As part of our ‘first issues’ series on the Node, Development’s in-house team are researching the authors of articles published in the first issues of Journal of Embryology and Experimental Morphology (JEEM) and its reincarnation, Development.
In this final post, we meet Tom R. Elsdale, a former embryologist at the Medical Research Council Human Genetics Unit in Edinburgh, UK, (Fig. 1) and an author in the first issue of Development, with an interest in understanding one of life’s big philosophical questions: time.
Born in 1929 in Somerset, UK, Tom Elsdale was raised by opera singer Vivian (née Miller) Elsdale and World War I marksman Henry Elsdale. Unusually, his dad also had a second home and family, fathering many children – one of whom accidentally blinded Elsdale in one eye by shooting him with an arrow (Bard, 2014). Despite this unorthodox upbringing, Elsdale would go on to be a remarkable developmental biologist and have fantastic taste in jumpers (Fig. 1).
Fig. 1. Tom Elsdale performing a nuclear transfer experiment in Xenopus. Still from a video posted by Denis Duboule (https://youtu.be/HhGvk5Aqh5k).
Following his undergraduate degree at the University of Reading, UK, Elsdale undertook his PhD studies with the renowned Conrad Waddington (Bard, 2014), a founding Editorial Board member of JEEM and likely one of its early supporters who pitched the journal to The Company of Biologists (Eve, 2025). Waddington published in the second-ever issue of the journal (Waddington and Carter, 1953) and several times thereafter, although Waddington and Elsdale never did publish together.
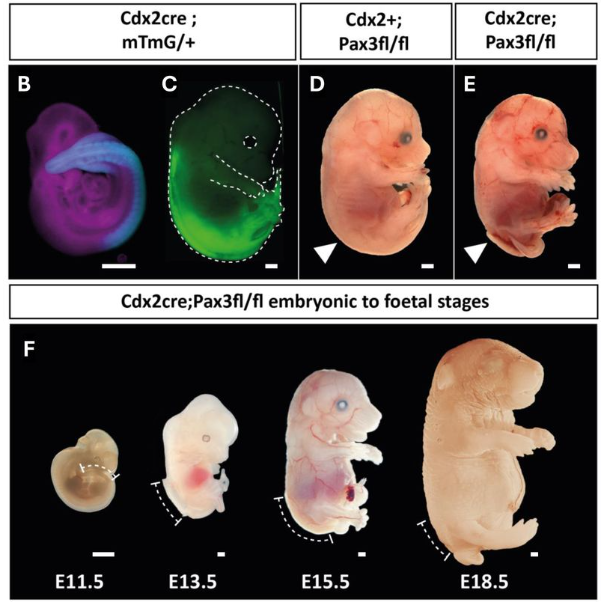
As a postdoc at the University of Oxford, UK, with Russian-born biologist Michail “Michael” Fischberg, Elsdale taught John Gurdon how to perform nuclear transfers in Xenopus laevis, following in the footsteps of Audrey Muggleton-Harris‘s work on amoebae. They adapted their approach from Briggs and King (1952), which involved taking nuclei from embryonic cells and transferring them into enucleated eggs, successfully cloning animals that survived into adulthood (Gurdon et al., 1958). Their technique was later published in JEEM (Elsdale et al. 1960), and you can watch Elsdale performing it as part of the Node’s ‘show and tell’ series (see Fig. 1, 2). Gurdon would use this approach to publish an article in 1962, also in JEEM, where he demonstrated that nuclei from differentiated cells (epithelia) could give rise to new embryos, successfully cloning an adult frog (Gurdon, 1962). It was for this work that Gurdon was awarded the 2012 Nobel Prize for Physiology or Medicine (Gurdon, 2013a,b), together with Shinya Yamanaka and his discovery of reprogramming cells to produce induced pluripotent stem cells (Takahashi and Yamanaka, 2006). Before being awarded the Nobel Prize, Gurdon also served as the Chair of the Board of Directors for The Company of Biologists from 2001 to 2011, demonstrating another indirect link Elsdale had with the Company (Bray et al., 2025).
Fig. 2. Figure from Elsdale et al. (1960) showing the apparatus for nuclear transplantation (A), performing the experiment (B, C) and the resulting cloned embryos (D, E).
Following his work on cloning amphibians, Elsdale started his own lab in the seemingly close-knit community at the Institute of Animal Genetics (IAG) in Edinburgh, UK, where he was colleagues with Ruth Clayton, and links him to The Company of Biologists yet again via the IAG’s founder, Francis Crew (Hankins and Rutledge, 2023; Knight, 2023). Elsdale became interested in optimising in vitro biology, demonstrating that frog embryonic cells could be cultured as aggregates. These studies revealed insights about gastrulation, germ layer formation and differentiation (Jones and Elsdale, 1963). One might also argue that these were prototype ‘xenobots’ – as published 60 years later to much media fanfare (Kriegman et al., 2020). Like the Wolffs, Elsdale was also a pioneer for culture conditions, using collagen rather than glass or plastic to aid fibroblast cell maintenance well before the days of Matrigel™ (Elsdale and Bard, 1972).
By the point of the latter study, Elsdale had moved from the IAG to the MRC Clinical & Population Cytogenetics Unit in Edinburgh, UK, where he then published a series of papers addressing somitogenesis. Returning – once again – to frogs, Elsdale and colleagues frequently used heat shock, which was known to cause developmental phenotypes in rats (Škreb and Frank, 1963), as a method of understanding more about developmental timing and robustness in morphogenesis (Elsdale et al., 1976; Pearson and Elsdale, 1979a,b; Cook and Elsdale, 1980; Elsdale and Davidson, 1983). Elsdale’s first paper on somitogenesis followed the landmark ‘clock and wavefront’ model of segmentation by Cooke and Zeeman, published the previous year, which became textbook in the field (Cooke and Zeeman, 1976). Today, although the model has not been immune from scrutiny (e.g. Murray et al., 2011; Klepstad and Marcon, 2024), it has formed the basis for many significant studies revealing the molecular mechanisms behind somitogenesis in vivo and, more recently, in vitro systems that undergo segmentation-like processes (reviewed in Miao and Pourquié, 2024). Collectively, these studies on the segmentation clock are a wonderful illustration of one form of timekeeping in development.
Before retiring in Spain, Elsdale published his final paper in the first issue of Development, ‘Timekeeping by frog embryos, in normal development and after heat shock’, in which Elsdale and Duncan Davidson explore how a population of embryos within a species develop at the same rate (Elsdale and Davidson, 1987). If you’ve ever seen a clutch of frog eggs developing, such as the Xenopus embryos undergoing cleavage in the video below (Fig. 3), you’ll appreciate that the synchrony between the individuals is extraordinary.
Fig. 3. Video of the synchronous cleavage of Xenopus laevis. From Xenbase.
In their paper, Elsdale and Davidson collected Rana temporaria spawn from a local pond and incubated the embryos at 15°C. By recording the development of three spawns from different locations, they report relatively little variation in developmental rate. Next, they use Elsdale’s favourite manipulation (heat shock by limited exposure to 37°C) to perturb the embryos, being careful to ensure that the extent of the heat shock does not induce developmental phenotypes that occlude accurate embryo staging through counting somites. They reveal that heat shock delays embryonic development, with longer exposures to high temperatures increasing the delay, roughly equivalent to one hour of delayed growth per minute of heat shock. Interestingly, though, the embryos go through a ‘rest and recover’ process: embryos stop developing during the heat shock during a ‘rest phase’ and then continue development at the same rate as controls. In other words, the delay in development caused by heat shock is not due to an overall slowing of the pace of embryogenesis, nor do embryos speed up following heat shock to recover lost time. Elsdale and Davidson speculate that some exceptionally stable ‘relaxation oscillator’ could account for this ‘rest phase’, resulting from the degradation of a factor during heat shock that must be replenished before development can continue; they propose that DNA might be the factor involved. Finally, they pursue an earlier observation that short exposure to heat shock-inducing temperatures protects against subsequent shocks (Pearson and Elsdale, 1979a). They show that embryos exposed to two shocks of 8 minutes each, given two days apart, have less developmental delay than embryos exposed to a single 16-minute shock. In the former condition, embryos have a shorter rest phase.
I think this is a really interesting paper that raises many questions relevant to modern research but, with only six citations, it seems to have largely gone unnoticed. Understanding how embryos ensure a steady developmental pace, perceive and interpret time for the synchronous development of organs and tissues, and modify regulatory mechanisms for heterochronic development in or between species is pertinent today. The field of developmental time has grown in recent years with regular meetings on the topic and is a flourishing area of research, both conceptually and experimentally (e.g. Garcia-Ojalvo and Bulut-Karslioglu, 2023; Ebisuya et al., 2024). The heat shock pathway Elsdale used to induce developmental phenotypes was later coopted for inducible transgenic expression systems in several models, including Xenopus (Wheeler et al., 2000), and temperature-sensitive manipulations remain important tools for studying development that continue to be developed (e.g. Benman et al., 2025). Similarly, the relationship between temperature and development is as relevant now as 40 years ago, particularly in the context of climate change (e.g. Moriyama et al., 2023; Kale et al., 2023, preprint). I think it’s safe to say that, in many ways, Tom Elsdale was ahead of his time.
And thus, this post concludes our ‘first issues’ series in honour of The Company of Biologists’ 100-year anniversary. We hope you’ve enjoyed another look at JEEM/Development’s archives, and maybe we’ve inspired you to take a look yourselves. If you find a hidden treasure, why not tell the community about it here on the Node?
