett, Laszlo G. Nagy
Systematic Characterization of RhoGEF/RhoGAP Regulatory Proteins Reveals Organization Principles of Rho GTPase Signaling
Paul Markus Mueller, Juliane Rademacher, Richard D Bagshaw, Keziban Merve Alp, Girolamo Giudice, Loise E Heinrich, Carolin Barth, Rebecca L Eccles, Marta Sanchez-Castro, Lennart Brandenburg, Geraldine Mbamalu, Monika Tucholska, Lisa Spatt, Celina Wortmann, Maciej T Czajkowski, Robert William Welke, Sunqu Zhang, Vivian Nguyen, Trendelina Rrustemi, Philipp Trnka, Kiara Freitag, Brett Larsen, Oliver Popp, Philipp Mertins, Chris Bakal, Anne-Claude Gingras, Olivier Pertz, Frederick P Roth, Karen Colwill, Tony Pawson, Evangelia Petsalaki, Oliver Rocks
Kilohertz frame-rate two-photon tomography
Abbas Kazemipour, Ondrej Novak, Daniel Flickinger, Jonathan S Marvin, Jonathan King, Philip Borden, Shaul Druckmann, Karel Svoboda, Loren L Looger, Kaspar Podgorski
Precise tuning of gene expression output levels in mammalian cells
Yale S. Michaels, Mike B Barnkob, Hector Barbosa, Toni A Baeumler, Mary K Thompson, Violaine Andre, Huw Colin-York, Marco Fritzsche, Uzi Gileadi, Hilary M Sheppard, David JHF Knapp, Thomas A Milne, Vincenzo Cerundolo, Tudor A Fulga
Reproducible big data science: A case study in continuous FAIRness
Ravi K Madduri, Kyle Chard, Mike D’Arcy, Segun C Jung, Alexis Rodriguez, Dinanath Sulakhe, Eric W Deutsch, Cory Funk, Ben Heavner, Matthew Richards, Paul Shannon, Gustavo Glusman, Nathan Price, Carl Kesselman, Ian Foster
Applications are invited for a four-year funded PhD studentship to conduct research in developmental neurobiology under the supervision of Dr Alexander Fletcher and Prof. Marysia Placzek at the University of Sheffield.
This project is on the development of the hypothalamus, a brain structure with very similar anatomy across vertebrate species. We know that the hypothalamus is very important for mediating physiological homeostasis, yet its development remains poorly understood. This project will address this through a combination of gain-and loss-of function studies in vivo and ex vivo 3D culture with computational modelling.
This project is a great opportunity for a student interested in developmental neurobiology, who is keen to tackle new techniques and work in a truly interdisciplinary environment, acquiring programming and modelling skills and gaining expertise in tissue culture and imaging.
A four-year fully-funded EPSRC studentship is available to home students (British or EU nationality based in the UK) starting in October 2018. Please get in touch with Alexander Fletcher (a.g.fletcher@sheffield.ac.uk) for more details.
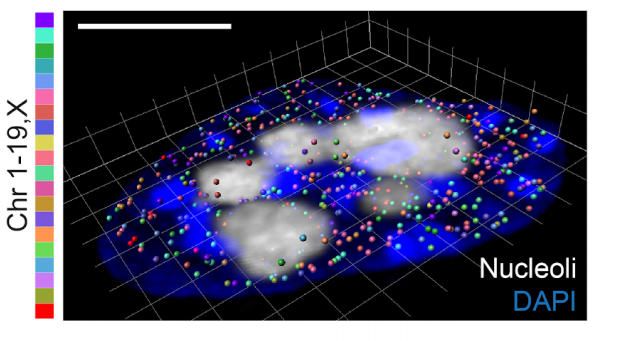
We are at the National Institute of Child Health and Human Development (NICHD) at NIH. Our lab is interested in understanding cell lineage differentiation, gene regulation and how non-coding DNA elements and the 3D architecture of chromosomes contribute to these processes during early mouse development.
Fully-funded postdoc positions for multiple years including health benefits
Opportunity to start your own research program or lead ongoing projects
The NIH research community is unparalleled in its size, diversity and resources
Possibility of living in a diverse, liberal and vibrant city: Washington DC
Or living in a calm residential area with great schools and good affordable housing , Bethesda and Rockville
Who you are:
You share our enthusiasm for epigenetics, gene regulation, nuclear organization and mouse development.
You have PhD-experience in one or more of the following: mouse development, mouse genetics, epigenetics, massively-parallel sequencing techniques or computational biology.
Apply: Send the following to gsrunit@gmail.com:
2 paragraph cover letter explaining your scientific trajectory and why you would like to join us.
CV and email contacts for 3 references.
The NIH is dedicated to building a diverse community in its training and employment programs.
