The team of B. Chazaud at Institut NeuroMyoGene at the Université Claude Bernard Lyon 1, CNRS, INSERM, at Lyon, is seeking to fill 2 postdoctoral research scientist positions in the area of skeletal muscle regeneration and homeostasis. One of the main goals of the Chazaud laboratory research programs is to understand the role of the close environment on muscle stem cell fate in normal regenerating skeletal muscle as well as during muscle diseases.
Requirements
PhD (or near PhD completion) in biological or biomedical sciences or post-doc scientists with no more than 2/3 years post-doc experience.
Scientific publications in peer-reviewed journals and presentations at scientific conferences. Published evidence of aptitude for high quality research is essential.
A strong background in the following fields:
Profile 1 – exploration of the role of extracellular matrix in skeletal muscle biology (PI: Dr. B Chazaud). The candidate must have an experience working in the biology and biochemistry of extracellular matrix. A background in skeletal muscle biology is a plus, but not mandatory.
Profile 2 – exploration of the role of myofiber contraction on skeletal muscle environment (PI: Dr. R Mounier). The candidate must have an experience working in the biology and physiology of skeletal muscle, with a reference to in vitro/ex vivo muscle cell contraction.
Terms and conditions
The post-doc researchers will conduct and nurture the research programs in close connection with the PI, until the submission of high-quality articles. Positions are open from the beginning of 2018. Salaries are available for 3 years, renewed every year according to progress, however the post-doc researchers are expected to be competitive and to be able to obtain their own independent financing.
Environment
B Chazaud team is part of Institut NeuroMyoGene, a new Institute dedicated to basic and translational research in the neuromuscular field that hosts 14 research teams encompassing 200 people and that is connected to numerous facilities (molecular biology, imaging, cell biology, physiology), guaranteeing a high quality and dynamic scientific environment.
As the second biggest town in France, Lyon hosts 60 laboratories (4500 people) in the area of Life and Health Sciences. Lyon is ideally located between the Alps and the Mediterranean Sea. Lyon is renowned for its quality of life including outdoor leisure closely the big city, and of course, its gastronomy and wines.
How to apply?
Please send to the PI: 1) a cover letter highlighting research interests, goals and previous scientific contributions; 2) a CV listing education, publications, meeting presentations and any other skills of interest; 3) at least 2 reference contacts.
The Company of Biologists (biologists.com) is launching a new preprint highlighting service for the biological community. We are looking for the right person to help us build and evolve this new service.
Joining an experienced and successful publishing team, this is an exciting opportunity for an enthusiastic and motivated team builder to take a step into publishing. Offered initially as a three-year appointment, we expect the role to evolve in new and interesting ways.
Applicants will have relevant research experience, ideally a PhD in a field relevant to one or more of the Company’s journals. They should have a good understanding of the needs of scientists and the growing impact of preprints as well as a demonstrated interest in science communication and experience with social media.
Core responsibilities include:
• building and maintaining a team of community contributors
• developing a good content strategy around preprint selection and commenting
• smooth running of the preprint highlighting service
• growing an active social media presence
• identifying opportunities that allow us to evolve this community service
Essential requirements for the job are a broad understanding of science and the scientific community, excellent communication and team-building skills, plus confidence in networking and managing relationships. The successful candidate will have a diplomatic style, enthusiasm, judgement and integrity. This position has an attractive salary and benefits and represents a unique career opportunity within a highly successful not-for-profit publisher. The role is based in our attractive modern offices on the outskirts of Cambridge, UK.
The Company of Biologists (biologists.com) exists to support biologists and inspire advances in biology. At the heart of what we do are our five specialist journals – Development, Journal of Cell Science, Journal of Experimental Biology, Disease Models & Mechanisms and Biology Open – two of them fully open access. All are edited by expert researchers in the field, and all articles are subjected to rigorous peer review. We take great pride in the experience of our editorial team and the quality of the work we publish. We believe that the profits from publishing the hard work of biologists should support scientific discovery and help develop future scientists. Our grants help support societies, meetings and individuals. Our workshops and meetings give the opportunity to network and collaborate.
To apply, please send your CV by email to recruitment@biologists.com along with a covering letter that states your current salary, summarises your relevant experience and explains why you are enthusiastic about this opportunity. You must be able to demonstrate your entitlement to work in the UK.
Applications should be made as soon as possible and by 8th January 2018. Late applications may be considered.
Following a generous donation, the BSDB has instituted the Dennis Summerbell Lecture, to be delivered at its annual Autumn Meeting by a junior researcher at either PhD or Post-doctoral level. The 2017 lecture awardee was Helen Weavers (School of Biochemistry, Faculty of Biomedical Sciences, University of Bristol) was with her submitted abstract “Understanding the inflammatory response to tissue damage in Drosophila: a complex interplay of pro-inflammatory attractant signals, developmental priming and tissue cyto-protection”. Her award lecture was presented at the Autumn Meeting 2017, jointly organised by the BSDB together with the Swedish, Finish, Norwegian and Danish Societies of Developmental Biology, 25-27 October 2017 in Stockholm.
