We are currently seeking an independent and motivated scientist to join the research group led by Dr Anestis Tsakiridis (https://www.sheffield.ac.uk/cscb/tsakiridis). His work focuses on dissecting the molecular basis of cell fate decisions in the developing embryo and aims to exploit this knowledge for disease-modelling and regenerative medicine applications. The group is particularly interested in the biology of neuromesodermal progenitors (NMPs), the bipotent cell population which drives embryonic axis elongation by generating spinal cord neuroectoderm and paraxial mesoderm.
We have recently described the efficient derivation of NMPs from human pluripotent stem cells (hPSCs) thus establishing a tractable system for studying these progenitors in vitro. The ideal candidate will employ this model in combination with various screens and complementary approaches aiming to define the intrinsic and extrinsic determinants of NMP maintenance and differentiation. Applicants should hold a PhD in a related field and have experience in hPSC culture/differentiation, immunocytochemistry, high content imaging and chromatin immunoprecipitation. A background in Crispr/Cas9-based genome editing and/or bioinformatics would be highly advantageous.
September was notable for the amount of new jobs posted, from Kansas to Copenhagen to King’s College London, and PhD to tenure-track. Check out our jobs page for the latest opportunities. In case you missed them, here are some of our other monthly highlights:
We continued our ‘People behind the papers’ series with Adam Johnston, who talked about his recent work on mouse digit tip regeneration and his new lab in Canada. We also featured an interview with Lewis Cantley from Disease Models and Mechanisms.
Life beyond the bench
A visual report from the Royal Society early career conference by Scriberian
The European Molecular Biology Laboratory is searching for Group Leaders. EMBL offers a highly collaborative, uniquely international culture. It fosters top quality, interdisciplinary research by promoting a vibrant environment consisting of young, independent researchers with access to outstanding graduate students and postdoctoral fellows.
Group Leader Developmental Biology
at EMBL Heidelberg, Germany
The Developmental Biology Unit studies the development of multicellular
organisms. Research in the unit covers all levels, from the cellular to
the whole organism, and is highly interdisciplinary, combining a wide
range of approaches and innovative techniques, with special emphasis
on quantitative and real-time imaging. Research in the unit is firmly
embedded within the overall EMBL research environment, with extensive
in-house collaborations and support from world-class services,
including the gene core, transgenic, metabolomics and mass-spectrometry
core facilities.
We are seeking outstanding candidates using model organisms to
address fundamental principles of multicellular development across the
entire spectrum of developmental biology. Complementary approaches,
including simplified systems, such as organoid and stem cell systems,
are also welcome. Candidates with strong background and research
using theoretical approaches are also encouraged to apply. There is the
possibility to hire two group leaders in the present call.
The successful candidate should have a strong motivation to work in
the multidisciplinary and collaborative environment of EMBL, grasping
the opportunity to interact with many other research groups. In general,
EMBL appoints group leaders early in their career and provides them
with a very supportive environment for their first independent position to
achieve highly ambitious and original research goals.
EMBL is an inclusive, equal opportunity employer offering attractive
conditions and benefits appropriate to an international research
organisation with a very collegial and family friendly working
environment. The remuneration package comprise a competitive
salary, a comprehensive pension scheme, medical, educational and
other social benefits, as well as financial support for relocation and
installation, including your family, and the availability of an excellent
child care facility on campus. EMBL is committed to achieving
gender balance and strongly encourages applications from women.
Appointment will be based on merit alone.
APPLICATION INSTRUCTIONS
Please apply online through www.embl.org/jobs and include a cover letter, CV
and a concise description of research interests and future research plans.
Please also arrange for 3 letters of recommendation to be emailed directly by
your referees to references@embl.de at the latest by 20 October 2016.
Interviews are planned for 19 to 21 December 2016.
For further information about the position can be obtained from
the Head of Unit Anne Ephrussi (anne.ephrussi@embl.de).
Further details on Group Leader appointments can
be found under www.embl.org/gl_faq.
Our monthly trawl for preprints from developmental biology and related research. Previous posts can be found here.
Preprints roll on, and this month’s selection is a reminder of how developmental biology is based on a variety of model organisms. We’ve got two investigations into zebrafish mesoderm from Deborah Yelon’s lab, quite a bit of cell mechanics (in zebrafish, fly and worm), updates on the underlying cell biology of cell division in c. elegans, a transcriptome for the spiny mouse, and much more besides.
Figure 1C from Robin, Michaux, et al. (Edwin Munro lab), showing dynamics of utrophin and myosin at the one cell stage of the c. elegans embryo
As in the last months, the preprints predominantly came from bioRxiv, but we also found a couple from PeerJ. If we missed anything out, let us know. Happy preprinting!
