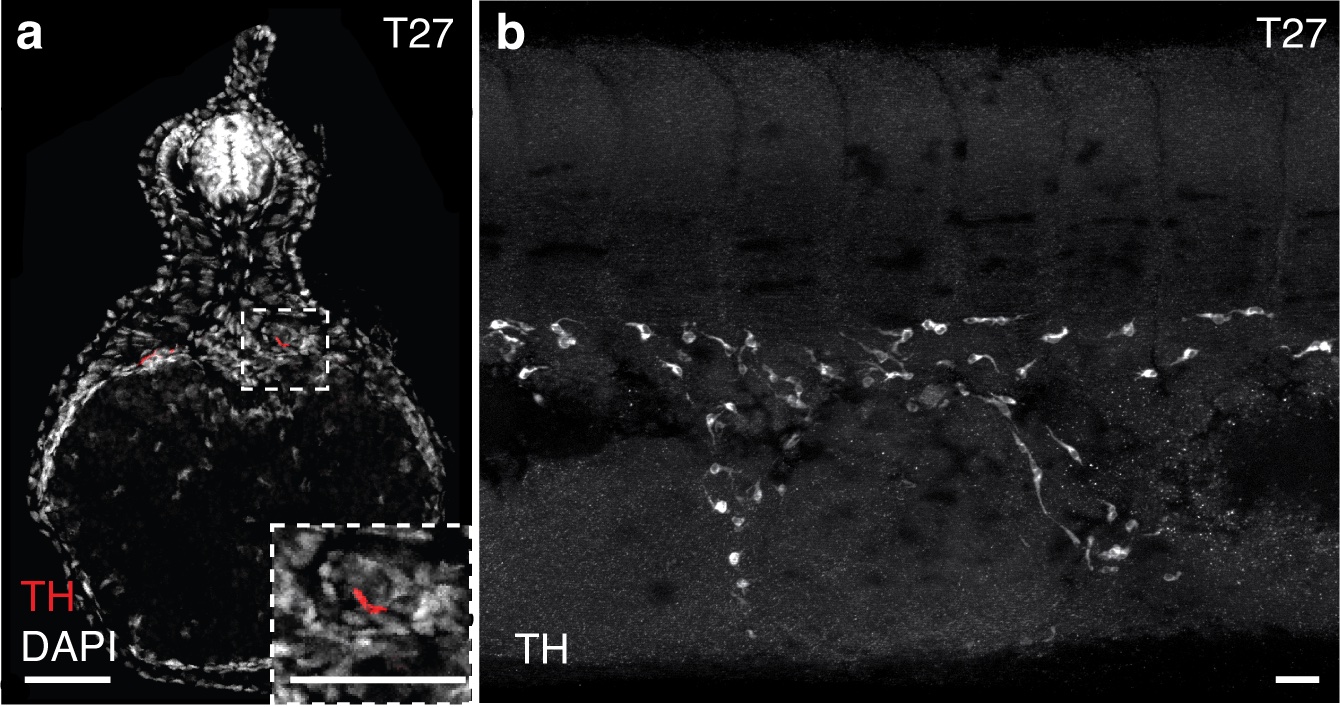
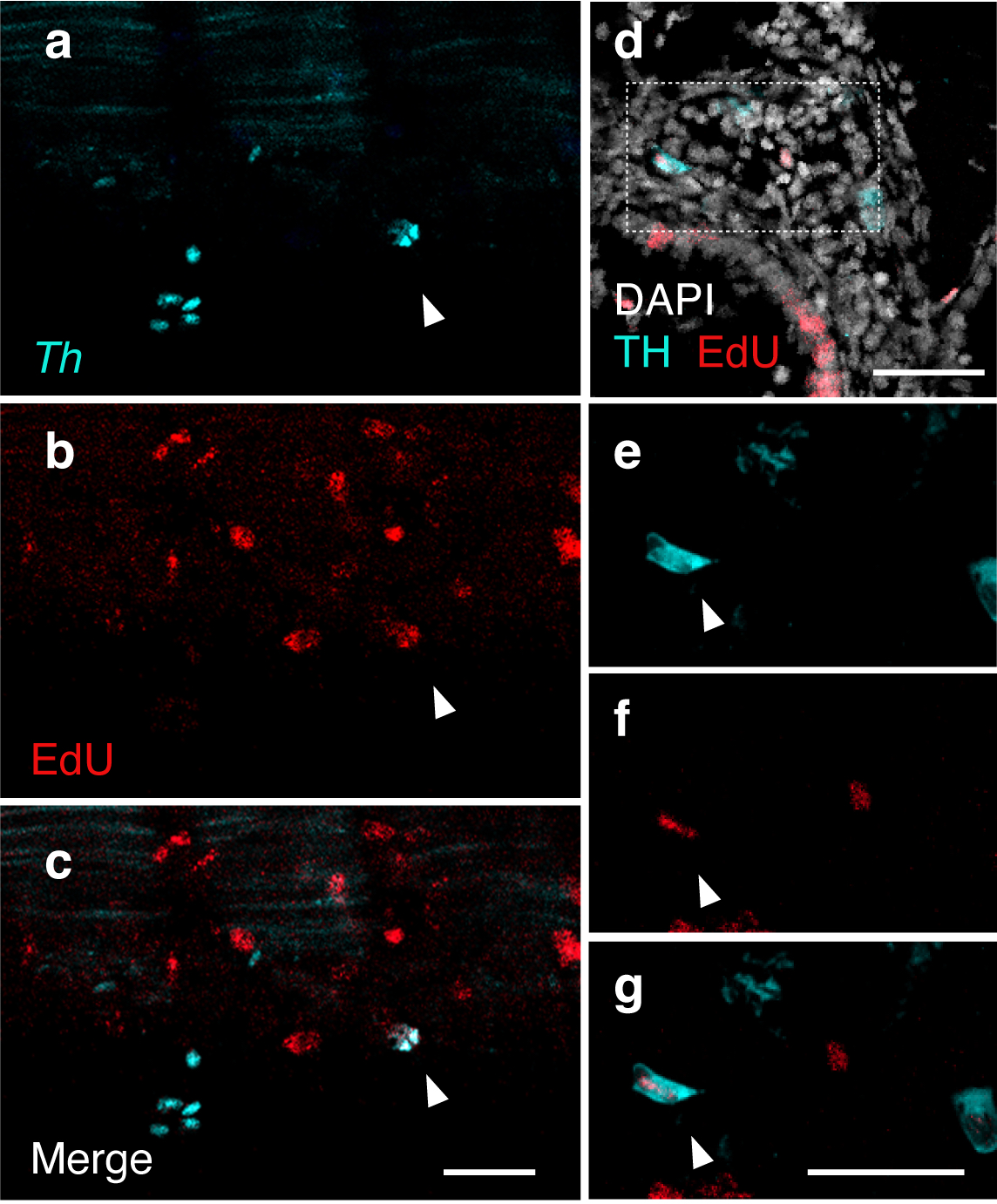
May in preprints
Posted by the Node, on 7 June 2024
Welcome to our monthly trawl for developmental and stem cell biology (and related) preprints.
The preprints this month are hosted on bioRxiv and arXiv – use these links below to get to the section you want:
- Patterning & signalling
- Morphogenesis & mechanics
- Genes & genomes
- Stem cells, regeneration & disease modelling
- Plant development
- Evo-devo
Research practice and education
Developmental biology
| Patterning & signalling
Diffusion barriers imposed by tissue topology shape morphogen gradients
Gavin Schlissel, Miram Meziane, Domenic Narducci, Anders S. Hansen, Pulin Li
Nicole E. Franks, Benjamin L. Allen
Luis Garcia-Alonso
Syeda Nayab Fatima Abidi, Sara Chan, Kerstin Seidel, Daniel Lafkas, Louis Vermeulen, Frank Peale, Christian W. Siebel
Erin Z. Aprison, Svetlana Dzitoyeva, Ilya Ruvinsky
A conserved chronobiological complex times C. elegans development
Rebecca K. Spangler, Guinevere E. Ashley, Kathrin Braun, Daniel Wruck, Andrea Ramos-Coronado, James Matthew Ragle, Vytautas Iesmantavicius, Daniel Hess, Carrie L. Partch, Helge Großhans, Jordan D. Ward
Regulation of brain development by the Minibrain/Rala signaling network
Melissa Brown, Erika Sciascia, Ken Ning, Wesam Adam, Alexey Veraksa
Jmaes Matthew Ragle, Ariela Turzo, Max T. Levenson, Keya Jonnalagadda, Anton Jackson, An A. Vo, Vivian T. Pham, Jordan D. Ward
Phased ERK-responsiveness and developmental robustness regulate teleost skin morphogenesis
Nitya Ramkumar, Christian Richardson, Makinnon O’Brien, Faraz Ahmed Butt, Jieun Park, Anna T Chao, Michel Bagnat, Kenneth Poss, Stefano Di Talia
BMP signalling facilitates transit amplification in the developing chick and human cerebellum
V Rook, P Haldipur, K Millen, T Butts, RJ Wingate
Camille Curantz, Ciara Doody, Helen R Horkan, Gabriel Krasovec, Paris K Weavers, Timothy Q DuBuc, Uri Frank
AMH protects the ovary from doxorubicin by regulating cell fate and the response to DNA damage
Ngoc Minh Phuong Nguyen, Eun Mi Chang, Maeva Chauvin, Natalie Sicher, Aki Kashiwagi, Nicholas Nagykery, Christina Chow, Phoebe May, Alana Mermin-Bunnel, Josephine Cleverdon, Thy Duong, Marie-Charlotte Meinsohn, Dadi Gao, Patricia K. Donahoe, David Pepin
Functional roles of neural aPKCs in mouse brain development and survival
Aicha El Ellam, Emily J. Alberto, Maria E. Mercau, Dimitrius T. Pramio, Krishna M. Bhat, William M Philbrick, Deborah Schechtman, Carla V. Rothlin, Sourav Ghosh
Chordin-mediated BMP shuttling patterns the secondary body axis in a cnidarian
David Mörsdorf, Maria Mandela Prünster, Grigory Genikhovich
LS Ee, D Medina-Cano, CM Uyehara, C Schwarz, E Goetzler, E Salataj, A Polyzos, S Madhuranath, T Evans, AK Hadjantonakis, E Apostolou, T Vierbuchen, M Stadtfeld
Emily Green, Akila Harishchandra, Prabha Ranasinghe, Richard Di Giulio, Nishad Jayasundara
