Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our eighth report from the 2017 group of student awardees comes from Nicole Serzhantova(student at The University of Edinburgh), who undertook her studentship withJennifer Nicholsat The University of Cambridge.
Understanding the first step in the formation of organismal complexity in the mouse embryo
As biologists, we are very used to poking and prodding very complex systems, be it drosophila or homo sapiens. Rarely is it a prerogative to take a step back and really understand how such complexity arises seemingly out of nowhere. How is that we start off from a single cell and over a period as short as 48 hours (in the mouse at least) we already start seeing complexity arising?
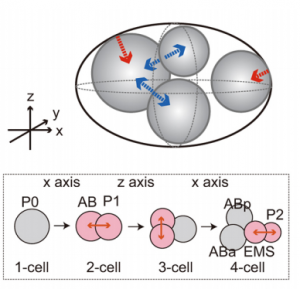
In mice, a single-cell zygote undergoes a series of cleavage cell divisions and morphogenetic changes to form a unipotent epithelial vesicle termed the trophectoderm (TE) enclosing a compact group of pluripotent stem cells known as the inner cell mass (ICM) (Fig.1F).
Figure 1. Development of the mouse embryo from one cell pre-implantation stage to a post-implantation stage embryo. (Rossant & Patrick, Development, 2009)
Restriction of cell fate takes place over an extended time period of 48 hours from the 8-cell (morula) stage (Fig.1C), when all cells are indistinguishable from one another and are capable of forming all three germ layers. This feature of cells is termed totipotency and is a transient characteristic of embryonic stem cells before they commit to become either TE or ICM.
At this 8-cell stage compaction of the embryo begins, in which the embryo surface smoothens because of an increase in intercellular adhesion. Cells epithelialise, forming tight junctions (which acts as a seal) between cells, limiting diffusion and what you are left with is fluid-filled epithelial vesicle enclosing a small number of cells (Fig. 1D-F).
It is at point that the first commitment of cells occurs, whereby the outer epithelium (Fig.1E) becomes committed to form the trophectoderm (TE), which subsequently gives rise to the trophoblast layers of the placenta and the trophoblast giant cells. Whilst the small population of enclosed cells commit to becoming the ICM, which is the pluripotent stem cell lineage of the embryo, giving rise to all of the primary germ layers of the foetus and its extraembryonic tissue.
At this stage of development, protein synthesis from maternal mRNA becomes more transient as the embryo’s own gene programs begin to switch on. Once commitment occurs, as the morulae becomes a blastocyst, differential gene expression is seen between the ICM and TE. At day 2 ½ post-fertilisation, all cells within the morula uniformly express most genes, including the pluripotency factor Oct4 and the TE marker Cdx2. Interestingly, at day 3 ½ the first real differential gene expression is observed, where the ICM begins to express Oct4 whilst loosing its ability to express Cdx2 and conversely the trophectoderm begins to express Cdx2 whilst loosing all Oct4 activity. There is a mutual repression by Oct4 and Cdx2 which further consolidates the TE and ICM segregation.
Elucidating the mechanism governing a cell’s ability to become either a unipotent TE cell or a pluripotent stem cell of the ICM can open up new avenues for generating and maintaining pluripotency in vitro. In an attempt to understand this initial lineage segregation, a stem cell line was cultured which removes the Oct4 gene upon treatment with tamoxifen. This cell line also had a florescent tdTomato reporter under the control of a constitutive promoter, meaning that these cells fluoresce red when exposed to light in the blue to green range.
The following cells were cultured in serum-LIF (a differentiation permissive medium) and different portions of the cells were treated with tamoxifen for either 24, 48 or 92 hours to allow for the excision of the Oct4 gene. Once these time points of treatment were reached, cells were microinjected into live mouse morulae and blastocysts. Approximately 5 morulae and blastocysts were microinjected with 8-10 stem cells deficient in Oct4 from each time point as well as un-induced cells form the same batch as a control. These morulae were left for two days to allow for compaction and the first lineage segregation to occur. These were then promptly fixed and immunostained to examine what effect the deletion of Oct4 had on the cell fate choices that the injected stem cells made.
Following what we already know about what happens during development, it can be hypothesized that stem cells that do not have the capacity to make Oct4 will commit to the trophectoderm lineage as they are missing a key pluripotency factor and therefore cannot become pluripotent cells competent of forming all 3 germ layers.
Unfortunately due to rising temperatures in the laboratory during summer (the air conditioning was of course broken on the days the microinjections were performed), upon imaging it was clear that in most cases only 1 stem cell, out of the 8-10 injected, integrated into the embryo. This was prevalent in the un-induced cells also, suggesting that it was not the tamoxifen treatment that caused toxicity to the cells but rather the experimental conditions. Integration rates were higher in morulae than in blastocysts as expected, since by the blastocyst stage cell commitment has already occurred making it harder for integration to ensue.
Out of the cells that did integrate, only 1 appeared to express Cdx2 markers showing that it has adopted a trophectodermal cell fate, whilst the others exhibited a mixture of pluripotency markers. The conclusion from this being that although Oct4 is necessary for cell commitment to the ICM lineage, additional molecular events must underpin this initial commitment in the mouse model. Inferring this from the data is difficult considering how the number of integrated stem cells was so low and future experiments with more tightly controlled conditions are advised.
These results however are concordant with other studies, with Wu and Schöler (2014) reporting that the “establishment of totipotency in maternal Oct4–depleted embryos was not affected, and that these embryos could complete full-term development without any obvious defect.” In addition to this, Wu and Schöler were able to form Oct4 expressing inner cell masses in embryos with complete inactivation of both maternal and zygotic Oct4 expression as well as reprogramming of fibroblasts into fully pluripotent cells by Oct4-deficient oocytes.
This, in conjunction with the results I obtained, tends to indicate that Oct4 is not essential for the initiation of pluripotency but in contrast to its role is critical for the maintenance of pluripotency.
I would like to thank Professor Jennifer Nichols, Ayaka Yanagida, Peter Baillie-Johnson, Thorsten Boroviak, Ken Jones, Tim Lohoff and everyone else at the Cambridge Stem Cell Institute for their invaluable guidance and support throughout my internship. I would also like to thank the British Society for Developmental Biology for granting me the Gurdon Studentship Award without which none of this would have been possible.
Built into the side of a tree covered hill on a remote Japanese island lies what at first sight appears to be a super-villain’s lair, or perhaps the HQ of Jurassic Park. In fact we had arrived at the Okinawa Institute of Science and Technology (OIST) for their 2 week Developmental Neurobiology Course. DNC is a 2 week intensive lecture and lab based course designed to teach, inspire and foster collaborations between the next generation of developmental neurobiologists.
OIST (Photo credit OIST website)
23 of us were accepted, a highly diverse collection of PhD students and Postdocs from all over the world to spend two weeks in paradise all there because of our love for developmental neurobiology. The course was split into two halves: Week one – lectures in the morning and a lab project in the afternoon. Week 2 – lecture bootcamp! 6-7 hours of fascinating lectures a day by some of the most distinguished researchers in the field on a vast variety of topics.
While the classes were intense there was plenty of downtime to interact, consolidate and simply think about the wider field beyond our individual specialities, with plenty of swimming off the beach attached to our accommodation! In this post I will try to summarise the course and hopefully explain why I think courses like this are so crucial for early career researchers to participate in.
Week 1:
After 27 hours of travelling I finally arrived at our Seaside House accommodation. OIST generously covered all course costs, accommodation, food and half of travel costs for all participants; a critical factor in making it accessible to early career researchers.
We started our first day with intensive short talks from all participants so we all knew a little bit about each other’s background. The range of specialities was huge, from pure molecular biologists to electrophysiologists, to using Zebrafish to study dyslexia, making for some fascinating discussions throughout the 2 week course. We got the chance to learn more about each other’s research the following day, where after a fascinating lecture on microscopy by Ulrike Engel, we gathered for a poster session to present our research to staff and participants.
A key part of DNC is the opportunity to learn new techniques that we might not otherwise get the opportunity to learn. This was taught through a choice of one of five lab projects, but before committing we got a brief rotation in each lab to learn more. The five lab project options available to us were:
Analysis of Drosophila embryonic neuromuscular development
Ca imaging and electrophysiology recording from Drosophila Larvae
In utero electroporation in mouse
Live imaging of zebrafish nervous system
Electrophysiology in zebra finch auditory system
I was lucky enough to be taught in utero electroporatation in mouse by Hiroshi Kawasaki, a powerful technique for highly spatially specific gene editing during development.
Having been assigned our lab projects the week continued with lectures in the morning and lab projects in the afternoon. Our lectures were fascinating and varied:
Cecilia Moens (Fred Hutchinson) spoke about cranial motor neuron development in the Zebrafish.
Lisa Goodrich (Harvard) explained about the peripheral nervous system, focusing on cochlear development.
Hiroshi Kawasaki (Kanazawa University) told us about his research focused on the molecular mechanisms controlling cortical development.
Michael Granato (U Penn) talked about axon guidance and regeneration in Zebrafish.
For me the length (3 hours) of the lectures was key to the immersion of the course. This time period gave the lecturers the chance to speak about their long research journey and all the side stories that would normally be left out of a shorter lecture. Although intense, every hour we were rewarded (behavioural conditioning?) with coffee and snacks, we had the opportunity to ask questions at any point, and the lecturers were able to enter into group discussions without fear of time constraints.
Embryos from the Zebrafish lab project
In the afternoons we utilised the fantastic laboratories at OIST to work on our lab projects with the help of our group leaders and the fantastic Postdoc tutors. Under their careful tuition we practised and perfected our techniques, and gathered our data to present at the end of the course. The resources available at OIST were staggering, we were able to perform CLARITY on our electroporated brains and image them with a light-sheet microscope, while the Drosophila group used super-resolution to image individual boutons in the Drosophila larvae!
This slideshow requires JavaScript.
Lab projects complete, we celebrated the end of the first week with a barbecue and fireworks at Seaside House, and of course Japanese karaoke with the course organisers!!
The in utero electroporation team with our instructor Hiroshi Kawasaki
Week 2: Lecture Bootcamp!!
Before the intensive days of lectures started, we were given a day off to recuperate and explore Okinawa. For a couple of us that meant a relaxing trip to an Onsen, for others Scuba-Diving, and for one adventurous group it involved a multi-stop tour to a pineapple farm, a Dragonball Z-themed butterfly park (…?) and the Okinawa aquarium.
The Dragonball-Z butterfly park
Re-energised after our free day we threw ourselves into the second week of lectures, learning about ideas and concepts that we had never encountered in our day to day research:
Yimin Zou(UCSD) spoke about how we must understand the processes controlling axon guidance during development, to investigate how circuits may repair themselves following injury.
