In this interview, we caught up with Indulekha M S, who joined the Indian Society of Developmental Biologists (InSDB) as their Community Manager at the end of 2023.
Indulekha giving a talk about InSDB to the students at TIFR-Hyderabad
What is your background and what made you decide to join the InSDB as their Community Manager?
I did an integrated bachelor’s and master’s of science from the Indian Institute of Science Education and Research (IISER) in Trivandrum, Kerala. I majored in biology and was working in a drosophila genetics lab at the time. Initially, like most of my peers, I wanted to go ahead with a graduate program. But somewhere in the five years of my time at IISE, I got interested in the communications side of science. I had a few stints with a couple of scicomm groups- I worked as an audio editor, produced video interviews, and so on. After graduation, I wanted to do this full-time, and that was when I came across the advert for the community manager position for InSDB. The job description seemed perfect for me, and I applied right away. That’s how I got in.
What does the role of InSDB’s Community Manager entail?
My role is to initiate new activities at InSDB and execute them. I engage with the members and produce stories, interviews, and other forms of content. I also take up new publications in developmental biology and publish lay summaries. Along with this, I manage the InSDB website and social media handles and keep an active online presence. I also coordinate and help organize the InSDB meetings. There’s also a small administrative side to the role: I help take care of the day-to-day functioning of the society. We want to initiate outreach activities that popularize developmental biology, and recently, as a part of this, I visited a few institutes and interacted with the students and faculty there to increase the visibility of InSDB.
Can you give an introduction about the InSDB to the Node readers?
InSDB is the national society that represents researchers in the area of developmental biology field in India. It was formed in the 1970s, and currently, we have members across all career stages, from undergrads to faculties. The society hosts its flagship meeting every two years, and researchers come together and present their work. We had our 2024 meeting in February this year, and the next meeting will be held in 2026. Along with that, we are now planning for smaller, focused meetings and more outreach initiatives. We also have an active website where you can access a variety of resources. When someone becomes a member of InSDB, you get access to the network of developmental biologists and can also avail reduced registration fees for meetings and workshops we organize.
There’s a recent revamp of InSDB’s website. What are the new features and how can researchers make use of the resources on the website?
The new website is a platform made for members to connect with each other and access resources. The site was designed keeping this in mind. One of the key features of the site is that once you become a member, you can ping other members directly from the website. There’s also the forum– a place where you can discuss relevant topics. The forum is your place if you want to discuss a newly published paper or get tips on something. We also publish events – meetings/workshops/journal clubs – and we advertise opportunities in devbio-related fields. The website also has a lot of teaching and learning resources that one can use. We try to add more resources frequently and all the things we curate are to help researchers/students in different ways.
Our audience can also put up events that they are organizing or that they know of. They can also feature Ph.D, postdoc, or internship openings in their labs. We also invite resource submissions that others can make use of.
Are there ways for researchers to contribute to InSDB?
Yes, of course! In fact, we want more people to contribute to InSDB. We welcome articles and perspectives from our readers. Our community blog has some articles written by graduate students. If anyone out there is interested in contributing, they can send a mail to info@insdb.in. It is our members who make the community, so if there is an idea for a new initiative that you would like to see InSDB implement, we are all ears for it.
Any exciting plans/ upcoming projects you’re doing for InSDB?
Yes! We want to do many things at InSDB, but first, we want to connect better with the members and then take InSDB and developmental biology to the more general audience. We have started this by showcasing our members’ stories on the website. You can find these interviews here. As I mentioned before, we want to initiate outreach activities for the non-specialist audience. We are building this up right now. We had our first outreach event of the year at the BLiSc campus in Bangalore, where we had a whole exhibition displayed for high school students. It was a fun event and there’ll be more such events in the future!
Finally, what do you like to do in your free time?
I am new to the city that I currently live in – Bangalore. My weekends are spent roaming around the city and indulging in the local cuisine. If not for that, I am a homebody and spend most of my time reading or trying to find new hobbies.
Indulekha interacting with high school students at the InSDB outreach day (No Ratings Yet) Loading...
While the vertebrate body consists of diverse structures formed during embryogenesis, there is a limited number of genetic regulatory modules that are repurposed in different developmental contexts. For example, the same gene, or group of genes, often play different roles in development of different embryonic structures. That is the case for the hindlimbs and external genitalia – appendages that share many developmental mechanisms and regulatory genes. In our recently published paper, we show that in the absence of Tgfbr1, precursor of external genitalia changes its response to common regulatory signals from genital-specific to limb-specific and adopts limb fate.
Tgf-beta signaling regulates the trunk to tail transition
When I started my PhD project I wanted to study the development of the main body axis. The vertebrate body consists of three main compartments: head, trunk and tail. During embryogenesis these compartments form sequentially, and together with timely transitions between them outline the body plan of an adult organism. I was particularly interested in the trunk to tail transition.
It has long been known that Growth Differentiation Factor 11 (Gdf11) is a key regulator of the trunk to tail transition [1]. In Gdf11 mutant embryos this transition is delayed, resulting in extended trunk length. However, the exact mechanism underlying Gdf11 activity remains unclear. Gdf11 is a signaling molecule of the transforming growth factor beta (Tgf-beta) superfamily and its activity regulating the trunk to tail transition is mediated by binding to a complex of membrane receptors. The partial redundancy with other Tgf-beta ligands has complicated studying molecular mechanism behind Gdf11 [2]. We though that removing one of the membrane receptors could solve this redundancy problem and help us understand genetic regulation of the transition. Previous studies have shown that Transforming growth factor beta receptor 1 (Tgfbr1) mediates Gdf11 activity in the context of trunk to tail transition, so we targeted this receptor and created a mutant mouse line [3], [4].