Landscape of X chromosome inactivation across human tissues. Taru Tukiainen, Alexandra-Chloé Villani, Angela Yen, Manuel A. Rivas, Jamie L. Marshall, Rahul Satija, Matt Aguirre, Laura Gauthier, Mark Fleharty, Andrew Kirby, Beryl B. Cummings, Stephane E. Castel, Konrad J. Karczewski, François Aguet, Andrea Byrnes, GTEx Consortium, Tuuli Lappalainen, Aviv Regev, Kristin G. Ardlie, Nir Hacohen, Daniel G. MacArthur
Nuclear pore-like structures in a compartmentalized bacterium. Evgeny Sagulenko, Amanda Nouwens, Richard I Webb, Kathryn Green, Benjamin Yee, Gary Morgan, Andrew Leis, Kuo-Chang Lee, Margaret K Butler, Nicholas Chia, Uyen Thi Phuong Pham, Stinus Lindgreen, Ryan Catchpole, Anthony M Poole, John A. Fuerst
Contrasting genome dynamics between domesticated and wild yeasts. Jia-Xing Yue, Jing Li, Louise Aigrain, Johan Hallin, Karl Persson, Karen Oliver, Anders Bergström, Paul Coupland, Jonas Warringer, View ORCID ProfileMarco Cosentino Lagomarsino, Gilles Fischer, Richard Durbin, Gianni Liti
Nanopore DNA Sequencing and Genome Assembly on the International Space Station. Sarah L Castro-Wallace, Charles Y Chiu, Kristen K John, Sarah E Stahl, Kathleen H Rubins, Alexa B. R. McIntyre, Jason P Dworkin, Mark L Lupisella, David J Smith, Douglas J Botkin, Timothy A Stephenson, Sissel Juul, Daniel J Turner, Fernando Izquierdo, Scot Federman, Doug Stryke, Sneha Somasekar, Noah Alexander, Guixia Yu, Christopher Mason, Aaron S Burton
This post first appeared on Annotations, the DMDD blog (blog.dmdd.org.uk).
Around a third of targeted gene knockouts in mice are embryonic-lethal. But not all deaths occur during gestation – a significant number of gene knockouts result in death at or shortly after the time of birth. Mice from these knockout lines provide a valuable animal model of human neonatal death and are the focus of a new systematic screen by the DMDD programme.
The study, to be carried out by consortium members at the Wellcome Trust Sanger Institute, will offer new insights into the genetic basis of death in neonates, complementing the efforts of large-scale human gene studies such as the DDD and UK10K.
UNDERSTANDING LETHALITY IN NEONATES
The DMDD programme studies embryonic-lethal knockout mouse lines, where lethal means that no pups are observed 14 days after birth (known as postnatal day P14). Detailed phenotyping of these lines can provide important clues about the genetic basis of human developmental disorders.
Our phenotyping efforts so far have focussed on in-utero development at embryonic days E9.5, E14.5 and E18.5 since, for the majority of embryonic-lethal lines, the embryos die well before birth.
In 5-10% of cases, however, the embryos die at the time of birth or shortly afterwards, and can provide an animal model of neonatal death due to genetic mutation. The pilot neonatal screen will study 20 of these lines during the period E18.5 to P14, gathering systematic phenotype data.
Nesting wild-type pups on their day of birth.
Common causes of lethality for neonates can include problems with partuition, breathing, suckling and achieving homeostasis. To understand the likely cause of death in each case, the neonatal screen will assess the pups’ gross dysmorphology, breathing, milk spot morphology (a test that reveals whether they have been able to feed), weight, measurements, locomotive skills, righting reflex (ability to correct their own body position) and blood glucose levels.
The first data is expected at the end of 2016. For more information about the DMDD programme, visit dmdd.org.uk.
Here are the highlights from the current issue of Development:
Defining digit number in the limb
During vertebrate limb development, multiple mechanisms act to ensure the appropriate number, identity and positioning of digits. In both the chick and the mouse, Sonic hedgehog (Shh) signalling emanating from the posterior polarising region provides positional information and regulates tissue expansion, while a self-organising Turing-type mechanism has been proposed to account for digit number determination. However, we still have an incomplete understanding of how these mechanisms interact, and how they have been manipulated during evolution to generate different species with different numbers and types of digit. On p. 3514, Joseph Pickering and Matthew Towers use temporally defined application of the Shh inhibitor cyclopamine to the chick wing to explore these questions. They find that, surprisingly, Shh inhibition at a specific time point can lead to an increased digit number, as well as increased proliferation of the polarising region. Moreover, they observe posterior expansion of the overlying apical ectodermal ridge (AER) – the source of Fgf signalling that is also key for digit formation. The data suggest that, if Shh signalling is inhibited during this critical period, there is a switch from Shh-dependent anterior-posterior expansion to AER-dependent expansion, with consequent effects on digit number and patterning. Together, these data help to shed light on the evolutionary mechanisms resulting in the divergence in digit patterning across vertebrates.
LEUTX: promoting genome activation in the human embryo
Embryonic genome activation (EGA) is a key event in the development of all organisms – when zygotic transcription takes over from maternally provided mRNAs. Relatively little is known about the control of EGA in human embryos, which occurs at the 4- to 8-cell stage. Juha Kere and colleagues have previously found that many paired-like homeobox transcription factors are expressed in the early human embryo, and identified a 36 bp motif associated with genes upregulated at the EGA. In a follow-up study (p. 3459), the authors now characterise one of these factors, LEUTX, in more detail. LEUTX is specifically expressed in early human embryos and cell lines derived from early blastomeres, with barely detectable or absent expression in later embryos, other tissues or cell lines. The authors then use expression profiling to analyse the consequences of overexpressing LEUTX in human embryonic stem cells. LEUTX induces expression of a large number of targets, including several pluripotency-associated genes. Intriguingly, the previously identified EGA-associated 36 bp motif is significantly enriched in theLEUTX-regulated gene set; in reporter assays, LEUTX can bind this motif and promote expression. A second paired-like homeobox factor, DPRX, is a putative target of LEUTX, and in turn, appears to bind the same motif and repress transcription. The authors therefore propose a two-stage model for EGA – whereby LEUTX-mediated induction followed by DPRX-mediated repression leads to transient expression of key EGA genes.