Jeff Wickens (OIST) gave a lecture on the neural mechanisms that underlie learning and adaptation in the mammalian brain.
Kang Shen (Stanford) talked about the molecular mechanisms behind dendrite patterning and how they wire into neural circuits.
Tomoyuki Takahashi (OIST) we were privileged to be given a lecture by one of the founders of electrophysiology, a fact made clear by the massive increase in audience size as many OIST researchers came to hear him speak.
Mei Zhen ( University of Toronto) described her research on understanding how the C.elegans neural motor system translates into movement.
Greg Basaw (U Penn) gave as another axon guidance lecture, this time from the perspective of receptor regulation during Drosophila development.
Anne Hart (Brown) gave one of my favourite lectures as she described using a wide range of models from C.elegans to mice to study disease mechanisms in a highly interactive manner calling on us to think of research ideas.
That Friday afternoon we gave our brains a chance to consolidate as we were taken on an excursion to Shurijo Castle to experience more of the island’s rich history before enjoying a traditional Okinawa dinner with the course organisers and lecturers in the capital Naha.
The next day we received news, after several days of speculation, that there was a typhoon passing between the Okinawa prefecture and mainland Japan. OIST is built for this weather so we continued on with the course albeit with an embargo on all water activities.
Hitoshi Okamoto (Riken) gave a fascinating lecture on social conflict in Zebrafish and the role of the habenula in this. To me this was particularly interesting as I had never considered this behavioural use for the Zebrafish before.
Tadashi Isa (NIPS) gave our penultimate lecture on how networks reorganise to recover movements following spinal cord injury.
Yoko Yazaki-Sugiyama (OIST) delivered our final lecture on critical periods in Zebra Finch auditory systems during song learning.
As lectures concluded on the final day we concluded the academic side of the course by giving presentations on our lab projects from the first week. This gave us an excellent opportunity to show the skills we had developed, consolidate our knowledge and thank our teachers and tutors for their guidance and support.
We gathered for a final dinner together in Seaside house with traditional Okinawan entertainment as we ate. After dinner we had final words from the course organisers who presented us with certificates of our participation. This also gave us time to thank them extensively for all their work organising the course and providing us with the most incredible learning opportunity that will shape our careers as neurobiologists. We ended the course as a team, with fireworks and karaoke as the lightening storm from the typhoon raged around us.
DNC 2017 team photo
This course is such a fantastic opportunity for young researchers. The vast diversity of lectures will broaden your horizons and bring new directions to your project and future plans. The people I met will be my friends and colleagues for the rest of my career and I hope one day I will get to publish alongside them as we collaborate together. I cannot thank the course organisers; Yoko Yazaki-Sugiyama, Ichiro Masai, David Van Vactor, Yimin Zou and Hiroshi Kohsaka enough for this opportunity as well as all the support staff who made it happen. I encourage all young developmental neurobiologists to apply to this wonderful course, you will not regret it!
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our seventh report from the 2017 group of student awardees comes from Agata Czap (student at University College London), who undertook her studentship withPaola Oliveri at University College London.
How to knockout a gene in sea urchin?
I am Agata Czap, an undergraduate student of MSci Human Genetics at UCL. The BDSB Gurdon Summer Studentship has given me an incredible opportunity to undertake research for 8 weeks at Oliveri lab in UCL. I applied to work with Dr. Oliveri to learn about the challenges of applying the CRISPRcas9 technology to new model systems.
My project focused on knocking out the transcription factor Pmar1 in sea urchin S. purpuratus using the CRISPRcas9 system. Pmar1 is a part of a characterized gene regulatory network (GRN) and its role has been dissected using ectopic and dominant negative approaches (1). Specifically, pmar1 represses HesC during early development, as part of a double negative gate (DNG) of skeletogenesis. HesC is another transcription factor, which represses the downstream skeletogenesis genes (2). This DNG enables specification of micromere cell fate in the correct cells (1), from which primary mesenchyme cells (PMCs) will arise and eventually form the larval skeleton. Furthermore, the DNG enables micromeres to release two signals. Early Signal (3), which is responsible for inducing archenteron invagination, and Delta, which activates the Notch receptor in secondary mesenchyme cells for endomesoderm patterning (4) (figure 1).
Figure 1: Pmar1 is a part of double negative gate (DNG) in the skeletogenesis pathway. This gene requires β-catenin, and another transcription factor Otx, both maternally expressed. These two determinants bind to promoter of Pmar1 for activation of its expression. Pmar1 represses transcription factor HesC in micromeres. This repression enables expression of skeletogenesis genes and release of two signalling proteins to induce endomesoderm patterning in secondary mesenchyme cells (SMCs) and archenteron invagination.
So far, there have been no functional knockouts of Pmar1, as the gene has duplicated 4 times in the S. purpuratus genome. Previous experiments deduced possible function with a series of downregulation and rescue indirect strategies (1,4). Micromeres are located on the vegetal plate of the urchin embryo. One of early experiments carried by Dr. Oliveri, involved removing and transplanting micromeres into animal pole of another healthy embryo (2). This has induced a secondary, ectopic gut (2). Additionally, an embryo from which micromeres have been removed had shown no development of skeleton nor gut (3). Pmar1 expression requires maternal determinant called β-catenin, and imparing its nuclerization results in an embryo similar to micromereless embryo (3). Thus, I have hypothesized that upon successful knockout, the embryo would lack skeleton as well as other endomesoderm derivatives (e.g archenteron). Other ectodermal tissues should be unaffected.
To tackle the issue of pmar1 duplication, I have collected all 4 (Pmar1a-d) FASTA sequences from the Echinobase transcriptome. I have used Jalview to identify conserved regions between duplicates within exon 1 and 2, and generated phylogram to visualize diversification of the sequences. My results agreed with that of Cavaleri’s team (4); Pmar1b has diverged to become a monophyletic group. The sequences are still highly conserved, the first 810 bp of each transcript show 86% similarity. I used this fragment of the Pmar1b sequence, as it had the best consensus when compared to cDNA. I have inserted this sequence into 4 algorithms including CHOPCHOP to generate gRNAs. Unfortunately, the algorithms used different criteria to rate gRNAs. To filter through all 98 gRNAs, I tested each against BLAST in Echinobase for off-targets. This allowed me to obtain top 5 gRNAs, which target every Pmar1 gene.
A more wet lab-based part of my project then followed. During the first two weeks, I have learned how to culture sea urchin embryos, spawn adults to obtain gametes and assess fertilisation rate. S. purpuratus has turned out to be a rewarding model system, embryos can be produced in large quantities and are transparent, allowing clear observation of tissues. Natalie, a PhD student who also worked on applying CRISPRCas9 technology to knockout neuropeptides, has taught me how to synthesize selected gRNAs using PCR and T7 in vitro translation. Once gRNAs were ready, we injected a pair of gRNAs and Cas9 mRNA into oocyte after fertilisation to induce a deletion mutation. The efficiency of knockout was tested with DNA extraction and agarose gels, which measured molecular weight of the target region. I carried out in situ hybridisation and antibody staining on genes acting downstream of Pmar1 to characterise mutants.
My results were promising. In each knockout, I have used PKS1 knockout in pigment cells as positive control, which proved Cas9 enzyme is non-toxic for embryos. PKS1 allows quick visual check that Cas9 works, as albino mutants are easily identified. Furthermore, it has high knockout efficiency of ~90% (5). My knockout attempt had lower PKS1 efficiency with 50% albino incidence at 45hpf. I have used additional positive control, knockout of Jun, a gene downstream of Pmar1, which caused delayed invagination in 42% at 45hpf. Pmar1 gRNA 1 and 5 have been very efficient; at both 45hpf and 70hpf, 100% of embryos were mutants. The embryos did have a blastopore and syncytial rods were present, but the gut nor skeleton were not developed (figure 2).
Figure 2: My attempt to knockout Pmar1. Jun and PKS1 knockouts act as positive controls. PKS1 knockout generated albino mutants. Jun knockout has resulted in delayed invagination characterised by undeveloped shorter gut. Pmar1 knockout caused lack of proper skeleton and gut, and round morphology of the embryos.
In conclusion, a successful knockout requires a lot of preparation, resilience and knowledge of molecular tools. The procedure gives you greater understanding of how Cas9 enzyme works and of gene function. My gRNAs will be further used by Oliveri lab to test efficiency of Pmar1 knockouts using different combinations of gRNA pairs and Cas9 concentrations.
I would like to thank Dr. Oliveri for giving me a chance to work in her lab. Also to PhD student Natalie Wood for being patient when teaching me all the techniques used. I greatly enjoyed being part of a research team and wish to pursue scientific career further. I would recommend Gurdon studentship to any student applying for a laboratory-based internship.
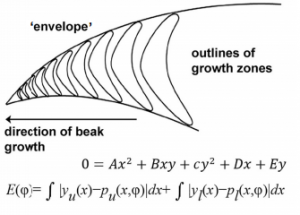
Pavement cells in plant leaves were identified as a puzzle which deviated from normal cell shape rules by D’Arcy Thompson in his classic text On Growth and Form. Now modern approaches allow researchers to revisit these problems and try to uncover the rules that govern pavement cell topology during leaf development. This week we feature a paper – published in the special issue of Development celebrating the centenary of On Growth and Form‘s publication – that uses live-imaging and modelling to understand the development of pavement cell topology. Co-first authors Ross Carter and Yara Sánchez-Corrales, and PIs Verônica Grieneisen and Athanasius (Stan) Marée, told us more.
Ross Carter, Yara Sánchez-Corrales, Verônica Grieneisen and Stan Marée
Stan, can you give us your scientific biography and the main questions your lab is trying to answer?
AM Despite enjoying mathematics and being a computer “nerd” during high school, I initiated my academic trail studying Biology at the Utrecht University, the Netherlands – as I was, and am, most fascinated in how life processes. After performing experimental work on neural vision in cats, I realised I really was a theoretical biologist. I modelled the full life-cycle of Dictyosteliumdiscoideum, and found the driving question for the rest of my carrier: how do individual cells operate collectively to form complex structures. In a broad sense, my lab uses multi-level modelling of morphogenesis to answer this, in both plants and animal systems. We find that it is particularly important to contrast independently evolved multi-cellular systems, and to work closely to experimental data. So my lab brings together intra- and intercellular communication, cell mechanics and motility, networks and cell differentiation, and cell shape and cell topology, to eventually understand how tissues, organs and even whole organisms are shaped.
And Verônica, what about you?