From the main body axis to appendages
This model helped us uncover many aspects of Tgf-beta regulation of the trunk to tail transition. Normally, during this transition, the embryo stops producing mesoderm associated with the development of internal organs and the body wall and induces caudal body appendages, such as hindlimbs and external genitalia. We showed that Tgfbr1 knock out embryos fail in all these processes [5]. However, due to Tgfbr1’s involvement in heart development and angiogenesis, which results in midgestational lethality, we could only evaluate the early phenotype in Tgfbr1 mutants [6]. To study the effects of Tgfbr1 deficiency at later developmental stages we designed another mouse model where this gene is inactivated after its requirement for heart development. We induced Tgfbr1 deletion in the embryo by administering tamoxifen to pregnant females.
Obtaining late-stage mutant embryos required crossing mice with compound genotypes and optimizing tamoxifen delivery. After months of adjustments in the protocol, we observed the first embryo with a distinct phenotype. It was very fragile and had multiple malformations in the body wall and neural tube, but the most striking feature was the duplication of the hindlimbs. When I showed the mutant to Moises, my supervisor, he said it was one of the most striking phenotypes that he had ever seen in his lab. Of course, we were curious to know how knocking out the Tgf-beta receptor led to the formation of additional hindlimbs. By then, I was already two years into my PhD, but the excitement around this finding made us pivot the project, and I had to learn a lot about limb development.
Phenotype of the Tgfbr1 cKO
Limbs and genitalia share more than we think
To understand the phenotype first we characterized the expression patterns of limb regulatory genes at midgestational stages during early limb bud development. Interestingly, we observed that the hindlimb field in the mutants was extended posteriorly, almost reaching the genital area. The genital primordium, in turn, was underdeveloped in the mutants. This reduction in genital growth could either coincide with hindlimb duplication due to independent regulation of the two structures by Tgfbr1, or alternatively, result from the recruitment of the genital primordium into the limb field, resulting in its development into an additional set of hindlimbs. Hindlimbs and external genitalia precursors are both induced at the trunk to tail transition and, despite developing into morphologically very different appendages, share many regulatory genes [7], [8]. The commonalities between hindlimb and genitalia development prompted us to explore the latter hypothesis. We tried to recreate the activation of limb genes in the pericloacal mesenchyme (the precursor of external genitalia) by overexpression. After generating several transgenics expressing early limb specific genes in the pericloacal mesenchyme, we found that the misexpression of a single gene was insufficient to recapitulate mutant phenotype. We decided to change our approach and focus on the general mechanisms of tissue response. Given that hindlimbs and external genitalia share the expression of so many genes, how are their developmental outcomes so different?
Tgfbr1 guides tissue response by acting on chromatin state
To answer this question, we decided to look at the cis-regulatory regions in the two structures in wild type embryos and Tgfbr1 conditional knock out (cKO). The simplest way to evaluate the activity of the chromatin regions is determining whether they are compacted into nucleosome (inactive) or are nucleosome-free (accessible for transcription factors). We used ATAC-seq, which only generates sequences of open, nucleosome-free chromatin regions [9]. This analysis identified a set of genital-specific regulatory regions that lost accessibility in mutant tissue collected from genital area. The loss of accessibility in these regions could contribute to the inability to activate expression of genes required for genital growth, despite the presence of upstream transcription factors. Further analysis of the mutant extra hindlimb showed that it shared more features with genital samples than with wild type limbs, in line with the hypothesis of its genital origin. Despite that, some chromatin regions in the extra hindlimb acquired limb-type patterns. We believe that gained accessibility in the limb specific regulatory regions of the mutant genital area contributed to its development into an ectopic limb.
Of course, not every nucleosome-free chromatin region is a regulatory region. To validate our findings, we tested several potential regulatory regions using a transgenic reporter assay. To narrow the search, we focused on regions near genes known to be involved in development of limbs and external genitalia. Another criterion for identifying potential regulatory regions was evolutionary conservation. In that way, we identified several regions that drive reporter expression in the genital tubercle (GT) among those that lost accessibility in mutant genital area.
One of the most interesting changes in chromatin was found in a well characterized enhancer driving Gremlin expression in the limb. Gremlin, a secreted Bmp inhibitor, is expressed in response to Sonic Hedgehog (Shh) [10]. Both genes are required for proper limb development [11], [12]. Shh is also a driver of genital growth [13]. However, Gremlin is not expressed in the developing genitalia, despite the presence of Shh signaling. Our results indicate that in wild type genitalia, Gremlin enhancers are inaccessible to Shh regulatory activity. In contrast, in Tgfbr1 cKO embryos, one of the Gremlin enhancers is accessible, leading to ectopic Gremlin activation in the pericloacal mesenchyme. Another regulatory region that we examined in more detail was associated with GT growth and contained binding sequences for the Wnt downstream transcription factor Lef1. By generating sequential deletions of Lef1 binding sequences, we showed that reducing their number decreases reporter activity in the GT. These results illustrate how Tgfbr1 modulates response of limb and genital precursor tissues to common regulatory factors, particularly signaling pathways. This explains how loss of Tgfbr1 leads to a systemic shift in pericloacal mesenchyme response, uncovering its potential to form limbs.