Helen’s work so far
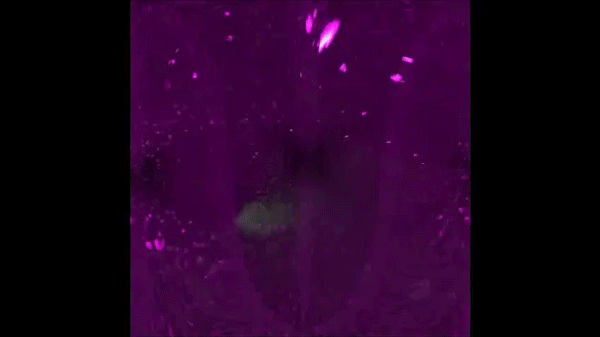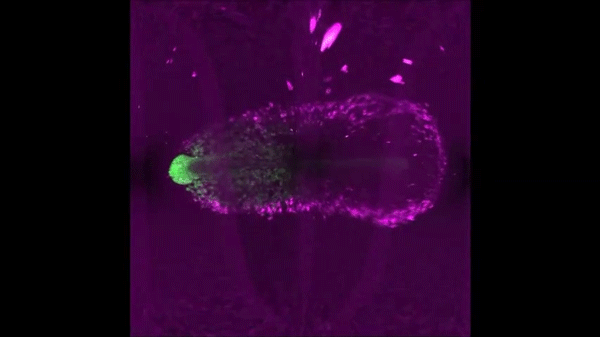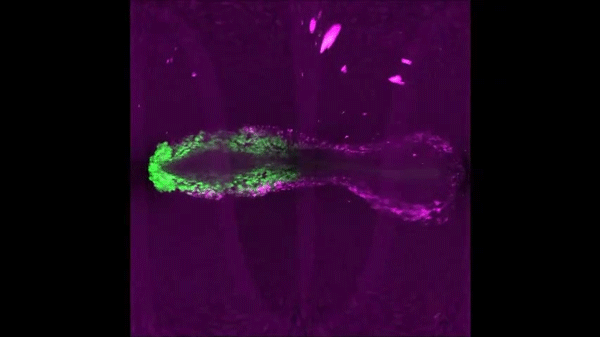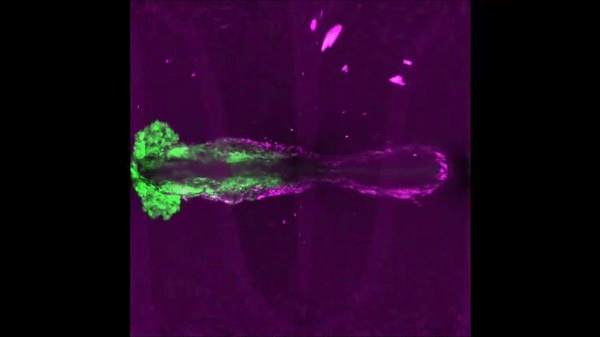
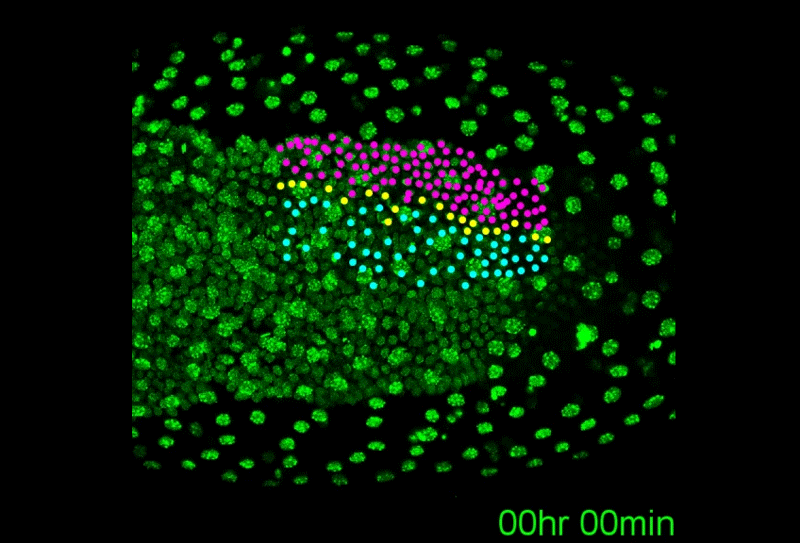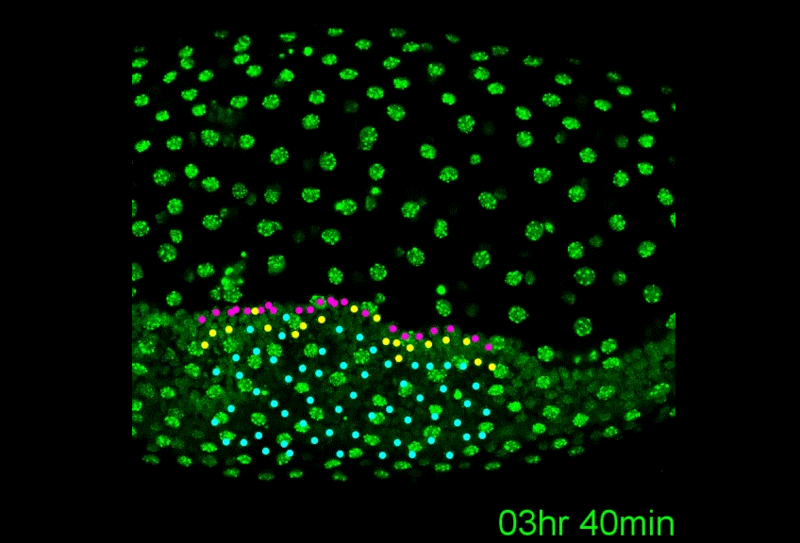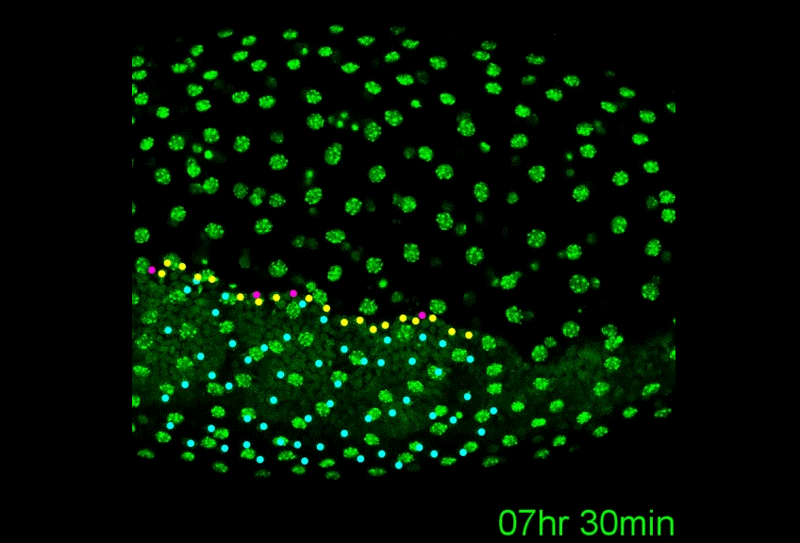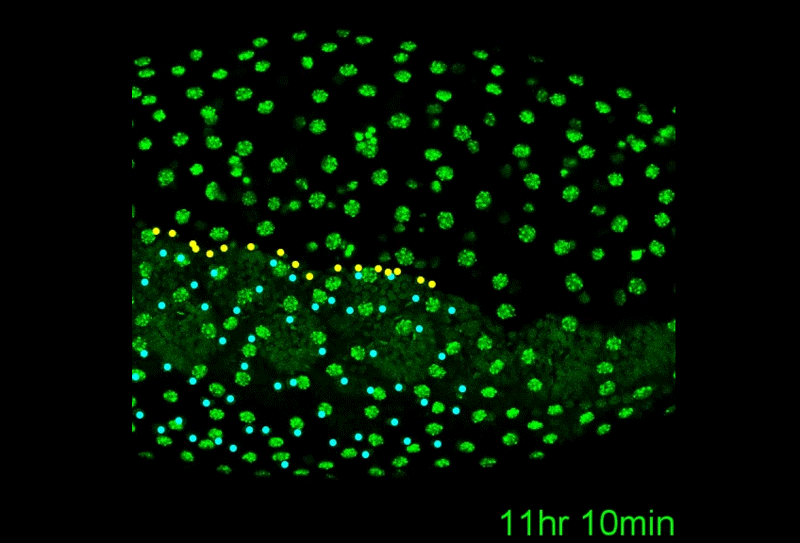
After completing her PhD studies investigating Drosophila nephrogenesis in Helen Skaer’s lab in Cambridge, Helen moved to Bristol in 2013 to take up a 5 year, MRC-funded post-doc position between Paul Martin’s and Will Wood’s labs. Her first publication from this work (Weavers et al., 2016, Cell 165, 1658ff.), showed that Drosophila macrophages (haemocytes), must first be “primed” by engulfing at least one dead cell, before they are responsive to wound attractants. These findings are important because the majority of human pathologies are a consequence of too little or too much inflammation. What really excited the judges of the Denis Summberbell Lecture award was the work which had led to her most recent paper entitled “Systems Analysis of the Dynamic Inflammatory Response to Tissue Damage Reveals Spatiotemporal Properties of the Wound Attractant Gradient” (Weavers et al., 2016, Curr Biol 26, 1974ff.). This was a true multidisciplinary study, using a combined approach of mathematics and biology to analyse macrophage behaviours in response to tissue damage. Although the identity of the wound attractant signal/s are still not clear, this study was able to determine several of the characteristics of the attractant(s). Building on this strong platform of work, Helen is currently developing her own research towards understanding tissue protection/resilience in Drosophila and man, and this was an exciting novel element of her award lecture. In her talk, she described in a stunningly visual and understandable way how successful tissue repair relies not only on the host’s ability to mount an effective inflammatory response, but also on its ability to limit it. Her talk was a fabulous highlight and a shining example of high quality research by members of the BSDB.