VG My origins lie in Physics, which I studied in my birth country, Brazil. There I got extremely interested in Statistical Mechanics and Complex Systems. Before I realised it, I ended up meeting Paulien Hogeweg – founder of Bioinformatics – with whom I did my PhD, in Theoretical Biology. This work was done in close collaboration with Ben Scheres, who introduced me to the hidden beauties of Arabidopsis thaliana. I never looked back. Plants are a great system to investigate problems of pattern formation and emerging morphogenesis, with an added twist: their interaction with the environment, for example foraging, light competition, etc, has to go through growth, which means that their development is extremely plastic. My lab seeks to understand this, and we have found that a key integrator of the necessary information processes stems from cell and tissue polarity. My lab explores issues of cell and tissue polarity (for example the development of the Pavement cell), by combining experiments and imaging with computer simulations (like the work here), and even taking an morphogenetic engineering standpoint.
Ross and Yara, how did you come to be involved in the project?
YSC I did my PhD at the John Innes Centre (JIC) under the supervision of Veronica Grieneisen and Enrico Coen. My research focused on investigating the role of different cellular processes during tissue growth. It was very important to obtain data on the dynamics of cell shape morphogenesis and divisions, so I devoted a considerable amount of my PhD to optimize live-imaging of the developing leaf for long time periods. In parallel, I had to find ways to extract meaningful information from the time-lapse data. I had a chance to develop some tools to analyse cell shape dynamics and was extremely lucky to get access to other computational tools that were being developed by Matthew Hartley and Stan’s lab, and were crucial for this research. We started addressing the problem of topology by the end of my PhD, but the real push for this work came when Ross joined Veronica and Stan’s labs. By then, I had already moved to Cambridge.
RC I’ve been interested in leaf development since the start of my PhD in 2010 and after joining Veronica and Stan’s groups in 2015 was given access to an amazing dataset collected by Yara. These time-lapse images collected throughout leaf development really allowed us to address some unanswered questions. Inspired by some work by Matthew Gibson (Nature 2006) we investigated whether our data fit with the topological models they were proposing. When we found that it didn’t we began to investigate why, and the fact that we heard about this special issue allowed us to frame the research to unanswered questions in ‘On Growth and Form’, specifically the puzzling shapes of leaf pavement cells.
Your paper features in Development’s Special Issue celebrating the centenary of D’Arcy Thompson’s On Growth and Form. How has Thompson’s work influenced your thinking about plant development, and what makes his ideas relevant today?
AM D’Arcy Thompson has influenced my work in more than one way, which is remarkable. One of the premises of his book is that living matter also needs to abide to physical laws, and he shows how surface tension can be regarded as an important driving force. While he provided proofs by analogies, my lab follows this inspiration, using the Cellular Potts Model, among others showing how formally it can be linked to Cell Surface Mechanics in Biology. Secondly, D’Arcy is sometimes criticised by not taking genes into account. In fact, he clearly mentions that differential activity of growth and mechanics is what is shaping the organism (and not the gene itself), and I think he is completely correct about that. The control information cannot simply be explained by a gene being expressed. My lab also takes this point of view, but linking gene regulatory networks – which we analyse through dynamical systems theory – to the actual growth and development of the tissue. And thirdly, it was D’Arcy which first showed in quantitative detail the similarities in the cellular distributions among neighbouring cells between biological and physical cellular matter. This publication is an example of what kind of unexpected discoveries we can make when following such an analysis by modern means.
VG I believe that D’Arcy Thompson was visionary in the way he was making links crossing disciplines, and bringing together very diverse species to show commonalities. I sometimes like to imagine how delighted he would be if he could see the approaches we currently have in the lab to actually follow up some of his insights: tracking of cell dynamics, computational models to test ideas, and powerful shape analysis methods. However, I am also constantly reminded of something he points out throughout his book: that observing something very elaborate and complex does not imply that the mechanism itself is complex to the same level. His message is something that guides my labs research constantly: are we capturing the phenomenon correctly, are we missing something essential, is the data distracted us?
And finally, D’Arcy Thompson’s ability to look across species to bring forth relevant relations is something we lost, as each field dived into its particular model system. With more advanced molecular methods, however, we will hopefully gain the ability again to zoom out, without losing depth. I am excited about these promising possibilities.
In fact, my first contact with the book was when I was an undergrad physics student, when I came across a worn-out, dog-eared copy of his book in the library. I immediately was totally fascinated by it, actually causing me to enter the field of biophysics during my masters. (Please note that I also gave a radio 4 interview on how he has inspired me.)
Why do pavement cells pose a particular problem or puzzle for those interested in cell topology?
AM & VG Historically, and based on physics, topology is often seen as emerging from cellular interactions that effectively capture surface-minimizing processes, together with mathematical constraints of tessellating structures. Pavement cells, with their extreme perimeter to area ratios, clearly do not fit in a picture of energy minimisation. Moreover, throughout the tissue and over time there are huge differences in cell area. Together this should give rise to a very broad topology distribution which moreover should change dramatically over time. The reality, however, turns out to be totally different.
YSC The peculiar morphology of these cells and the fact that they cannot exchange neighbours made them a very intriguing model to test whether they could follow the typical topological pattern found in animal cells. I was very “puzzled” by these cells, I wrote a blogpost about them some time ago for Botany One.
RC A lot of previous work on cellular topologies has been done either on animal tissues, where membrane tension dominates or on tissues with regular cellular shapes. Pavement cells have neither of these properties and allowed us to address the question of whether these processes are important. These cells start off as quite regular oblong shaped cells and gain their characteristic jigsaw puzzle shape once they have differentiated. We were interested in whether the topological fingerprints we see were due to the shape of the cells or how they ‘choose’ to divide.
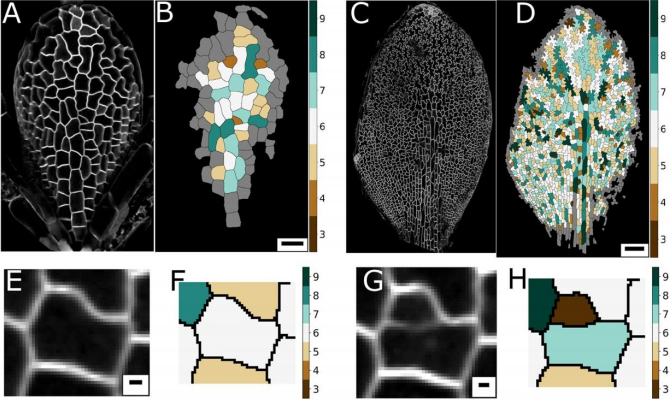
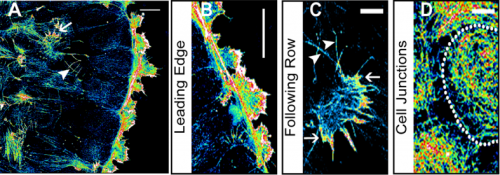
Confocal image of pavement cells. From Figure 1, Carter et al, 2017
Can you give us the key results of the paper in a paragraph?
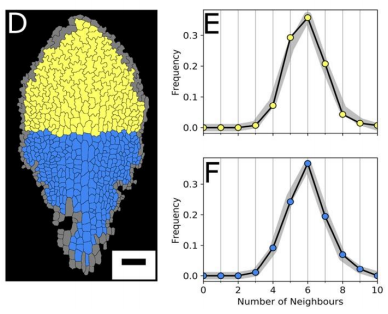
AM & VG Our results surprisingly showed that PC topology establishes very early on during development, independent on the rounds of cell division and pre-established before the cells acquire complex shapes and large differences in cell sizes. At the same time, we found that the topology is unique, compared to other species or other Arabidopsis tissues. This could only be explained if cell divisions are actually (directly or indirectly, this remains an open, exciting question) taking the topological organisation into account when they divide. So, the cell wall is carefully placed while “counting” its neighbours. We show this both through modelling the division dynamics and comparing it to the real date of more than 50,000 cells, and through further confirming it by tracking the division events themselves. The observation that tissues “care” about their topology offers a whole new search image to think about tissue development and functionality.
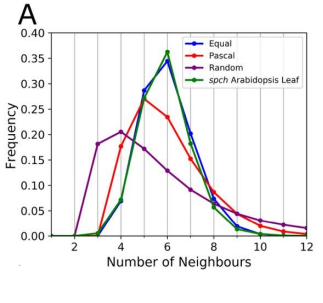
YSC We used live-imaging and modelling to study cell topology in a developing plant tissue. Surprisingly, the leaf epidermis has a unique topological signature that is independent of stage, position along the leaf and cell shape. We tested different hypothesis and found that neighbourhood relationships in the leaf can be recapitulated by implementing simple rules of cell division. Therefore, we concluded that the topology in the leaf is mostly determined by cell division rather than cell-surface mechanics.
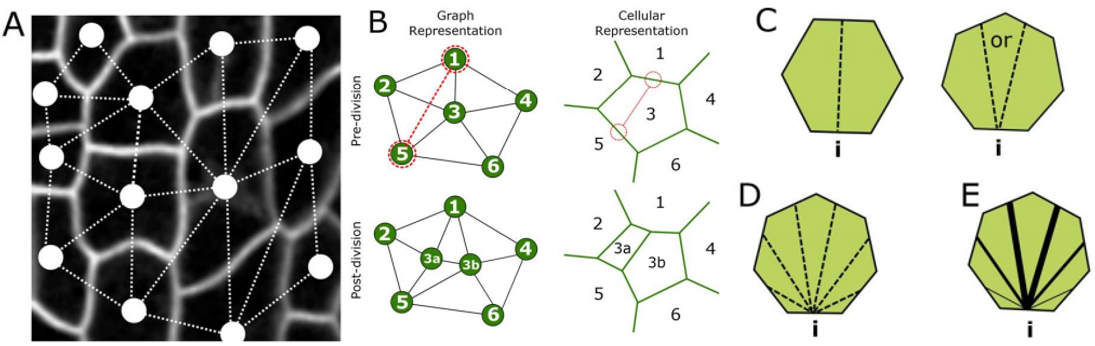
RC We developed an abstract topological model of cell division to test whether or not this can recapitulate the topological distributions we see in the developing leaf. Surprisingly we found that a simple division model, which splits a cell’s neighbourhood in half, was able to reproduce the topological distributions we see with remarkable accuracy. We hope that this will encourage people to think about topology when looking at cell division as apposed to cell geometry and mechanics.
Leaf topology is generated through divisions. From Figure 2, Carter et al, 2017
Do you think your model’s insights are restricted to pavement cells (or similar ‘misfits’), or can it explain other topologies in plant and animal tissues?
AM & VG We think that the particular way tightly coordinated division decisions strictly preserve the pavement cells’ unique neighbourhood distribution, might be quite specific for this system, likely linked to the particular cell shapes the PCs later acquire. However, from our study as well as other studies, it starts to emerge that complex mechanisms might operate in order to establish the right topology. The lack of surface tension-driven processes and the large variation in cell sizes allowed us to lay this bare much more clearly for this system, but topology control itself might be much more generic. Future collaborations and interdisciplinary studies will be needed to shed more light on both the how and the why.