Concluding remarks
The interesting phenotype of our mutant, and (I hope) the novelty of the mechanism we discovered brought a lot of attention to our preprint. Although the manuscript was rejected by two journals, it finally found its place in Nature Communications, which was immensely gratifying. I am very grateful to the reviewers for their comments and suggestions that helped us improve the manuscript.
Unexpectedly for me, the published work received a lot of media attention, likely due to the extraordinary phenotype of the Tgfbr1 cKO. While it was exciting to see my work getting attention, it made me think about the importance of science communication, especially when working with animal models, and even more when conducting basic research. The lack of direct medical application in our research made some people question the justification of our work. This experience underscored the need for clear and effective communication to convey the value and purpose of scientific research to the public.
References
[1] A. C. Mcpherron, A. M. Lawler, and S. Lee, ‘Regulation of anterior / posterior patterning of the axial skeleton by growth / differentiation factor 11’, Nature, vol. 22, no. july, pp. 1–5, 1999.
[2] A. C. McPherron, T. V. Huynh, and S. J. Lee, ‘Redundancy of myostatin and growth/differentiation factor 11 function’, BMC Dev. Biol., vol. 9, no. 1, pp. 1–9, 2009, doi: 10.1186/1471-213X-9-24.
[3] O. Andersson, E. Reissmann, and C. F. Ibáñez, ‘Growth differentiation factor 11 signals through the transforming growth factor-beta receptor ALK5 to regionalize the anterior-posterior axis.’, EMBO Rep., vol. 7, no. 8, pp. 831–7, 2006, doi: 10.1038/sj.embor.7400752.
[4] A. D. Jurberg, R. Aires, I. Varela-Lasheras, A. Nóvoa, and M. Mallo, ‘Switching axial progenitors from producing trunk to tail tissues in vertebrate embryos’, Dev. Cell, vol. 25, no. 5, pp. 451–462, 2013, doi: 10.1016/j.devcel.2013.05.009.
[5] A. Lozovska et al., ‘Tgfbr1 regulates lateral plate mesoderm and endoderm reorganization during the trunk to tail transition’. Mar. 19, 2024. doi: 10.7554/eLife.94290.1.
[6] J. Larsson et al., ‘Abnormal angiogenesis but intact hematopoietic potential in TGF-β type I receptor-deficient mice’, EMBO J., vol. 20, no. 7, pp. 1663–1673, 2001, doi: 10.1093/emboj/20.7.1663.
[7] C. Lin et al., ‘Delineating a Conserved Genetic Cassette Promoting Outgrowth of Body Appendages’, PLoS Genet., vol. 9, no. 1, pp. 1–12, 2013, doi: 10.1371/journal.pgen.1003231.
[8] M. J. Cohn, ‘Development of the external genitalia: Conserved and divergent mechanisms of appendage patterning’, Dev. Dyn., vol. 240, no. 5, pp. 1108–1115, May 2011, doi: 10.1002/dvdy.22631.
[9] J. D. Buenrostro, B. Wu, H. Y. Chang, and W. Greenleaf, ‘ATAC-seq: A Method for Assaying Chromatin Accessibility Genome-Wide’, Curr Protoc Mol Biol, vol. 48, no. 6, pp. 1197–1222, 2016, 10.1002/0471142727.mb2129s109
[10] J. Malkmus et al., ‘Spatial regulation by multiple Gremlin1 enhancers provides digit development with cis-regulatory robustness and evolutionary plasticity’, Nat. Commun., vol. 12, no. 1, p. 5557, Dec. 2021, doi: 10.1038/s41467-021-25810-1.
[11] R. D. Riddle, R. L. Johnson, E. Laufer, and C. Tabin, ‘Sonic hedgehog mediates the polarizing activity of the ZPA’, Cell, vol. 75, no. 7, pp. 1401–1416, Dec. 1993, doi: 10.1016/0092-8674(93)90626-2.
[12] A. Zúñiga et al., ‘Mouse limb deformity mutations disrupt a global control region within the large regulatory landscape required for Gremlin expression’, Genes Dev., vol. 18, no. 13, pp. 1553–1564, Jul. 2004, doi: 10.1101/gad.299904.
[13] C. L. Perriton, N. Powles, C. Chiang, M. K. Maconochie, and M. J. Cohn, ‘Sonic hedgehog Signaling from the Urethral Epithelium Controls External Genital Development’, Dev. Biol., vol. 247, no. 1, pp. 26–46, Jul. 2002, doi: 10.1006/dbio.2002.0668.
We are looking for a motivated, talented and energetic PhD student to join our lab under the supervision of Dr. Benedetta Artegiani and Dr. Delilah Hendriks.
Our lab has expertise and interest in liver and brain biology and their associated diseases.
To this end, in the past years, we have developed 3D human organoid culture systems to grow mini-livers and mini-brains in a dish. We use these models together with state-of-the-art technologies such as for instance CRISPR-Cas9, to model different diseases, such as cancer and genetic disease, and to understand mechanisms underlining the pathogenesis as well as regulating proper organ development.
This PhD position is fully funded (4-years) by a recent grant from the Dutch cancer society (KWF). This project will deal with understanding how chronic liver disease develops into cancer. To read more about the project, visit the link below:
An official post to apply will be opened soon on the institutional webpage, but for informal inquiries we appreciate if you can reach out to us both in a joint email, and reviewing of interesting candidates can start earlier:
On 19 June 2024, Development’s Editor James Wells (Cincinnati Children’s Hospital Medical Center) hosted a Development presents… webinar with three early-career researchers studying mechanics and morphogenesis. Catch up on the recordings of the talks.