Claire Arata, Sandeep Paul, Simone Schindler, Mathi Thiruppathy, Mackenzie Flath, Dion Giovannone, Zack Hammer, Dev Subramanie, Gage Crump
| Morphogenesis & mechanics
Kei Nagura, Takafumi Ikeda, Takashi Hasebe, Yumeko Satou-Kobayashi, Sumio Udagawa, Shuji Shigenobu, Atsuko Ishizuya-Oka, Masanori Taira
Mechanical signaling through membrane tension induces somal translocation during neuronal migration
Takunori Minegishi, Honami Hasebe, Tomoya Aoyama, Keiji Naruse, Yasufumi Takahashi, Naoyuki Inagaki
Yusuke Mori, Sierra Smith, Jiacheng Wang, Akankshi Munjal
E-cadherin tunes tissue mechanical behavior before and during morphogenetic tissue flows
Xun Wang, Christian M. Cupo, Sassan Ostvar, Andrew D. Countryman, Karen E. Kasza
Annabel May, Katja Röper
Endogenous OptoRhoGEFs reveal biophysical principles of epithelial tissue furrowing
Andrew D. Countryman, Caroline A. Doherty, R. Marisol Herrera-Perez, Karen E. Kasza
Kristen Kurtzeborn, Vladislav Iaroshenko, Tomáš Zárybnický, Julia Koivula, Heidi Anttonen, Darren Brigdewater, Ramaswamy Krishnan, Ping Chen, Satu Kuure
Kathryn Berg, Joshua Gorham, Faith Lundt, Jonathan Seidman, Martina Brueckner
Vascular development of fetal and postnatal neocortex of the pig, the European wild boar Sus scrofa
Eric Sobierajski, Katrin Czubay, Christa Beemelmans, Christoph Beemelmans, Martin Meschkat, Dennis Uhlenkamp, Gundela Meyer, Petra Wahle
Takanobu A. Katoh, Tim Lange, Yoshiro Nakajima, Kenta Yashiro, Yasushi Okada, Hiroshi Hamada
Igor Kondrychyn, Haymar Wint, Liqun He, Christer Betsholtz, Li-Kun Phng
| Genes & genomes
miR214 regulates sex determination through gsdf in zebrafish
N. Wittkopp, A.M. de Jesus Domingues, R.F. Ketting
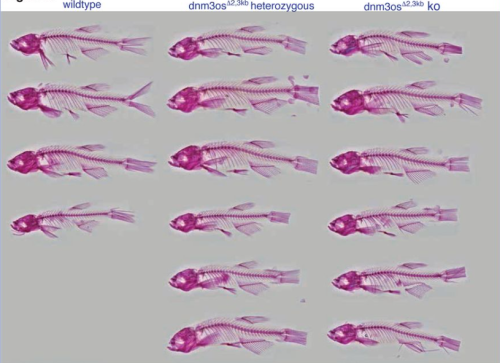
Effects of HMGCR deficiency on skeletal muscle development
Mekala Gunasekaran, Hannah R. Littel, Natalya M. Wells, Johnnie Turner, Gloriana Campos, Sree Venigalla, Elicia A. Estrella, Partha S. Ghosh, Audrey L. Daugherty, Seth A. Stafki, Louis M. Kunkel, A. Reghan Foley, Sandra Donkervoort, Carsten G. Bönnemann, Laura Toledo-Bravo de Laguna, Andres Nascimento, Daniel Natera-de Benito, Isabelle Draper, Christine C. Bruels, Christina A. Pacak, Peter B. Kang
A microRNA that controls the emergence of embryonic movement
Jonathan A. C. Menzies, Andre M. Chagas, Tom Baden, Claudio R. Alonso
Mitochondrial DNA removal is essential for sperm development and activity
Zhe Chen, Fan Zhang, Annie Lee, Michaela Yamine, Zong-Heng Wang, Guofeng Zhang, Chris Combs, Hong Xu
The emerging H3K9me3 chromatin landscape during zebrafish embryogenesis
Katherine L. Duval, Ashley R. Artis, Mary G. Goll
Normal male fertility in a mouse model of KPNA2 deficiency
Franziska Rother, Dalia Abu Hweidi, Enno Hartmann, Michael Bader
Chiemela Ohanele, Jessica N. Peoples, Anja Karlstaedt, Joshua T. Geiger, Ashley D. Gayle, Nasab Ghazal, Fateemaa Sohani, Milton E. Brown, Michael E. Davis, George A. Porter Jr., Victor Faundez, Jennifer Q. Kwong