Polycomb: promoting stem cell differentiation in the germline
The Drosophila ovary is a valuable model for the investigation of niche-stem cell interactions. In this model, BMP (Dpp) signals from the niche cap cells inhibit differentiation of germline stem cells (GSCs), at least partially through repression of the transcription factor bam. Away from the niche, germline cells are in contact with somatic escort cells, which support GSC differentiation and do not express dpp. Several mechanisms have been uncovered that contribute to the regulation of dpp expression and signal transduction, and on p. 3449, Rongwen Xi and co-workers reveal an important role for Polycomb group (PcG)-mediated modulation of dpp. Depletion of Polycomb repressive complex 1 (PRC1) components in escort cells causes aberrant expression of dpp, leading to tumour-like accumulation of GSCs and differentiation failure. This non-autonomous phenotype could be rescued by expression ofbam in the GSCs. PcG usually acts in antagonism to Trithorax group (TrxG) complexes, and the authors further show that knockdown of the TrxG component brm reverses the PcG-depletion phenotype, so GSCs can once again differentiate. Unlike many other cases of PcG-TrxG antagonism, however, brmknockdown or overexpression alone has no effect, suggesting that PcG-mediated dpp repression may be the default state in escort cells. This work adds to our understanding of the mechanisms underlying niche-stem cell interactions and GSC differentiation, and underlines the importance of dynamic chromatin regulation.
PLUS:
Blood stem cells: from beginning to end
In June 2016, scientists from all over the world gathered at EMBL headquarters in Heidelberg, Germany to discuss the recent advances in hematopoietic stem cell research. In their Meeting Review, Anna Bigas andClaudia Waskow summarize the exciting work that was presented and the main themes that emerged from the meeting.
Making sense out of spinal cord somatosensory development
Research over the past couple of decades has identified transcription factor networks that define and instruct the generation of diverse neuronal populations within the spinal cord. A number of studies have now started to connect these developmentally-defined populations with their roles in somatosensory circuits. In their Review, Helen Lai, Rebecca Seal and Jane Johnson review how neuronal diversity in the dorsal spinal cord is generated and discuss the logic underlying how these neurons form the basis of somatosensory circuits.
From today and until Wednesday, 150 scientists from around the world will debate plasticity and cell migration in the new gathering in the Barcelona Biomed Conference Series, organised by IRB Barcelona and the BBVA Foundation.
The 18th conference in this series brings together biologists devoted to the study of development in animal models, specialists in cancer, and paediatricians.
How do paediatric cancers differ from those in adults? What makes a static cell migrate and promote metastasis? How do differentiated cells become tissue repair cells? These are all
A Drosophila melanogaster embryo (credit:IRB Barcelona)
questions that can be answered through the study of cell biology and development in animal models such as flies, mice and zebra fish.
Today, around 20 speakers from around the world and 150 participants will debate cutting-edge research into plasticity and cell migration—two key aspects in paediatric cancer, metastasis, and regeneration. From today and until Wednesday, these scientists will gather at the Institut d’Estudis Catalans on the occasion of the 18th Barcelona Biomed Conference, an event organised by the Institute for Research in Biomedicine (IRB Barcelona), with the support of the BBVA Foundation.
Jordi Casanova, IRB Barcelona scientist and CSIC researcher, and Jean Paul Thiery, from the University of Singapore, co-organisers of the event, which is entitled “Epithelial transitions and cell migration: learning from development to tackle disease”, have brought together scientists working on the transitions that certain static cells undergo to become migratory cells, and vice versa, to cover specific requirements.
The list of guest speakers includes Jaume Mora, a physician-scientist at the Hospital San Joan de Déu specialised in paediatric cancer. “Adult cancers arise as a result of various mutations, but in children, a single mutation can cause cancer by conferring cell plasticity. However, this same property can also allow cancer to go into remission without treatment,” explains Casanova. “This is why research into cell plasticity during development is particularly relevant for physicians working in paediatric cancer,” he adds.
Cell reversibility and movement are also critical for the spread of cancer.“A static cancer cell cannot colonise other organs,” explains Casanova. These cells undergo two transitions: from epithelial cells (static cells) to mobile cells that leave the tumour, and after, the opposite process, whereby the latter anchor and colonise a new organ. Alternatively, the cells remain in an intermediate state, which can be particularly dangerous. A complete understanding of the genetic bases of these transitions is critical to further our knowledge of metastasis in epithelial cancers, such as in breast, lung, skin and colon cancer, among others. Eduard Batlle, ICREA researcher at IRB Barcelona devoted to colon cancer, will be giving a talk on the progression of this kind of tumour.
Other guest speakers include Maria Leptin, director of EMBO, the main organisation representing European biologists, and a specialist in epithelial transitions, and Jean Paul Thiery, who, together with Ángela Nieto, from the Instituto de Neurociencias de Alicante, have laid the groundwork for international research in these fields.
“As cells become specialised, they lose plasticity. But some of them are in fact able to gain plasticity,” explains Casanova. Unravelling the variety and complexity of these processes is one of the key objectives of developmental biology.
Finally, another feature associated with plasticity is regeneration. Using animal models with a high regenerative capacity to study the genetic programmes that are activated to recover lost or damaged tissue will bring about advances in regenerative medicine.
During our research as biologists we endure numerous problems, failures, and setbacks in our efforts to advance scientific knowledge. But sometimes the very problems we encounter are opportunities in disguise. This was my experience as a postdoc studying regeneration in the planaria Schmidtea mediterranea in the lab of Alejandro Sánchez Alvarado at the Stowers Institute for Medical Research.