Lecture abstract:
Understanding the inflammatory response to tissue damage in Drosophila: a complex interplay of pro-inflammatory attractant signals, developmental priming and tissue cyto-protection
Helen Weavers, Bristol, UK
An effective inflammatory response is pivotal to fight infection, clear debris and orchestrate the repair of injured tissues; however, inflammation must be tightly regulated since many human disease pathologies are a consequence of inflammation gone awry. Using a genetically tractable Drosophila model, I use precise genetic manipulation, live imaging and computational modelling to dissect the mechanisms that activate the inflammatory response to tissue damage and those that simultaneously protect the regenerating tissue from immunopathology. Upon tissue damage, immune cells (particularly neutrophils and macrophages) are recruited into the damaged area by damage signals (danger-associated molecular patterns, DAMPs) released from the injured tissue. In collaboration with computational biologists, we employ a sophisticated Bayesian statistical approach to uncover novel details of the pro-inflammatory wound attractants, by analysing the spatio-temporal behaviour of Drosophila immune cells as they respond to wounds. We show that the wound attractant is released by wound edge cells and spreads slowly through the tissue, at rates far slower than small molecule DAMPs such as ATP and H2O2. Strikingly, we also find that immune cells must be developmentally ‘primed’ by uptake of apoptotic corpses before they can respond to these damage attractant signals. Such corpse-induced priming is an example of “innate immune memory” and may serve to amplify the inflammatory response in situations involving excessive cell death – and otherwise limit an overzealous and damaging immune response. Indeed, whilst inflammation is clearly beneficial, toxic molecules (e.g. reactive oxygen species, ROS) generated by immune cells to fight infection, can also cause significant bystander damage to host tissue and delay repair – and may underpin chronic wound-healing pathologies in the clinic. To counter this, I find that wounded Drosophila tissue employs a complex network of cyto-protective pathways that promote tissue ‘resilience’, which both protect against ROS-induced damage and stimulate damage repair. Successful tissue repair, therefore, not only relies on the host’s ability to mount an effective inflammatory response, but also its ability to finely tune it and limit associated immunopathology.</div?
Our latest monthly trawl for developmental biology (and other cool) preprints. Let us know if we missed anything.
In their end of year round up, Science magazine picked ‘Biology preprints take off’ as a runner up 2018 Breakthrough of the Year, and ran a quote from Ron Vale –
“It’s a major cultural change in communication.”
In compiling this list over 2017 (which month by month gets longer and longer), it’s been exciting to witness the buzz around preprints grow and watch this cultural change take place.
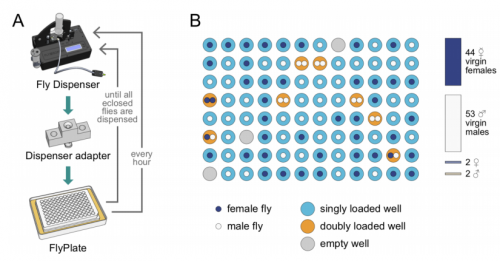
As for December, two organs seem to predominate – brains and kidneys! Also plenty of beautiful evo-devo and cell biology work, a good chunk of modelling, and the Drosophilists dream –a machine that collects your virgins for you!
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
Cardiac directed differentiation using small molecule Wnt modulation at single-cell resolution. Clayton Friedman, Quan Nguyen, Samuel Lukowski, Abbigail Helfer, Han Chiu, Holly Voges, Shengbao Suo, Jing-Dong Han, Pierre Osteil, Guangdun Peng, Naihe Jing, Greg Ballie, Anne Senabouth, Angelika Christ, Timothy Bruxner, Charles Murry, Emily Wong, Jun Ding, Yuliang Wang, James Hudson, Ziv Bar-Joseph, Patrick Tam, Joseph Powell, Nathan Palpant
Human-specific NOTCH-like genes in a region linked to neurodevelopmental disorders affect cortical neurogenesis. Ian T Fiddes, Gerrald A Lodewijk, Meghan M Mooring, Colleen M Bosworth, Adam D Ewing, Gary L Mantalas, Adam M Novak, Anouk van den Bout, Alex Bishara, Jimi L Rosenkrantz, Ryan Lorig-Roach, Andrew R Field, Maximillian Haeussler, Lotte Russo, Aparna Bhaduri, Tomasz J Nowakowski, Alex A Pollen, Max L Dougherty, Xander Nuttle, Marie-Claude Addor, Simon Zwolinski, Sol Katzman, Arnold Kreigstein, Evan E Eichler, Sofie R Salama, Frank MJ Jacobs, David Haussler
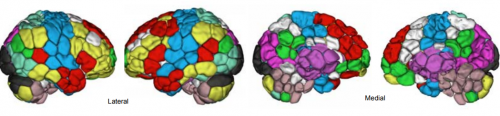
Shared and distinct transcriptomic cell types across neocortical areas. Bosiljka Tasic, Zizhen Yao, Kimberly A Smith, Lucas Graybuck, Thuc Nghi Nguyen, Darren Bertagnolli, Jeff Goldy, Emma Garren, Michael N Economo, Sarada Viswanathan, Osnat Penn, Trygve Bakken, Vilas Menon, Jeremy A Miller, Olivia Fong, Karla E Hirokawa, Kanan Lathia, Christine Rimorin, Michael Tieu, Rachael Larsen, Tamara Casper, Eliza Barkan, Matthew Kroll, Seana Parry, Nadiya V Shapovalova, Daniel Hirchstein, Julie Pendergraft, Tae Kyung Kim, Aaron Szafer, Nick Dee, Peter Groblewski, Ian Wickersham, Ali Cetin, Julie A Harris, Boaz P Levi, Susan M Sunkin, Linda Madisen, Tanya L Daigle, Loren Looger, Amy Bernard, John Phillips, Ed Lein, Michael Hawrylycz, Karel Svoboda, Allan R Jones, Christof Koch, Hongkui Zeng
A single-cell catalogue of regulatory states in the ageing Drosophila brain. Kristofer Davie, Jasper Janssens, Duygu Koldere, Uli Pech, Sara Aibar, Maxime De Waegeneer, Samira Makhzami, Valerie Christiaens, Carmen Bravo Gonzalez-Blas, Gert Hulselmans, Katina Spanier, Thomas Moerman, Bram Vanspauwen, Jeroen Lammertyn, Bernard Thienpont, Sha Liu, Patrik Verstreken, Stein Aerts
Genome Architecture Leads a Bifurcation in Cell Identity. Sijia Liu, Haiming Chen, Scott Ronquist, Laura Seaman, Nicholas Ceglia, Walter Meixner, Lindsey A. Muir, Pin-Yu Chen, Gerald Higgins, Pierre Baldi, Steve Smale, Alfred Hero, Indika Rajapakse
Distinct SoxB1 networks are required for naïve and primed pluripotency. Andrea Corsinotti, Frederick CK Wong, Tulin Tatar, Iwona Szczerbinska, Florian Halbritter, Douglas Colby, Sabine Gogolok, Raphael Pantier, Kirsten Liggat, Elham S Mirfazeli, Elisa Hall-Ponsele, Nicholas Mullin, Valerie Wilson, Ian Chambers