Nida Ozarslan, Corina Mong, John Ategeka, Lin Li, Sirirak Buarpung, Joshua F. Robinson, Jimmy Kizza, Abel Kakuru, Moses R. Kamya, Grant Dorsey, Phillip J. Rosenthal, Stephanie L. Gaw
Destabilization of mRNAs enhances competence to initiate meiosis in mouse spermatogenic cells
Natalie G. Pfaltzgraff, Bingrun Liu, Dirk G. de Rooij, David C. Page, Maria M. Mikedis
Patterning, regulation, and role of FoxO/DAF-16 in the early embryo
Michael S. Mauro, Sophia L. Martin, Julien Dumont, Mimi Shirasu-Hiza, Julie C. Canman
Sonia Guha, Andrew M. Nguyen, Alejandra Young, Ethan Mondell, Debora B. Farber
Transcriptional Regulators with Broad Expression in the Zebrafish Spinal Cord
Samantha J. England, Paul C. Campbell, Santanu Banerjee, Richard L. Bates, Ginny Grieb, William F. Fancher, Katharine E. Lewis
Shweta Verma, Sujit Dalabehera, Subhash Gowda, Koushika Chandrasekaran, Dayanidhi Singh, Bhavana Prasher, Sharmila Bapat, Sivaprakash Ramalingam, Chetana Sachidanandan
Seth M. Kelly, Allison Paschack, Gargi Mishra, Emma Smith, Katherine Shelmidine, Andre Yazhbin, Young Ji
Haluk Lacin, Yuqing Zhu, Jose T. DiPaola, Beth A. Wilson, Yi Zhu, James B. Skeath
Ke Zhang, Botong Li, Zhiling Deng, Yong Dong, Yuanyuan Li, Bingyu Chen, Ming Zhao, Mao Lu, Xingdong Liu, Zhenhua Guo, Sizhou Huang
Xupeng Chen, Xiaowei Ning, Chenguang Lu, Han He, Yingpeng Yao, Yanli Ni, Jie Zhou, Bing Liu, Siyuan Hou, Yu Lan, Zongcheng Li
Chromatin remodeling protein BPTF regulates transcriptional stability in planarian stem cells
Prince Verma, Alejandro Sánchez Alvarado, Elizabeth M. Duncan
Gita Gajjar, Hayden P. Huggins, Eun Suk Kim, Weihua Huang, Frederic X. Bonnet, Dustin L. Updike, Brett D. Keiper
Irina Lazar-Contes, Rodrigo G. Arzate-Mejia, Deepak K. Tanwar, Leonard C. Steg, Kerem Uzel, Olivier Ulrich Feudjio, Marion Crespo, Pierre-Luc Germain, Isabelle M. Mansuy
The Iroquois (Iro/Irx) homeobox genes are conserved Hox targets involved in motor neuron development
Catarina Catela, Stavroula Assimacopoulos, Yihan Chen, Konstantinos Tsioras, Weidong Feng, Paschalis Kratsios
Histone H2B isoform H2bc27 is expressed in the developing brain of mouse embryos
Saki Egashira, Kazumitsu Maehara, Kaori Tanaka, Mako Nakamura, Tatsuya Takemoto, Yasuyuki Ohkawa, Akihito Harada
Luis Hernandez-Huertas, Ismael Moreno-Sanchez, Jesús Crespo-Cuadrado, Ana Vargas-Baco, Gabriel da Silva Pescador, José M. Santos-Pereira, Ariel A. Bazzini, Miguel A. Moreno-Mateos
| Stem cells, regeneration & disease modelling
Xiexiang Shao, Xingzuan Lin, Hao Zhou, Lili Han, Xin Fu, Sheng Li, Siyuan Zhu, Shenao Zhou, Jianhua Wang, Ping Hu
Andrew S. Hagan, Scott Williams, Casey J. N. Mathison, Shanshan Yan, Bao Nguyen, Glenn C. Federe, Guray Kuzu, Joseph C. Siefert, Janice Hampton, Victor Chichkov, S. Whitney Barnes, Frederick J. King, Brandon Taylor, John R. Walker, Rui Zhao, Jimmy Elliott, Dean P. Phillips, Bin Fang, Rebekah S. Decker