Planarians are free-living flatworms that have the remarkable capability of regenerating any and all of their tissues throughout their seemingly endless adult life (Elliott and Sánchez Alvarado 2013). The negligible senescence and regenerative capabilities of planarians can be traced to an abundant population of persistent pluripotent stem cells that constantly fulfill the tissue homeostasis and regeneration demands of these organisms. This population of cells has the additional feature of enabling amputated tissue fragments containing at least one pluripotent stem cell to regenerate into an entire clonal organism (Wagner, Wang et al. 2011).
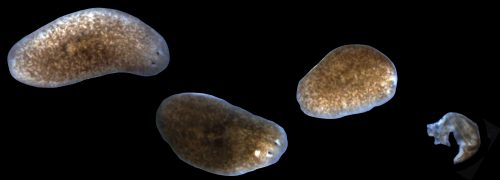
While these properties earned planaria the classic moniker of being “immortal under the edge of the knife” (Dalyell, 1814), our laboratory would receive a potent lesson in planarian mortality. In order to expand our planarian colony, animals were subjected to a vigorous amputation and feeding regimen. Initially this aggressive expansion strategy seemed to be working, but soon it was discovered that the stage had also been set for an equally aggressive bacterial infection. Visible tissue lesions formed and amputated fragments were lysing with increasing frequency (Figure 1). The outbreak transitioned from acute to chronic and in some cases entire boxes of worms amputated one day would be lysed by the next morning. At first, treatment with antibiotics stabilized the colony and kept death at bay, but the propensity for bacteria to evolve antibiotic resistance made this truce a fragile one.
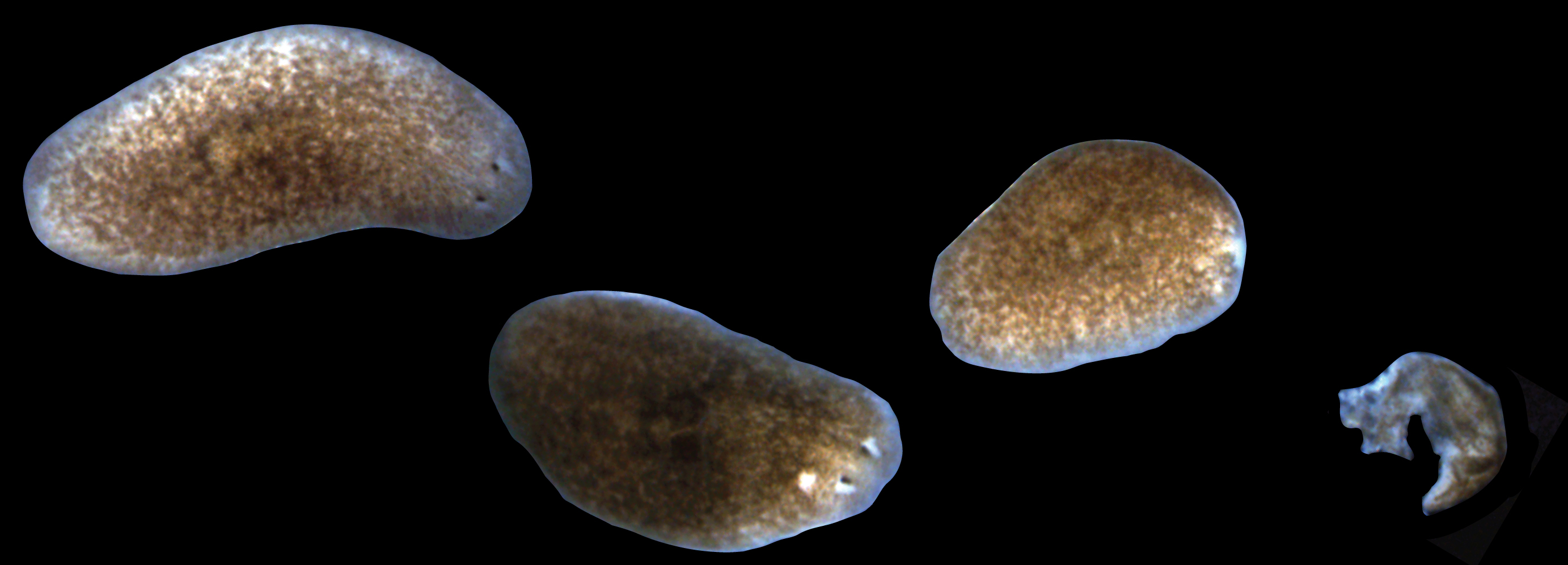
Figure 1. Progression of lesions and tissue resorption in sick planarians. From left to right, a healthy planarian, an animal with a head lesion, an animal that has already degenerated its head, and a planarian that has undergone complete tissue lysis.
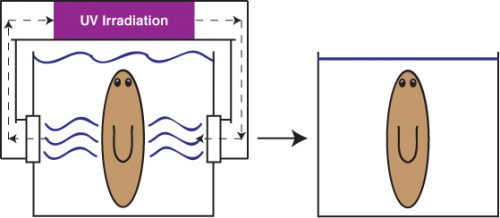
Alejandro sought the assistance of the Stowers Institute for Medical Research Aquatics Facility to develop a long-term solution that could allow us to indefinitely maintain planaria in a stable, low septic environment. The answer was a novel recirculation culture system that utilized constant UV irradiation of recirculating planaria water to prevent uncontrolled bacterial outgrowth (Figure 2). The system was able to rear biomass levels of healthy worms ripe for experimentation. I had just begun my postdoc in the Sánchez lab in the wake of this worm crisis and was excited to receive my first box of worms from this new system.