YSC I think the insights gained with our set of graph models would help contrasting topological profiles in other epidermal tissues.
RC The rules we implemented in our model give the shallowest and broadest topological distributions (neighbour distributions) of any model we have seen, both the geometrical and the markov models. We did compare our results to some previously published data, and found that there were some similarities, for example with Anthurium, but this data is quite limited. The benefits of the modelling framework we’ve introduced here are that it’s relatively easy to add new topological division rules and, as long as you have a segmented image of a tissue, to compare it with cellular topologies of other organisms, both animal and plant.
Graph models for cell division. From Figure 4, Carter et al, 2017
If cell division history can explain the observed topology of pavement cells, do you have any idea about the molecular rules of the topology? (One might propose spindle regulators/cell division kinases etc. to have a role)
AM & VG A friend and collaborator of ours, Alan Champneys, proposed to us the idea of a “tent model”, in which neighbours can be “counted” by assessing the location of the triple junctions, similarly to how a tent is positioned by pegs. Molecular processes could thus arise due to cytoskeletal components. Alternatively, it is also possible that substances diffuse amongst cells, for example through plasmodesmata, reaching certain distributions only if the topology is correct, biasing division otherwise.
YSC Our Equal split neighbour division rule fits very well the observed topological distribution in the leaf. There are two possibilities to explain this in biological terms: 1) cells are assessing their number of neighbours after division using an unknown molecular mechanism or 2) our observations are a side effect of other underlying division and growth mechanisms acting together.
In the first scenario, there are some examples of molecular players described in the fruit fly that are expressed at tricellular junctions and could work as ‘topological sensors’. In plants, some level of coordination (and neighbour ‘counting’) could be achieved through plasmodesmata.
We favoured the second scenario and believe that our work calls for explanations that reconcile and integrate cell shape and topology into account. Regarding the molecular basis of this, I think it is still an open question.
RC One idea we did come up with we termed the ‘tent model’ of cell division. The number of triple junctions formed on the cell membrane, where it joins with other cells, is equal to the number of neighbours for that cell. These junctions have been shown to exert a tension on the cell membrane. If a cell can somehow measure this tension locally, possibly by use of mechanosensitve ion channels, it could place the mitotic spindle as to divide these neighbours in half.
Ross and Yara, when doing the research, did you have any particular result or eureka moment that has stuck with you?
YSC I was very surprised to know that the complex shape of the PC cells has no influence in the topological profile, as the signature is very similar at early and late stages. The other surprising result for me was that a very simple rule could recapitulate very well the topological pattern we observe in the data.
RC I remember when we were investigating division rules and looking at the resulting neighbour distributions we couldn’t find one that matched our data very well. I was in a meeting with Stan and we ended the meeting by saying ‘I wonder what happens if we just split the neighbourhood in half?’. I added the rule to our model and half an hour later checked the results and the distributions matched almost exactly! I recall going back to Stan’s office and both getting quite excited!
Cell age has no effect on topological profile. From Figure 3, Carter et al, 2017
And what about on the flipside: any moments of frustration or despair?
YSC It was very challenging to acquire time-lapse data of a slowly growing tissue. I had all the support from the Coen’s lab, especially from Samantha Fox. But, I still tried many times and make all possible mistakes before acquiring good data sets.
RC As above! We got to the point where none of the division rules we tried were matching our data, as a last attempt we tried the equal split rule, and it worked!
The data best fits the “equal split” rule. From Figure 5, Carter et al, 2017
What are you career plans following this work?
YSC In my postdoctoral work, I am looking at a related aspect of topology: the case when cells exchange neighbours in a polarised manner.
RC I’m currently in another post-doc position at the Sainsbury Laboratory, Cambridge University. Here I’m doing research into modelling the mechanics of morphogenesis in the group of Prof. Henrik Jönsson. I hope to be applying for fellowships next year and to become a bit more independent in the future.
Verônica and Stan, where do you think this work will take your labs?
AM & VG Other groups, like the one of George Bassel, have found interesting topological hierarchies and control for other plant tissues. We think that the community is now realising that these topologies, which are all unique and can only be explained as being tightly regulated on the topological level, points to some form of functionality. As we are also working with nutrient uptake in roots, we wish to explore how topological connectedness of cells could be related to the transport dynamics. In the light of the leaf, we hope to continue to explore how changes in mitosis could distort these topological patterns, and possibly, how this might influence other patterning, communication, and developmental aspects.
Finally, what do you like to do when you are not in the lab?
AM & VG We both love hiking through forests, mountains, marsh lands etc. We now enjoy these nature walks with our two small children, carrying the smallest one, who is one-year-old, on our back, while our 4-year-old jumps ahead like a mountain goat. They are teaching us to “re-see” the world around us, as well as our human condition – with a renewed wonder and a fresh set of questions – from a whole new perspective.
YSC I like cooking (especially Mexican food) and enjoy walking in the countryside.
RC I really enjoy getting out in to the countryside on my bike, painting and drawing, and live music!
Thank you!
Ross Carter, Yara E. Sánchez-Corrales, Matthew Hartley, Verônica A. Grieneisen, Athanasius F. M. Marée. 2017. Pavement cells and the topology puzzle.Development. Volume 144, Issue 23, p4386-4397.
This is #33 in our interview series. Browse the archive here.
In preparation of our upcoming 70th anniversary to be celebrated at the special Spring Meeting in Warwick (April 15th-18th 2018), the BSDB announces a writing competition for its graduate student and postdoc members. We are curious to hear about your thoughts (in not more than 500 words) on one of the following topics:
The future of Developmental Biology
What Developmental Biology has contributed to society
The experiment/paper in Developmental Biology that most inspired you
Further details of the competition will be formally announced shortly, but we thought it might help if we give you some head-start. The deadline will be Sunday, 4th of March 2018, and the winner is going to be announced at the Spring Meeting. The first prize is a free trip to and attendance of the 77th Annual Society of Developmental Biology meeting in Portland, Oregon, USA.
An NIH R01-funded postdoctoral position is available in the laboratory of Dr. Beth Roman in the Department of Human Genetics at the University of Pittsburgh. We study the interplay between molecular and biomechanical pathways in the control of endothelial cell behavior and vascular development and disease. Our primary focus is on endothelial bone morphogenetic protein (BMP)/ALK1 signaling, which is disrupted in the genetic vascular disorder, hereditary hemorrhagic telangiectasia (HHT). We are looking for a motivated scientist to lead studies exploring the molecular, cellular, and biomechanical basis for BMP ligand requirements in HHT, using zebrafish and primary endothelial cell culture models.
Qualified candidates must have a PhD in a relevant field, a strong publication record, and excellent oral and written English communication skills. Expertise in molecular biology and cell culture is required. Experience with confocal microscopy and the zebrafish model is strongly preferred. Interested candidates should send a brief cover letter summarizing scientific accomplishments and outlining motivation for this position; a full CV; and full contact information for three professional references to Dr. Beth Roman at romanb@pitt.edu.
The University of Pittsburgh is a collaborative, collegial environment for biomedical research. Our laboratory is part of a large, interactive zebrafish community, and we are affiliated with the Heart, Lung, and Blood Vascular Medicine Institute (VMI). The VMI offers numerous opportunities for collaboration and provides access to competitive T32 postdoctoral positions.
Today marks the publication of Development’s special issue celebrating the centenary of D’Arcy Thompson’s ‘On Growth and Form’. Given the continuing influence of the book’s graphical elements – from its captivating transformation diagrams to its drawings of geometrically stunning invertebrate shells – the Development team knew that the cover of the issue had to reflect this visual influence. A standard cover – for instance, a confocal image of an embryo – might not have been suitable. Luckily, as the Development team were discussing possible options, Mukul Tewary, first author of one of the papers featured in the issue, got in touch to let us know about the artistic and graphical talents of a graduate student in the Zandstra lab, Jen Ma. A look at some of her work led the Development team to commission her to design the cover for us, and in another piece of luck, Jen was able to involve Matthew Spremulli, a researcher with a background in architecture, in the project.
We caught up with Jen and Matthew to hear about the cover’s conception, design and completion.
Jen Ma and Matthew Spremulli
Jen, can you tell us a little about your scientific background?
JM I am currently doing my PhD in Dr. Peter Zandstra’s Stem Cell Bioengineering Lab at the University of Toronto. It has been an eye-opening experience being in a such a diverse group, where our interests span from cell fate acquisition to cell competition, with expertise in advanced techniques like computational modeling and micropatterning (both employed in the study by my colleagues Tewary et al. published in this special issue). My research focuses on developing a platform for rapid enumeration of rare hematopoietic stem cells based on mRNA signatures.
Have you always been artistically inclined?
JM Come to think of it, everyone in my immediate family are involved in the STEM fields professionally, but also deeply immersed in the arts! Although I never had any formal training, I have always enjoyed drawing and painting, photography, and graphic design. About a year ago, I picked up lettering and illustration as a hobby. Having a creative outlet helps me balance the busy grad school life and has brought many interesting opportunities. I share my work on social media (Instagram: @ItsLikePudding) and volunteer my skills whenever my friends allow me, which have led to many fruitful collaborations and commissions (including this cover illustration). I’m especially interested in combining my passion in science and art for science communication and outreach.
Some of Jen’s creative output. See her Instagram page for more
Matthew – what’s your background, and how did you come to be involved in the project?
MS I am an Architect by training having completed a Masters of Architecture (focusing on digital media, fabrication, and landscape) and a Hon.B.A. (specializing in architecture and fine-art history). However, I decided to professionally explore experimental/research activities rather than traditional licensed practice since graduating, in other words I have not constructed a building. Instead I have worked with various groups on projects that have required expertise in research, design and digital media. I was invited into this project by Jen who upon mentioning the special issue of “On Growth and Form” clearly peaked my interest since the book is so influential to my disciplinary training.
So you were familiar with the book before? And how influential is it in architecture?
MS Yes, I had heard of the book and actually own a copy (the 1971 abridged edition by John Tyler Bonner)! The publication (all editions) is very influential to the discipline, specifically to designers who are interested in structure, form-finding and form-making. There are a wide range of historically notable designers who were directly inspired by the text and illustrations such as Frei Otto and Buckminster Fuller. In the case of Buckminster Fuller you can look at the Montreal Biosphère (originally built as the American Pavilion of the World Expo ‘67 in Montreal) to see some resemblance to the Radiolarian Skeletons or Hexagonal Skeletons from Thompson. On a more broad level, within the last decade architecture (and many of the allied design disciplines) has been heavily influenced by what we informally call ‘parametric’ and ‘generative’ design. These notions imply that a designer can set-up underlying mathematical relationships between geometric features of an object. The resultant forms of these relationships – which are usually managed through software applications such as Rhinoceros-Grasshopper can resemble some of the diagrams and illustrations from Thompson’s book.