Clémentine Villeneuve (Max Planck Institute for Molecular Biomedicine)
The Royal Microscopical Society (RMS) are currently running a Training Needs Survey, which will be live until 30th July, 2024. Fill in the survey here: https://www.surveymonkey.com/r/66TLZQW
In a recent review exercise the RMS established that in 2014 over 30 training courses for different microscopy modalities and techniques were running in the UK. By 2024 this had shrunk to 10. The impact of the Covid-19 pandemic, changes in the funding landscape and steps forwards in Microscopy have all made a profound difference to the skills development landscape.
This 15 minute survey will help the RMS get a better understanding of what scientists who use imaging (including PhD Students, Postdocs, Staff Scientists and Research Technical Professionals and more) are looking for in terms of training and career development. Which techniques would you value learning, how far will you travel, how long would you like the course to be…etc?
We need your input to help shape tomorrow’s microscopy community! Let us know how we can serve you best.
This is an international survey that is open to everyone who uses microscopy for their research, and we are interested in capturing all areas of science. We will publish the results on the RMS website within 6 months.
As an extra incentive there is a prize draw for those who complete the survey with a chance to win a £100 Amazon voucher!
We recently caught up with Delan Alasaadi, who received the 2024 British Society for Developmental Biology (BSDB) Beddington Medal for the best PhD thesis in developmental biology defended in the year previous to the award. Delan gave a talk about his PhD work at the joint BSDB/ Genetics Society Spring Meeting in Warwick in April 2024, and was presented with the Beddington Medal.
Where were you born, and where did you grow up?
I started school in Baghdad, Iraq, where I was born, and continued my studies in Amman, Jordan. My next academic adventure was in Lebanon, where I did my undergraduate degree at the American University of Beirut (AUB).
When did you first become interested in science?
I first became interested in science during my early teenage years. My curiosity about the world around me was growing by the day, and I was constantly puzzled about a wide range of questions like: Where do ideas come from?How can we stimulate more? And how can we decide which notions to follow and which to let go of? Too many questions that involve too many fields like psychology, sociology, science, and more. The underlying theme, regardless of the topic, was this passion for questioning and the attempt to find answers, which became a driving force in my life beyond my “working” hours. The word working is highlighted because if you have an interest in exploring what is around you, beyond your Petri dish or animal model, then do you ever stop?
One direct influence on this way of thinking was the movie “A Beautiful Mind,” which I watched when I was about 15 years old. Avoiding spoiling this marvel of a movie, it initially had a grip on me for eliciting how appealing the outcome of being a scientist was. As I continued my education, this blend of curiosity and inspiration from stories like Nash’s (the main character) drove me to explore various fields of science. Certainly, years later, my view of the movie’s ending changed, and I came to the realization of the importance of grit in the pursuit of what is meaningful knowledge over what is expedient despite obstacles. This mindset has stayed with me, fuelling my continuous quest for knowledge and my love for science.
How did you come to do a PhD with Roberto Mayor at University College London (UCL)?
My undergraduate degree was in biomedical sciences, and my master’s degree at the University of Chester was in differentiation and de-differentiation of macrophages. Despite enjoying doing both, I constantly was asking How (in what) I would develop a niche? And What does that question really constitute? It is easier, saying to myself, to start in a new or emerging field because there is a lot to discover – no matter what, I’ll make it. I was completely wrong, yet the outcome of this approach worked great. That thinking made me choose to explore how physical forces influence the biological behavior of cells. With that interest, the city, and the diverse expertise and pivotal discoveries in mind, I applied to Roberto’s lab. Roberto and his team, throughout decades, specialized in untangling the physical microenvironment and cell signaling pathways that dictate the induction, migration, and differentiation of neural crest cells. The lab has access to a wide range of animal models, cell lines, and biophysical tools, which meant the only limitation was the questions I would ask. I was stunned in my first meeting with Roberto, where we had a well-rounded, simple, down to earth conversation about the possibility of me joining his team. The freedom to express ideas freely and discuss them constructively is a predisposition to any place I aim to be in – so I joined. One week into my PhD, Roberto taught my first fundamental lesson; he said: “The frog is the boss; we will ask the frog if our question is right or wrong.” I knew I was in the right place to get it done.
Your recent NCB paper summarises your main PhD project on neural crest competence. How did the project get started?
This project started when I was around ten years old. One of Roberto Mayor’s students was investigating the resilience of neural crest cells in a Petri dish. The student put a small glass (coverslip) on top of these cells, and to their surprise, the neural crest cells did not die, yet they did not retain their identity either. Although this was a fascinating result, the cause behind this outcome remained unclear – it got shelved. Two decades later, I joined Roberto’s lab and decided to revisit this phenomenon. The immediate hypothesis was that the coverslip applied pressure to the neural crest cells (a mechanical stimulus), changing their nature and preventing them from retaining their neural crest identity—so many questions to answer and so many avenues we could have taken. One question came right at the top of the list: Does this happen in vivo as well? If so, How does that actually happen? Given that neural crest cells in developing embryos are exposed to hydrostatic pressure generated from a liquid-filled cavity (blastocoel cavity), I explored hydrostatic pressure’s role on neural crest cells, an emerging mechanical stimulus in the field of biophysics.
The work involved a wide range of techniques – do you have a favourite?