Figure 2. Diagram of Novel Planarian Recirculation Culture System. Left, planaria reared in low septic culture with constant UV-irradiation of recirculation planaria culture media. Right, planaria housed in traditional static culture.
To my dismay, the very first planarians under my care quickly got sick. As the whole lab would discover, worms withdrawn from the recirculation system were highly susceptible to the development of tissue lesions and degeneration characteristic of sick worms (Figure 1). Just as before, these symptoms could be managed with reduced feedings, antibiotics, and time. Instead of allowing disappointment to gain the upper hand, we saw this as an opportunity to explore the potential role of bacteria and the immune system in animal regeneration.
The role of the immune system in regeneration is an area of intense and ongoing research. Significant evidence spanning a diverse array of organisms and tissues indicated that the immune response can play a central role in either promoting or hindering wound healing and tissue regeneration (Eming, Hammerschmidt et al. 2009, Karin and Clevers 2016). The precise determinants facilitating these diametrically opposed outcomes have important implications in the treatment of chronic non-healing wounds, scar formation, and inflammatory disorders. This reproducible and robust means of inducing tissue degeneration via bacterial infections in planarians struck us as a viable experimental vulnerability to decipher the relationship between microbiota, the immune system, and regeneration in planaria.
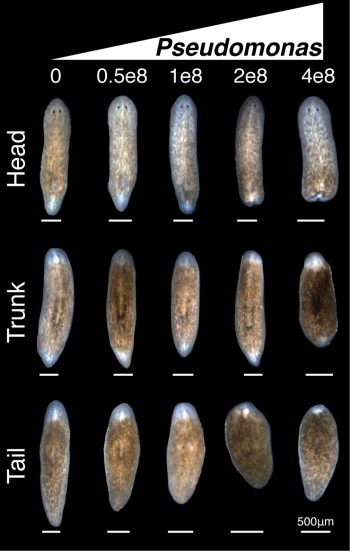
We used a two-pronged strategy of bacterial 16s deep sequencing and plating of planaria homogenate to identify bacterial changes after removal of planarians from the recirculation system. Full analyses of the 16s sequencing was not completed until much later in the project. While the deep sequencing strategy was ongoing, culture of planarian homogenate on LB media plates yielded a strain of Pseudomonas that expanded in worms upon removal from the recirculation system. Infection of healthy worms with Pseudomonas was sufficient to induce the tissue degeneration we observed in sick animals. Furthermore, worms infected with Pseudomonas failed to properly regenerate amputated tissues (Figure 3).
Figure 3. Bacterial infection compromises planarian regeneration. Effects of increasing concentrations of Pseudomonas on head, trunk, and tail fragment regeneration 8 days after infection and 7 days after amputation.
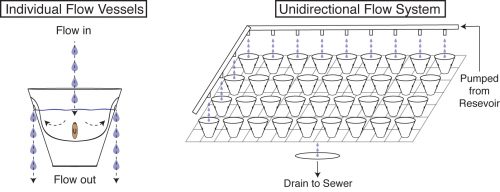
We now had an inducible, highly penetrant, and visibly score-able tissue degeneration phenotype, which combined with the amenability of planaria to RNAi afforded us with the opportunity to dissect genes involved in modulating infection, and to uncover any roles such genes may play in regulating tissue lesions and/or regeneration. Healthy worms directly isolated from the recirculation system were the ideal source to study this process. Unfortunately, our initial attempts were unsuccessful as worms fresh from the recirculation system subjected to the 10 day RNAi feeding protocol developed tissue lesions and degeneration prior to Pseudomonas infection. This presented us with a new challenge: preserving the reduced septic state of the recirculation culture system in a means conducive to the RNAi feeding protocol. To solve this problem, we once again worked with the Aquatics Core of the Stowers. We developed a flow culture system that allows us to house small groups of planaria in individual vessels supplied with a continuous flow of fresh planaria water (Figure 4). This permitted worms removed from the recirculation system to be subjected to the RNAi feeding protocol while preventing the pathogenic bacterial bloom. Worms could then be infected and the results of the gene knockdown could be assayed.
Figure 4. Diagram of Novel Flow Culture System. Left, individual planarian flow culture vessels. Right, integrated unidirectional flow system (planaria water is administered to all cups but depicted only in the left column and upper row to simplify visualization).
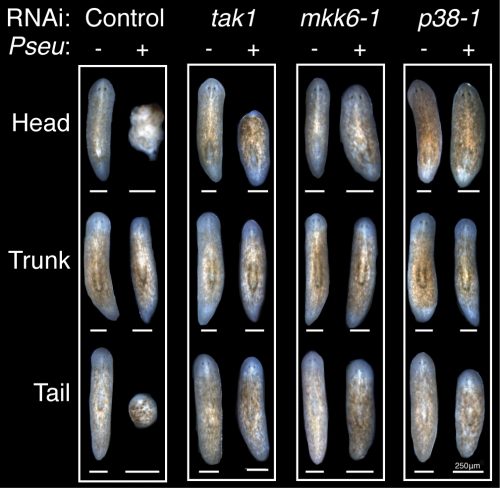
With this new methodology, I performed a candidate screen that quickly identified members of the TAK1/MKK/p38 pathway, a conserved innate immune/inflammatory-signaling module, as the primary mediator of tissue degeneration in response to infection. RNAi of activators of TAK1/MKK/p38 signaling prevented bacterial induced tissue degeneration while RNAi of inhibitors of this signaling pathway accelerated tissue degeneration. Additionally, this innate immune signaling impeded the regeneration of worms during infection, but had no discernible effect on the phenotypic regeneration of uninfected worms (Figure 5).