Whole Genomes Define Concordance of Matched Primary, Xenograft, and Organoid Models of Pancreas Cancer. Deena M.A. Gendoo, Robert E. Denroche, Amy Zhang, Nikolina Radulovich, Gun Ho Jang, Mathieu Lemire, Sandra Fischer, Dianne Chadwick, Ilinca M. Lungu, Emin Ibrahimov, Ping-Jiang Cao, Lincoln D. Stein, Julie M. Wilson, John M.S. Bartlett, Ming-Sound Tsao, Neesha Dhani, David Hedley, Steven Gallinger, Benjamin Haibe-Kains
Human iPSC-derived RPE and retinal organoids reveal impaired alternative splicing of genes involved in pre-mRNA splicing in PRPF31 autosomal dominant retinitis pigmentosa. Adriana Buskin, Lili Zhu, Valeria Chichagova, Basudha Basu, Sina Mozaffari-Jovin, David Dolan, Alastair Droop, Joseph Collin, Revital Bronstein, Sudeep Mehrotra, Michael Farkas, Gerrit Hilgen, Kathryn White, Dean Hallam, Katarzyna Bialas, Git Chung, Carla Mellough, Yuchun Ding, Natalio Krasnogor, Stefan Przyborski, Jumana Al-Aama, Sameer Alharthi, Yaobo Xu, Gabrielle Wheway, Katarzyna Szymanska, Martin McKibbin, Chris F Inglehearn, David J Elliott, Susan Lindsay, Robin R Ali, David H Steel, Lyle Armstrong, Evelyne Sernagor, Eric Pierce, Reinhard Luehrmann, Sushma-Nagaraja Grellscheid, Colin A Johnson, Majlinda Lako
HNF1A is a Novel Oncogene and Central Regulator of Pancreatic Cancer Stem Cells. Ethan Abel, Masashi Goto, Brian Magnuson, Saji Abraham, Nikita Ramanathan, Emily Hotaling, Anthony A. Alaniz, Chandan Kumar-Sinha, Michele L. Dziubinski, Sumithra Urs, Lidong Wang, Jiaqi Shi, Meghna Waghray, Mats Ljungman, Howard C Crawford, Diane M. Simeone
Evolutionary Origin of the Mammalian Hematopoietic System Found in a Colonial Chordate. Benyamin Rosental, Mark A. Kowarsky, Jun Seita, Daniel M. Corey, Katherine J. Ishizuka, Karla J. Palmeri, Shih-Yu Chen, Rahul Sinha, Jennifer Okamoto, Gary Mantalas, Lucia Manni, Tal Raveh, D. Nathaniel Clarke, Aaron M. Newman, Norma F. Neff, Garry P. Nolan, Stephen R. Quake, Irving L. Weissman, Ayelet Voskoboynik
Firefly genomes illuminate the origin and evolution of bioluminescence. Timothy R Fallon, Sarah E Lower, Ching-Ho Chang, Manabu Bessho-Uehara, Gavin J Martin, Adam J Bewick, Megan Behringer, Humberto J Debat, Isaac Wong, John C Day, Anton Suvorov, Christian J Silva, David W Hall, Robert J. Schmitz, David R Nelson, Sara Lewis, Shuji Shigenobu, Seth M Bybee, Amanda M Larracuente, Yuichi Oba, Jing-Ke Weng
Improved Aedes aegypti mosquito reference genome assembly enables biological discovery and vector control. Benjamin J Matthews, Olga Dudchenko, Sarah Kingan, Sergey Koren, Igor Antoshechkin, Jacob E Crawford, William J Glassford, Margaret Herre, Seth N Redmond, Noah H Rose, Gareth D Weedall, Yang Wu, Sanjit S Batra, Carlos A Brito-Sierra, Steven D Buckingham, Corey L Campbell, Saki Chan, Eric Cox, Benjamin R Evans, Thanyalak Fansiri, Igor Filipovic, Albin Fontaine, Andrea Gloria-Soria, Richard Hall, Vinita S Joardar, Andrew K Jones, Raissa G G Kay, Vamsi Kodali, Joyce Lee, Gareth J Lycett, Sara N Mitchell, Jill Muehling, Michael R Murphy, Arina Omer, Frederick A Partridge, Paul Peluso, Aviva Presser Aiden, Vidya Ramasamy, Gordana Rasic, Sourav Roy, Karla Saavedra-Rodriguez, Shruti Sharan, Atashi Sharma, Melissa Smith, Joe Turner, Allison M Weakley, Zhilei Zhao, Omar S Akbari, William C Black IV, Han Cao, Alistair C Darby, Catherine Hill, J. Spencer Johnston, Terence D Murphy, Alexander S Raikhel, David B Sattelle, Igor V Sharakhov, Bradley J White, Li Zhao, Erez Lieberman Aiden, Richard S Mann, Louis Lambrechts, Jeffrey R Powell, Maria V Sharakhova, Zhijian Tu, Hugh M Robertson, Carolyn S McBride, Alex R Hastie, Jonas Korlach, Daniel E Neafsey, Adam M Phillippy, Leslie B Vosshall
ZMYND10 functions in a chaperone relay during axonemal dynein assembly. Girish R Mali, Patricia Yeyati, Seiya Mizuno, Margaret A Keighren, Petra zur Lage, Amaya Garcia-Munoz, Atsuko Shimada, Hiroyuki Takeda, Frank Edlich, Satoru Takahashi, Alex von Kriegsheim, Andrew Jarman, Pleasantine Mill
Quantitative mass imaging of single molecules in solution. Gavin Young, Nikolas Hundt, Daniel Cole, Adam Fineberg, Joanna Andrecka, Andrew Tyler, Anna Olerinyova, Ayla Ansari, Erik G Marklund, Miranda P Collier, Shane A Chandler, Olga Tkachenko, Joel Allen, Max Crispin, Neil Billington, Yasuharu Takagi, James R Sellers, Cedric Eichmann, Philip Selenko, Lukas Frey, Roland Riek, Martin R Galpin, Weston B Struwe, Justin L P Benesch, Philipp Kukura
Equivalent high-resolution identification of neuronal cell types with single-nucleus and single-cell RNA-sequencing. Trygve E Bakken, Rebecca D Hodge, Jeremy M Miller, Zizhen Yao, Thuc N Nguyen, Brian Aevermann, Eliza Barkan, Darren Bertagnolli, Tamara Casper, Nick Dee, Emma Garren, Jeff Goldy, Lucas T Gray, Matthew Kroll, Roger S Lasken, Kanan Lathia, Sheana Parry, Christine Rimorin, Richard H Scheuermann, Nicholas J Schork, Soraya I Shehata, Michael Tieu, Kimberly A Smith, Hongkui Zeng, Ed S Lein, Bosiljka Tasic
Resolving the Full Spectrum of Human Genome Variation using Linked-Reads. Patrick Marks, Sarah Garcia, Alvaro Martinez Barrio, Kamila Belhocine, Jorge Bernate, Rajiv Bharadwaj, Keith Bjornson, Claudia Catalanotti, Josh Delaney, Adrian Fehr, Brendan Galvin, Haynes Heaton, Jill Herschleb, Christopher Hindson, Esty Holt, Cassandra B. Jabara, Susanna Jett, Nikka Keivanfar, Sofia Kyriazopoulou-Panagiotopoulou, Monkol Lek, Bill Lin, Adam Lowe, Shazia Mahamdallie, Shamoni Maheshwari, Tony Makarewicz, Jamie Marshall, Francesca Meschi, Chris O’keefe, Heather Ordonez, Pranav Patel, Andrew Price, Ariel Royall, Elise Ruark, Sheila Seal, Michael Schnall-Levin, Preyas Shah, Stephen Williams, Indira Wu, Andrew Wei Xu, Nazneen Rahman, Daniel MacArthur, Deanna M. Church
The Jensen group (affiliated to DanStem) is looking to recruit a highly motivated and talented postdoctoral researcher to our highly dynamic research group.