It has to be inflation and deflation. These two techniques involved artificially changing the hydrostatic pressure within the blastocoel cavity by changing its volume. We called aspirating fluid from the blastocoel Deflation and adding fluid Inflation. Although these two assays were a challenge to optimize from scratch to suit our model of choice, the Xenopus embryos, I enjoyed them the most, at the end of the day, it felt like inflating or deflating a balloon – it is OK, arguably a must, to stay in touch with your inner child-like exploratory mode.
Were there any frustrating moments?
Sure! I wonder if there is anyone who embarked on a scientific journey without being utterly and completely frustrated. However, throughout my PhD, I approached these challenging moments with a more unbiased mindset and as objective as possible I can be. In these moments, your choice of the “right” environment is vital – you get to pick the premise of what “right” means here. Thus, I often mention it is important to pick an environment where you can build a story out of the things that have worked, but it is even more important to be in a place where it is ok when things are not going well too, and you can find means of support through these time(s)!
“When the confocal is not confocalling, a day with some struggles.” — Delan
In your medal award talk at the BSDB meeting, you mentioned your collaborative visit to the Netherlands was supported by a travelling fellowship from The Company of Biologists. Can you talk more about your experience of the visit?
In the interview for my PhD, a panellist said: “Congratulations on this step, but do not forget to get out of the lab.” I did not appreciate its significance till I had the chance to collaborate with the Hiiragi group in Utrecht, Netherlands. Indeed, as I mentioned, we were exploring the possibility of hydrostatic pressure’s role on neural crest cells. We needed to measure directly the change (if any) in Blastocoel’s hydrostatic pressure during gastrulation. During my stay, I had the privilege of meeting new people, where we discussed the struggles and triumphs we went through during our PhDs and the current limitations in our understanding of biological systems. Prof. Takashi Hiiragi and Dr. Moghe Prachiti’s expertise and advice throughout this collaboration were crucial for the successful outcome we achieved in this approach. This collaboration would not have been possible without the generous funds from The Company of Biologists. I highly recommend that, if you are reading this, you find your question, apply for funds, and explore beyond what is known – how exciting. Indeed, the science that we conduct at our benches builds our careers, but the science we do in collaboration builds scientific character.
What other project(s) did you work on during your PhD?
It is an exhilarating feeling to see your colleagues’ projects evolve and to have the chance to contribute with a suggestion, idea, or even a figure. I joined my colleague as a co-first author to investigate the role of RanBP1 on neural crest migration and its role in front-rear polarity. After neural crest induction (the focus of my main story), migration of these cells begins a widely studied phenomenon that has been previously linked to different signaling pathways (e.g., Wnt pathway). However, this work explored the possibility of a new player (RanBP1) in regulating this phenomenon by controlling cellular processes such as cell polarity.
Further, I have written a review of the most recent finding on the role of biomechanical cues on cell fate during early development. In this summary, we explore the latest in vitro, ex vivo, and in vivo studies on how mechanical cues interlink with biochemical signaling to promote tissue-specific fate and function of various cells. I found it is necessary to step out from my main work from time to time and come back with a refreshed mindset to tackle the next set of challenges.
If you took one abiding memory with you from your PhD, what would it be?
This is definitely a hard question to answer. There are so many different memories that led to this moment. Indeed, memories like surviving a global pandemic, walking in empty streets of central London, having the entire London tube for myself, the chance to go to the Netherlands, numerous conversations and mentorship from mentors and friends, and so many countless other memories. Simply, I cannot be dishonest to all of these career and character-building experiences by selecting just one!
What have you been working on since you completed your PhD? What’s next for you?
Neil deGrasse Tyson, world-renowned astrophysicist, said, “Ignorance is the natural state of mind for a research scientist … the boundary between what is known and unknown in the universe.” (Astrophysics for People in a Hurry; p32-33). In that boundary is where I aim to be. To this end, I joined the Acton Lab as a postdoctoral fellow, aiming to bridge the gap between our understanding of normal developmental processes and the transition into a pathogenic state. Generally, my interest remains the same: How do cells make their decisions? Can we influence that process for the betterment of our future?
What techniques or areas in developmental biology excite you the most lately?
Arguably, molecular biology spawned from developmental biology, and now we live in an era overwhelmed by the vast amount of information generated daily by this field. This invokes a paralyzing feeling of “Where do we go from here…?” Thus, I am not generally excited about particular techniques; however, I am quite interested in ideas. Indeed, I am very keen to see how developmental biology will piece together the intricate and detailed information we have of a certain cellular behavior to a much wider picture of understanding biological systems. “Chase ideas and not techniques, Delan!” was the second fundamental lesson I learned from Roberto Mayor during my first year as a PhD.
Outside of the lab, what do you like to do?
I did my PhD in London, so there is always something happening and to do! My general interests are photography, tennis, climbing, and swimming. However, my favourite remains a cup of joe and a conversation. So, If you are in town, let’s go for one?!
“When the Lab is not Labbing, a day to reset everything and go at it again!” — Delan
Alasaadi DN, Mayor R. Mechanically guided cell fate determination in early development. Cell Mol Life Sci. 2024 May 30;81(1):242. doi: 10.1007/s00018-024-05272-6. PMID: 38811420; PMCID: PMC11136904.
Alasaadi DN, Alvizi L, Hartmann J, Stillman N, Moghe P, Hiiragi T, Mayor R. Competence for neural crest induction is controlled by hydrostatic pressure through Yap. Nat Cell Biol. 2024 Apr;26(4):530-541. doi: 10.1038/s41556-024-01378-y. Epub 2024 Mar 18. PMID: 38499770; PMCID: PMC11021196.