Figure 5. Conserved TAK1/MKK/p38 pathway components mediate the inhibition of regeneration during infection. Regenerating head, trunk, and tail fragments following control or tak1, mkk6-1, and p38-1 RNAi (16 days post-infection and 14 days post-amputation).
Further molecular analyses revealed a role for TAK1/MKK/p38 in promoting apoptosis in the anterior region of the worm following infection and preceding tissue degeneration. Unexpectedly TAK1/MKK/p38 signaling repressed apoptosis normally coincident with tissue remodeling in un-infected amputated worms. The opposing regulation of apoptosis mediated by TAK1/MKK/p38 in infection versus normal regeneration mirrored the duality of the role of inflammatory signaling in regeneration in other experimental paradigms, suggesting potential cell type specific or context dependent activation underlying its seemingly contradictory roles.
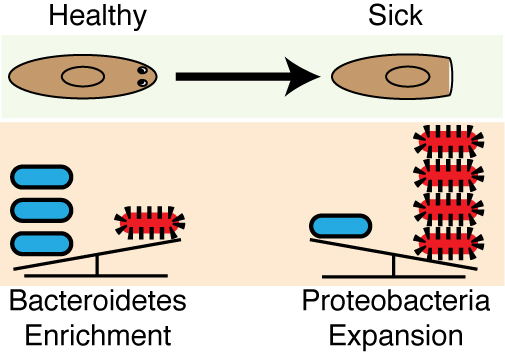
Analyses of the 16s sequencing initiated earlier on in the project was completed and revealed that a high Bacteroidetes to Proteobacteria ratio within the planaria from the recirculation culture system mirrored the distribution of these phyla within the human lower intestinal tract (Human Microbiome Project 2012). This phyla composition is not conserved in other invertebrate (e.g., C. elegans and D. melanogaster) or even vertebrate (e.g., D. rerio) model systems (Lee and Brey 2013, Berg, Stenuit et al. 2016). This was the icing on the cake. The enrichment in Bacteroidetes in planaria was significant as this phylum represents a diverse array of key symbionts known to support proper tissue homeostasis, the development of the immune system, as well as the prevention of inflammatory diseases in mammals, and, intriguingly, have even been linked to the early evolution of multicellular organisms (Rakoff-Nahoum, Paglino et al. 2004, Mazmanian, Liu et al. 2005, Mazmanian, Round et al. 2008, Alegado, Brown et al. 2012). Removal from the recirculation system triggered an expansion in Proteobacteria, including Pseudomonas. Proteobacteria expansion has been linked to a myriad of human inflammatory disorders and coincided with the susceptibility of worms to tissue degeneration (Frank, St Amand et al. 2007, Shin, Whon et al. 2015) (Figure 6). This discovery provided our work with a unique parallel to poorly understood disease-relevant shifts in the human microbiome.
Figure 6. Pathogenic shifts in the planarian microbiome underlie tissue degeneration and impeded regenerative capabilities. Left, bacteria of healthy worms from the recirculation culture are enriched in the phyla Bacteroidetes. Right, following transition to static culture, Proteobacteria rapidly expands coinciding with tissue degeneration.
This work began as a systemic problem of worms becoming sick and evolved into a study linking shifts in endogenous microbiota, the innate immune system, and regeneration. Maybe there are very few problems that we as biologist encounter in our research that can become opportunities. Maybe there are many more. But in the end, it is our attitude as curious scientists that will always help us to ultimately turn lemons into lemonade.
Contributors: Christopher Arnold and Alejandro Sánchez Alvarado
Alegado, R. A., L. W. Brown, S. Cao, R. K. Dermenjian, R. Zuzow, S. R. Fairclough, J. Clardy and N. King (2012). “A bacterial sulfonolipid triggers multicellular development in the closest living relatives of animals.” Elife 1: e00013.
Berg, M., B. Stenuit, J. Ho, A. Wang, C. Parke, M. Knight, L. Alvarez-Cohen and M. Shapira (2016). “Assembly of the Caenorhabditis elegans gut microbiota from diverse soil microbial environments.” ISME J.
Elliott, S. A. and A. Sánchez Alvarado (2013). “The history and enduring contributions of planarians to the study of animal regeneration.” Wiley Interdiscip Rev Dev Biol 2(3): 301-326.
Eming, S. A., M. Hammerschmidt, T. Krieg and A. Roers (2009). “Interrelation of immunity and tissue repair or regeneration.” Semin Cell Dev Biol 20(5): 517-527.
Frank, D. N., A. L. St Amand, R. A. Feldman, E. C. Boedeker, N. Harpaz and N. R. Pace (2007). “Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases.” Proc Natl Acad Sci U S A 104(34): 13780-13785.
Human Microbiome Project, C. (2012). “Structure, function and diversity of the healthy human microbiome.” Nature 486(7402): 207-214.
Karin, M. and H. Clevers (2016). “Reparative inflammation takes charge of tissue regeneration.” Nature 529(7586): 307-315.
Lee, W. J. and P. T. Brey (2013). “How microbiomes influence metazoan development: insights from history and Drosophila modeling of gut-microbe interactions.” Annu Rev Cell Dev Biol 29: 571-592.