Benman, W., Huang, Z., Iyengar, P., Wilde, D., Mumford, T. R. and Bugaj, L. J. (2025). A temperature-inducible protein module for control of mammalian cell fate. Nat Methods. https://doi.org/10.1038/s41592-024-02572-4
Bray, S. J., Royle, S. J., Shiels, H. A. and St Johnston, D. (2025). The Company of Biologists: celebrating 100 years. Development; 152 (1): dev204567. doi: https://doi.org/10.1242/dev.204567
Briggs, R. and King T. J. (1952). Transplantation of living nuclei from blastula cells into enucleated frogs’ eggs * Proc Natl. Acad Sci USA; 38 (5) 455-463, https://doi.org/10.1073/pnas.38.5.455
Ebisuya, M., Rayon, T., Diaz-Cuadros, M., Chalut, K. J., Wu, G., Dodd, A. N., Torres-Padilla, M. E., Levine, M. and Gladyshev, V. N. (2024). Understanding how cells and organisms keep time during development. Dev Cell; 59(13):1623-1627. doi: 10.1016/j.devcel.2024.05.029
Elsdale, T. R., Gurdon, J. B. and Fischberg, M. (1960). A Description of the Technique for Nuclear Transplantation in Xenopus laevis. Development; 8 (4): 437–444. doi: https://doi.org/10.1242/dev.8.4.437
Elsdale, T. and Davidson, D. (1987). Timekeeping by frog embryos, in normal development and after heat shock. Development; 99 (1): 41–49. doi: https://doi.org/10.1242/dev.99.1.41
Garcia-Ojalvo, J. and Bulut-Karslioglu, A. (2023). On time: developmental timing within and across species. Development; 150 (14): dev201045. doi: https://doi.org/10.1242/dev.201045
Gurdon, J., Elsdale, T. and Fischberg, M. (1958). Sexually Mature Individuals of Xenopus laevis from the Transplantation of Single Somatic Nuclei. Nature; 182, 64–65. https://doi.org/10.1038/182064a0
Gurdon, J. B. (1962). The Developmental Capacity of Nuclei taken from Intestinal Epithelium Cells of Feeding Tadpoles. Development; 10 (4): 622–640. doi: https://doi.org/10.1242/dev.10.4.622
Gurdon, J. B. (2013b). The egg and the nucleus: a battle for supremacy. Development; 140 (12): 2449–2456. doi: https://doi.org/10.1242/dev.097170
Hankins, L. E. and Rutledge, C. E. (2023). Class of 1923: looking back at the authors of JEB’s first issue. J Exp Biol; 226 (1): jeb245424. doi: https://doi.org/10.1242/jeb.245424
Jones, K. W. and Elsdale, T. R. (1963). The Culture of Small Aggregates of Amphibian Embryonic Cells in vitro. Development; 11 (1): 135–154. doi: https://doi.org/10.1242/dev.11.1.135
Kale, G., Agarwal, P., Diaz-Larrosa, J. J. and Lemke, S. (2023). Elevated temperature fatally disrupts nuclear divisions in the early Drosophila embryo. bioRxiv; 2023.09.17.558127; doi: https://doi.org/10.1101/2023.09.17.558127
Klepstad, J. and Marcon, L. (2024). The Clock and Wavefront Self-Organizing model recreates the dynamics of mouse somitogenesis in vivo and in vitro. Development; 151 (10): dev202606. doi: https://doi.org/10.1242/dev.202606
Knight, K. (2023). Journey through the history of Journal of Experimental Biology: a timeline. J Exp Biol; 226 (22): jeb246868. doi: https://doi.org/10.1242/jeb.246868
Miao, Y. and Pourquié, O. (2024). Cellular and molecular control of vertebrate somitogenesis. Nat Rev Mol Cell Biol; 25, 517–533. https://doi.org/10.1038/s41580-024-00709-z
Moriyama, D. F., Makri, D., Maalouf, M.-N., Adamova, P., de Moraes, G. F. A., Pinheiro, M. de O., Bernardineli, D. L., Massaia, I. F. D. S., Maalouf, W. E. and Lo Turco, E. G. (2022). The effects of temperature variation treatments on embryonic development: a mouse study. Scientific Reports; 12(1), 2489. https://doi.org/10.1038/s41598-022-06158-y
Murray, P. J., Maini, P. K. and Baker, R. E. (2011). The clock and wavefront model revisited. J Theor Biol; 283(1):227-38. doi: https://doi.org/10.1016/j.jtbi.2011.05.004. Epub 2011 May 27. PMID: 21635902
Škreb, N. and Frank Z. (1963). Developmental Abnormalities in the Rat Induced by Heat Shock. Development; 11 (2): 445–457. doi: https://doi.org/10.1242/dev.11.2.445
Takahashi, K. and Yamanaka, S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell; 126(4):663-76. doi: https://doi.org/10.1016/j.cell.2006.07.024
Continuing the ‘First issues’ series, in this post we’ll find out more about Audrey Muggleton-Harris, who published in Development’s first issue in 1987.
Audrey Muggleton-Harris was born in London, England, in 1932. Throughout her prolific career spanning over 40 years, she moved across the Atlantic several times while continuing her research into in vitro systems, early human embryonic development and improvements in assisted reproductive technologies.
While the reasons behind Muggleton-Harris’ multiple relocations are not clear (but probably involved a combination of professional opportunities and personal motivations), moving abroad is still very common, and at times challenging, for academic researchers in the present day. For Muggleton-Harris, the UK and the USA no doubt played a vital role in shaping her professional and personal lives.
Photo of Audrey Muggleton-Harris while she was working at Worcester Polytechnic Institute in the 1970s (From WPI Historical Images, made available under the CC BY-NC 4.0 license.)
Early experiments on nuclear transfer
Muggleton-Harris was originally trained as a teacher, but after reading about the work of cell biologist James Danielli, she joined his team as a research assistant at King’s College London in 1952 [1]. James Danielli was a Director of The Company of Biologists, and helped find a publisher for the previous incarnation of Development – Journal of Embryology and Experimental Morphology (JEEM).
Together, Muggleton-Harris and Danielli performed some of the earliest cloning experiments, first in amoebae [2] and later in frog embryos [3]. They transferred nuclei between cells to test the ability of the transferred nucleus to direct development in a different cytoplasmic environment. These early experiments paved the way for future cloning techniques.
In the early 1960s, Danielli was invited to lead the Center for Theoretical Biology at the State University of New York. Most of Danielli’s group at King’s College London, including Muggleton-Harris, crossed the Atlantic to join him. This move foreshadowed the multiple relocations between the USA and the UK that Muggleton-Harris would make over the next few decades.
In the late 1960s, Muggleton-Harris was a Research Associate at the Institute for Cancer, and then became the Research Director of the Cell Biology program at the Wills Eye Hospital. Muggleton-Harris and her family went back ‘home’ to the UK in 1972 and she worked in Cambridge University. But after just 2 years, they returned to the USA, where Muggleton-Harris was a Research Fellow and Professor of Biology and Biotechnology at Worcester Polytechnic Institute from 1976 to 1983. From 1983 till her retirement in 1997, she would remain in the UK, working at the MRC Experimental Embryology and Teratology Unit in Surrey [1].
Developing in vitro systems to understand development and disease
In her paper in Development issue 1, Muggleton-Harris and co-author N. Higbee found that fragments of the lens capsule, a thin membrane around the eye, could promote mouse lens epithelial cells to synthesize the collagen, glycoproteins and proteoglycan components required for further lens cell differentiation in vitro. This paper was part of a body of work by Muggleton-Harris on culturing mice lens epithelial cells to understand normal lens development and congenital cataracts. Since this Development issue 1 paper, Muggleton-Harris published four more papers in Development during her time working at the MRC in the UK.
Left: A clone of cultured mouse lens epithelial (MLE) cells; Right: A monolayer of MLE cells grown from cloned cells. Figures from Muggleton-Harris & Higbee, 1987 [4]
Preimplantation development and genetic testing
Her work in lens development was in fact only one part of Muggleton-Harris’ significant contribution to in vitro systems and early development. Almost a decade prior to the Development issue 1 paper, during her two-year stint in Cambridge Univeristy, she published a paper in JEEM in 1976 on ‘surface alloantigens on preimplantation mouse embryos’, investigating immune recognition and compatibility during early development, which is important in understanding maternal-fetal interactions.
In 1982, Muggleton-Harris et al. reported the first successful mammalian transfer of cytoplasm in mice, demonstrating that an unidentified cytoplasmic factor(s) can prevent embryo arrest at the two-cell stage in vitro [5]. In 1988, together with Marilyn Monk and colleagues, Muggleton-Harris developed the trophectoderm biopsy technique in mice [6]. This involved extracting cells from the trophectoderm layer of a blastocyst-stage embryo for preimplantation genetic diagnosis, while minimising damage to the embryo. A few years later, Muggleton-Harris and Findlay tested this method in ‘spare’ human preimplantation embryos in culture [7].
These are just a few examples of Muggleton-Harris’ effort into developing and refining methods for genetic analysis of biopsied cells from preimplantation embryos. Her work has contributed significantly to improving outcomes in assisted reproductive techniques and enhancing the accuracy and safety of preimplantation genetic testing.
An active member of scientific and local communities
Apart from her research, Muggleton-Harris was also a valued member of the scientific community, serving as a committee member of the British Society for Developmental Biology (BSDB) and organising the 1992 BSDB Autumn meeting on the developmental basis of inherited diseases [8].
After many prolific years at the MRC in the UK, Muggleton-Harris retired in 1997 and moved across the Atlantic one last time to Cape Cod, Massachusetts. During her retirement years, according to her obituary [1], she “volunteered as a docent at the Cape Cod Museum of Art… and will be remembered by visitors to the museum as the very proper Englishwoman who led museum tours and the annual historic house and garden bus tours” – a testament to how she left a mark on both sides of the pond, scientifically and socially.
[2] Muggleton A, Danielli JF. Inheritance of the “life-spanning” phenomenon in Amoeba proteus. Exp Cell Res. 1968;49(1):116-120. doi:10.1016/0014-4827(68)90524-7
[3] Muggleton-Harris AL. Cellular changes occurring with age in the lens cells of the frog (Rana pipiens) in reference to the developmental capacity of the transplanted nuclei. Exp Gerontol. 1970;5(3):227-232. doi:10.1016/0531-5565(70)90042-2
[4] A. L. Muggleton-Harris, N. Higbee; Factors modulating mouse lens epithelial cell morphology with differentiation and development of a lentoid structure in vitro. Development 1 January 1987; 99 (1): 25–32. doi: https://doi.org/10.1242/dev.99.1.25
[5] Muggleton-Harris A, Whittingham DG, Wilson L. Cytoplasmic control of preimplantation development in vitro in the mouse. Nature. 1982;299(5882):460-462. doi:10.1038/299460a0
[6] Monk M, Muggleton-Harris AL, Rawlings E, Whittingham DG. Pre-implantation diagnosis of HPRT-deficient male and carrier female mouse embryos by trophectoderm biopsy. Hum Reprod. 1988;3(3):377-381. doi:10.1093/oxfordjournals.humrep.a136711
[7] Muggleton-Harris AL, Findlay I. In-vitro studies on ‘spare’ human preimplantation embryos in culture. Hum Reprod. 1991;6(1):85-92. doi:10.1093/oxfordjournals.humrep.a137264
As part of our ‘first issues’ series to mark The Company of Biologists’ 100th anniversary, Development’s in-house team are researching the authors of articles published in the first issues of the Journal of Embryology and Experimental Morphology (JEEM) and its reincarnation, Development. In this post, we meet Ruth Clayton, a biologist who worked at the University of Edinburgh and who published an article in the first issue of JEEM.