Barriga EH, Alasaadi DN, Mencarelli C, Mayor R, Pichaud F. RanBP1 plays an essential role in directed migration of neural crest cells during development. Dev Biol. 2022 Dec;492:79-86. doi: 10.1016/j.ydbio.2022.09.010. Epub 2022 Oct 4. PMID: 36206829.
On the 8th-10th of May Disease Models and Mechanisms organised their first journal meeting entitled Preclinical Modelling of Human Disease and Therapy to showcase the latest advances in modelling human genetics diseases.
**Pics free to use**
The MRC National Mouse Genetics Network was a key organiser and participant with Owen Sansom (Director) and Pleasantine Mill (PI in the Congenital Anomalies Cluster) as members of the organising committee and many speakers and poster presenters to highlight the common thread that drives our scientific interests.
The meeting was also another chance to promote the Congenital Anomalies Variant of Uncertain Significance submission portal to the many national and international colleagues interested in human congenital diseases. Read our meeting report for further insights into a great programme and hugely stimulating three days.
This is part of the ‘Lab meeting’ series featuring developmental and stem cell biology labs around the world.
Where is the lab?
We are part of the Biomedical Research Institute, in the main campus of the National Autonomous University of Mexico in Mexico City (Instituto de Investigaciones Biomédicas, Ciudad Universitaria, Universidad Nacional Autónoma de México, UNAM).
Our aim is to integrate morphogenesis from a subcellular to a tissue-scale approach. We address this using the Drosophila respiratory system as model. This tissue forms complex morphologies that exploit basic cellular processes to build many types of tubes. In addition, these tubes interact with a range of tissues with different chemical and physical properties, making it a great system to compile sub- and supracellular mechanisms of development.
The Tissue Interactions Lab. From left to right: Pedro (astral projection), Samantha, Daniel, Berenice, Fernanda, Luis, and Rebeca (also astrally projected… we found it tough to gather everyone at the same time).
Lab roll call
Daniel: I’m the proud leader of this group.
Berenice: I recently joined this incredible lab. I’m academic technician and my participation has consisted of experimentally supporting the projects.
Luis: I’m a PhD student in the lab, part of the Biochemistry program of UNAM. I’m studying how different tissues can interact during embryogenesis to form different functional structures, from a mechanical point of view. As with all the lab members, I use the fruit fly as a model.
Fernanda: Undergraduate student (Biomedical Research), studying the role of protein secretion during tracheal development.
Pedro: In my BSc program in Biomedical Research we do one-year lab rotations. This is my first rotation and I’m studying apoptosis in the tracheal system.
Samantha: I am a Biology BSc graduate and I am currently applying to Master’s programs to officially join the lab. My research interests lie in studying the mechanisms that coordinate the development of the respiratory system with other tissues.
Rebeca: I am a Neuroscience BSc student interested in developmental biology.
Favourite technique, and why?
Daniel: Tough choice! We do a lot of live imaging in the lab and I really love watching, processing and analysing those movies. But I’m also fascinated by electron microscopy and the wonders you find in electron microcraphs.
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Daniel: Non-model organism studies. The diversity of developmental processes in nature is truly fascinating.
How do you approach managing your group and all the different tasks required in your job?
Daniel: I think open communication with all lab members is key; also, an electronic calendar. We actually have a shared calendar where we schedule our meetings. the calendar includes deadlines, confocal bookings, things to buy, etc. So that we are all aware of what’s going on in the lab.
Are there any challenges unique to running a lab in Mexico?
Daniel: We have great talent, and funding from the University is good (at least for a fly lab). The main challenge is the speed of importing strains and reagents. We really have to plan ahead.
What is the best thing about where you work?
Daniel: Our campus is enormous! We have 10 bus lines just to get around. Besides that, our institute has hired many new faculty and we are building a great community among ourselves and with the rest of the labs here.
Berenice: Our campus is always open to the general public, it has a very active cultural life to which everyone has access. Particularly in our Institute, there are many activities that keep us together such as competitions in which the student community participates with different ideas, we all learn a lot, I love that!
Luis: This university has an infinity of opportunities to do whatever you want. Since it is the largest university in Latin America, it has a lot of cultural, sport and social activities to do. Just in our institute, here we have very diverse labs researching very different topics, and we have the opportunity of learning about other’s research with the different seminars organized every week.
Fernanda: The campus is considered a World Heritage site. It offers diverse activities and services to the entire community, including sports, scientific and cultural events, free language courses, etc. Plus, beautiful scenery can be enjoyed when traveling by bus or bike inside the campus. At the institute, our collaborative environment with experts from various fields promotes the development of exciting research projects.
Pedro: The whole campus is really nice; especially I like that it is surrounded by nature and grass to lie on.
Samantha: I haven’t had the opportunity to enjoy the entire campus yet, but from the times I’ve been there, I can say that it’s a big institution. This facilitates collaboration with other labs for mutual support, and the campus provides excellent resources to achieve research goals. Additionally, the university offers a free bus service to travel all around the campus.
Rebeca: I like that laboratories have diverse research lines because it allows us to learn about many scientific topics and participate in outreach events that enrich our research projects.
What’s there to do outside of the lab?
Daniel: Even though Mexico City is huge, I find everything I need just within my neighbourhood: A bouldering gym, good spaces for biking, and above all, amazing restaurants.
Berenice: Near the institute, we have the university cultural zone with a wide variety of entertainment at low-cost, even free: theater, cinema, concerts, museums.