Mazmanian, S. K., C. H. Liu, A. O. Tzianabos and D. L. Kasper (2005). “An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system.”Cell 122(1): 107-118.
Mazmanian, S. K., J. L. Round and D. L. Kasper (2008). “A microbial symbiosis factor prevents intestinal inflammatory disease.” Nature 453(7195): 620-625.
Rakoff-Nahoum, S., J. Paglino, F. Eslami-Varzaneh, S. Edberg and R. Medzhitov (2004). “Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis.” Cell 118(2): 229-241.
Shin, N. R., T. W. Whon and J. W. Bae (2015). “Proteobacteria: microbial signature of dysbiosis in gut microbiota.” Trends Biotechnol 33(9): 496-503.
Wagner, D. E., I. E. Wang and P. W. Reddien (2011). “Clonogenic neoblasts are pluripotent adult stem cells that underlie planarian regeneration.” Science 332(6031): 811-816.
Baldwin Wallace University invites applicants for a tenure-track position as an Assistant Professor in Neuroscience with a preferred research focus in electrophysiology using an invertebrate or fish model system beginning August 1, 2017. This position is a joint appointment between the Biology Department and the Neuroscience Program.
Qualified applicants will have completed a Ph.D. in Neuroscience, Physiology or a related discipline and preference will be given to candidates who have teaching experience and postdoctoral research. Teaching responsibilities will include existing courses on Principles of Neuroscience and Neuroscience Methods, as well as newly developed courses on Cell and Molecular Neuroscience, Neurophysiology or other courses related to the candidate’s area of expertise. Start-up funds will be made available to obtain electrophysiology equipment. The successful candidate will be expected to mentor undergraduate research and to seek extramural grants to support neuroscience research.
Candidates will be expected to develop and teach a seminar course for the first year experience program, share in the supervision of Neuroscience senior thesis projects, serve as academic advisors and participate in a shared governance system.
Founded in 1845, Baldwin Wallace University is an independent, coeducational college in the liberal arts tradition. Located 15 miles southwest of downtown Cleveland, Baldwin Wallace enrolls approximately 3,100 full-time undergraduate day students, 800 part-time students in evening and weekend programs, and 700 graduate students.
Candidates should apply online at https://www.bw.edu/employment. A single Word or PDF file containing cover letter, curriculum vitae, teaching philosophy, research statement, and contact information for 3 current references must be uploaded at time of application. Official transcripts will be requested before a campus interview is scheduled. Full consideration will be given to applications received prior to October 21, 2016 and review will continue until the position is filled. All inquiries regarding the position should be directed to Diana Barko, Search Chair (dbarko@bw.edu or 440-826-2489).
Baldwin Wallace University is an EEO/AA employer and educator. At BW, we support and encourage diversity in a variety of forms. We value and appreciate inclusive excellence in the classroom, within extracurricular activities, and as we engage our community. Learn more at https://www.bw.edu/about/diversity/
Researchers working with flies at IRB Barcelona describe that the concentration of some small intracellular organelles determines the branching capacity of tracheal cells.
Tracheal cells are analogous to the cells that form blood vessels in the human body. The inhibition or stimulation of new blood vessels has implications in cancer and in tissue regeneration.
Barcelona, 29th September 2016.- Published today in Current Biology, a study by Sofia J. Araújo, associate researcher at the Institute for Research in Biomedicine (IRB Barcelona), reveals that the number of centrosomes—small intracellular structures—in cells determines the final shape that cells adopt and their function.
The effects of variations in the number of centrosomes are studied mainly in dividing cells because of the key function of centrosomes in this process and their correlation with cancer when defects are present. Araújo’s team, which works in Jordi Casanova’sDevelopment and Morphogenesis in Drosophila Lab, has now addressed the influence of variations in centrosome number on cells that have already divided and differentiated. They demonstrate that centrosomes are also determinant organelles in cells that have already left the cell-division cycle.
The greater the number of centrosomes, the greater the branching in the trachea
The scientists have studied centrosomes in the tracheal cells of the fly Drosophila melanogaster. In the article, they demonstrate that cells that hold more than two centrosomes form more branches (a single cell is like a small tree with many branches). In contrast, those that have two (the usual number) form standard branches, while those that have none show practically no branching.
This finding indicates variations in the number of centrosomes affect the morphology of tracheal cells. In addition, the study describes that the number of centrosomes is related to the first developmental stages of these structures in the fly respiratory system. “Through our work, we have been able to modify the capacity of these cells to branch in function of the number of centrosomes that we introduce. This finding may have biomedical implications,” says Araújo.
Of biomedical interest
Tracheal cells are homologous structures to the cells that form the delicate blood vessels of the human body. The formation of new vessels (from those already existing (angiogenesis) or new ones (neovascularisation) has two implications in human health. On the one hand, medical practice seeks how to inhibit pathological angiogenesis, for example in cancer, when tumour cells generate more blood vessels to facilitate access to more oxygen and nutrients, thus ensuring their growth. On the other hand, and in contrast, it may be of interest to stimulate the formation of new vessels, for example for wound healing or in regenerative processes.
Beyond the widely studied effects of centrosomes on cell division, the work by Araújo provides new information on the function of these organelles and their possible contribution to disease in differentiated cells.