Ruth Clayton was born in London in 1925 [1], the same year that our publisher, The Company of Biologists, was founded. Since Clayton would have turned 100 this year, it feels appropriate to honour her as part of the Company’s own 100th birthday celebrations. Clayton studied Zoology at Oxford before moving to the Institute of Animal Genetics (IAG) at the University of Edinburgh. Interestingly, this links Clayton to The Company of Biologists, since the IAG was initially established as the Department of Research in Animal Breeding by Francis Crew, who happens to have been the Managing Editor of our sister journal, Journal of Experimental Biology (JEB). I found out a bit about Crew and the institute a couple of years ago when researching the authors from JEB’s first issue [2]. The journal, which was founded in 1923 as the British Journal of Experimental Biology, quickly ran into financial difficulties, and George Parker Bidder III founded The Company of Biologists to safeguard its future [3].
By the time Clayton joined the IAG, it had come under the leadership of C. H. Waddington. Waddington is of course best known for his iconic epigenetic landscape [4], but he was also a member of JEEM’s first Editorial Board [5]. Clayton helped produce a gift for Waddington’s 50th birthday: a commemorative photo album [6]. This is worth a mention, mostly because it features candid shots of other IAG researchers [7] including Charlotte Auerbach and Tom Elsdale (who we’ll hear about on Friday, in the final post of this series), but also because the album documents the institute’s “Drosophila ballet”, which looks just as bizarre as it sounds [8]. The photo album was presented to Waddington at his birthday party, which seems to have been a similarly eccentric event, apparently featuring a pinball machine modelled on Waddington’s landscape [9]. Sadly, I could not find photographic evidence of the original machine, but during my search I did stumble across a paper that redraws Waddington’s epigenetic landscape as a pinball machine [10; the idea seems to be that the machine’s flippers can promote dedifferentiation]. So, maybe the party organisers were onto something.
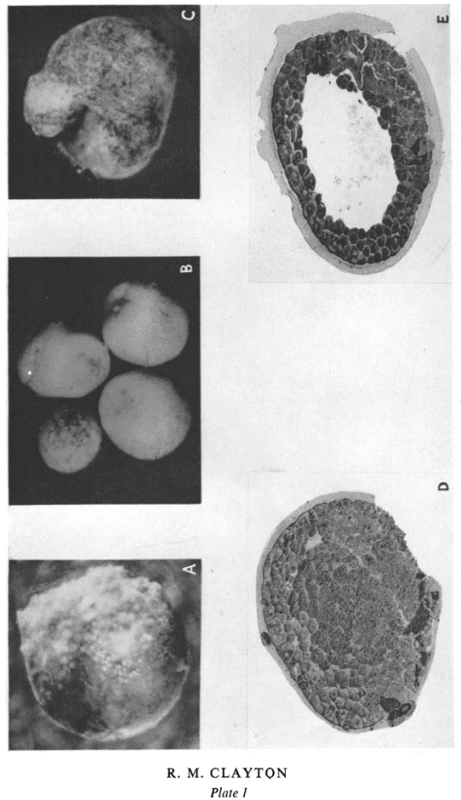
Clayton’s article in JEEM’s first issue focuses on antigen specificity in the alpine newt (Triturus alpestris) embryo [11]. She begins her paper with a statement that still rings true for developmental biologists over 70 years later: “The mechanism of differentiation presents one of the central problems of embryology.” Clayton’s study aimed to address one aspect of this problem by investigating how antigens in the early embryo differ from those in the adult, to give hints as to what changes during the process of differentiation. To do this, she dissected different sections of newt embryos (e.g. ectoderm, mesoderm and tailbud) at different stages (e.g. blastula, gastrula) and homogenised them for injection into rabbits, to produce antisera.
Fig 1: Plate from Ruth Clayton’s paper in the first issue of JEEM [11] showing examples of embryos placed in different combinations of antiserum.
The use of antisera in biology goes back a remarkably long way; at the turn of the 20th century, Emil von Behring pioneered ‘serum therapy’, injecting mammals including guinea pigs with diphtheria and tetanus toxins and ultimately using the resulting antiserum to treat patients [12, 13]. Nowadays, developmental biologists might be most familiar with using antibodies for Western blots, or for immunostaining. In her paper, Clayton mixed the antisera and antigens produced from different embryonic structures and recorded whether they cross-reacted. She went on to test the effects of placing embryos (Fig. 1) and explants in sera, antisera or absorbed antisera (i.e. the antigen and antisera mixes from earlier in the paper). The results are all a bit of a puzzle to tease out, because the experiments are done in multiple combinations, but they allow Clayton to deduce the existence of common antigens (which are present throughout development) and antigen fractions that arise at specific timepoints. For example, she notes that blastulae die or exhibit perturbed development when placed in “anti-gastrula serum absorbed with blastula” and suggests that this “is due to antibodies left in the medium after removal of antibodies to blastula antigens, i.e. that at gastrulation a new fraction appears”.
Clayton explains that she chose to work with the alpine newt for a couple of reasons: she needed “an embryo whose dissection might prove relatively easy and large numbers of which were available”. Today, other salamander species, such as the axolotl, are widely used to study regeneration. By contrast, the alpine newt doesn’t seem to have endured in developmental biology research. Indeed, a very quick search of Development’s archives suggests that it’s been nearly 50 years since we last published a research paper about alpine newts (to be fair, it’s a memorable article, featuring a chimeric newt with six legs (Fig. 2, 14). However, there is evidence that JEEM used to feature a pleasing range of newt species, including the California newt [15], the Iberian ribbed newt [16] and the Japanese fire-bellied newt [17]. Clayton’s rationale is a reminder of the importance of selecting the most appropriate organism to address a particular research question, so maybe the alpine newt will make a comeback when the circumstances are right. We highlighted the use of unconventional model organisms in our 2024 Special Issue [18].
Fig. 2: A six-legged alpine newt featured in a later JEEM paper [14]
Clayton published in JEEM a total of four times. She continued to make use of antisera in her investigations, but her subsequent work focused on cell differentiation in the context of the developing chick eye. This included research into the transdifferentiation of neural retina cells into pigment and lens cells [19]. Today, transdifferentiation is an active area of research that may hold promise for stem cell-based therapies. Clayton remained at the IAG, where Waddington later won funding to establish an Epigenetics Research Group. However, according to an obituary written by Alan Robertson, much of the project’s resources were used for molecular biology research that had no clear implications for development [9]. Indeed, Robertson mentions Clayton as “a notable exception” whose work on avian lens development “fitted into Waddington’s original concept”. Later in her career, Clayton’s expertise in lens development saw her lead an interdisciplinary collaboration to investigate risk factors for cataract formation [1]. She retired in 1993 and was appointed Reader Emeritus. Clayton’s obituary was published in the University of Edinburgh’s eBulletin in February 2003. It states that “though never formally a Director of Studies, she was often the member of staff to whom students turned for guidance”, suggesting that she was a dedicated mentor as well as a great scientist.
[2] Hankins, L. E. and Rutledge, C. E. (2023) Class of 1923: looking back at the authors of JEB’s first issue. J Exp Biol; 226 (1): jeb245424. doi: https://doi.org/10.1242/jeb.245424
[3] Knight, K. (2023) Journey through the history of Journal of Experimental Biology: a timeline. J Exp Biol; 226 (22): jeb246868. doi: https://doi.org/10.1242/jeb.246868
[9] Robertson, A. (1977) Conrad Hal Waddington, 8 November 1905-26 September 1975. Biogr. Mems Fell. R. Soc; 23, 575–622. doi: https://doi.org/10.1098/rsbm.1977.0022
[10] Sareen, D. and Svendsen, C.N. (2010) Stem cell biologists sure play a mean pinball. Nature biotechnology; 28(4), 333-335. doi: https://doi.org/10.1038/nbt0410-333
[11] Clayton, R. M. (1953) Distribution of Antigens in the Developing Newt Embryo. Journal of Embryology and Experimental Morphology; 1 (1): 25–42. doi: https://doi.org/10.1242/dev.1.1.25
[12] Behring, E. V. (1890) Ueber das zustandekommen der diphtherie-immunität und der tetanus-immunität bei thieren. Dt. Med. Wochenschr; 49, 1113–1114.
[13] Behring, E. V. (1913) Ueber ein neues Diphtherieschutzmittel. Dt. Med. Wochenschr; 39: 873-876.
[14] Houillon, C. (1977) Tractus uro-génital des chimères chez l’amphibien modèle Triturus alpestris Laur. Journal of Embryology and Experimental Morphology; 42 (1): 15–28. doi: https://doi.org/10.1242/dev.42.1.15
[15] Tucker, R. P. and Erickson, C. A. (1986) The control of pigment cell pattern formation in the California newt, Taricha torosa. Journal of Embryology and Experimental Morphology; 97 (1): 141–168. doi: https://doi.org/10.1242/dev.97.1.141
[16] Deparis, P. and Jaylet, A. (1984) The role of endoderm in blood cell ontogeny in the newt Pleurodeles waltl. Journal of Embryology and Experimental Morphology; 81 (1): 37–47. doi: https://doi.org/10.1242/dev.81.1.37
[17] Matsuda, M. (1980) Cell surface properties of amphibian embryonic cells. Journal of Embryology and Experimental Morphology; 60 (1): 163–171. doi: https://doi.org/10.1242/dev.60.1.163
[18] Extavour, C., Dolan, L., Sears, K. E. (2024) Promoting developmental diversity in a changing world. Development; 151 (20): dev204442. doi: https://doi.org/10.1242/dev.204442
[19] De Pomerai, D. I. and Clayton, R. M. (1978) Influence of embryonic stage on the transdifferentiation of chick neural retina cells in culture. Journal of Embryology and Experimental Morphology; 47 (1): 179–193. doi: https://doi.org/10.1242/dev.47.1.179
Launched in 2020, Development presents… is a monthly Zoom webinar series covering the fields of developmental biology, stem cells and regeneration. Each webinar features three speakers who present short talks loosely grouped on a particular topic, and have been hosted by a Development Editor who mediates written questions from the audience. With the speakers’ permission, webinars are recorded and are available to watch on demand.
Since 2023, speakers have been selected from the pool of postdocs applying for their first independent position as part of Development’s Pathway to Independence (PI) programme. Although this is a fantastic way to highlight and support postdocs during their transition to becoming a principal investigator with additional exposure for them and their research, applications to the programme have not always reflected the geographical diversity of the field.
Therefore, we would like to supplement the applicants of the PI programme with additional self-nominations from postdocs – based outside of the US and Europe – looking to transition to secure an independent position within the next year or so.
If you would like to be considered as a Development presents… speaker in 2025 (and early 2026), please follow the link to the self-nomination form below. The deadline for nominations is Friday, 28 March 2025.