Our research/The group
We wish to understand how the intestinal epithelium forms and how adult stem cells in the forming organ are specified during development. The intestinal epithelium is associated with numerous disorders and we believe that insights into normal tissue development will allow us to harness the potential of both fetal and adult intestinal stem cell for the development of new treatment option for patients with intestinal disorders. In order to address these key questions we use a number of different techniques including in vivo fate mapping to define cellular heritage, state of the art cell culture systems to identify signaling pathways that controls directed differentiation during embryonic and fetal stages and transplantation techniques (Wong et al., 2012, Nature Cell Biology; Page et al., 2013, Cell Stem Cell; Fordham et al., 2013, Cell Stem Cell; Yui et al., 2018, Cell Stem Cell).
Project/The research project(s)
We are looking for a postdoctoral candidate with a strong cell biological and cell signaling background. The candidate will participate in an ERC funded project aimed at developing a transplantation strategy as a cure for inflammatory bowel disease. Here the candidate will use state-of-the-art genetic, cell biological and tissue-engineering strategies the candidate to identify key gene regulatory networks that control tissue maturation.
Start: Preferably May 2018 or after agreement
Duration: 3 years
Hours per week: 37
Qualifications
We expect you to be a highly motivated and highly ambitious scientist with the following qualifications:
A PhD in Life Sciences
Excellent track record with at least 1 peer reviewed first authors paper in a high-impact journal
A strong background in cellular biology is essential, and experience with transplantation technologies is an advantage
The ability to independently envision, plan and execute a research project
Excellent technical skills
Excellent English skills written and spoken
For further information regarding the position, please contact Associate Professor Kim Jensen on e-mail kim.jensen@bric.ku.dk
Place of employment
The employment is at BRIC, University of Copenhagen. BRIC is located in the Biocenter, close to the centre of Copenhagen. We offer creative and stimulating working conditions in a dynamic and international research environment. Our research facilities include modern laboratories and a number of core facilities shared between the 23 research groups at BRIC and the neighboring Finsen Laboratory. We have weekly journal clubs, data clubs, seminars with invited speakers and a young researchers club ASAP and our own PhD programme, MoMeD and our own Postdoc Career Programme. BRIC actively participates in the European alliance, EU-life consisting of 13 excellent life science research institutions http://eu-life.eu/
Salary, pension and terms of employment will be in accordance with the agreement between the Ministry of Finance and The Academics Central organization. Currently, the monthly salary starts at 33,224 DKK/ca. 4,463 Euro plus pension. Depending on qualifications, a higher salary may be negotiated.
Non-Danish and Danish applicants may be eligible for tax reductions, if they hold a PhD degree and have not lived in Denmark the last 10 years.
The position is covered by the “Memorandum on Job Structure for Academic Staff at the Universities” of June 28, 2013.
Application
Your application must be submitted electronically by clicking ‘Apply now’ below or via BRIC’s website on http://www.bric.ku.dk/jobs/. The application must include the following documents/attachments – all in PDF format:
Motivated letter of application (max. one page) detailing the basis on which the applicant scientific qualifications meet the requirements for this position.
CV incl. education, work/research experience, language skills and other skills relevant for the position.
A certified/signed copy of a) PhD certificate and b) Master of Science certificate. If the PhD is not completed, a written statement from the supervisor will do.
List of publications.
Letters of recommendation
Furthermore, the applicant should provide two letters of recommendation. To be taken into consideration, the letter must be signed by the supporting person on paper with institutional letter head and in PDF-format. The letters should be received before deadline on kim.jensen@bric.ku.dk with subject “name candidate-…-postdoc-…2018; preferably by the supporting person. Please make sure that an easy match between the recommendation letter and the individual applicant is possible.
Application deadline: 15 February 2018.
We reserve the right not to consider material received after the deadline, and not to consider applications or letters of recommendation that do not live up to the above-mentioned requirements.
The further process
Shortlist: After expiry of the application deadline, the superior with power to appoint selects a number of applicants for assessment on the advice of the Appointments Committee. All applicants are immediately notified whether their application has been passed for assessment. Applicants who were not passed for assessment should not expect further with regard to their application unless the shortlist is revised.
Assessment: The selected applications will be assessed according to the Ministry Order on the Appointment of Academic Staff at Universities 2012 and the University of Copenhagen’s guidelines 2013. The Assessment Committee makes a non-prioritized assessment of the academic qualifications and experience with respect to the above mentioned area of research, techniques, skills and other requirements listed in the advertisement.
BRIC and University of Copenhagen wish to reflect the diversity of society and welcome applications from all qualified candidates regardless of personal background.
The BSDB will soon publish its next newsletter. An important topic in that issue will be communication within our community and advocacy of Developmental Biology. See here a preview of the contribution by our communications officer Andreas Prokop describing the BSDB’s advocacy strategy.
The BSDB’s advocacy strategy
As argued in a recent PLoS Blog, there are alarming indications of communication fatigue in our community which weakens our ability to coordinate our activities and promote the importance of our science. But why do we turn off in this way at the worst possible time when conditions for fundamental research are worsening? As my colleague Sam Illingworth and I have argued in an editorial for a recent special issue about science communication, the likely reasons include (1) lack of awareness about the means and power of communication, (2) lack of incentives and external rewards for participation in science communication, and (3) lack of time: as academics we usually have more than 5 professions rolled into one, and the time demand in each of these professional spheres is steadily increasing, suffocating our productivity as scientists, let alone as communicators.