Outside of our campus -located south of Mexico City- is Coyoacán, a space with a lot of history, and Xochimilco, full of traditions and with a lake that you can explore.
Luis: Mexico City is beautiful and it’s not a secret that its population is huge. Because of this you can find any activity or group of people that matches your interests. Personally, I love musical theater and this city has amazing local and international productions. Also, if you want to have fun at night, there are so many options of bars and clubs for everyone.
Fernanda: Mexico City is a place full of museums, theaters, parks, and restaurants. This means you can find all sorts of activities and delicious food to eat. Moreover, these places are open nearly every day and are easily accessible by subway.
Pedro: In general, I like spending my afternoons playing basketball on campus, but overall Mexico City is very diverse and you can find all kinds of activities.
Samantha: There are plenty of activities to enjoy around campus, such as visiting the botanical garden, exploring the museums, relaxing in the green spaces, or taking a long walk. These are just a few examples.
Rebeca: In the university, there are a variety of cultural spaces. You can attend art exhibitions, museums, theater plays, or concerts. Additionally, you can take art or dance classes. Personally, I enjoy learning languages, and the university offers that too. I’m currently learning Italian.
On 5 June 2024, Development’s Deputy Editor Steve Wilson (UCL) hosted a Development presents… webinar with three early-career researchers studying brain development. Catch up on the recordings of the talks.
Vertebrates, including humans, possess a “head” comprising cranial bones, the central nervous system, and sensory organs. It is believed that the emergence of the vertebrate “new head” is closely linked with the evolutionary acquisition of two cell populations: neural crest cells (NCCs) and cranial placode cells. Therefore, understanding the evolutionary origin and history of NCCs and cranial placode cells is crucial for understanding the evolution of vertebrates.
In vertebrate embryos, both NCCs and cranial placode cells arise from the border region between the neural plate and the epidermis. NCCs are unique because they produce not only cell types of ectodermal origin, such as sensory neurons and melanocytes, but also cell types of mesodermal origin, including smooth muscle cells, osteocytes, and chondrocytes.
Ascidians: fascinating model organisms for evolutionary developmental biology
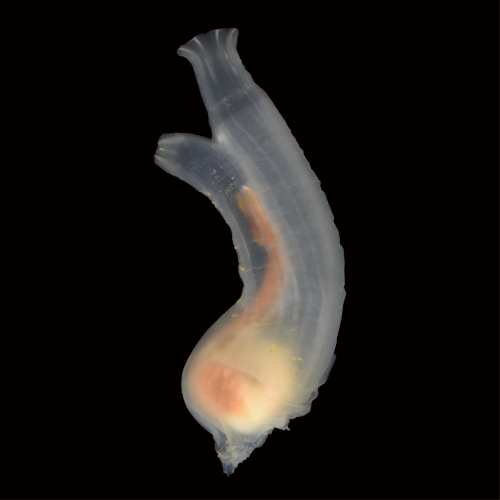
Ascidians, commonly known as sea squirts, belong to the subphylum Urochordata or Tunicata, the sister group of vertebrates. They have been providing key insights into chordate developmental mechanisms and their evolution (FIGURE 1). Recent studies suggested that ascidian embryos have cells that share an evolutionary origin with vertebrate NCCs[1-3]. For example, ascidian cells called a9.49, located in the neural plate border, likely share an evolutionary origin with vertebrate NCCs[1]. Indeed, this cell pair expresses orthologous genes that specify the neural plate border cells and NCCs in vertebrate embryos. Furthermore, a9.49 cells can be reprogrammed to migratory pigment cells by overexpression of Twist, which encodes a transcription factor for mesenchyme specification. However, unlike vertebrate NCCs, ascidian NCC-like cells identified thus far do not produce cell types that are commonly of mesodermal origin. Therefore, it is believed that the multipotency of NCCs has been acquired within the vertebrate lineage after the split from the ascidian lineage.
FIGURE1 The sea squirt Ciona robusta (Ciona intestinalis type A)
A key observation made nearly 40 years ago
In 1987, Nishida found that ascidian cells called b8.17 and b8.19 give rise to muscle cells, nerve cord cells, and endodermal cells near the tip of the tail of embryos[4]. Both b8.17 cells and b8.19 cells are located in the neural plate border, which abuts the neural plate cells that give rise to the central nervous system. These cells express many orthologous genes that specify the neural plate border cells and NCCs in vertebrates. Therefore, if b8.17 and b8.19 cells share an evolutionary origin with vertebrate NCCs and produce cell types that are commonly ectodermal and mesodermal origin, the potential of NCCs to produce cells of multiple germ layers may date back to the last common ancestor (LCA) of vertebrates and ascidians, contrary to the prevailing hypothesis explained above.
In light of this context, we have decided to investigate the possibility that ascidian b8.17 and b8.19 cells share an evolutionary origin with vertebrate NCCs. First, we confirmed that b8.17 cells indeed produced muscle cells, as Nishida showed previously[4]. Second, we showed that these ascidian cells expressed Msx, Zic, Pax3/7, and Snai, which encode orthologs of key transcriptional factors specifying neural plate border cells of vertebrate embryos. We indeed showed that these genes were involved in specifying these ascidian cells. The location and the gene circuit for specification indicate that this ascidian cell population shares an evolutionary origin with vertebrate NCCs.
Do neural plate border cells of ascidian embryos share the evolutionary origin with vertebrate neuromesodermal progenitors (NMPs)?