• work on a topic within the scope of Development. • have a first-author publication or preprint from their postdoctoral research. • be applying, or ready to apply, for their first independent position in the next year. • be based outside of the US or Europe.
This post is part of our ‘first issues’ series on the Node – looking at some of the papers, and the researchers behind them, that appeared in the first issues of Journal of Embryology and Experimental Morphology (JEEM) and Development.
This piece turns the spotlight on an article published in the first ever issue of JEEM, over 70 years ago, with a surprisingly modern theme. The paper is written in French, a common occurrence in the early days of JEEM, but now accessible in English thanks to our recent digitisation of the journal’s full archive and the wonders of Google translate. So let’s dive in…
Many of you working with stem cells and organoids have likely used Matrigel or similar cell-derived products for cell and tissue culture. And you will likely be familiar with the frustrations associated with its heterogeneity – one batch can differ from another, making reproducibility a significant problem. For this reason, many researchers have now turned to fully synthetic hydrogels, whose composition is defined and consistent, for in vitro culture.
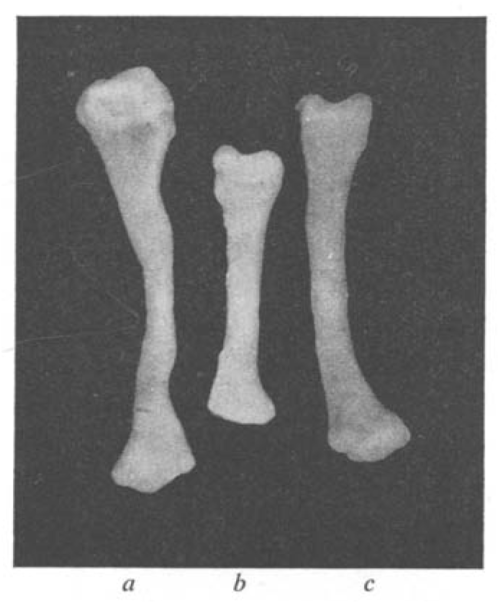
But you may not be aware that researchers of the 1950s faced similar issues. The search for a synthetic medium that could support organ growth in vitro is the topic of one of the papers in the first ever issue of JEEM. Published by husband-and-wife team Etienne and Emilienne Wolff and others, “Essais de cultures in vitro d’organes embryonnaire en milieux synthetiques” (translated as “In vitro culture tests of embryonic organs in synthetic media”) reports the ability of various synthetic media to support the in vitro growth and differentiation of avian explanted organs. Etienne Wolff, along with Katy Haffen (also an author on the JEEM paper), had previously published a string of papers describing successful short-term culture of a wide range of chicken and duck embryonic organs in their ‘standard medium’ – which contained chicken embryo extract and a glucose-containing solution. In the JEEM paper, they looked to replace this extract with media containing defined concentrations of amino acids and vitamins. These experiments demonstrated the feasibility of such an approach – showing that a fully synthetic medium could support the survival, growth and differentiation of explanted chick embryo gonad, syrinx (the bird vocal organ – I had to look that one up!) and tibia, at least over the course of a few days. The set of papers that established effective in vitro culture conditions for organs opened up opportunities for studying and manipulating organogenesis in ways that were not – at the time – possible in the embryo.
Figure 5 from Wolff et al. (1953) a shows a tibia cultured in ‘standard medium’; b shows a tibia cultured in glucose medium without nitrogenous substances; c shows a tibia cultured in synthetic medium containing amino acids.
So, who were the team behind this study? Etienne Wolff seems to have been something of a renaissance man. Born in 1904, he first graduated with a degree in literature before turning his attention to science. Although his subsequent career was devoted to biology, he was also made a member of the Académie Française (the body responsible for the rules of the French language). Having graduated with a PhD in 1936, his career was interrupted by the Second World War, where he was captured by the Germans and spent most of the war in prisoner of war camps before being liberated in 1945. Wolff was not idle during his captivity – he created a university within the camp, and wrote two books, “La Science des Monstres” and “Les Changements de Sex”. After the war, he resumed his research in Strasbourg. Having set out in his PhD to reproduce ‘spontaneous monstrosities’ in the avian gonad, to study both the generation of teratomas and the development of these organs, teratology and sexual differentiation were research themes that continued throughout his career. Indeed, through his work in birds, Wolff was (I believe) the first to demonstrate conclusively that intersexuality or sex reversal could be experimentally induced by sex hormones. The desire to better understand male versus female development of the gonad was the driving force for his work on in vitro organ culture – though he also applied these techniques to many other organs.
Etienne Wolff. Image reproduced with permission from Galperin, 2005.
In 1955, Wolff was appointed Chair of Experimental Embryology at the Collège de France and founded the Nogent Institute of Experimental Embryology and Teratology, where he spent the rest of his research career. For a decade, he also served as the Director of the Collège de France. In 1976, he was succeeded as Director of the Nogent institute by his former student and another developmental biology great, Nicole le Douarin. After his retirement from research, he became active in supporting animal rights, serving as Chair of the Animal Rights, Ethics and Science Foundation in the 1980s. He died in 1996.
Much less information is available about Emilienne Wolff, though she was clearly a distinguished scientist in her own right – serving as a CNRS Director and a laureate of the French Academy of Sciences. At a symposium organised in their honour at the time of their retirement, their colleague Etienne Lasfargues described their research programme at the Nogent Institute thus: “This huge coordinated research, acutely aimed at a deeper knowledge of life, functioned very much like a symphonic orchestra in which Professor [Etienne] Wolff was the maestro, and Madame [Emilienne] Wolff, the concertmaster”.
While the JEEM paper is over 70 years old, its core concern – how to recapitulate embryonic development in an in vitro setting – is a topic that resonates today, with hundreds of labs worldwide working to grow and study organoids of all different types. Though the Wolffs’ approach and outcomes were much less sophisticated than what we can now achieve, their body of work is part of the long history of what is often viewed as a very recent field.
Sources:
Essais de cultures in vitro d’organes embryonnaires en milieu synthétiques Etienne Wolff, Katy Haffen, Madeline Kieny and Emilienne Wolff. J. Embryol. Exp. Morphol. 1 (1): 55-84 (1953) doi: 10.1242/dev.1.1.55
L’École de Nogent: the contributions of Etienne Wolff and Nicole Le Douarin Charles Galperin Int. J. Dev. Biol. 49: 79-83 (2005) doi: 10.1387/ijdb.041944cg
The Nogent Institute – 50 Years of Embryology Nicole le Douarin Int. J. Dev. Biol. 49: 85-103 (2005) doi: 10.1387/ijdb.041952nl
Introduction of Madame Emilienne Wolff and Professor Etienne Wolff Etienne Lasfargues In Vitro. 1979 Jan;15(1):3-5. doi: 10.1007/BF02627072
The elusive importance of NR5A2 and ESRRB as pluripotency factors
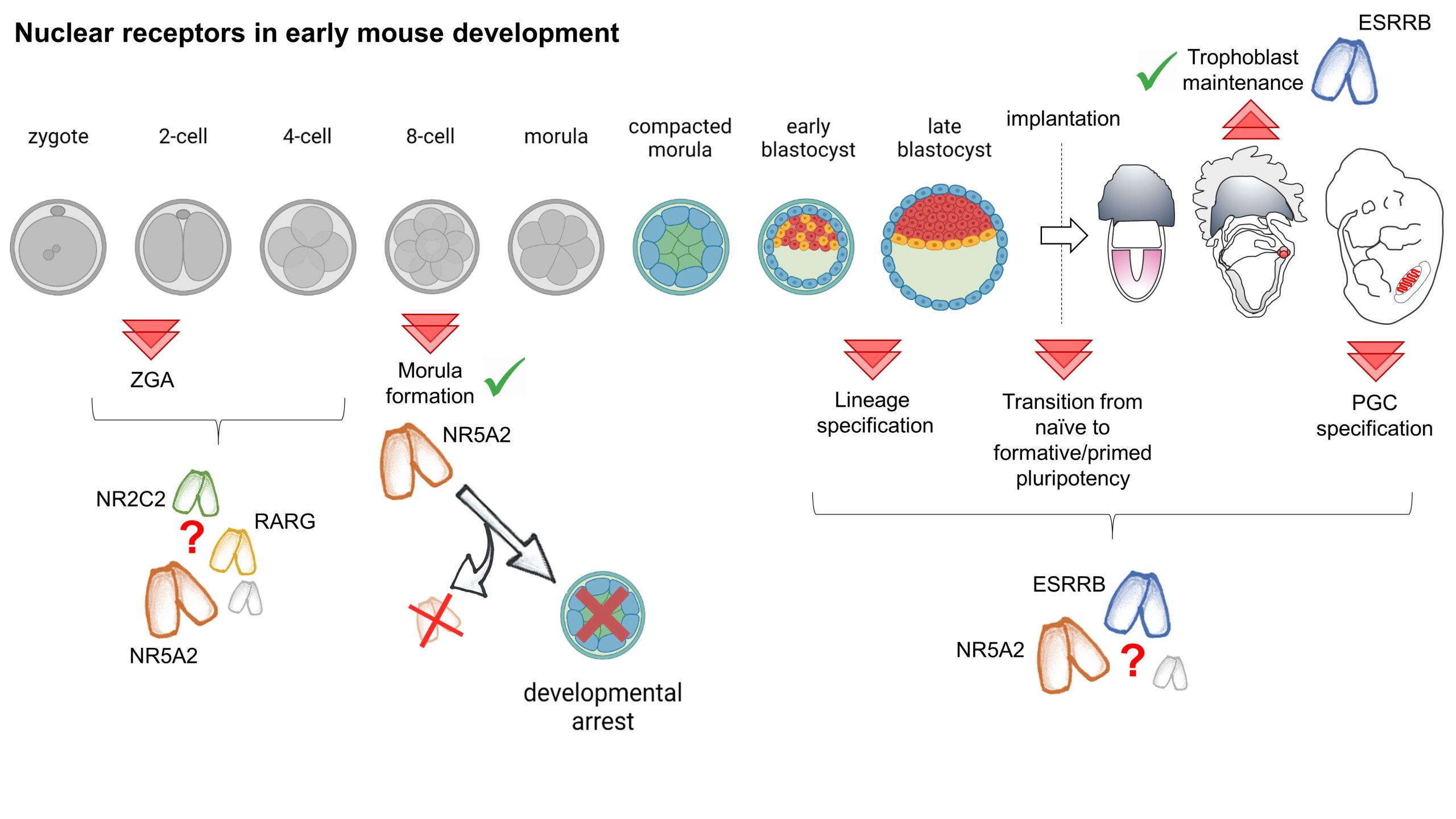
Our paper entitled “Nr5a2 is dispensable for zygotic genome activation but essential for morula development” is the culmination of a long scientific journey working on mouse embryonic stem cells (ESCs), which I initiated during my PhD in the group of Ian Chambers, in Edinburgh, where we showed that the nuclear receptor Esrrb acts as a pluripotency transcription factor (TF) [1]. Back then, we also first asked the question that eventually led to our recent publication: if ESRRB is important for pluripotency, how can embryos specify an epiblast in the absence of this TF? In fact, the available literature showed that Esrrb KO embryos do not manifest evident defects in the pluripotent lineage [2, 3]. This contradiction became more strident in the following years. My work, first as a postdoc and then as a permanent CNRS researcher in the group of Pablo Navarro in Paris, together with results from several other groups, showed that ESRRB performs unique functions in ESCs: it conveys external signals[4, 5], binding and maintaining the activity of key nodes of the pluripotency network[6], including during mitosis [7-9]. As a consequence, the loss of ESRRB marks the beginning of differentiation and, conversely, ESRRB expression drives the completion of somatic cell reprograming [1, 10-12].