Notwithstanding, I argued in the above mentioned PLoS Blog that current circumstances cry out for communication and we MUST find feasible and effective ways to do so. As I argued, this is possible through the formation of collaborative networks of science communication. To achieve this, we need to communicate within our own communities to be able to coordinate our action. We need to make our individual contributions to science communication; if we are prepared to share the fruits of our activities, for example via The Node, this can then lead to the cumulative build-up of high quality and freely available resources and strategies. Finally, we need to make active use of and further improve existing resources and strategies; by reaching out jointly we will have a higher chance of gaining momentum and impact – all with the common goal of promoting dialogue about the science we love.
To lead the way in this direction, the BSDB has started an advocacy campaign together with The Node. The first step is simple and consists in putting together the best arguments for Developmental Biology and powerful examples illustrating these statements. The first draft of this document has been published on the BSDB site and on The Node. This resource can now be capitalised on by us all, but it also requires further community input to refine and complement the arguments – in particular also in the areas of Plant Biology and Evo-Devo which are not well represented. To catalyse this process, the editorial team of Development has complementary plans that will be announced in due term, and the BSDB has initiated a writing competition for PhD students and postdocs focussing on advocacy.
The gradually improving advocacy resource is intended to provide us with effective elevator pitches that can be used in dialogue with the public, students, other scientists, clinicians and politicians – and many of the arguments may fly well also on grant applications or in scientific publications. The overarching goal is to achieve wider recognition of fundamental Developmental Biology research as an important science branch that deserves public funding support.
But we should not stop there, and hopefully more members of our community will join in and help to develop creative science communication initiatives that carry dialogue proactively into the relevant target groups. Ideally, this is done through collaboration and long-term objective setting which has a higher chance of achieving sustainability, momentum and impact. To illustrate this point, a recent special issue on science communication describes examples of existing initiatives, explaining their origins and gradual developments. To facilitate the task, the BSDB and The Node have collaborated to put together a link collection (originally published on the BSDB site) which provides ideas, advice and resources that can be used and followed. We hope that these actions taken by the BSDB will help to raise the awareness of and participation in science communication and advocacy within our community for the benefit of all.
We are looking for an organised and enthusiastic research technician to join Professor St Johnston’s research group at the Gurdon Institute. The group works on how epithelial cells polarise using Drosophila as the model organism.
The post holder will be responsible for maintaining the fly stocks for the group and helping with research projects, so candidates should have experience of working with Drosophila. The ideal candidate will be organised and methodical with an HNC, first degree (or equivalent) in a biological science subject and should have some experience of working in a research laboratory. They will also be involved in the day-to-day running of the lab. Excellent communication skills are essential as the role involves working with all members of the group. Good IT skills are also required.
Although this is a full-time position, we welcome applications from part-time candidates and would consider a job-share, should two suitable candidates apply. Closing date 12th January 2018
Applications should be submitted through http://www.jobs.cam.ac.uk/job/15907/
We are seeking for highly motivated and competitive graduate students or postdocs to apply to the following calls:
AYUDAS PARA LA FORMACIÓN DE PROFESORADO UNIVERSITARIO (FPU) 2017
CONTRATOS JUAN DE LA CIERVA (JDC) INCORPORACION Y FORMACION 2017
Two projects are available:
– Uncovering the cellular mechanisms and the mechanics underlying the acquisition of the final shape of the embryonic Central Nervous System in Drosophila
– Exploring how different cell layers (epithelia, muscles and nerves) coordinate in a single morphogenetic process: the replacement of obsolete larval tissues to generate the adult Drosophila abdomen.
The completion of these projects will involve super-resolution imaging and cells tracking in combination with genetic studies and development of molecular markers and optogenetic tools.
Applications should include a CV and letter of motivation and must be sent by email as soon as possible:
I studied molecular mechanisms of faithful chromosome segregation in mitosis and meiosis during my PhD and wanted to continue studying chromosome segregation but from a different angle. While looking for a postdoc lab, I learned about a phenomenon called meiotic drive where the selfish genetic element preferentially segregates into the egg during female meiosis in violation of Mendel’s Law of Segregation. I became quickly fascinated by meiotic drive and decided to join Dr. Michael Lampson’s lab at the University of Pennsylvania where they study the cell biological basis of meiotic drive in mammals.
Female meiosis is inherently asymmetric, producing only one gamete, and only chromosomes that segregate into the egg will be inherited to the next generation, while the rest will be degraded in the polar bodies. This asymmetric fate of the chromosomes creates an opportunity for them to compete during the meiotic cell division to remain in the egg, providing the framework to drive the evolution of genetic elements that regulate the segregation such as centromeres. Indeed, centromeric DNA is the most rapidly evolving DNA sequence in eukaryotic genomes despite its conserved function in chromosome segregation. Although examples of meiotic drive are widespread across eukaryotic species (plants, insects, and mammals including humans) with significant impacts on chromosome evolution, the underlying cell biological mechanisms are largely unknown.