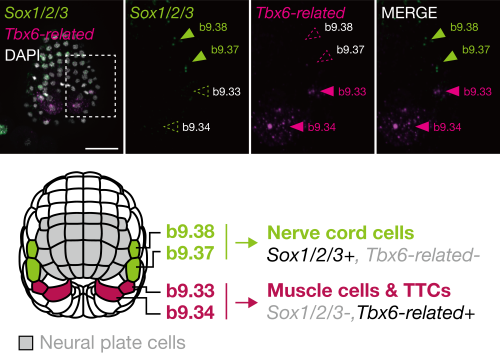
In the middle gastrula stage, the ascidian neural plate border consists of four cells: b9.34, b9.33, b9.37, and b9.38, in order from posterior to anterior. In later embryos, the anterior two cells (b9.37 and b9.38) give rise to nerve cord cells (commonly of ectodermal origin), and the posterior two cells (b9.34 and b9.33) give rise to muscle cells (commonly of mesodermal origin) and other cells near the tip of the tail region[4]. On the basis of this observation, we hypothesized that these cells may share an evolutionary origin with vertebrate neuromesodermal progenitors (NMPs).
In vertebrates, Tbx6 is expressed in NMP-derived mesodermal cells and Tbx6 negatively regulates Sox2, which is expressed in NMP-derived spinal cord cells[5]. If our hypothesis is correct, the gene regulatory circuit consisting of Tbx6 (or its orthologs) and Sox2 (or its orthologs) will also be used for fate decisions in the neural plate border cells of ascidian embryos. Indeed, the anterior cells, which give rise to the nerve cord, expressed Sox2 ortholog (Sox1/2/3), and the posterior cells, which give rise to muscle, expressed Tbx6 ortholog (Tbx6-related) (FIGURE 2). Overexpression of Tbx6-related downregulated Sox1/2/3, and promoted muscle fate. Thus, the ascidian neural plate border cells and vertebrate NMPs share the gene regulatory circuit of Sox2 and Tbx6. In addition, a comparative single-cell transcriptome analysis also supported a close relationship between these ascidian cells and NMPs of zebrafish embryos.
FIGURE2 Gene expression pattern and fate decision of neural plate border of ascidian embryos
In this way, this ascidian cell population has properties of both vertebrate NMPs and NCCs. Therefore, the LCA of tunicates and vertebrates likely had cells with a hybrid property of NCCs and NMPs, and such ancestral cells may have produced both ectodermal and mesodermal cells.
Chordate origin of NCCs and NMPs
A logical follow-up question to ask is whether the Cephalochordata, the sister group of Olfactores, possessed NCC-like cells and NMP-like cells. Cephalochordates, commonly known as lancelets or amphioxus, are filter-feeding marine animals and are believed to retain ancestral features of chordates. Amphioxus is believed to lack cells homologous to vertebrate NCCs[6], although a recent preprint indicated that amphioxus possesses migratory NCC-like cells[7].
Interestingly, somites, notochord cells, dorsal neural tube, and hindgut of the posterior part of amphioxus embryos are produced from a cell population near the tip of the tail[8]. Therefore, amphioxus may possess NMP-like cells. Elucidating the developmental mechanism of this cell population should shed light on the evolution of the body plan of chordates.
A possible evolutionary history of the stemness of NCCs/NMPs
The ascidian NCCs/NMP-like cell population does not possess stemness: they do not show the ability of self-renewal, although they produce cell types that are commonly ectoderm and mesoderm origin. In vertebrates, the high stem cell-like potential of NCCs may depend on pluripotent factors or Yamanaka factors[9-12]. Among Yamanaka factor genes, only Sox1/2/3 was known to be expressed in the ascidian NCCs/NMP-like cells. This may be a reason why the ascidian cells do not have self-renewal ability.
Altogether, we propose a two-step model for the evolution of stemness of NCCs/NMPs: 1) the ability to produce ectodermal and mesodermal cells came first, and 2) the self-renewal ability, which led to acquisition of bona fide NCCs and NMPs. Future works on non-ascidian tunicates (e.g., Oikopleura), amphioxus, and cyclostomes will shed light on the evolution of the stemness of NCCs/NMPs. It would be particularly important to associate the evolution of the stemness of NCCs/NMPs with the evolutionary acquisition of pluripotent factor genes and whole genome duplications that occurred in the vertebrate lineages.


 (No Ratings Yet)
(No Ratings Yet)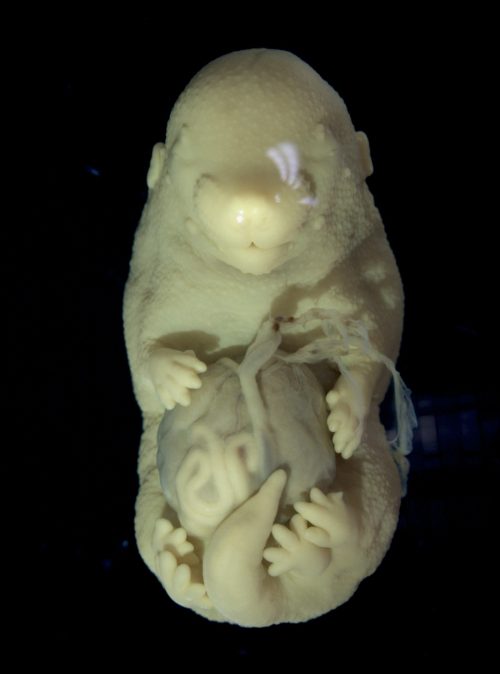
 (1 votes)
(1 votes)