An engineer position in cell biology and biomaterials is available starting from September 2018 in the group of André Le Bivic, at the Institute of Biology of Development of Marseille (IBDM), France. The project aims at understanding the impact of extracellular matrix stiffness on the development of human colon organoids, and in particular on the crosstalk between the matrix and the polarity complexes. The initial contract will be made for 12 months, with possible extension to up to 2 years.
We are seeking for a highly motivated candidate who will use a wide range of methods to support this interdisciplinary research project on the impact of extracellular matrix stiffness on epithelial morphogenesis. The main tasks of the engineer will be the preparation of organoid cultures in 3D from human colon samples and the development / optimization of different types of biocompatible matrices (collagen, BME, PEG or PAA etc.). Specifically, the job requires expert knowledge in the synthesis of biocompatible polymers. Some knowledge in cell biology (cell culture, transfection and immunostaining) and imaging (confocal microscopy, image analysis) would be a plus. The working language in the laboratory is English. Candidates are expected to be able to communicate in English.
A cover letter, a CV and the names of two referents must be sent to the following addresses: Elsa Bazellières: elsa.bazellieres@univ-amu.fr and André Le Bivic: andre.le-bivic@univ-amu.fr
The story behind melanocyte BACE2, posted by Yan Zhang and Richard White. You can read our recently published full article at Developmental Cell using this link.
Our story began six years ago when my mentor, Dr. Richard White, opened the zebrafish facility and showed me those swimming creatures. He pointed to one type with pigmented stripes and told me those are wild-type fish named AB. He caught one fish with a net and that fish very quickly jumped out of the net and escaped to a water reservoir, before I could have a closer look at her. I did not know that after that day, I would officially join a fish lab and have days and nights to observe those free swimming, free jumping animals.
That escaped fish has a stunning array of pigment patterns which is composed of three types of pigment cells: black melanophores, yellow or orange xanthophores and silvery iridophores (Parichy, 2003). While the tank next to it is filled with a transparent version, casper, where the black melanophores and the silvery iridophores are absent (White et al., 2008), some of the other fish have fuzzy pigmentation with a black tumor on the back. Those are melanomas due to uncontrolled growth of melanophores (Patton et al., 2005). I was fascinated by how diverse a pigment pattern can look like and why animals evolve them.
Animals do this for a reason. Dolphins and marlin have a darker upper surface and a white lower belly. They countershade themselves so that when seen from the top, the dark dorsal matches with deep water darkness and when seen from the bottom, the light colored belly mixes into a sunlit water above. This is one example of camouflage in front of predators. Female guppies prefer male guppies with more orange coloration, possibly because males fed on high-carotenoid diet could better reject interspecific allografts of scales and resist parasite infection, suggesting they have better immune function (Houde, 1997) – a nice example of how guppies use pigment color as a honest signal for health during sexual selection.
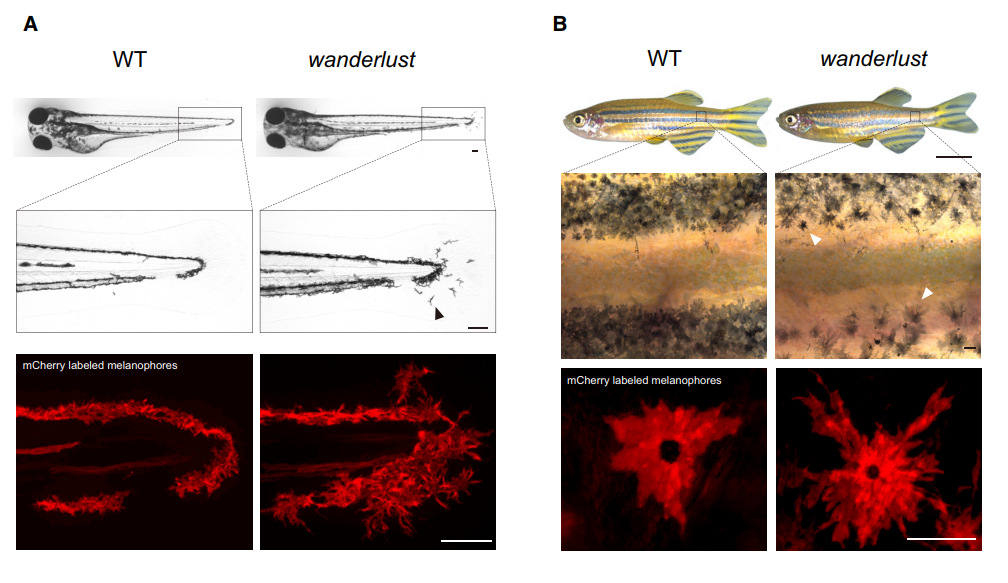
Our interests into Bace2 started when Dr. Richard White found out this gene is highly enriched in human melanoma, suggestive of oncogenic effects. We thought knocking it out would make melanomas less aggressive. But when we got zebrafish Bace2 mutants from the Sanger Center, the melanophores looked a lot “more” aggressive, an initially counterintuitive observation. It turned out this pigment mutant has super elongated melanophore cell projections, a structure named the dendrite. In mammals, melanocytes use these long dendrites to transfer melanin to neighboring keratinocytes, a process involved in tanning response to protect keratinocytes from UV-induced damage (Yamaguchi et al., 2007). Unlike mammals, zebrafish melanophores do not transfer melanin across cells, but instead traffic melanin intracellularly to modulate fish appearance (Logan et al., 2006). When melanin is aggregated around the nucleus, the fish looks lighter, and this typically occurs when fish are raised in a daylight environment. When the fish is raised in dark, melanin tends to be dispersed so that melanin covers more area and fish can match their dark-looking color with the environment. Even though zebrafish no longer use dendrites as a channel for melanin delivery, their melanophores can still be very dendritic especially when young melanophores are still actively developing. They likely use these dendrites for patterning and other ways of communicating with their neighbors. Mature melanophores lose those dendrites for unknown reasons, but not for the bace2 mutant, where their melanophores keep dendricity from embryos to adults (Figure 1).