What about your experience of the book, Jen?
JM I was briefly introduced to On Growth and Form at a lab meeting a few years back, and borrowed the book from Matthew when I found out that he has a copy. I’m embarrassed to admit that it has been sitting at my desk and I never had a chance to study and appreciate the influence of the book until now. It was very interesting to learn that many familiar concepts like surface-area-to-volume ratio and the logarithmic spiral are studied in this classic, and that its influence is seen in areas outside of developmental biology such as architecture. I actually visited the Montreal Biosphère earlier this month when I was in town for a conference based on Matthew’s recommendation!
Once Development had approached you about the project, what were your initial thoughts?
JM & MS Inspired by On Growth and Form, we wanted to implement concepts of rule-based form development to create the cover illustration. We also wanted to highlight the important role of computational modelling in advancing our understanding of complex physical form in the field of developmental biology. Graphic-wise, we thought it was important to have a strong reference to the iconic and beautifully hand drawn illustrations from the book. The idea was to take one of the specific images as a departure point to develop our own interpretative graphics that were going to be clearly ‘contemporary’ and unapologetically ‘digital’. There was going to be a fusion between these two ideas – historical and projective – we just did not know how this would specifically turn out.
We wanted to highlight the important role of computational modelling in advancing our understanding of complex physical form in the field of developmental biology
I’m interested how the process worked exactly?
JM & MS A key to our ‘start’ was selecting one of the illustrations from the book that we wanted to work with. We debated between a few in particular – the Nautilus Shell, the Grid-Warped Fish, and the Radiolarian Skeletons. Ultimately we settled on the Nautilus Shell because we felt it was the most iconic and easily identified as being from the book. We then started sketching some layout options to integrate the biologically illustrated and digitally invented shells. The layouts provided us with a framework to move forward in our digital studies and, on a practical level, an estimation of how many digital forms we were going to make.
To digitally create our own shells, we sampled a number of scripts in Grasshopper, a visual programming plug-in for the Rhinoceros 3D design software, from various authors exploring spirals and shell type forms (big shout-out to the Grasshopper Forum and it’s amazing group of contributors and members). Ultimately, we cobbled together our own little Frankenstein-script, which could generate 3D structures that feature a logarithmic spiral and inner/outer profiles based on a few parameters and curves.
This allowed us to design an array of new shapes by experimenting with the parameters systematically. Each of the digital shells was then rendered to generate 2D images from the 3D models.We experimented with a number of rendering treatments which we then composited together in Photoshop to achieve the final shell images. This includes the mesh lines – the distinctive blue graphics you see on the cover – that visualize the framework the 3D forms were constructed based on.
The composited shells were then arranged to emulate the sketch layouts we made earlier.
And how did you settle on the final image?
JM & MS The layout was chosen for its incorporation of the concept of size and clean design. The shells on the left, while starkly different in terms of form and complexity, were created by tweaking just one parameter of the model. The shells on the right, on the other hand, share the same spiral structure but different profiles to create various forms. This demonstrates the power of the computational model to easily generate new structures. As for the renders, we implemented a textured finish and shading that’s different from stereotypical computer graphics to give the final image a unique look.
And ultimately, this combination received the most votes from the Development editors so the winner it is :)
Has the experience taught anything you about each other’s fields and the ways they work?
MS I think the experience taught me that scientists/engineers are very rigorous with their process … much more rigorous than designers. I was deeply influenced by the way in which Jen wanted to conduct the creation of the shells and their categorization. It also taught me to be a lot more clear and careful with my language and descriptions of design processes. To be honest I am still recovering from what we call in our industry “archi-speak”, which constitutes a number of words and phrases that designers use amongst each other, but which I realized are difficult for others to understand.
JM While there are some differences due to our backgrounds and personalities, we also share many commonalities in our approaches, which made the process quite smooth and fun! I think Matthew and I both appreciate designs that are aesthetically pleasing, but share the sentiment that developing a concept and representing it through compelling visuals is a more exciting challenge. We are both very open to experimenting with different media and methods, and enjoy the process of evolving our ideas through iterations. Implementing these approaches in the context of 3D modelling is outside of my expertise (my experience is limited to creating simple microfluidic devices and modelling a pineapple once) but well within Matthew’s, and it’s a pleasure to watch him work his magic. Learning parametric design and visual scripting from Matthew and seeing firsthand how he set up the digital framework also means I got a private lesson from one of the best teachers on the subject!
Where next for you both?
MS Over the years I have been moving more into the realm of digital media and research, and in fact I have just recently joined Autodesk (technology and software company). I will be working within one of their Research groups where I will have the opportunity to further explore my interests. I will also continue teaching digital media within the department of Landscape Architecture at the University of Toronto.
JM I’m working on wrapping up my thesis, and exploring opportunities in both research and science communication/illustration. I have also taken an interest in data visualization more recently, which may also allow me to combine my interest in graphics, communication, and science.
What explains the lasting legacy of D’Arcy Thompson’s ‘On Growth and Form’? A century on from the publication of the first issue, we reached out to authors from Development’s special issue celebrating the centenary and asked what the book meant to them.
A rare first edition of On Growth and Form held in the D’Arcy Thompson Zoology Museum in the University of Dundee. Find out more about the museum in our interview with its curator Matthew Jarron here.
Natural History Museum (London) &Imperial College London
“There are ideas which make their impact and give way to new and better ideas. They have their day and disappear into history. However, there also ideas that have a much more profound and enduring effect and continue to influence the way we think for many yeas, sometimes for generations. “On Growth and Form” by D’Arcy Thompson is a book full of such brilliant everlasting ideas (and so eloquently written). I remember reading it for the first time as a half-fledged undergraduate awed by the notion that living organisms and their parts do not have “random” shapes but could be related to each other in precise mathematical terms. What could be a more powerful and visual evidence for evolution than a gradual transformation of a skull of a primitive horse into a modern one via a series of shapes of the intermediate species? This book asked many important questions but gave few concrete answers, instead outlining ways of addressing problems. My own current research is very much driven by the quest to explain Thompson’s “laws of growth” and their role in evolution while using his “theory of transformations” in the form of modern geometric morphometrics. For example, the origin and subsequent diversification of the uniquely-shaped skull of modern birds can be better understood by comparing avian skull shapes with each other and with their reptilian ancestors – all within the same “morphospace”, the concept invented by D’Arcy Thompson. Thus, his ideas in many ways are still surprisingly relevant and stimulating today as ever before. I hope that the 100th anniversary will bring this book more attention from the new generation of scientists curious about biological shapes.”
“I spent several summers in the early 1980’s undertaking my doctoral research on early sea urchin development at the Marine Biological Laboratory (MBL) in Woods Hole. I remember fondly the diverse array of sea life that John Valois and his crew in the supply department would collect for seasonal investigators over the course of each summer. Their labors afforded opportunities to observe and handle not only adult marine animals – many of which I had never seen alive before – but also, on occasion, their eggs and embryos. It was against this backdrop that I developed a career-long fascination with the problem of morphogenesis, and first became acquainted with D’Arcy Thompson’s “On Growth and Form”. Back then, there was a small bookstore on Water Street that was unusual for carrying not only the light summer reading demanded by island-bound tourists, but also a selection of titles reflective of the greater MBL and Woods Hole Oceanographic Institute scientific communities. One August, as the exodus of summer investigators, students and tourists began to transform Woods Hole back to its less frenetic state, I purchased a paperback edition of Thompson’s book, which I began reading on the beach and would continue back at my home institution that Fall. Thompson’s perspective inspired me to appreciate the problem of development as more than an unfolding of genetically-encoded programs but one that is also subject to physical rules and open to mathematical description. In many ways, I feel that my research and that of others interested in morphogenesis has moved inexorably toward a view of embryos as biomechanical “machines”. Thompson likely could not have imagined the modern tools and approaches now being developed to measure, analyze and model forces generated by cells and tissues but it is safe to say he would have approved of where the field has headed since the publication of his influential work.”
Université Denis Diderot, Paris and IGBMC, Strasbourg
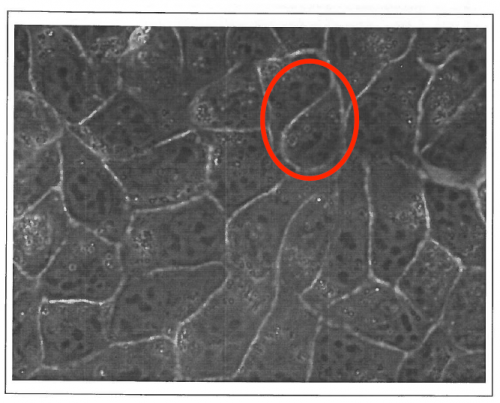
“DR is an experimentalist biophysicist, working on acto-myosin as well as cell adhesion and motility in vitro; FG is at the interface between tissues and foams, and between theory and experiments. We have been knowing each other 20 years ago, while we were in the same lab in Grenoble, and our first discussions in 1999 made already clear that MDCK cells monolayers cellular outlines were not following rules of shapes for bubbles (see our original MDCK image below).
Tissues as foams ? The original 1999 image with MDCK epithelial cells observed by phase contrast microscopy which triggered the first discussions between the authors. In the highlighted zone, these cellular outlines would not be possible in foams.
We both then moved to different scientific sites, and we started to interact again in 2010 around a new Master program launched by DR in Strasbourg University, where we gave interdisciplinary lectures to the same students. DR suggested to share classics pages with students on considering living matter as a matter, such as Thompson book proposes, with modern echoes in developmental biology. FG was reluctant in considering these pages as actually seminal: he rather considered that they had been causing a waste of time over the century, by triggering inadequate searches for simplicity and perfection. These distinct appreciations of ‘On Growth and Form’ nurtured between us an animated debate. We decided to focus on two chapters which were central for us, i.e. on shape and packing of cells in tissues. Our disagreement was so strong that it was difficult to imagine that we would agree on a common flow; initially, we had no agenda whatsoever to wrap up this discussion in an article one day. We turned this debate into a scientific approach, by carefully listing specific arguments and counter-arguments.
And then, we both moved from our original respective postures. DR recognized that Thompson had been merging phenomena under the same umbrella, with often analogy as a guideline and limited care about physical mechanisms. FG acknowledged the influence of Thompson in general and even on his own work for several aspects, and particularly the illuminating ability of Thompson to act as honest experimentalist representing tissues through their essential read-outs – cell contours, representing them as they actually are. We had just decided to write an article when, by coincidence, Development launched a call for its special issue.