Derivation of human trophoblast stem cells from placentas at birth
Victoria Karakis, John W. Britt, Mahe Jabeen, Adriana San Miguel, Balaji M Rao
Sox5 controls the establishment of quiescence in neural stem cells during postnatal development
Cristina Medina-Menéndez, Lingling Li, Paula Tirado-Melendro, Pilar Rodríguez-Martín, Elena Melgarejo-de la Peña, Mario Díaz-García, María Valdés-Bescós, Rafael López-Sansegundo, Aixa V. Morales
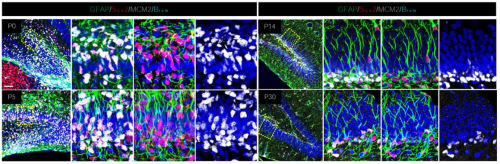
Hiroki Nagai, Yuya Adachi, Tenki Nakasugi, Ema Takigawa, Junichiro Ui, Takashi Makino, Masayuki Miura, Yu-ichiro Nakajima
Generation of spermatogonia from pluripotent stem cells in humans and non-human primates
Eoin C. Whelan, Young Sun Hwang, Yasunari Seita, Ryo Yokomizo, N. Adrian Leu, Keren Cheng, Kotaro Sasaki
Wenqing Li, Sara McCurdy, Miguel A. Lopez-Ramirez, Ho-sup Lee
Amalie Holm Nygaard, Alrik L. Schörling, Zehra Abay-Nørgaard, Erno Hänninen, Yuan Li, Adrian Santoja, Gaurav Singh Rathore, Alison Salvador, Charlotte Rusimbi, Yu Zhang, Agnete Kirkeby
Impaired yolk sac NAD metabolism disrupts murine embryogenesis with relevance to human birth defects
Kayleigh Bozon, Hartmut Cuny, Delicia Z. Sheng, Ella M. M. A. Martin, Paul Young, David T. Humphreys, Sally L. Dunwoodie
Marloes Blotenburg, Lianne Suurenbroek, Vivek Bhardwaj, Peter Zeller
Zhixin Ma, Wenshu Wang, Xiaojing Yang, Menglong Rui, Su Wang
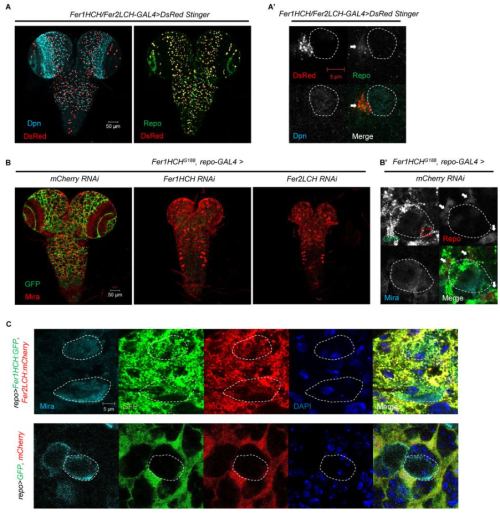
Cell Competition Eliminates Aneuploid Human Pluripotent Stem Cells
Amanda Ya, Chenhui Deng, Kristina M. Godek
Elizabeth Elder, Anthony Lemieux, Lisa-Marie Legault, Maxime Caron, Virginie Bertrand-Lehouillier, Thomas Dupas, Noël Raynal, Guillaume Bourque, Daniel Sinnett, Nicolas Gévry, Serge McGraw
Srivatsava Viswanadha, Manuel Gómez-González, Celine Labouesse, Valeria Venturini, Xavier Trepat, Kevin Chalut, Zanetta Kechagia, Pere Roca-Cusachs
Embryonic diversification of adult neural stem cells and ependymal cells
Shima Yamaguchi, Takaaki Kuniya, Hanae Omiya, Yutaka Suzuki, Masahide Seki, Hideki Ukai, Lingyan Fang, Yujin Harada, Daichi Kawaguchi, Yukiko Gotoh
Archana Kamalakar, Brendan Tobin, Sundus Kaimari, M. Hope Robinson, Afra I. Toma, Timothy Cha, Samir Chihab, Irica Moriarity, Surabhi Gautam, Pallavi Bhattaram, Shelly Abramowicz, Hicham Drissi, Andrés J. García, Levi B. Wood, Steven L. Goudy
Samuel R. Alper, Deeptha Vasudevan, Maya K. Wheeler, Samin Panahi, Richard I. Dorsky
Jonathan M. LaCombe, Kourtney Sloan, Jared R. Thomas, Matthew P. Blackwell, Isabella Crawford, Joseph M. Wallace, Randall J. Roper
BMP signaling promotes heart regeneration via alleviation of replication stress
Mohan Dalvoy Vasudevarao, Denise Posadas Pena, Michaela Ihle, Chiara Bongiovanni, Pallab Maity, Simone Redaelli, Kathrin Happ, Dominik Geissler, Hossein Falah Mohammadi, Melanie Rall-Scharpf, Chi-Chung Wu, Arica Beisaw, Karin Scharffetter-Kochanek, Gabriele D’Uva, Lisa Wiesmüller, Gilbert Weidinger