We soon came to realise that the role of ESRRB in pluripotent cells cannot be understood without considering its interaction with other nuclear receptors, especially NR5A2. Indeed, while ES cells tolerate the individual loss of either ESRRB or NR5A2, in the simultaneous absence of these two nuclear receptors the pluripotency network collapses remarkably fast [9, 13]. Thus, while remaining distinct TFs, ESRRB and NR5A2 act as a single functional unit, which rivals in importance with that of the core components of the pluripotency network: their redundant activity is strictly required for pluripotency.
Shared and unique roles of NR5A2 in building a morula
Having identified two essential players in the maintenance of ES cell identity, we now had the knowledge needed to test the role that these two orphan nuclear receptors play in driving the establishment of pluripotency in vivo. We hypothesized that the lack of a phenotype in Esrrb KO embryos was likely due to a compensation by NR5A2. To test this, we joined efforts with the team led by Michel Cohen Tannoudji, who had joined Pablo’s group. Together, we generated animals carrying floxed alleles for both genes and expressing Zp3:Cre, which drives recombination in growing oocytes. We crossed these females with heterozygous males, and recovered control, single KO, and double KO embryos at the blastocyst stage. This experiment, a few years in the making, generated the promised exciting results… but unceremoniously contradicted our expectations! While controls had almost completed lineage specification, Nr5a2 KO embryos appeared severely degenerated, and this is… irrespective of Esrrb expression [14].
Following on this unexpected observation, our work, together with the results of two independent reports from the groups of Wei Xie [15] and Kikue Tachibana [16], revealed an essential developmental role for Nr5a2. Rapidly upregulated after the activation of transcription from the zygotic genome (ZGA), which occurs at the 2-cell stage (2C-stage) in mouse, NR5A2 controls the first wave of embryonic gene expression. NR5A2 targets include many key developmental regulators, such as Nanog, Klf5, Sox15, Tead4, Gata3 and Cdx2, but also genes supporting housekeeping functions, such as cell division, and DNA repair. As a consequence, during the two rounds of cell divisions that follow the activation of transcription, NR5A2 performs a dual function: it lays the ground for the beginning of lineage specification, all while maintaining genomic stability. While the contribution of ESRRB appears minor, NR5A2 does not act alone during this developmental window. At least two other TFs expressed just after ZGA, KLF5 [17, 18] and TCFP2AC [19], have been shown to contribute in preparing the segregation of the first two embryonic lineages, endowing blastomeres with the potential to generate both trophectoderm and the inner cell mass (ICM). NR5A2 activates Tcfp2a and Klf5 expression, and seems to act in conjunction with Krüppel-like factors to establish chromatin accessibility in 8C-stage embryos [14]. Reciprocally, TCFP2A activates Nr5a2 [19]. Therefore, an interconnected regulatory network, characteristic of the 8C-stage, and governed by the interaction between regulators of extraembryonic and embryonic fates, is open to future research.
The role of NR5A2 in maintaining genome stability might, instead, be specific to this TF. After the 8-cell stage, Nr5a2 KO embryos start displaying cell division defects, which progressively exacerbate, and eventually arrest before forming a blastocyst. Besides activating expression of genes that are required for the execution of cell division, NR5A2 might execute functions that are directly linked to its activity as a bookmarking TF. Is the ability of orphan nuclear receptors to remain bound to specific genomic region during mitosis playing an unforeseen role in ensuring the faithful partitioning of the genetic material? Irrespectively, despite opening a number of novel questions, these results establish that NR5A2 is required for the formation of a developmentally competent and viable morula.
Orphan nuclear receptors and the first wave of gene expression
While the transcriptional effect of the KO of Nr5a2 manifest fully only at the 8C-stage, the earlier role of this TF remains less clear. Treating embryos with chemical inhibitors targeting NR5A2, among other nuclear receptors, or depleting NR5A2 protein in ex-vivo experiments has been shown to disrupt ZGA and cause developmental arrest starting at the 2C-stage [16]. A role as an initiator of ZGA implies a determinant function of maternally inherited NR5A2. However, our experiments show that females producing oocytes devoid of Nr5a2 can give birth to live pups, and the Zp3:Cre-driven maternal or maternal and zygotic KO of Nr5a2 affects only a few hundred genes at the late 2C-stage, and compromises the activation of a minority of the genes that start being transcribed during ZGA. Similarly mild effects have been reported after the siRNA-mediated knockdown of Nr5a2 [15, 16]. In addition, we showed that the developmental defects of Nr5a2 KO embryos can be rescued by injecting Nr5a2 mRNA in 4C-stage embryos. Conversely, triggering the loss of Nr5a2 after ZGA, by the use of an inducible Cre driver, mimics the phenotype of the maternal and zygotic KO. What is the base of the discrepancy between different studies? Extending our original hypothesis, it is possible to envisage functional interactions with yet other nuclear receptors expressed at this stage of development. Among these, two stand out: NR2C2, which is also sensitive to NR5A2 inhibitors [16], and the retinoic acid receptor RARG, which has been shown to play a role in 2C embryos, and in driving ZGA [20]. Both nuclear receptors present an expression profile analogous to Nr5a2 in early embryos. In addition, among the few genes downregulated by the KO of Nr5a2 at the onset of ZGA we found a number of know regulators of the 2C transcriptional programme, including Zscan4 genes, themselves involved in activating embryonic transcription. Curiously, while NR5A2 seems to activate these genes at the time of ZGA, at later stages mutant embryo fail, to a variable extent, to correctly repress their expression. Therefore, the interplay between different nuclear receptors and other classes of TFs in controlling the 2C-stage transcriptional programme, ZGA, and the progression past this stage of development remain an active area of investigation. Despite these uncertainties, it is however clear that NR5A2 accumulates in the blastomeres as early as the late 2C stage; if not playing a role in initiating ZGA, this TF surely starts regulating embryonic gene expression soon after the onset of embryonic transcription.
Open Questions
Despite the excitement brought by these results, our work leaves our original question standing. New tools need to be developed to bypass the requirement for NR5A2 in the formation of a morula able to begin lineage specification, and assess the conjunct role that ESRRB and NR5A2 may play during the execution of these cell fate choices. Indeed, beside an essential role as a pluripotency TFs, and potentially in the establishment of pluripotency, experiments in ESCs indicate that ESRRB and NR5A2 might have both overlapping and distinct functions in the segregation of the epiblast from the primitive endoderm [21-24].
Looking beyond pre-implantation development, ESRRB and NR5A2 might also play a role in germline specification. ESRRB expression is extinguished in the epiblast at implantation [3, 14]. Studies in ESCs indicate that, before being downregulated, ESRRB may prime the transition between the gene regulatory networks that support pre- and post-implantation pluripotency [25]. These changes are in turn required for the activation of the germ cell programme in a few cells in the posterior region of the epiblast. Both Esrrb and Nr5a2, together with a number of other pluripotency regulators, are then re-expressed in these nascent primordial germ cells [26]. In-vivo and in-vitro studies hinted at the involvement of the two orphan receptors in multiple aspect of germ cell specification [3, 25, 27] but did not report an essential function in supporting germ cell identity. However, functional compensation might have masked the role of orphan receptors also in this context.
Developmental TF: lone wolves or social animals?
Our work illustrates both the individual and collective importance of orphan nuclear receptors in regulating cell identity and developmental processes. These TFs, able to access the very same sites on DNA, show sign of a functional redundancy that goes beyond the generic cooperativity between TFs [13]. However, if ESRRB and NR5A2 are collectively required for the maintenance of pluripotency in ESCs, and possibly the epiblast, we have observed that NR5A2 alone is essential for morula development. Could then redundancy be inbuilt in the control of some developmental processes and not others? And if so with what implications? Redundancy provides a fail-safe mechanism to ensure the robustness of crucial gene regulatory processes, and has been observed in other TF families active during early development such as Krüppel-like and Gata factors [17, 28, 29]. Studies ranging from the analysis of stress response TF in yeast [30] to work on retinoic acid receptors [31], indicate that redundant TF pairs, while showing the ability to regulate a substantial fraction of common target genes, often maintain some degree of specificity. This property can be exploited to achieve robust expression of a common set of genes, while retaining the ability to modulate either of the uniquely regulated gene sets. One could imagine that some of the genes exclusively regulated by a TF operating redundantly with NR5A2, for instance ESRRB, must not be activated or repressed during morula formation, when NR5A2 expression peaks, and its effects are dominant. Ensuring strong activation of a common gene set, like that supporting the pluripotency programme, might instead be later required during epiblast specification, when NR5A2 levels are declining and ESRRB is still robustly expressed. With potential functions spanning through events that link fertilisation to the establishment of the germline, acting across cell cycle phases, and in a collective or individual fashion, studying nuclear receptors promises to teach us much about how redundancy contributes to the making and unmaking cell identity during development. Are developmental TFs lone wolves or social animals?
References
1. Festuccia, N., et al., Esrrb is a direct Nanog target gene that can substitute for Nanog function in pluripotent cells. Cell Stem Cell, 2012. 11(4): p. 477-90.
2. Luo, J., et al., Placental abnormalities in mouse embryos lacking the orphan nuclear receptor ERR-beta. Nature, 1997. 388(6644): p. 778-82.
3. Mitsunaga, K., et al., Loss of PGC-specific expression of the orphan nuclear receptor ERR-beta results in reduction of germ cell number in mouse embryos. Mech Dev, 2004. 121(3): p. 237-46.
4. Martello, G., et al., Esrrb is a pivotal target of the gsk3/tcf3 axis regulating embryonic stem cell self-renewal. Cell Stem Cell, 2012. 11(4): p. 491-504.