A long way has been done since our very first discussions, and we view a posteriori our debates as one of the most productive discussions we had in recent years, with no other ambition than understanding the text and its scope a century after its first publication. And we can only encourage others to perform the same patient task for other chapters of this book or other classical texts in Biology.”
“The field of development has reinvented itself several times over the past decades, building from grafting/morphogen-centered experiments to molecular genetics, cell biology and now including more quantitative and predictive aspects, through biophysics and computational modeling. With the rise of quantitative approaches, the current era is very much in line with D’Arcy Thompson persistent words:
“Dreams apart, numerical precision is the very soul of science, and its attainment affords the best, perhaps, the only criterion of the truth of theories and the correctness of experiments”
D’Arcy Thompson certainly was a (maybe too) strong advocate of the role of physical forces in development, but as we are collecting more and more evidence of a widespread role of mechanical signals in morphogenesis and cell behavior, his take on development also offers a source of inspiration for the young generation of interdisciplinarians in development.
One key quote that was influential to me is a simple one:
“The form of an object is a “diagram of forces””
The simplicity of these words has many implications. It underlines the fact that changing shape is not only a geometrical problem: it also involves a structural change, and thus, that changing patterns of forces must be managed by cells. Not as immediately obvious, this statement also implies that a living object will usually try to resist such forces. There are now many examples of cells displaying such resistance strategies, from changes in cell growth rate, to cell polarity or extracellular matrix stiffening. This also is true at tissue and organ scales, the most obvious example being Wolff’s law, which is mentioned in D’Arcy’s book, where bones build their intricate trabeculae network along maximal mechanical stress lines. We, and others, found similar responses in plants, with cells and tissues reinforcing their structure in the direction of maximal stress too, making this statement from physics, a true biological theorem. D’Arcy Thompson was a supporter of the organismal theory of development, and this quote is also consistent with that option, diagrams of forces spreading, and arguably, synchronizing, populations of cells. While this leaves out many subtleties, it remains a very powerful reminder of the fact that cells manage communication, nutrition, reproduction, etc. and while doing all this, also need to maintain their own structural integrity, which calls for synergies between biochemical and mechanical signalling.
One last take home message from D’Arcy Thompson’s book is his didactic choice: the use of many analogies makes his book very accessible to a wide audience, truly bridging biology, mathematics and physics. This is a remarkable piece of work, and despite several questionable interpretations 100 years down the road, “On growth and form” remains a landmark in developmental biology, and very much worth a read today, for biologists, mathematicians and physicists alike.”
SOKENDAI & National Institute of Genetics, Mishima, Japan
AK “When I was a post-doc, I first read “On Growth and Form” in the Japanese version translated by Dr. Tomomichi Yanagida et al. and published in 1973. In the translators’ preface, Dr. Yanagida wrote the following: “Initially, we were not sure whether the book is worth translating, as the book is about 70-years old and the biology had been progressed dramatically since then.” For me, having read many textbooks on molecular biology, reading this book was an eye-opening experience. Explaining features in sizes and shapes of various organisms at the cellular level (or beyond), using mathematics and physics was an area I wished to pursue as a scientist. Many topics of study in this 100-year-old book are currently pursued by many quantitative biologists. In fact, our manuscript in this special issue of Development was inspired from the chapter “The Forms of Tissues, or Cell-aggregates.” I hope that our manuscript has provided a novel insight into the topic covered in this chapter. Dr. Yanagida concluded the preface as follows: “Nowadays, there is a flood of biology textbooks, but this book is one of a few books that is truly inspirational”; I completely agree with this statement.”
KY “I learned about this book for the first time when I was a graduate student. At that time, I was skeptical of pursuing research in molecular biology because I found it difficult to believe that the essence of the organisms I wished to work on could be revealed by simply focusing on the function of a single gene. The book looked at the question from a different point of view. Therefore, I found the philosophy of the book attractive. Indeed, for many people who like to watch organisms including myself, one of the interesting aspects of organisms is the various beautiful structures they create. The premise of the book that the shape of different structures, such as a cluster of cells, the skeleton of planktons and sponges in the ocean, and the body shape of jellyfish, might be explained by surface properties of liquids, is indeed enthralling. The association of surface tension with the broad range of biological shapes, from cell shapes (micron-size) to animal body shapes (meter-size), intrigues me. Now, I deeply respect D’Arcy Thompson’s abundant knowledge and speculations regarding organismal morphology based on physical and mathematical viewpoints, and I shall remember him every time I observe dynamic behaviors of organisms under a microscope.”
Keck School of Medicine, University of Southern California & Children’s Hospital Los Angeles
“I first ‘read’ Thompson’s ‘On Growth and Form’ as an undergrad. I found it rather daunting in a kind of War and Peace thick science text way. I tepidly thumbed through the pages, mostly stopping at interesting figures, especially the ones about spirals in nature and the accompanying mathematics. Soon I was up the N. California coast abalone diving with friends. We were there to collect and enjoy the yummy gastropods with local spirits. We always enjoyed the iridescent nacre that lines the inner shell. However, on this trip, my mind drifted back to Thompson’s figures of Haliotis, and its precise patterns of growth. My perspectives had forever changed, even on road trips with friends, to see patterns and morphometrics everywhere.”
“I was phD student the first time I heard from on Growth and Form. Thomas Lecuit gave a very nice seminar in our institute. I remember him showing the famous drawings describing how environmental forces may affect the overall shape of an adult fish (it should correspond to the Figure 517 to 524). As a PhD in developmental biology, it had a strong impact on me. At the time, we were struggling with in situ hybridization and our understanding of how shape is emerging out of these expression profiles was clearly not satisfactory. I cannot say if it was the drawings themselves or if the field had already shifted, but the idea of forces affecting shape led to lots of discussion in the institute. It definitely influenced my post doctoral work and current research.”
“Throughout the book, which I read in a French translation of the abridged John Tyler Bonner version, D’Arcy Thompson tried to use physics and mathematics to explain the shape of life forms. This was an audacious project, since the living world remained a big mystery at that time. D’Arcy Thompson foresaw the great utility of physics and mathematics in studying growth and form, while admitted that they could not explain all the living phenomena. He stressed the importance of material and mechanical nature of life, considering that a living form was nothing else than “a force diagram”. D’Arcy Thompson illustrated the beauty of a wide variety of species through mathematical description. He pointed out the similarities between seemingly unrelated things, linking the natural phenomena with physical laws. Modern research on morphogenesis has proved that many of these intuitions were correct.
As a researcher working on a particular morphogenetic problem (elongation of C. elegans embryo), I also look at the biological phenomenon from a physical point of view, in terms of “materials”, “forces” and “force distribution and evolution”. Like D’Arcy Thompson, I also try constantly to relate the morphogenetic mechanisms with physical laws and physical phenomena that we see in daily life. Some paragraphs of the book are directly related to my work, for example the one stating that the formation of shapes different from a perfect sphere may results from either a difference in internal pressure (inside the object) or the anisotropy of the surface envelope. The relevance to my particular case on the anisotropic deformation C. elegans embryos shows that D’Arcy Thompson’s observations can still inspire future generations of scientists. Although many of the theories in “On Growth and Form” were simplistic and unproven, the observations and reasoning are a good incentive to further investigate the nature of the phenomenon. The book deserves to be read broadly, not only for its pioneer ideas on morphogenesis but also for its enjoyable elegant writing style.”
In late September I boarded a tiny propeller plane to Dundee to meet Matthew Jarron, who curates the University of Dundee’s D’Arcy Thompson Zoology Museum. We had seen Matthew speak about On Growth and Form at a Royal Society event over the summer, and realised he would be a perfect person to interview to give the work some context in our special issue celebrating its centenary (published today!). On the taxi from the airport, through the fog I could just about make the Tay Rail Bridge, which was rebuilt in 1887 after collapsing in a storm in 1879. D’Arcy Thompson’s book contains a section comparing dinosaur skeletons to railway bridges – how does the form of an object reflect the mechanical strains placed on it? – so I could almost imagine a sleeping dinosaur stretching south towards St. Andrews. Matthew was a great source of insight into the book and the man behind it; I’d encourage anyone who visits Dundee to go see the museum and its wonderful collection of items, many of which inspired Thompson’s ideas.
Matthew Jarron (left) and a portrait of D’Arcy Thompson by David Shanks Ewart (right; courtesy of the University of Dundee Museum Services).
D’Arcy Thompson was born in 1860, trained in Edinburgh and Cambridge, and held positions in Dundee and St Andrews, where he worked until his death in 1948. On Growth and Form, his classic work on the mathematical patterns and physical rules underlying biological forms, was first published in 1917. To learn more about the book’s context, we met Matthew Jarron, Curator of Museum Services at the University of Dundee, in the University’s D’Arcy Thompson Zoology Museum. Surrounded by specimens, many of which were collected by Thompson himself, we discussed the legacy of On Growth and Form and the life of the man behind it.
Matthew, can you tell me how you first came to encounter D’Arcy Thompson’s work?
Before I came to Dundee, I was a curator of a local history museum in St Andrews, which had been gifted a medicine chest from his days as a student in Edinburgh by the last surviving of his three daughters. This wonderful chest was my first encounter with D’Arcy, and as I gradually read more stories about him, I realised what a fascinating character he was. When I came to Dundee, I discovered we had what was left of his zoology collection. This was once one of the largest collections of zoological specimens in the country but had become dispersed in various displays and was not really accessible to the public. It wasn’t until the department of Life Sciences moved building in 2007 that we were able to create a proper version of the museum, have it open to the public and start telling people more about D’Arcy’s work. The more I read about him, the more I realised how incredible his influence has been in so many different fields in biology and beyond. In the museum, we’ve been very keen to use his collection in as many different ways as we can, to get people interested in his life and work. The centenary of his book has provided a great opportunity to do that, and you can find out about the various celebratory events here in Dundee as well as globally on the anniversary website (www.ongrowthandform.org/).
How did the idea for On Growth and Form come about?
From very early childhood, D’Arcy was fascinated by the natural world, partly because a number of his family were vets, but also because his father was a great classical scholar and had introduced him to classical biologists like Aristotle. Originally, he went to Edinburgh University to study medicine, but quite quickly realised that he wanted to specialise in biology, and transferred to Cambridge, which was really the only place where you could focus on the natural sciences at the time. He came to the University of Dundee (known then as University College Dundee, which had opened in 1883) at a very early age – he was only 24 when he was appointed to the first chair of biology in 1884. It was quite a small university, but they assembled a really extraordinary and dynamic group who were able to think in interdisciplinary ways. So, from very early on I think he had that broader interest, paired with his classical training and the associated idea that naturalists could look at broader themes.