Reference article:
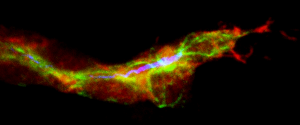
Terminal tracheal cell during the initial ramification. In red, the cells of the trachea, in green the microtubules and in blue the lumen. (Delia Ricolo and Sofia J. Araújo)
Centrosome amplification increases single-cell branching in post-mitotic cells
Delia Ricolo, Myrto Deligiannaki, Jordi Casanova and Sofia J. Araújo
Current Biology (2016) doi: 10.1016/j.cub.2016.08.020
 (1 votes)
(1 votes)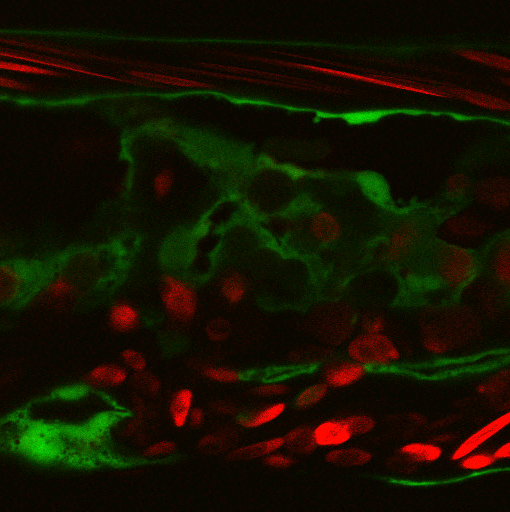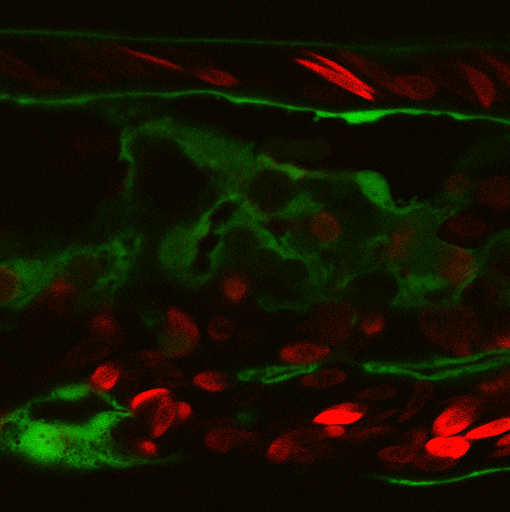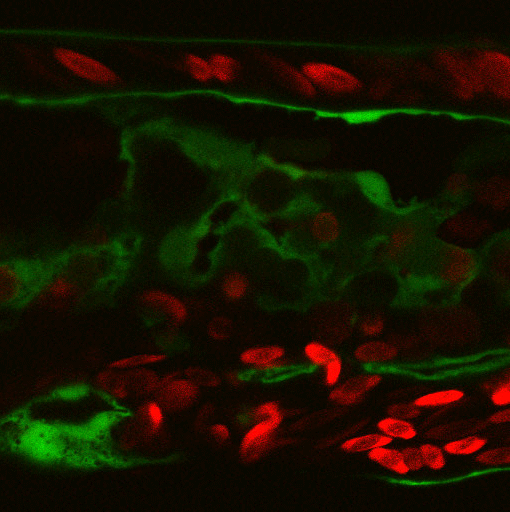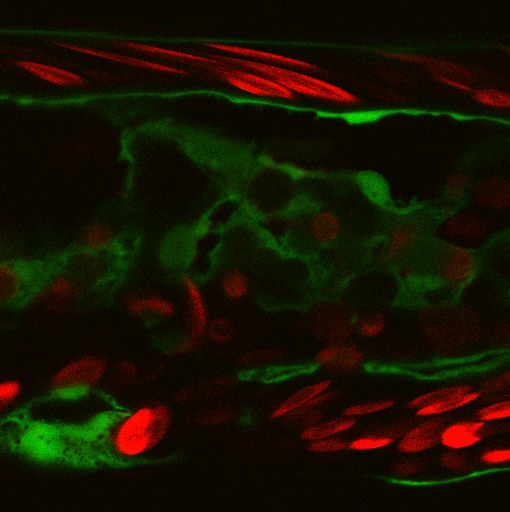


 (No Ratings Yet)
(No Ratings Yet)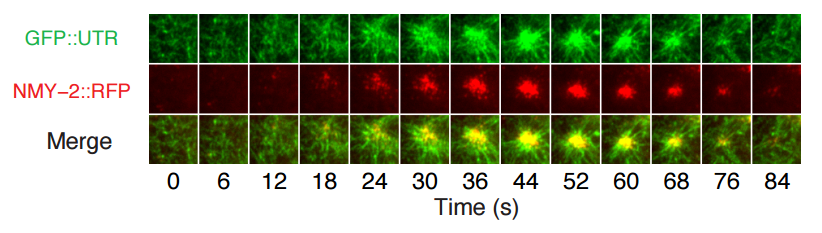




 In June 2016, scientists from all over the world gathered at EMBL headquarters in Heidelberg, Germany to discuss the recent advances in hematopoietic stem cell research. In their
In June 2016, scientists from all over the world gathered at EMBL headquarters in Heidelberg, Germany to discuss the recent advances in hematopoietic stem cell research. In their  Research over the past couple of decades has identified transcription factor networks that define and instruct the generation of diverse neuronal populations within the spinal cord. A number of studies have now started to connect these developmentally-defined populations with their roles in somatosensory circuits. In their
Research over the past couple of decades has identified transcription factor networks that define and instruct the generation of diverse neuronal populations within the spinal cord. A number of studies have now started to connect these developmentally-defined populations with their roles in somatosensory circuits. In their 
 (16 votes)
(16 votes)