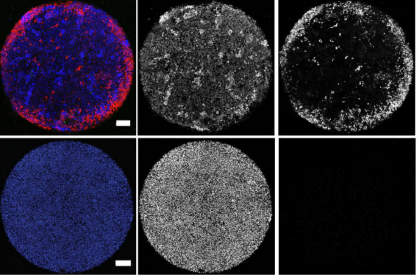
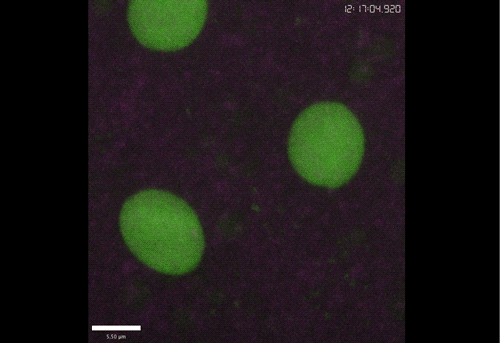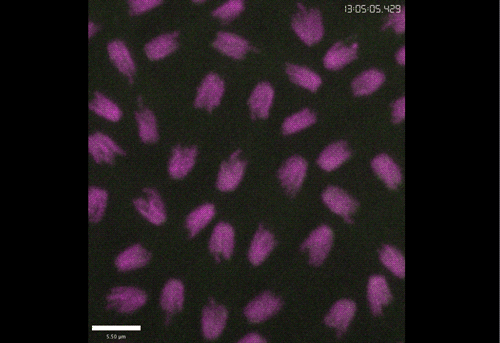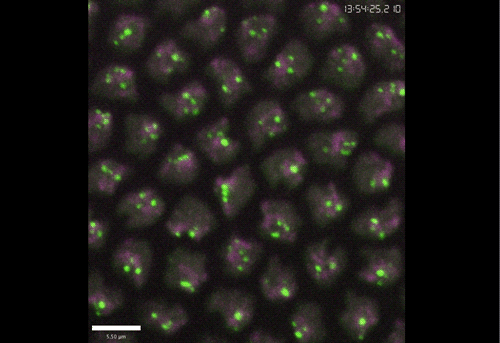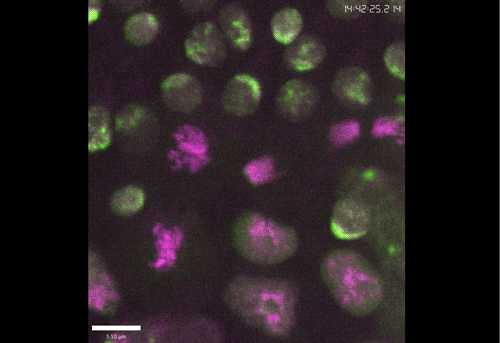
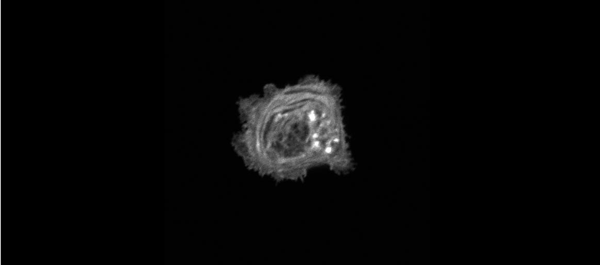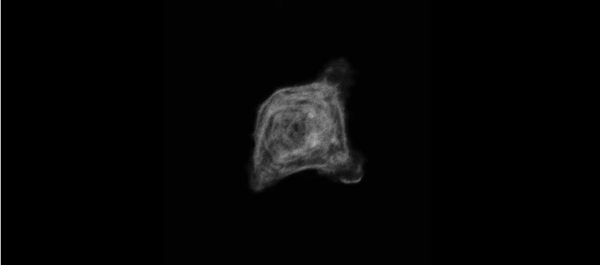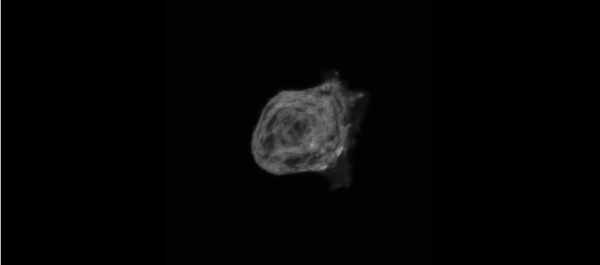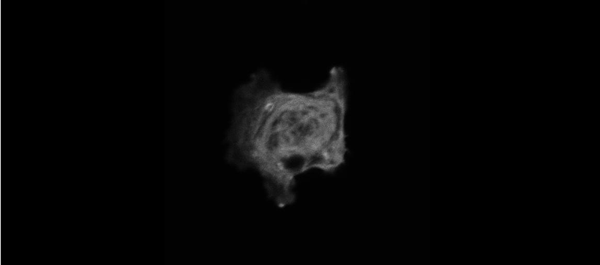
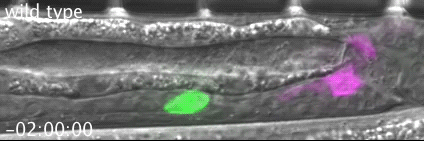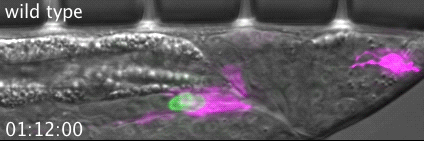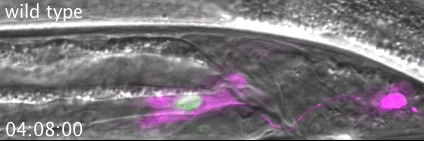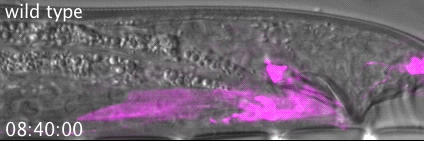
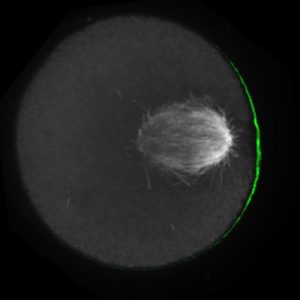
The meiotic drive project was a perfect fit for me, where I could use my expertise in chromosome segregation to tackle an exciting evolutionary biology question. When I joined the Lampson lab, I overlapped for several months with a former PhD student, Lukáš Chmátal, who established the system to study meiotic drive in mouse oocytes. This was a great opportunity for me to learn about meiotic drive and the mouse oocyte system, both of which were new to me. My long-term goal is to understand how a selfish centromere interacts with the microtubule spindle to preferentially remain in the egg. In theory, a selfish centromere needs to find out which side of the spindle will take it to the egg. This implies some kind of asymmetry within the spindle, but the mechanisms were mysterious. So, my first goal was to understand the molecular basis of the spindle asymmetry, which is the main topic of this post. This spindle asymmetry project was originally started by Lukáš who established the basis before I joined the lab. It was also strongly supported by the collaboration with Dr. Carsten Janke at Curie Institut, who is one of the leaders in post-translational modifications (PTMs) on tubulin, which composes microtubules. Carsten suggested that we test the possibility that a tubulin PTM is asymmetric within the spindle, and indeed this was the case! Tyrosinated (Tyr) α-tubulin was enriched on the cortical side of the spindle, which will end up in polar body, whereas detyrosinated (dTyr) α-tubulin was enriched on the interior side of the spindle, which will remain in the egg. Subsequently, I found that CDC42 GTPase that localizes to the polarized cortex (green in the figure below) is required for the asymmetry in Tyr α-tubulin (white in the figure). Therefore, we proposed a model where CDC42 is the master regulator of the spindle asymmetry, increasing Tyr α-tubulin specifically on the cortical side of the spindle.
To support this model, I aimed to induce an artificial spindle asymmetry by targeting CDC42 to one of the spindle poles. I decided to take an optogenetic approach, which was previously developed in our lab using culture cells. So, I thought, “Okay, I just need to apply it to mouse oocytes”. But it turns out, it was not that simple. Science is not that easy… But thankfully, I had great colleagues Huaiying Zhang (Lampson lab) and Chanat Aonbangkhen (David Chenoweth lab at Penn Chemistry), who were working on this optogenetic tool in cell culture and helped me a lot to optimize the light-induced targeting in mouse oocytes. After multiple rounds of trial and error, optimizing the laser intensity and the constructs for the targeting, I successfully targeted CDC42 specifically to one spindle pole and found that it is sufficient to induce asymmetry in Tyr α-tubulin consistent with our model. (I personally think this optogenetic tool will be useful to study female meiosis in general because it allows us to control protein localizations spatially and temporally, which is of course important in mouse oocytes as well as in other systems)
Then, the obvious question coming next is how this tyrosination asymmetry helps a selfish centromere orient towards the egg side of the spindle. By carefully analyzing the attachments between selfish centromeres and the asymmetric spindle, I noticed that attachments were unstable when the selfish centromere faces the cortical side enriched in Tyr α-tubulin, which is the wrong direction for the selfish behavior going to the polar body. Also, I found that tyrosination destabilize microtubules. Therefore, a selfish centromere is more likely to detach from the spindle when facing the wrong side because of the high tyrosination levels, providing an opportunity to re-orient to the right side, where it can stably attach because of low tyrosination levels (see the movie below by AAAS, which nicely summarize this work).