As for On Growth and Form, we know that in 1889 he wrote a letter to one of his students saying that he had taken to mathematics. He had been looking at foraminifera, single-celled organisms that grow these extraordinary ‘tests’ (external shells) that often form geometric shapes, and he realised that there were very specific patterns that kept recurring. That was probably when he started to think that not everything in biology could be explained by Darwin’s ideas of evolution, which by that point had become quite widely accepted. These foraminifera, which presumably had similar evolutionary pressures, adopted quite diverse geometric forms; for D’Arcy this suggested that there was clearly something more going on here. He began to think about the physical forces acting on these organisms and realised that they formed specific mathematical patterns during their development.
Thompson started to think that not everything in biology could be explained by Darwin’s ideas of evolution
At the time he realised that these ideas would be quite controversial, and also that no one would really have much interest in applying them. He wasn’t completely keeping it to himself, but it wasn’t until 1908 that he published anything at all – a paper in Nature on the shape of eggs. Then, in 1915, he published a fairly substantial paper in the Royal Society of Edinburgh’s Transactions called ‘Mathematics and Morphology’, which is essentially what forms the final and most celebrated chapter of On Growth and Form. In 1911, he had been asked by Cambridge University Press to write a small book on the biology of growth and form. They had a series of popular science works that cost something like a shilling and were perhaps a hundred pages long, but as he started writing, it just got bigger and bigger, taking many years to write, and when he finally sent the text back to Cambridge they must have been somewhat horrified to find how enormous it had become! They did, however, agree to publish it separately.
The shell of a nautilus, one of the species that D’Arcy Thompson included in his section on spirals in nature, from the D’Arcy Thompson Zoology Museum (University of Dundee, UK).
And what does the book actually contain?
D’Arcy introduces the book with his case for looking at biology from a mathematical standpoint, to understand form from the point of view of physics and of mathematical laws, even though this was something that naturalists didn’t particularly want to do. The next chapter is concerned with size, and he makes the point that gravitational forces are much more important at larger scales, while surface forces, and in particular surface tension, play a much more important role at smaller scales. He starts with the smallest organisms, individual cells and their internal structures, and gradually works his way up, for instance exploring how the formation of clusters of cells is analogous to how soap bubbles come together. Throughout the book, and particularly here, he takes an organic form and compares it with an inorganic one; where he sees the same patterns, he reasons that the same physical forces are causing these shapes to appear. He goes on to look at individual parts of organisms – for instance the shells of foraminifera and nautilus, or antlers, horns and tusks – and explores how growth rates might cause their different curves and spirals. Then he looks at larger organisms, and famously compares the structure of quadruped skeletons with the structure of bridges. There is a chapter (which is missed out of the abridged edition) on plants and their phyllotaxis in relation to the Fibonacci sequence, and then in the final chapter he presents his theory of transformations. When dealing with large entire organisms, he admits he clearly can’t explain all their differences through mathematics, but he can take two related organisms and see how one could mathematically have changed into the other. He looks at the organism as a system, and explores how that whole system will transform; the famous diagrams are his attempt to explain that. He ends the book by saying that what he has described in the book is ‘a field which few have entered and no man has explored’.
A rare first edition of On Growth and Form, held in the D’Arcy Thompson Zoology Museum (University of Dundee, UK).
How was the book received?
When the book came out it was very widely praised. Everyone was hugely impressed by the vastness of his learning; one of the most extraordinary things about the work is how he brings together examples from recent research with examples from ancient history, that he views the biology of Aristotle as equally relevant to the biology of the time. There were reviews of the book not only in biology but also in engineering journals, as well as Country Life of all places, and I think this reflects a recognition that the ideas in the book could have a much wider application. But it’s notable that many of the reviewers were basically saying how well written it was, rather than how it was going to transform biology. While an enormous amount of learning had gone into the work, very few people could actually grasp how to take it forward. A key issue was that many biologists didn’t really know much mathematics; similarly, he struggled to get many mathematicians interested in the work. But there were some key people who picked up on the book and took forward the idea of mathematical biology, and with whom D’Arcy was in regular correspondence. Interestingly, these people were often working in different fields and using completely different kinds of mathematics from D’Arcy’s; for instance, early ecologists interested in using mathematical models to look at populations.
The D’Arcy Thompson Zoology Museum (University of Dundee, UK).
More and more people appear to be turning back to his work and finding useful things in it today
The first edition had sold out in 1923 – as far as we know it was limited to 500 copies, one of which is displayed here in the museum – but it was 20 years before he got round to writing his second edition. It’s no coincidence that both editions were published during wartime – it was only then that he had time to write, as so many of his students were away at the front and his administrative role lightened. The second edition includes more illustrations and examples – for instance, a section on animal coat markings that was a key influence on Alan Turing – but it does not actually contain that many new areas of research. In particular, he was criticised for ignoring all the developments in genetics since the first edition was published. It wasn’t that he didn’t know anything about genetics – he was completely up to speed with everything that was going on – rather, it was that he couldn’t see how he could incorporate the work into his thesis, and so he just ignored it. While he could get away with this in the first edition, by the time of the second edition it was less easy, and that rather doomed his work for a lot of biologists. Conversely, while the second edition was problematic for some biologists, it got very quickly picked up by artists, engineers, architects, geographers and anthropologists. So from the 1950s there was a renewed interest that slowly picked up, until recently when it seems to have taken off exponentially. It’s quite amazing that more and more people appear to be turning back to his work and finding useful things in it today.
The book argues that one can look at biology through a mathematical lens – what type of mathematics did Thompson employ?
The mathematics he knew was pretty much all classical: geometry and algebra. He certainly didn’t have any kind of detailed knowledge of contemporary mathematics, and indeed he acknowledges that himself. I think it was a real problem for him, but he just didn’t have the time to get into detailed mathematics of the kind that later biomathematicians did. He probably did not have the inclination either: I don’t think he ever felt that mathematical biology was the topic he’d spend the rest of his life on, as his main interest was in fisheries. Indeed most of his work was taken up with international diplomacy about fishing quotas and that sort of thing; he was well known as a great diplomatist in helping to resolve disputes while countries were dividing up the sea for different fishing rights. With On Growth and Form, he felt that he’d opened a gate into this new field, and was happy for other people to go in and explore it.
Richard Dawkins once said it was a great shame that the computer wasn’t invented in D’Arcy’s lifetime because his work cries out for it, and perhaps this is one of the reasons why so few people at the time actually picked up on what he was doing. It took more sophisticated mathematicians like Alan Turing and the development of computers to allow you to test these theories and build models.
What do you think are the book’s key contributions?
Largely, I think it’s the general idea that you can apply mathematics to biology. This has been so influential in biology as a whole, and particularly now in developmental biology, where its specific influence might not have been that great at the time. His holistic approach was influential for the whole organicist tradition of biology, and particularly for people like Conrad Hal Waddington, who was enormously influenced by the book and whose ideas about epigenetics have become so crucial to development. The idea of taking this vast and complex world of nature and reducing it down to fundamental rules is also obviously a key part of systems theory, and was hugely influential in its development, not to mention cybernetics and the start of computing, artificial intelligence and so on. These are all linked to D’Arcy’s work, and his ideas are still influential in art, architecture, geography and anthropology. Actually there are whole new areas of science that trace an influence back to him – things like nanochemistry; one of the field’s pioneers, Geoffrey Ozin, credits D’Arcy as an inspiration. It’s clearly something that people in a wide range of fields are continually coming back to, even if they are just taking general ideas rather than specifics.
Model of a radiolarian skeleton (Actinomma inerme) from the D’Arcy Thompson Zoology Museum (University of Dundee, UK); D’Arcy ordered a set of these from the Czech model-maker and natural history dealer Vaclav Fric.
Considering Stephen J. Gould’s opinion that biologists regarded On Growth and Form as ‘an unusable masterpiece doomed by excessive length and difficulty of application’, why should the modern reader pick it up?
It certainly is the case that very few people have read the book from cover to cover, in order, but in a sense that doesn’t really matter. For me there are two reasons to pick it up: one is that even if you’re never going to read through the mathematical bits, or the stretches of Latin, Greek and German that he doesn’t bother to translate for you, there are lots of very beautiful, poetic passages that still read very well, and which are just inspiring; the other is that it has so many amazing illustrations that are also just as inspirational. Frankly, if all you do is pick it up, flick through it and look at the pictures, that’s great – you’ll still be inspired by the mathematical beauty of nature, as D’Arcy wants you to be. There’s still lots there for the modern reader.
And what about D’Arcy the man – you’ve read much of his correspondence and what people wrote about him. Do you have any feel for his personality?
He had a great personality, and was a larger than life character in every sense. For one thing he was a large, tall man, standing six foot three, described as a veritable lion due to his beard. His students loved him as an eccentric teacher who would use bizarre props to help illustrate his points. There are many anecdotes about how learned he was, how in tutorials he would translate from Medieval Italian, that kind of thing. I think he had a great sense of humour, which certainly comes across in letters to his friends. He corresponded with so many different people, and though he was this great diplomatist, always very polite to people, when he was speaking to his closest friends he could be incredibly and hilariously rude about others! He was also a great populariser of science – he loved giving public lectures, and talking to children and showing them round his museum. Famously, in his old age he had a pet parrot that perched on his shoulder as he wandered the streets of St Andrews. So by all accounts he was a great character.
In a sense he was the last of an era. In the 19th century, it was quite common to have great erudite polymaths, but in the 20th century less so. I think he very much felt of himself as a man out of time, and this was perhaps one of the reasons he loved classics so much – he looked back on great figures like Aristotle who could take a broad look at life. The breadth of his knowledge was extraordinary, and he was constantly championing ideas of interdisciplinarity and emphasising how important it is to look beyond your own field. Holistic overviews like the one provided by D’Arcy are, I think, still important, and it would be a shame if we totally lost them.
Do you think there are any misconceptions about D’Arcy Thompson and his work?
Well, one is that he is somehow anti-evolution. It’s not that he thinks that evolution is wrong or doesn’t exist, it’s just that he’s saying that Darwinian evolution can’t explain everything. The transformation diagrams were, I guess, the part that most obviously appeared to contradict Darwinian evolution – whereas Darwin emphasised slow, gradual change, with each particular part responding to natural selection, D’Arcy says that that can’t always be the case, and there must have been times when there was more of a sudden transformation from one kind to another. But even if you argue with this point, his work emphasises that, however something evolves, it will evolve according to certain mathematical patterns.
There’s also a general misconception – and of course I’d say this being here in Dundee – that he was some kind of maverick loner in a remote backwater writing this book without any support. This simply isn’t true – he drew hugely on his fellow professors here in Dundee; for instance, the engineering professor Thomas Claxton Fidler helped him with his ideas of dinosaurs and bridges, and the physics professor William Peddie was enormously valuable in helping to shape the entire book.
If Thompson were alive today, what would you ask him?