5. Hamilton, W.B., et al., Dynamic lineage priming is driven via direct enhancer regulation by ERK. Nature, 2019. 575(7782): p. 355-360.
6. Whyte, W.A., et al., Master transcription factors and mediator establish super-enhancers at key cell identity genes. Cell, 2013. 153(2): p. 307-19.
7. Festuccia, N., et al., Mitotic binding of Esrrb marks key regulatory regions of the pluripotency network. Nat Cell Biol, 2016. 18(11): p. 1139-1148.
8. Festuccia, N., et al., Transcription factor activity and nucleosome organization in mitosis. Genome Res, 2019. 29(2): p. 250-260.
9. Chervova, A., et al., Mitotic bookmarking redundancy by nuclear receptors in pluripotent cells. Nat Struct Mol Biol, 2024.
10. Huang, D., et al., LIF-activated Jak signaling determines Esrrb expression during late-stage reprogramming. Biol Open, 2018. 7(1).
11. Buganim, Y., et al., Single-Cell Expression Analyses during Cellular Reprogramming Reveal an Early Stochastic and a Late Hierarchic Phase. Cell, 2012. 150(6): p. 1209-22.
12. Polo, J.M., et al., A molecular roadmap of reprogramming somatic cells into iPS cells. Cell, 2012. 151(7): p. 1617-32.
13. Festuccia, N., et al., The combined action of Esrrb and Nr5a2 is essential for murine naive pluripotency. Development, 2021. 148(17).
14. Festuccia, N., et al., Nr5a2 is dispensable for zygotic genome activation but essential for morula development. Science, 2024. 386(6717): p. eadg7325.
15. Lai, F., et al., NR5A2 connects zygotic genome activation to the first lineage segregation in totipotent embryos. Cell Res, 2023. 33(12): p. 952-966.
16. Gassler, J., et al., Zygotic genome activation by the totipotency pioneer factor Nr5a2. Science, 2022. 378(6626): p. 1305-1315.
17. Kinisu, M., et al., Klf5 establishes bi-potential cell fate by dual regulation of ICM and TE specification genes. Cell Rep, 2021. 37(6): p. 109982.
18. Lin, S.C., et al., Klf5 regulates lineage formation in the pre-implantation mouse embryo. Development, 2010. 137(23): p. 3953-63.
19. Li, L., et al., Lineage regulators TFAP2C and NR5A2 function as bipotency activators in totipotent embryos. Nat Struct Mol Biol, 2024.
20. Iturbide, A., et al., Retinoic acid signaling is critical during the totipotency window in early mammalian development. Nat Struct Mol Biol, 2021. 28(6): p. 521-532.
21. Knudsen, T.E., et al., A bipartite function of ESRRB can integrate signaling over time to balance self-renewal and differentiation. Cell Syst, 2023. 14(9): p. 788-805 e8.
22. Uranishi, K., et al., Esrrb directly binds to Gata6 promoter and regulates its expression with Dax1 and Ncoa3. Biochem Biophys Res Commun, 2016. 478(4): p. 1720-5.
23. Olivieri, D., et al., Cooperation between HDAC3 and DAX1 mediates lineage restriction of embryonic stem cells. EMBO J, 2021. 40(12): p. e106818.
24. Herchcovici Levy, S., et al., Esrrb is a cell-cycle-dependent associated factor balancing pluripotency and XEN differentiation. Stem Cell Reports, 2022. 17(6): p. 1334-1350.
25. Carbognin, E., et al., Esrrb guides naive pluripotent cells through the formative transcriptional programme. Nat Cell Biol, 2023. 25(5): p. 643-657.
26. Zhang, M., et al., Esrrb Complementation Rescues Development of Nanog-Null Germ Cells. Cell Rep, 2018. 22(2): p. 332-339.
27. Hackett, J.A., et al., Tracing the transitions from pluripotency to germ cell fate with CRISPR screening. Nat Commun, 2018. 9(1): p. 4292.
28. Home, P., et al., Genetic redundancy of GATA factors in the extraembryonic trophoblast lineage ensures the progression of preimplantation and postimplantation mammalian development. Development, 2017. 144(5): p. 876-888.
29. Yamane, M., et al., Overlapping functions of Kruppel-like factor family members: targeting multiple transcription factors to maintain the naive pluripotency of mouse embryonic stem cells. Development, 2018. 145(10).
30. Wu, Y., et al., Yeast cell fate control by temporal redundancy modulation of transcription factor paralogs. Nat Commun, 2021. 12(1): p. 3145.
31. Kastner, P., M. Mark, and P. Chambon, Nonsteroid nuclear receptors: what are genetic studies telling us about their role in real life? Cell, 1995. 83(6): p. 859-69.
In the latest issue of Development, you can read about the journal’s history since launching as Journal of Embryology and Experimental Morphology (JEEM) in 1953 (Eve, 2025). To complement the Editorial, here on the Node, we compare some ‘milestone’ issues: the first issue of JEEM, the first issue of the journal after being renamed Development in 1987 and the first issue of Development published in 2025, which marks the 100th anniversary year of the journal’s publisher, The Company of Biologists (Bray et al., 2025). Inspired by our colleagues at Journal of Experimental Biology (Hankins and Rutledge, 2023), we look at how changing publishing trends have evolved through the decades, all from the perspective of the table of contents. In addition, some of Development’s in-house team will highlight papers of their choice published in the first issue of JEEM or Development over the week, with a short series of vignettes about the authors and their research. We hope you find the series interesting and that you enjoy learning a little more about the people who’ve contributed to the journal during its history.
A comparative approach
Table 1 summarises some of the statistics from the three issues. Let’s first acknowledge that journal articles vary widely from one issue to the next, so examining a single issue isn’t representative of overall trends in biology or publishing. So, for example, just because the three issues selected here don’t contain an article from India doesn’t mean the journal never publishes work from Indian researchers (indeed, we do!) or that India doesn’t have a flourishing developmental biology community (indeed, it does!). However, this minimalistic approach helps highlight some potential leads for further discussion.
7 countries: Canada (1), France (1), Germany (1), Japan (1), Switzerland (1), UK (6), USA (10)
Editorial Board members
23
48
107
Estimated gender ratio of Editorial Board members (men:women)*
21:2
38:10 (approx. 3:1)
71:36 (approx. 2:1)
Table 1. Comparing information in issues of JEEM/Development at different stages of the journal’s history. *We’ve had to estimate when authors or members provide only forename initials or have names that do not identify gender. We also acknowledge that some individuals may not identify as a man or a woman, but we cannot always determine them by name alone.
Trends in papers
One important trend that is seen across the publishing industry is that there are more authors per paper (Jakab et al., 2024). In the 1950s, when JEEM was launched, it was common – standard, even – to have publications with just one or two authors. This pattern is still true in the 1980s when Development was (re)launched but, occasionally, papers with several authors do crop up. Today, single-author research papers from the field of developmental and stem cell biology are very rare. Two-author papers are still seen, although there aren’t any of these in the first issue of 2025; it’s much more common to see papers with at least three authors. Issue 1 of 2025 also contains a couple of papers with 10 contributing authors, which I don’t think would surprise any readers. The increasing number of authors likely reflects the increasing content per paper; comparing a research article published today from one 70 years ago, it’s clear that more experiments, techniques and models are employed today, all of which can be complex and require time, money and expertise (see later posts in this series, as well as the Regeneration retrospective and Forgotten classics series). More authors are also evidence of the strong collaboration culture that exists in today’s science.
Over the years, there’s also been an increase in the geographical range of authors. JEEM was founded as a developmental biology journal for European research to complement the launch of Embryologia in Japan, so it’s no surprise that the papers in its first issue derive from European authors (Medawar, 1980). However, as a UK-based journal, it’s a little unexpected to see only one paper from the UK in this inaugural issue; looking at the first volume, there are a few more papers from British authors in later issues, as well as papers from the USA and India. UK-based authors are well represented in the first issue of Development, possibly because of the strong links between the journal and the British Society for Developmental Biology established by this point (Eve, 2025). Part of the discussion around the rebranding of the journal as Development considered the need to capture more papers from North America as a research powerhouse, reflected in more authors from Canada and the USA in the first issue – as well as the appointment of two USA-based Editors (Eve, 2025). In 2025, Development has a global reach and publishes papers from developmental and stem cell biologists worldwide. The increasing research output from Asia is illustrated by increasing numbers of papers and authors based in China and Japan, although publications from the UK and USA still make large contributions.
In early issues of JEEM, there was no such thing as a model organism; researchers used all sorts of species, things they could dredge from ponds (Pasteels, 1953), find under rocks (Brøndsted, 1953) or fish during trips to Naples (Van de Kamer and Schuurmans, 1953). Not a single article in the first issue of JEEM uses mice, although it is worth pointing out that in vitro systems (of chick cells, in this case) are represented (Wolff et al., 1953). By the first issue of Development, the field had coalesced around a handful of model systems for which there were genetic tools and large research communities, including papers from giants in the field, such as Jim Smith using Xenopus (Smith, 1987) and Rosa Beddington and Patrick Tam using mice (Beddington and Tam, 1987). The first issue of Development also features studies on moths, which shows how the journal has always supported developmental biology regardless of the experimental system (Kato et al., 1987), which remains true today – see the latest Special Issue: Uncovering Developmental Diversity. Table 1 also highlights the prominence of zebrafish papers in modern issues of Development, gaining popularity since the genetic screens were published in the ‘zebrafish issue‘ of the journal in 1996. Interestingly, the two issues of Development here feature articles on protists, Tetrahymena (Frankel and Nelsen, 1987) and Dictyostelium (Yamashita et al., 2025), which we can probably consider outliers! Plant models don’t appear in these three specific issues, although Development publishes many plant papers. In fact, when JEEM was first launched, the press suggested that ‘animal’ was added to the journal’s title, but the journal’s Editor, Michael Abercrombie, opposed this addition because it would ‘preclude the journal from ever accepting papers [that] integrate botanical and zoological aspects of the subject’ (The Company of Biologists’ Board minutes). However, it was when the journal was renamed Development in 1987 that it started actively inviting plant papers. Similarly, human models, including stem cells and organoids, although absent in this selection, are now frequently seen in Development following active recruitment of the papers as part of Olivier Pourquie’s tenure as Editor-in-Chief (Eve, 2025) – although early volumes of JEEM also contain studies on primary human tissue (e.g. O’Rahilly, 1963).
Trends in the journal
The organisation and management of the journal have also evolved alongside the papers it publishes. When JEEM was launched, Michael Abercrombie was the only (UK-based) Editor who handled research papers. The number of Editors expanded up to a team of four during the following decades, all of whom were located within Britain. As I mentioned earlier, a key focus at the time of Development’s launch was receptivity to research from the USA, and so the British team was supported by two new USA-based Editors (Eve, 2025). This year, Development has Editors from North America, Europe and Asia, which better represent the research community.