(A) Brightfield imaging shows that the bace2-/- melanophores are hyperdendritic compared with WT fish (arrowhead) in the tail fin at 72 hpf. Labeling of the melanophore cell membranes (bottom panel) using the Tg(tyrp1b: membrane-mCherry) line demonstrates that this is due to a change in cell morphology rather than a redistribution of melanin.Figure 1. The zebrafish bace2-/- mutant has hyperdendritic melanophores. (B) These hyperdendritic melanophores are maintained into adulthood, and yield irregular stripe boundaries (arrowhead). Scale bars: (A, B middle and bottom) 100 μm; (B top) 0.5 cm.
We were intrigued by this out of control problem and sought to study why. We showed that Bace2 works during melanophore maturation, a time frame when melanophores turn on pigment genes and gain melanin. The question was how does it work? Bace2 is a cell-intrinsic sheddase which modulated cell morphology inside the melanophore lineage. We further sought to find out which protein is cleaved by Bace2 to exhibit all those phenotypes. We had no luck in the beginning. Pmel and Gpnmb are the two initial guesses as both of them are involved in melanin production and PMEL can be cut by BACE2 in mice (Rochin et al., 2013; Shimshek et al., 2016). But we soon found out those two substrates could not explain the melanophore dendricity. The research was stuck for a while. When I was even trying to grab everything I could find in our chemical room and threw them into fish to have a try, Dr. White came to me and said, why don’t we try an unbiased chemical screen?
The breakthrough came with a change in methodology. The chemical screen gave us an unexpected but fruitful hit. We found four chemicals able to convert bace2 mutant melanophores into normal looking ones, all of which converge on the same pathway-one that contains insulin, PI3K and mTOR. We realized this is something never studied before, a new PI3K/mTOR regulator that has a melanophore-specific consequence. All of the pieces came together. It turned out Bace2 itself cleaves the insulin receptor and this cleavage modulates how many functional insulin receptors are left on the cell membrane. In the mutant fish, Bace2 no longer cleaves it and melanophores have hyperactivated insulin/PI3K/mTOR which drives this uncontrolled dendricity. The driving force came from long distance, as we found that a brain-derived insulin peptide (insb) is the stimulating ligand that feeds into insulin receptor in this context.
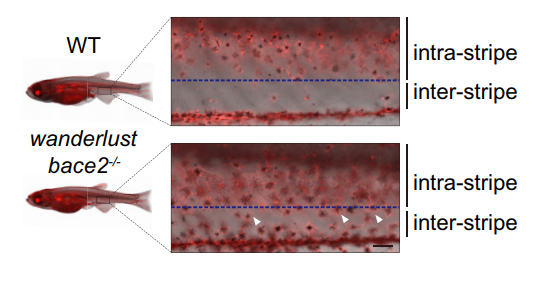
There are other consequence associated with uncontrolled dendricity: those bace2 mutant melanophores are actively differentiating, actively dividing and wandering around to ectopic locations (Figure 2). We decided to name this bace2 mutant wanderlust as those melanophores like to explore the world, travel to new sites and are free of constraints.
Figure 2. bace2-/- mutants develop melanophores outside of the stripe during metamorphosis at 24 days post fertilization (dpf) (arrowhead). Scale bars: 100 μm.
One thing that emerged from this research is the power of unbiased approaches to a problem. We were stuck for a while, but the screen turned out to be the most efficient and rapid way to get to the answer. It’s one of the greatest things about the zebrafish, and has allowed us to connect things – insulin and melanophores – that would have been hard to guess otherwise.
 (4 votes)
(4 votes) (No Ratings Yet)
(No Ratings Yet)