Direct lineage conversion of postnatal mouse cortical astrocytes to oligodendrocyte lineage cells
Justine Bajohr, Erica Y. Scott, Arman Olfat, Mehrshad Sadria, Kevin Lee, Maria Fahim, Hiba T. Taha, Daniela Lozano Casasbuenas, Ann Derham, Scott A. Yuzwa, Gary D. Bader, Maryam Faiz
Anish Bose, Keaton Schuster, Chandril Kodali, Surabhi Sonam, Rachel Smith-Bolton
Anna Kasprzyk-Pawelec, Mingjun Tan, Raneen Rahhal, Alec McIntosh, Harvey Fernandez, Rami Mosaoa, Lei Jiang, Gray W. Pearson, Eric Glasgow, Jerry Vockley, Christopher Albanese, Maria Laura Avantaggiati
| Plant development
Transit amplifying cells balance growth and differentiation in above-ground meristems
Jessica Joossens, Denia Herwegh, Reinout Laureyns, Julie Pevernagie, Tom Van Hautegem, Lotte Pollaris, Samik Bhattacharya, Christian Korfhage, Thomas Depuydt, Kirin Demuynck, Klaas Vandepoele, Yvan Saeys, Clinton Whipple, Josh Strable, Hilde Nelissen
Dual role of BdMUTE during stomatal development in the model grass Brachypodium distachyon
Roxane P. Spiegelhalder, Lea S. Berg, Tiago D. G. Nunes, Melanie Dörr, Barbara Jesenofsky, Heike Lindner, Michael T. Raissig
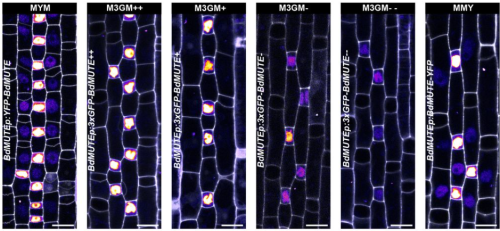
The major nucleoid-associated protein WHIRLY1 promotes chloroplast development in barley
Karin Krupinska, Jürgen Eirich, Urska Repnik, Christine Desel, Monireh Saeid Nia, Anke Schäfer, Ulrike Voigt, Bationa Bennewitz, Wolfgang Bilger, Iris Finkemeier, Götz Hensel
Enric Bertran Garcia de Olalla, Gabriel Rodriguez-Maroto, Martina Cerise, Alice Vayssieres, Edouard Severing, Yaiza Lopez-Sampere, Kang Wang, Sabine Schaefer, Pau Formosa-Jordan, George Coupland
Kumi Matsuura-Tokita, Takamasa Suzuki, Yusuke Kimata, Yumiko Takebayashi, Minako Ueda, Takeshi Nakano, Hitoshi Sakakibara, Akihiko Nakano, Tetsuya Higashiyama
Vikas Garhwal, Sreya Das, Sreeramaiah N Gangappa
James Ronald, Sarah C.L. Lock, Will Claydon, Zihao Zhu, Kayla McCarthy, Elizabeth Pendlington, Ethan J. Redmond, Gina Y.W. Vong, Sanoj P. Stanislas, Seth J. Davis, Marcel Quint, Daphne Ezer
Jae-Hyung Lee, Thu Minh Doan, Sandhya Senthilkumar, Chan Yul Yoo
Arabidopsis ABCC4 encodes a cytokinin efflux transporter and is involved in root system development
Takuya Uragami, Takatoshi Kiba, Mikiko Kojima, Yumiko Takebayashi, Yuzuru Tozawa, Yuki Hayashi, Toshinori Kinoshita, Hitoshi Sakakibara
Simona Crivelli, Kai Bartusch, M. Aguila Ruiz-Sola, Mario Coiro, Signe Schmidt Kjølner Hansen, Elisabeth Truernit
Kang Xu, Haoran Zeng, Feiyang Lin, Emi Yumoto, Masashi Asahina, Ken-ichiro Hayashi, Hidehiro Fukaki, Hisashi Ito, Masaaki K. Watahiki
Mathias Höfler, Xiaomin Liu, Thomas Greb, Karen Alim
Fahad Aldowigh, Rodrigo Matus, Haozhan Gao, Julien Agneessens, Jennifer Topping, Keith Lindsey