In addition to geographical range, the journal has improved its team’s gender diversity over the years. Although there was one Editor for the first issue of JEEM, there was an Editorial Board (which can support Editors with making decisions on specific papers or general journal strategy) of 23 members – only two of whom were women. Although one might argue there were fewer women in leadership positions in the 1950s, the gender of the authors published in the same issue is almost 50–50, indicating the Editorial Board is not representative. Gender balance is better in the first issue of Development, with a ratio of three men to one woman on the Editorial Board, the same as the authors. However, the eight-person Editorial team is 100% men! Now, the team at Development is roughly equal, with 11 men and 10 women Editors, which much better represents our society, and an Editorial Board of two men for every woman, which is in line with that of authors and suggestive of representing the field.
One aspect of the journal that has become less diverse over the years is language. Many articles in JEEM are written in French (e.g. Pasteels, 1953) and some other languages, such as German (e.g. von Gerhart Wagner), are also present. Although I don’t know if he was a polyglot himself, Abercrombie was a proponent for multilingual science, introducing a practice that all papers must have dual-language abstracts, frequently reporting to the publisher the number of articles published in each language, as well as the journal eventually providing some translation services to authors (Gurdon, 2013; The Company of Biologists’ Board minutes). This policy was slowly phased out between 1960-1970 as English became the de facto language for research communication.
Finally, as already alluded, the journal has expanded hugely during its lifetime to keep up with rapidly increasing research output. From a single Editor in JEEM, Development now has 21 Editors handling research papers. Similarly, the Editorial Board has roughly doubled from 23 in JEEM’s first issue to 48 in the first issue of Development and then doubled again to 107 in the journal’s current volume. The number of research papers published has remained largely stable, but modern Development supports these with a strong ‘front section’ featuring review-type articles and magazine-type content, such as interviews and perspectives, which is handled by an in-house team.
If you’re interested in learning more about the science published in these first two issues, check back again tomorrow!
References
Bray, S. J., Royle, S. J., Shiels, H. A. and St Johnston D. (2025). The Company of Biologists: celebrating 100 years. Development; 152 (1): dev204567. doi: https://doi.org/10.1242/dev.204567
Brøndsted, H. V. (1953). Rate of Regeneration in Planarians after Starvation. Development; 1 (1): 43–47. doi: https://doi.org/10.1242/dev.1.1.43
Frankel, J. and Nelsen, E. M. (1987). Positional reorganization in compound janus cells of Tetrahymena thermophila. Development; 99 (1): 51–68. doi: https://doi.org/10.1242/dev.99.1.51
Hankins, L. E. and Rutledge, C. E. (2023). Class of 1923: looking back at the authors of JEB’s first issue. J Exp Biol; 226 (1): jeb245424. doi: https://doi.org/10.1242/jeb.245424
Jakab, M., Kittl, E. & Kiesslich, T. (2024). How many authors are (too) many? A retrospective, descriptive analysis of authorship in biomedical publications. Scientometrics; 129, 1299–1328. https://doi.org/10.1007/s11192-024-04928-1
O’Rahilly, R. (1963). The Early Development of the Otic Vesicle in Staged Human Embryos. Development; 11 (4): 741–755. doi: https://doi.org/10.1242/dev.11.4.741
Pasteels, J. (1953). Les effets de la centrifugation sur la blastula et la jeune gastrula des Amphibiens: I. Mécanisme de la formation des organes secondaires aux dépens de l’ectoblaste. Development; 1 (1): 5–24. doi: https://doi.org/10.1242/dev.1.1.5
Smith, J. C. (1987). A mesoderm-inducing factor is produced by a Xenopus cell line. Development; 99 (1): 3–14. doi: https://doi.org/10.1242/dev.99.1.3
Tam, P. P. L and Beddington R. S. P. (1987). The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development; 99 (1): 109–126. doi: https://doi.org/10.1242/dev.99.1.109
Van de Kamer, J. C. and Schuurmans, A. J. (1953). Development and Structure of the Saccus Vasculosus of Scylliorhinus caniculus (L.). Development; 1 (1): 85–96. doi: https://doi.org/10.1242/dev.1.1.85
von Gerhart Wagner. (1955). Chimaerische Zahnanlagen aus Triton-Schmelzorgan und Bombinator-Papille: Mit Beobachtungen über die Entwicklung von Kiemenzähnchen und Mundsinnesknospen in den Triton-Larven. Development; 3 (2): 160–188. doi: https://doi.org/10.1242/dev.3.2.160
Wolff, E., Haffen, K., ;Kieny, M. and Wolff, E. (1953). Essais de cultures in vitro d’organes embryonnaires en milieux synthétiques. Development; 1 (1): 55–84. doi: https://doi.org/10.1242/dev.1.1.55
Yamashita, K., Shimane, K. and Muramoto T. (2025). Optogenetic control of cAMP oscillations reveals frequency-selective transcription factor dynamics in Dictyostelium. Development; 152 (1): dev204403. doi: https://doi.org/10.1242/dev.204403
We’re only just over a month into 2025, and lots have happened already. In this ‘Developing news’ post, we’ve collated a few Bluesky posts about topics that have been on people’s minds lately.
Apart from Bluesky, you can also find the Node on X, LinkedIn and Facebook. If you’ve decided to take a break from social media, you can subscribe to our weekly emails so you get notified about the latest posts and job listings on the Node.
What’s happening in the US…
Are you based in the US? Is your research affected by the recent events? We appreciate the situation is evolving very quickly, but if you would like to share your experience and thoughts, please leave a comment below.
The US National Science Foundation, a major funder of basic academic research, has reopened a website that distributes money from research grants to scientistshttps://go.nature.com/4hhd5Aq
Disruptions to the NIH impact more than just science; they will hit all Americans in our wallets. I spoke yesterday with Becky Fogel from @kutnews.bsky.social about the important role NIH-funded research plays in the economy. 🧵 1/nwww.kut.org/education/20…
I wrote some thoughts about why peer review mattersIt shapes scientific standards, maintains field coherence & trains new researchersYes, it needs improvement—but it's the glue that holds scientific progress togetherbriscoelab.org/2024/12/11/i…
Dear fellow scientists: Have you used generative AI such as chatGPT to help writing papers, particularly introductions? If so what are the ethical considerations around that? Should this help be acknowledged? I feel like this will become increasingly prevalent & we need clearly defined rules/norms.
And lastly, an appreciation of the beauty of developmental biology
Here is the 2025 embryo alphabet from alligator to zebrafish. Developmental biology is stunning & leads to important discoveries for human medicine. @socdevbio.bsky.social
Welcome to embryo alphabet. Each day I will post an unannotated embryo image. We begin with the letter A, the next day will be B, etc. Repost this tweet w/an image of a different species embryo & add a quote with name that begins with same letter of the day. Enjoy! @SocDevBiopic.twitter.com/QBdXp8jjFG
First post – starting with letter A. Check out the reposts for images of other embryos that begin with a letter A and subsequent posts for the rest of the alphabet. (No Ratings Yet) Loading...
To accompany the Biologists @ 100 conference, we are launching the Node–FocalPlane image competition.
Enter your best biological research images for your chance to win £250. All the shortlisted images will be presented in our gallery at Biologists @ 100 at ACC Liverpool, 24-27 March 2025 and on the Node and FocalPlane.
Voting will begin the week before the conference and will continue until Thursday 27 March, when the winner will be announced at the conference and online.
Entries are open to all researchers whether you are attending Biologists @ 100 or not.
Deadline for submissions: 24 February 2025
Registration for Biologists @ 100 is open until 28 February. Join us in Liverpool for the chance to see your image displayed in our gallery! The programme overview is available here and the details of the cell and developmental biology track can be found here.
Competition details:
Email your image to thenode@biologists.com with ‘Biologists @ 100 image competition’ in the subject line.
You can submit up to three biological scientific research images that fall within the scope of The Company of Biologists‘ journals.
In the email, include a description of the image and imaging modality used to acquire the image or software used to reconstruct or analyse it.
We’ll require a high-resolution version if you image is shortlisted. You can submit downsampled images for the initial selection.
There is no theme and no restriction content-wise; it can be a raw, reconstructed, filtered or analysed image of any type of biological sample.
Deadline for image submission is 24 February 2025.
Submitted images should not have already been published elsewhere unless under a CC-BY license and should not have been submitted in another image competition.
One first place prize – £250 and two runners up prizes – £125
Image credit: Antara Chakraborty (No Ratings Yet) Loading...