Now we know a little bit more about meiotic drive at least in our system, but there are many big questions remaining. For example, we still don’t know how CDC42 regulates tubulin tyrosination. The link between CDC42 and the tyrosination is new, and the underlying mechanisms are completely unknown not just in mouse oocytes but also in other systems. It would be interesting in future to examine whether CDC42 regulates the enzyme that catalyzes tyrosination. Also, it is mysterious why the spindle is asymmetric in the first place. There should be some function in female meiosis in general, and the selfish centromere is just exploiting that asymmetry. Furthermore, we don’t know what activity at the centromere confers selfishness. Previously, our lab in collaboration with Ben Black’s lab (Penn Medicine) showed that a selfish centromere has expanded satellite repeat sequences and recruits more centromere proteins compared to the centromere of the homologous chromosome. I am very curious which protein(s) at the centromere is actually important for their selfish behavior. The above projects would help answer my long-term question − how does a selfish centromere interacts with the spindle to win the competition in female meiosis.
Biased chromosome segregation is not restricted to meiotic drive, but also present in some asymmetric stem cell divisions (e.g. muscle stem cell), where the old DNA strand preferentially remains in the stem cell and the newly synthesized strand segregates to the differentiated cell. Since the mechanisms of biased segregation in these systems are unclear, it would be interesting to examine whether we can extend the concept of biased segregation that we learned from meiotic drive to other systems.
The aim of the Nice Conference is to provide a niche for presentations of the latest basic research in stem cell and developmental biology and to stimulate exchange of ideas.
Organized by Novo Nordisk Fonden together with DanStem, the aim of the Nice Conference is to provide a niche for presentations of the latest basic research in stem cell and developmental biology and to stimulate exchange of ideas by providing ample time for both formal and informal discussions.
The conference theme is the concept of the “niche”, in the broadest possible sense of the word, encompassing all sources of inputs stem and progenitor cells receive from their environment to expand or differentiate. This topic is both important and timely in light of the need for improved therapies against devastating diseases, such as diabetes, cancer, neurodegenerative and cardiovascular disorders. It is difficult to predict which basic research will lead to major medical breakthroughs, so future applications in medicine will rely on continued strong basic research.
Date: 27 May 2018, 5:00 pm to 31 May 2018, 9:00 am
Registration fee, accommodation and local costs for all approved applicants are covered by the Novo Nordisk Foundation. Only travel expenses to and from the conference venue at own expense.
 (No Ratings Yet)
(No Ratings Yet) (1 votes)
(1 votes) After completing her PhD studies investigating Drosophila nephrogenesis in Helen Skaer’s lab in Cambridge, Helen moved to Bristol in 2013 to take up a 5 year, MRC-funded post-doc position between Paul Martin’s and Will Wood’s labs. Her first publication from this work (
After completing her PhD studies investigating Drosophila nephrogenesis in Helen Skaer’s lab in Cambridge, Helen moved to Bristol in 2013 to take up a 5 year, MRC-funded post-doc position between Paul Martin’s and Will Wood’s labs. Her first publication from this work ( An effective inflammatory response is pivotal to fight infection, clear debris and orchestrate the repair of injured tissues; however, inflammation must be tightly regulated since many human disease pathologies are a consequence of inflammation gone awry. Using a genetically tractable Drosophila model, I use precise genetic manipulation, live imaging and computational modelling to dissect the mechanisms that activate the inflammatory response to tissue damage and those that simultaneously protect the regenerating tissue from immunopathology. Upon tissue damage, immune cells (particularly neutrophils and macrophages) are recruited into the damaged area by damage signals (danger-associated molecular patterns, DAMPs) released from the injured tissue. In collaboration with computational biologists, we employ a sophisticated Bayesian statistical approach to uncover novel details of the pro-inflammatory wound attractants, by analysing the spatio-temporal behaviour of Drosophila immune cells as they respond to wounds. We show that the wound attractant is released by wound edge cells and spreads slowly through the tissue, at rates far slower than small molecule DAMPs such as ATP and H2O2. Strikingly, we also find that immune cells must be developmentally ‘primed’ by uptake of apoptotic corpses before they can respond to these damage attractant signals. Such corpse-induced priming is an example of “innate immune memory” and may serve to amplify the inflammatory response in situations involving excessive cell death – and otherwise limit an overzealous and damaging immune response. Indeed, whilst inflammation is clearly beneficial, toxic molecules (e.g. reactive oxygen species, ROS) generated by immune cells to fight infection, can also cause significant bystander damage to host tissue and delay repair – and may underpin chronic wound-healing pathologies in the clinic. To counter this, I find that wounded Drosophila tissue employs a complex network of cyto-protective pathways that promote tissue ‘resilience’, which both protect against ROS-induced damage and stimulate damage repair. Successful tissue repair, therefore, not only relies on the host’s ability to mount an effective inflammatory response, but also its ability to finely tune it and limit associated immunopathology.</div?
An effective inflammatory response is pivotal to fight infection, clear debris and orchestrate the repair of injured tissues; however, inflammation must be tightly regulated since many human disease pathologies are a consequence of inflammation gone awry. Using a genetically tractable Drosophila model, I use precise genetic manipulation, live imaging and computational modelling to dissect the mechanisms that activate the inflammatory response to tissue damage and those that simultaneously protect the regenerating tissue from immunopathology. Upon tissue damage, immune cells (particularly neutrophils and macrophages) are recruited into the damaged area by damage signals (danger-associated molecular patterns, DAMPs) released from the injured tissue. In collaboration with computational biologists, we employ a sophisticated Bayesian statistical approach to uncover novel details of the pro-inflammatory wound attractants, by analysing the spatio-temporal behaviour of Drosophila immune cells as they respond to wounds. We show that the wound attractant is released by wound edge cells and spreads slowly through the tissue, at rates far slower than small molecule DAMPs such as ATP and H2O2. Strikingly, we also find that immune cells must be developmentally ‘primed’ by uptake of apoptotic corpses before they can respond to these damage attractant signals. Such corpse-induced priming is an example of “innate immune memory” and may serve to amplify the inflammatory response in situations involving excessive cell death – and otherwise limit an overzealous and damaging immune response. Indeed, whilst inflammation is clearly beneficial, toxic molecules (e.g. reactive oxygen species, ROS) generated by immune cells to fight infection, can also cause significant bystander damage to host tissue and delay repair – and may underpin chronic wound-healing pathologies in the clinic. To counter this, I find that wounded Drosophila tissue employs a complex network of cyto-protective pathways that promote tissue ‘resilience’, which both protect against ROS-induced damage and stimulate damage repair. Successful tissue repair, therefore, not only relies on the host’s ability to mount an effective inflammatory response, but also its ability to finely tune it and limit associated immunopathology.</div?





 (6 votes)
(6 votes)