Isaia Vardanega, Jan Eric Maika, Edgar Demesa-Arevalo, Tianyu Lan, Gwendolyn K. Kirschner, Jafargholi Imani, Ivan F. Acosta, Katarzyna Makowska, Götz Hensel, Thilanka Ranaweera, Shin-Han Shiu, Thorsten Schnurbusch, Maria von Korff Schmising, Rüdiger Simon
Rasik Shiekh Bin Hamid, Fruzsina Nagy, Nikolett Kaszler, Ildikó Domonkos, Magdolna Gombos, Eszter Molnár, Aladár Pettkó-Szandtner, László Bögre, Attila Fehér, Zoltán Magyar
| Evo-devo
Abdull J. Massri, Alejandro Berrio, Anton Afanassiev, Laura Greenstreet, Krista Pipho, Maria Byrne, Geoffrey Schiebinger, David R. McClay, Gregory A. Wray
Brennan D. McDonald, Abdull J. Massri, Alejandro Berrio Escobar, Maria Byrne, David R. McClay, Gregory A. Wray
Daniel J. Stadtmauer, Silvia Basanta, Jamie D. Maziarz, Alison G. Cole, Gülay Dagdas, Gilbecca Rae Smith, Frank van Breukelen, Mihaela Pavličev, Günter P. Wagner
Haidee Tinning, Alysha Taylor, Dapeng Wang, Anna Pullinger, Georgios Oikonomou, Miguel A. Velazquez, Paul Thompson, Achim Treumann, Peter T. Ruane, Mary J O’Connell, Niamh Forde
Simon Rethemeier, Sonja Fritzsche, Dominik Mühlen, Gregor Bucher, Vera S. Hunnekuhl
Archita Mishra, Niveda Udaykumar, Suvimal Kumar Sindhu, Jonaki Sen
Evolution of the sex-determination gene Doublesex within the termite lineage
Kokuto Fujiwara, Satoshi Miyazaki, Kiyoto Maekawa
Priscila K. F. Santos, Karen M. Kapheim
Sperm competition favours intermediate sperm size in a hermaphrodite
Santhosh Santhosh, Dieter Ebert, Tim Janicke
Nathan D. Harry, Christina Zakas
Anyi Mazo-Vargas, Alan Liang, Brian Liang, Jeanne M.C. McDonald, Arnaud Martin, Robert D. Reed

Olivia Perotti, Gabriel Viramontes Esparza, David S. Booth
Cara Van Der Wal, Shane T. Ahyong, Maxim W.D. Adams, Nathan Lo, Simon Y.W. Ho
Benjamin C. Klementz, Georg Brenneis, Isaac A. Hinne, Ethan M. Laumer, Sophie M. Neu, Grace M. Hareid, Guilherme Gainett, Emily V.W. Setton, Catalina Simian, David E. Vrech, Isabella Joyce, Austen A. Barnett, Nipam H. Patel, Mark S. Harvey, Alfredo V. Peretti, Monika Gulia-Nuss, Prashant P. Sharma
Amelia RI Lindsey, Jason M Tennessen, Michael A Gelaw, Megan W Jones, Audrey J Parish, Irene LG Newton, Travis Nemkov, Angelo D’Alessandro, Madhulika Rai, Nicole Stark
Hiroki Yoshikawa, Yoshiaki Morino, Hiroshi Wada
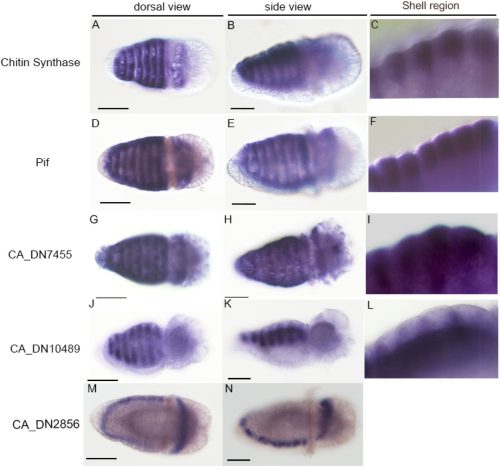
Chhavi Choudhary, Divyanshu Kishore, Keshav Kumar Meghwanshi, Vivek Verma, Jayendra Nath Shukla
Cell Biology
An actomyosin network organizes niche morphology and responds to feedback from recruited stem cells
Bailey N. Warder, Kara A. Nelson, Justin Sui, Lauren Anllo, Stephen DiNardo
Helen Lamb, Małgorzata Liro, Krista Myles, McKenzi Fernholz, Holly Anderson, Lesilee S. Rose
Mohammad Zeeshan, Ravish Rashpa, David J. Ferguson, George Mckeown, Raushan Nugmanova, Amit K. Subudhi, Raphael Beyeler, Sarah L. Pashley, Robert Markus, Declan Brady, Magali Roques, Andrew R. Bottrill, Andrew M. Fry, Arnab Pain, Sue Vaughan, Anthony A. Holder, Eelco C. Tromer, Mathieu Brochet, Rita Tewari