Luuli N. Tran, Ashwini Shinde, Kristen H. Schuster, Aiman Sabaawy, Emily Dale, Madalynn J. Welch, Trevor J. Isner, Sylvia A. Nunez, Fernando García-Moreno, Charles G. Sagerström, Bruce H. Appel, Santos J. Franco
C. Schwayer, S. Barbiero, D. B. Brückner, C. Baader, N. A. Repina, O. E. Diaz, L. Challet Meylan, V. Kalck, S. Suppinger, Q. Yang, J. Schnabl, U. Kilik, J. G. Camp, B. Stockinger, M. Bühler, M. B. Stadler, E. Hannezo, P. Liberali
Marek J. van Oostrom, Yuting I. Li, Wilke H. M. Meijer, Tomas E. J. C. Noordzij, Charis Fountas, Erika Timmers, Jeroen Korving, Wouter M. Thomas, Benjamin D. Simons, Katharina F. Sonnen
Shariq S. Ansari, Miriam E. Dillard, Mohamed Ghonim, Yan Zhang, Daniel P. Stewart, Robin Canac, Ivan P. Moskowitz, William C. Wright, Christina A. Daly, Shondra M. Pruett-Miller, Jeffrey Steinberg, Yong-Dong Wang, Taosheng Chen, Paul G. Thomas, James P. Bridges, Stacey K. Ogden
Iona G. Thelwall, Carola M. Morell, Dominika Dziedzicka, Lucia Cabriales, Andrew Hodgson, Floris J.M. Roos, Louis Elfari, Ludovic Vallier, Kevin J. Chalut
Eunah Chung, Fariba Nosrati, Mike Adam, Andrew Potter, Mohammed Sayed, Benjamin D. Humphreys, Hee-Woong Lim, Yueh-Chiang Hu, S. Steve Potter, Joo-Seop Park
Rui Yan, Ludwig A. Hoffmann, Panagiotis Oikonomou, Deng Li, ChangHee Lee, Hasreet Gill, Alessandro Mongera, Nandan L. Nerurkar, L. Mahadevan, Clifford J. Tabin
Indhujah Thevarajan, Maria F. Osuna, Sonia Fuentes Lewey, Eustolia Sauceda, Sayra Briseno, Caylah Griffin, Bareun Kim, R. Grant Rowe, Edroaldo Lummertz da Rocha, Jihan K Osborne
Meghana S. Oak, Marco Stock, Matthias Mezes, Tobias Straub, Antony M. Hynes-Allen, Jelle van den Ameele, Ignasi Forne, Andreas Ettinger, Axel Imhof, Antonio Scialdone, Eva Hörmanseder
Anita Adami, Raquel Garza, Patricia Gerdes, Pia A. Johansson, Fereshteh Dorazehi, Symela Koutounidou, Laura Castilla-Vallmanya, Diahann A.M. Atacho, Yogita Sharma, Jenny G. Johansson, Oliver Tam, Agnete Kirkeby, Roger A. Barker, Molly Gale-Hammell, Christopher H. Douse, Johan Jakobsson
Alexander Stanton, Selcan Aydin, Daniel A. Skelly, Dylan Stavish, Kim Leonhard, Seth Taapken, Erik McIntire, Matthew Pankratz, Anne Czechanski, Tenneille Ludwig, Ted Choi, Steven P. Gygi, Ivana Barbaric, Steven C. Munger, Laura G. Reinholdt, Martin F. Pera
Rajini Chandrasegaram, Antony M. Hynes-Allen, Beitong Gao, Abhilesh Dhawanjewar, Michele Frison, Stavroula Petridi, Patrick F. Chinnery, Hansong Ma, Jelle van den Ameele
Valentina Sica, Jacob G Smith, Oleg Deryagin, Eva Andres, Vera Lukesova, Mirijam Egg, Nina Cabezas-Wallscheid, Salvador Aznar Benitah, Antonio L. Serrano, Eusebio Perdiguero, Pura Muñoz-Cánoves
Yuanyuan Qin, Parth Chhetri, Elizabeth Theusch, Grace Lim, Sheila Teker, Yu-Lin Kuang, Shahrbanoo Keshavarz Aziziraftar, Mohammad Hossein Mehraban, Antonio Munoz-Howell, Varun Saxena, Dounia Le Guillou, Aras N. Mattis, Jacquelyn J. Maher, Marisa W. Medina
Sarah Stucchi, Lessly P. Sepulveda-Rincon, Camille Dion, Gaja Matassa, Alessia Valenti, Cristina Cheroni, Alessandro Vitriolo, Filippo Prazzoli, George Young, Marco Tullio Rigoli, Martina Ciprietti, Benedetta Muda, Zoe Heckhausen, Petra Hajkova, Nicolò Caporale, Giuseppe Testa, Harry G. Leitch
F. Soares-da-Silva, G. Nogueira, Marie-Pierre Mailhe, L. Freyer, A. Perkins, S. Hatano, Y. Yoshikai, P. Pereira, A. Bandeira, R. Elsaid, E. Gomez-Perdiguero, A. Cumano
Tae Wan Kim, Jinghua Piao, Vittoria D Bocchi, So Yeon Koo, Se Joon Choi, Fayzan Chaudhry, Donghe Yang, Hyein S Cho, Emiliano Hergenreder, Lucia Ruiz Perera, Subhashini Joshi, Zaki Abou Mrad, Nidia Claros, Shkurte Ademi Donohue, Anika K. Frank, Ryan Walsh, Eugene V. Mosharov, Doron Betel, Viviane Tabar, Lorenz Studer
Andrzej Kubiak, Natalia Bryniarska-Kubiak, Mehmet Eren, Kacper Kowalski, Kinga Gawlińska, Patrycja Kwiecińska, Martine Biarnes-Pelicot, Marie Dessard, Jana El Husseiny, Ti-Thien N-Guyen, Pawel Kożuch, Olga Lis, Marta Targosz-Korecka, Pierre-Henri Puech, Krzysztof Szade
Antonella F.M. Dost, Katarína Balážová, Carla Pou Casellas, Lisanne M. van Rooijen, Wisse Epskamp, Gijs J.F. van Son, Willine J. Wetering, Carmen Lopez-Iglesias, Harry Begthel, Peter J. Peters, Niels Smakman, Johan H. van Es, Hans Clevers
Xi Chen, Krishnan Raghunathan, Bin Bao, Elsy Ngwa, Andrew Kwong, Zhongyang Wu, Stephen Babcock, Clara Baek, George Ye, Anoohya Muppirala, Qianni Peng, Michael Rutlin, Mantu Bhaumik, Daping Yang, Daniel Kotlarz, Unmesh Jadhav, Meenakshi Rao, Eranthie Weerapana, Xu Zhou, Jose Ordovas-Montanes, Scott B. Snapper, Jay R. Thiagarajah
Alejandro Roisman, Leonardo Rivadeneyra, Lindsey Conroy, Melissa M. Lee-Sundlov, Natalia Weich, Simon Glabere, Shikan Zheng, Katelyn E. Rosenbalm, Mark Zogg, George Steinhardt, Anthony J. Veltri, Joseph T. Lau, Tongjun Gu, Hartmut Weiler, Ramon C. Sun, Karin M. Hoffmeister
Connor J. Powell, Hani D. Singer, Ashley R. Juarez, Ryan T. Kim, Duygu Payzin-Dogru, Aaron M. Savage, Noah J. Lopez, Steven J. Blair, Adnan Abouelela, Anita Dittrich, Stuart G. Akeson, Miten Jain, Jessica L. Whited
Maria Jassinskaja, Daniel Bode, Monika Gonka, Theodoros I Roumeliotis, Alexander J Hogg, Juan A Rubio Lara, Ellie Bennett, Joanna Milek, Bart Theeuwes, M S Vijayabaskar, Lilia Cabrera Cosme, James L C Che, Sandy MacDonald, Sophia Ahmed, Benjamin A Hall, Grace Vasey, Helena Kooi, Miriam Belmonte, Mairi S Shepherd, William J Brackenbury, Iwo Kucinski, Satoshi Yamazaki, Andrew N Holding, Alyssa H Cull, Nicola K Wilson, Berthold Göttgens, Jyoti Choudhary, David G Kent
Rohan Kulkarni, Chinmayee Goda, Alexander Rudich, Malith Karunasiri, Amog P Urs, Yaphet Bustos, Ozlen Balcioglu, Wenjun Li, Sadie Chidester, Kyleigh A Rodgers, Elizabeth AR Garfinkle, Ami Patel, Katherine E Miller, Phillip G Popovich, Shannon Elf, Ramiro Garzon, Adrienne M Dorrance
Li’ang Yu, Giovanni Melandri, Anna C. Dittrich, Sebastian Calleja, Kyle Palos, Aparna Srinivasan, Emily K Brewer, Riley Henderson, Ciara Denise Garcia, Xiaodan Zhang, Bruno Rozzi, Andrea Eveland, Susan J. Schroeder, David Stern, Aleksandra Skirycz, Eric Lyons, Elizabeth A. Arnold, Brian D. Gregory, Andrew D. L. Nelson, Duke Pauli
Sissy E. Wamaitha, Ernesto J. Rojas, Francesco Monticolo, Fei-man Hsu, Enrique Sosa, Amanda M. Mackie, Kiana Oyama, Maggie Custer, Melinda Murphy, Diana J. Laird, Jian Shu, Jon D. Hennebold, Amander T. Clark
Vanesa Fernández-Majada, Jordi Comelles, Aina Abad-Lázaro, Verónica Acevedo, Anna Esteve-Codina, Xavier Hernando-Momblona, Eduard Batlle, Elena Martinez
Hong Jiang, Emilie Derisoud, Denise Parreira, Nayere Taebnia, Paulo R. Jannig, Reza Zandi Shafagh, Allan Zhao, Congru Li, Macarena Ortiz, Manuel Alejandro Maliqueo, Elisabet Stener-Victorin, Volker M. Lauschke, Qiaolin Deng
Tatsuya Tsukamoto, Ji Kent Kwah, Mark E. Zweifel, Naomi Courtemanche, Micah D. Gearhart, Katherine M. Walstrom, Aimee Jaramillo-Lambert, David Greenstein
Giada Rossignoli, Michael Oberhuemer, Ida Sophie Brun, Irene Zorzan, Anna Osnato, Anne Wenzel, Emiel van Genderen, Andrea Drusin, Giorgia Panebianco, Nicolò Magri, Mairim Alexandra Solis, Chiara Colantuono, Sam Samuël Franciscus Allegonda van Knippenberg, Thi Xuan Ai Pham, Sherif Khodeer, Paolo Grumati, Davide Cacchiarelli, Paolo Martini, Nicolas Rivron, Vincent Pasque, Jan Jakub Żylicz, Martin Leeb, Graziano Martello
Nagham Khouri-Farah, Emma Wentworth Winchester, Brian M. Schilder, Kelsey Robinson, Sarah W. Curtis, Nathan G. Skene, Elizabeth J. Leslie-Clarkson, Justin Cotney
Peter M Luo, Neha Ahuja, Christopher Chaney, Danielle Pi, Aleksandra Cwiek, Zaneta Markowska, Chitkale Hiremath, Denise Marciano, Karen K Hirschi, M Luisa Iruela-Arispe, Thomas J Carroll, Ondine Cleaver
Giada Rossignoli, Michael Oberhuemer, Ida Sophie Brun, Irene Zorzan, Anna Osnato, Anne Wenzel, Emiel van Genderen, Andrea Drusin, Giorgia Panebianco, Nicolò Magri, Mairim Alexandra Solis, Chiara Colantuono, Sam Samuël Franciscus Allegonda van Knippenberg, Thi Xuan Ai Pham, Sherif Khodeer, Paolo Grumati, Davide Cacchiarelli, Paolo Martini, Nicolas Rivron, Vincent Pasque, Jan Jakub Żylicz, Martin Leeb, Graziano Martello

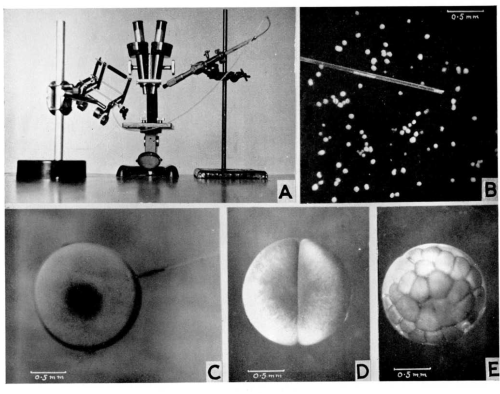
 (3 votes)
(3 votes)
 (No Ratings Yet)
(No Ratings Yet)