From a purely personal point of view, working here in the museum, I would ask him where he got each one of these specimens – he never catalogued his museum, it was just all in his head! Occasionally, we’ll find a letter where he’ll describe some gorilla he’d just acquired, but for a vast amount of stuff we just have no written record.
The other thing I find mysterious about him was his position on the great debates in biology at the time around ideas of vitalism, mechanism and organicism. It is never entirely clear where D’Arcy sits with regard to them – on the one hand, On Growth and Form seems to be quite a mechanistic book, reducing nature down to rules, but at the same time D’Arcy is very keen to point out that mechanism doesn’t have all the answers, that there are things that you can’t explain in nature. In that sense, he takes elements of vitalism, while a lot of the organicists saw him as a key influence. It probably comes back to his diplomacy – he very deliberately doesn’t come down on one side or the other. So I’d like to sit him down in a room and find out what he really thought about these issues.
Morphogenesis, the study of how forms arise in biology, has attracted scientists for aeons. A century ago, D’Arcy Wentworth Thompson crystallized this question in his opus On Growth and Formusing a series of biological examples and geometric and physical analogies to ask how biological forms arise during development and across evolution. In light of the advances in molecular and cellular biology since then, a succinct modern view of the question states: how do genes encode geometry?
Understanding this fascinating problem requires insight into how shape emerges when molecular information and physical forces are regulated over many different scales in space and time. To address this requires an appreciation of the enormous ‘morphospace’ of potential shapes and sizes that living forms can take up. In parallel, we need to consider the large diversity in the genetic space of potential regulatory interactions that influence form. While the conceptual framework of developmental patterning explains how cells acquire information and how this defines their behaviours, Thompson’s agenda of describing biological processes in mathematical terms is based on understanding how instabilities and patterns in physical systems might be harnessed by evolution. Consequently, the subjects of morphological (phenotypic) and regulatory (genotypic) diversity that are separated by many orders in length scales, have not been sufficiently coupled intellectually.
100 years after the publication of On Growth and Form, we are in a position to better encapsulate phenotypes and genotypes under a unified conceptual and mechanistic framework
Now, 100 years after the publication of On Growth and Form, we are in a position to better encapsulate phenotypes and genotypes under a unified conceptual and mechanistic framework. This entails a search for a potentially low-dimensional phase space for the description and control of shape over developmental and evolutionary time scales. Any parametrisation of the processes at play must have both physical as well as regulatory bases in terms of biomolecular processes that respond to and control these physical parameters. A fundamental challenge therefore is to connect these different scales while deducing the dimensionality of these ‘morpho-genetic’ spaces underlying the development and evolution of shape.
The past two decades have seen an increasing influx of physicists, mathematicians, engineers and computer scientists into the field of developmental biology, who are all attempting to determine the correspondence between the parameters that describe shape and those that define its generation and transformation. There are three major areas in which they have made contributions. First, they have developed mathematical and algorithmic tools for the quantitative description of shape, i.e. morphometrics, directly inspired by the last chapter in Thompson’s book, titled ‘The Theory of Transformations’. This has led to the modern field of pattern theory and statistical shape analysis. Second, they have contributed theoretical and experimental tools to describe and measure the collective biophysical properties, instabilities and patterns of active living matter. Finally, they have pushed forward the efforts to describe morphogenesis using a limited set of relevant physical or mechanical parameters and relate them to biological regulatory processes, initiating a transition towards a predictive developmental biology.
This Special Issue celebrates this synergy by providing insights into the genetic underpinning of embryo and tissue patterning, the biological basis of cell and tissue dynamics, and a physical framework to capture these processes operating across scales. The issue begins with an interview with Matthew Jarron, curator of the D’Arcy Thompson museum in Dundee (Maartens, 2017). This Spotlight article sets the scene – introducing the reader to Thompson’s life and ideas, as well as his legacy.
The Review and Research papers that follow cover a wide spectrum of topics across developmental biology. Coen and colleagues (Coen et al., 2017) directly address the question laid out at the beginning of this Editorial – how genes regulate geometry – with a particular focus on plant tissues. Continuing the theme of plant morphogenesis, three research papers apply mathematical approaches to phyllotactic patterning (Fal et al., 2017), sepal growth (Tsugawa et al., 2017) and cell packing and topology in the leaf (Carter et al., 2017). Graner and Riveline review chapters VII and VIII of Thompson book, ‘The Forms of Tissues, or Cell-aggregates’, providing an overview of the mathematical and physical principles underlying epithelial cell shape and packing in both historical and modern contexts (Graner and Riveline, 2017).
Also focussing on epithelial tissue, a research paper from Dye and colleagues (Dye et al., 2017) provides a quantitative analysis of growth and patterning in Drosophila wing, which is one of the best-studied epithelial tissues, and Irvine and Shraiman (2017) review our understanding of tissue growth from a mathematical and mechanical perspective, using planar shapes such as wings and leaves as examples.
In their Review, Heer and Martin (2017) discuss how tension and contractility influence morphogenesis. Complementing this, but on a very different scale, Felsenthal and Zelzer (2017) review how the developing musculoskeletal system influences its final form and function. Revisiting the last chapter of Thompson’s book, ‘Theory of Transformations’, Abzhanov (2017) argues that to understand both ontogeny (development) and phylogeny (evolution), an appreciation of the intrinsic ‘laws of growth’ is essential to frame our models of adaptation and speciation across evolutionary time. Complementing this, Sharpe (2017) provides an overview of the kinds of computational tools and problems that Thompson might have used and addressed had he been alive today.
The mechanics of developmental processes involves multiple scales, and a number of papers discuss examples of this: Boselli and colleagues consider the role of fluid flows and shear stress in orienting tissue movements (Boselli et al., 2017), Nelson and colleagues look at the effects of pressure on early branching processes (Nelson et al., 2017), Ruiz-Herrero and colleagues provide a general framework for size control of growing tissue cysts (Ruiz-Herrero et al., 2017), Lefevre and colleagues analyse multi-scale branching in the mammalian kidney (Lefevre et al., 2017), and axis elongation in the avian embryo is the focus of the work of Bénazéraf and colleagues (Bénazéraf et al., 2017).
In their Review, Engler and colleagues (Kumar et al., 2017) provide an overview of the latest advances of our understanding of how such forces can regulate stem cell fate. Several papers directly address the role of mechanics in growth, form and fate, focusing on the role of the extracellular matrix (Chlasta et al., 2017; Vuong-Brender et al., 2017), cytoskeletal dynamics and cell-cell contacts (Sonavane et al., 2017), and the mechanical phenotype of cells during reprogramming and differentiation (Urbanska et al., 2017). In addition to some of those studies already mentioned above, problems ranging from gradient establishment in the Drosophilaembryo (Carrell et al., 2017), eggshell shape in Caenorhabditis elegans (Yamamoto and Kimura, 2017), and patterning of stem cell colonies in culture (Tewary et al., 2017) showcase the variety of physical and mathematical approaches that modern development is beginning to embrace.
Problems yield only when appropriate tools can be developed to solve them
Problems yield only when appropriate tools can be developed to solve them. Thompson’s pioneering vision was exemplified in his statement that ‘[The] problems of form are in the first instance mathematical problems, and [the] problems of growth are essentially physical problems; and the morphologist is, ipso facto, a student of physical science’. Today’s developmental biologists have much more sophisticated tools at their disposal, relative to those a century ago, when we had almost no way to measure gene expression patterns, to image at the subcellular level, to understand physical instabilities and patterns in nonlinear systems, or to use computers to help simulate them. Indeed, looking at the papers in this issue, we can see the influence of quantitative experiments coupled with mathematical modelling and simulations everywhere. For example, the mathematical and computational tools deployed range from topology (Carter et al., 2017; Graner et al., 2017), complex analysis (Irvine et al., 2017), reaction-diffusion theory (Carrell et al., 2017), agent-based models (Yamamoto et al., 2017) and graph theory and lineage analysis (Lefevre et al., 2017) among others.
On Growth and Form raised the question of the origin of biological shape in a physical framework. Since then, advances in our understanding of the biochemical basis of the laws of heredity have provided the modern conceptual understanding for how shapes develop anew at each generation, from a single cell – thus surviving the death of an individual through its offspring. As this Special Issue illustrates, we are now beginning to understand how genes encode geometry. As morphology both enables and constrains function, a natural next question is how biology creates functional (and plastic) shape that begins to link morphology to physiology and behaviour. As you mull this question, we would like to thank all the authors and referees of the articles in this Special Issue for their contributions, and we hope you enjoy reading it!
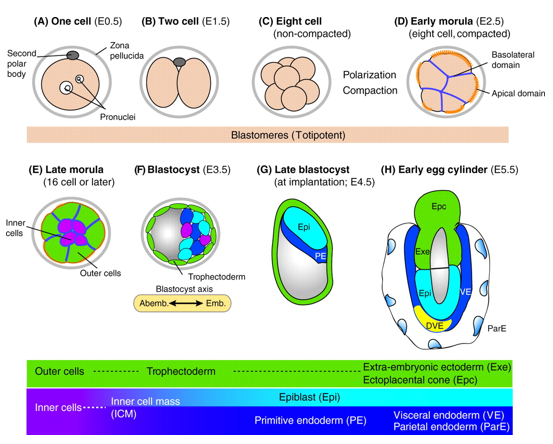
 (3 votes)
(3 votes)



 (3 votes)
(3 votes)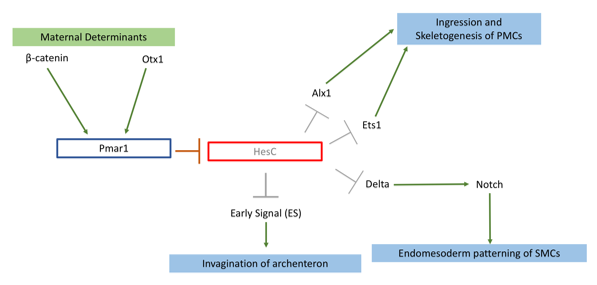
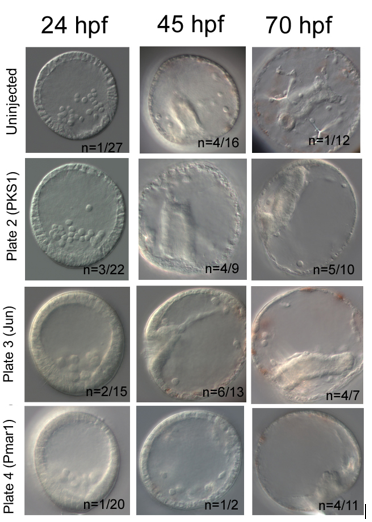
 (No Ratings Yet)
(No Ratings Yet)