Lisandra Vila Ellis, Jonathan D Bywaters, Jichao Chen
Elevated temperature fatally disrupts nuclear divisions in the early Drosophila embryov
Girish Kale, Pratika Agarwal, J Jaime Diaz-Larrosa, Steffen Lemke
Yan Wu, Yiling Lan, Favour Ononiwu, Abigail Poole, Kirsten Rasmussen, Jonah Da Silva, Abdalla Wael Shamil, Li-En Jao, Heidi Hehnly
Haploidy-linked cell proliferation defects limit larval growth in Zebrafish
Kan Yaguchi, Daiki Saito, Triveni Menon, Akira Matsura, Miyu Hosono, Takeomi Mizutani, Tomoya Kotani, Sreelaja Nair, Ryota Uehara
Chang Zhang, Xiaoqing Sun, Deyi Wu, Guoxia Wang, Hainan Lan, Xin Zheng, Suo Li
Owen H. Funk, Daniel L. Levy, David S. Fay
Differential behavior of pericytes and adipose stromal cells in vasculogenesis and angiogenesis
Julian Gonzalez-Rubio, Hiltrud Königs-Werner, Christian G. Cornelissen, Anja Lena Thiebes
Yuhkoh Satouh, Takaki Tatebe, Isei Tanida, Junji Yamaguchi, Yasuo Uchiyama, Ken Sato
Modelling
Growth regulation bringing modularity to morphogenesis of complex three-dimensional exoskeletons
Hiroshi C. Ito, Yu Uchiumi
How a reaction-diffusion signal can control spinal cord regeneration in axolotls: A modelling study
Valeria Caliaro, Diane Peurichard, Osvaldo Chara
An adaptive numerical method for multi–cellular simulations of tissue development
James M. Osborne
Life history shapes variation in egg composition in the blue tit Cyanistes caeruleus
Cristina-Maria Valcu, Richard A. Scheltema, Ralf M. Schweiggert, Mihai Valcu, Kim Teltscher, Dirk M. Walther, Reinhold Carle, Bart Kempenaers
Stochastic dynamics of two-compartment models with regulatory mechanisms for hematopoiesis
Ren-Yi Wang, Marek Kimmel, Guodong Pang
Waves, patterns and bifurcations: a tutorial review on the vertebrate segmentation clock
Paul François, Victoria Mochulska
Tools & Resources
Alwyn Dady, Lindsay Davidson, Nicolas Loyer, Timothy Sanders, Jens Januschke, Kate G. Storey
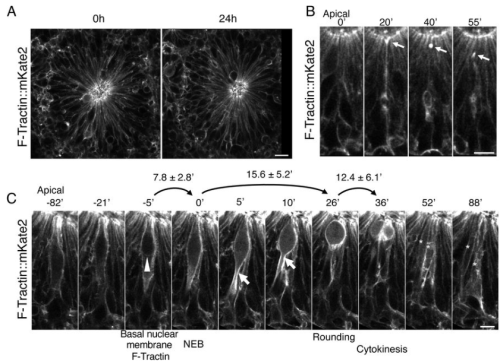
Elizabeth Abraham, Mikel Zubillaga, Thomas Roule, Eleonora Stronati, Naiara Akizu, Conchi Estaras
Nil Üresin, Valdemaras Petrosius, Pedro Aragon-Fernandez, Benjamin Furtwängler, Erwin M. Schoof, Bo T. Porse
Reconstruction of artificial nuclei with nuclear import activity in living mouse oocytes
Nao Yonezawa, Tomoko Shindo, Haruka Oda, Hiroshi Kimura, Yasushi Hiraoka, Tokuko Haraguchi, Kazuo Yamagata
Topological data analysis of pattern formation of human induced pluripotent stem cell colonies
Iryna Hartsock, Eunbi Park, Jack Toppen, Peter Bubenik, Elena S. Dimitrova, Melissa L. Kemp, Daniel A. Cruz
Chungha Lee, Geon Kim, Taeseop Shin, Sangho Lee, Jae Young Kim, Kyoung Hee Choi, Jieun Do, Jaehyeong Park, Jaephil Do, Ji Hyang Kim, YongKeun Park
Engineering an fgfr4 knockout zebrafish to study its role in development and disease
Emma N. Harrison, Amanda N. Jay, Matthew R. Kent, Talia P. Sukienik, Collette A. LaVigne, Genevieve C. Kendall
Dhruv Khatri, Chaitanya A Athale
Yuang Ma, Bo Gou, Yuetong Xu, Muya Shu, Falong Lu, Xiang Li
Barcoding Notch signaling in the developing brain
Abigail Siniscalco, Roshan Priyarangana Perera, Jessie E Greenslade, Aiden Masters, Hannah M Doll, Bushra Raj
Peter Zeller, Marloes Blotenburg, Vivek Bhardwaj, Buys Anton de Barbanson, Fredrik Salmen, Alexander van Oudenaarden
OoCount: A Machine-Learning Based Approach to Mouse Ovarian Follicle Counting and Classification
Lillian Folts, Anthony S. Martinez, Corey Bunce, Blanche Capel, Jennifer McKey
Ashley RG Libby, Tiago Rito, Arthur Radley, James Briscoe
Analysis of developmental gene expression using smFISH and in silico staging of C. elegans embryos
Laura Breimann, Ella Bahry, Marwan Zouinkhi, Klim Kolyvanov, Lena Annika Street, Stephan Preibisch, Sevinç Ercan
Valentina Gandin, Jun Kim, Liang-Zhong Yang, Yumin Lian, Takashi Kawase, Amy Hu, Konrad Rokicki, Greg Fleishman, Paul Tillberg, Alejandro Aguilera Castrejon, Carsen Stringer, Stephan Preibisch, Zhe J. Liu
Camilla Mapstone, Helen Hunter, Daniel Brison, Julia Handl, Berenika Plusa
Mesendodermal cells fail to contribute to heart formation following blastocyst injection
Biyi Li, Chulan Kwon
Erik A. Ehlers, Kyle N. Klein, Margaret A. Fuqua, Julia R. Torvi, Javier Chávez, Lauren M. Kuo, Jacob McCarley, Jacqueline E. Smith, Gaea Turman, Danielle Yi, Ruwanthi N. Gunawardane, Brock Roberts
Marsupial single-cell transcriptomics provides an atlas of developmental heterochrony
Sergio Menchero, Christopher Barrington, Gregorio Alanis-Lobato, Wazeer Varsally, Kathy K. Niakan, James M. A. Turner
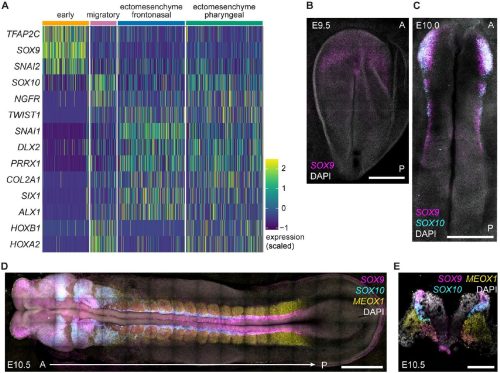
Barbara Varnum-Finney, Adam M. Heck, Sanjay R. Srivatsan, Stacey Dozono, Rachel Wellington, Cynthia Nourigat-McKay, Tessa Dignum, Cole Trapnell, Brandon Hadland
Spatiotemporal profiling of neural crest cells in the common wall lizard Podarcis muralis
Robin Pranter, Nathalie Feiner
Yuting Fu, Xiaoqi Zeng, Yifang Liu, Shikai Jia, Yujia Jiang, Jia Ping Tan, Yue Yuan, Tianchang Xia, Yun Mei, Shan Wen, Xiaojing Liu, Yue You, Weike Pei, Chengshuo Yang, Sida Shao, Saifeng Cheng, Luyi Tian, Longqi Liu, Xiaoyu Wei, Xiaodong Liu
Research practice & education
Jessica Allen, Ekland Abdiwahab, Meghan D. Morris, Claude Jourdan Le Saux, Paola Betancur, K. Mark Ansel, Ryan D. Hernandez, Todd G. Nystul
Building Brains for Robots: A Hands-On Approach to Learning Neuroscience in the Classroom
Raha Kannan, Maribel Gendreau, Alex Hatch, Sydney K. Free, Kithinji Muriungi, Yash A. Garje, Jennifer DeBoer, Gregory J. Gage, Christopher A. Harris
Lauren A. Maggio, Natascha Chtena, Juan Pablo Alperin, Laura Moorhead, John M. Willinsky
Enabling preprint discovery, evaluation, and analysis with Europe PMC
Mariia Levchenko, Michael Parkin, Johanna McEntyre, Melissa Harrison
 (No Ratings Yet)
(No Ratings Yet)







 (3 votes)
(3 votes)