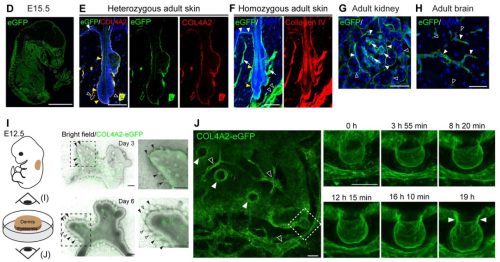
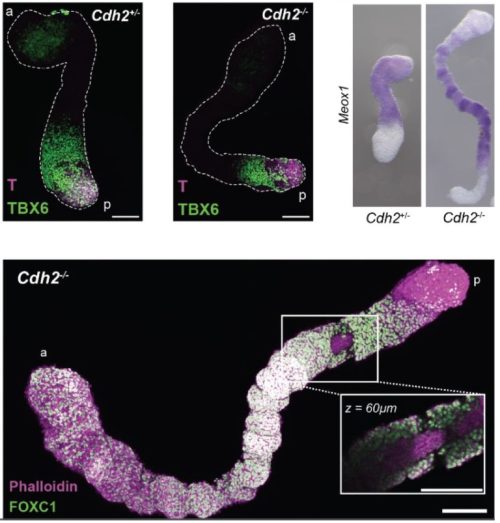
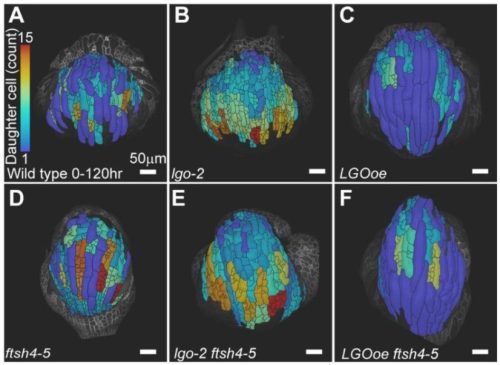
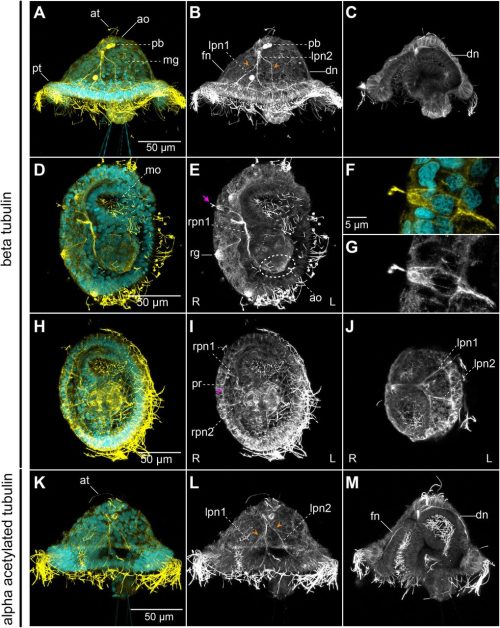
Throughout my years in science, I have been drawn to biological questions across scales and have been struck by how often they reflect aspects of societal phenomena. In this piece, I share with you some of my recent work, and how I view it as a lesson on how reductive or myopic definitions can overlook some of the most impactful discoveries and individuals in a collective.
Like many developmental biologists, I am fascinated by our bodies’ extraordinary cell type diversity. The genetic and epigenetic codes in each type of cell will dictate which unique sets of proteins are expressed. Until recently, the role of a large class of genes, now called micropeptides (or microproteins), was largely overlooked. Protein-coding genes were initially defined using a size cutoff of 100 codons; proteins smaller than that were assumed to not fold properly or carry out functions. Starting in 1990, we realized that this biased definition was filtering out potentially functional genes 1–3. Partnered with technological advances, this shift in mindset has allowed the identification of thousands of small open reading frames (sORFs) that may encode functional tiny proteins.
In recently published work, my colleagues and I set out to investigate whether some previously identified long noncoding RNAs in fact encoded micropeptides 4–6. Many of these RNAs were enriched in developing zebrafish brains and could represent uncharacterized small proteins that play important roles in vertebrate neurodevelopment. If this were the case, the loss of these micropeptides could manifest as behavioral phenotypes, a useful means of screening and prioritization. In this study, we showed that two previously identified long noncoding RNAs actually encode micropeptides with homology to a chromatin regulator found exclusively in vertebrates, called Hmgn1. In humans, this chromatin architectural protein is critically overexpressed in Down syndrome 7, and has been identified as a gene linked to autism 8. Through a series of behavioral, pharmacological, cellular, and molecular assays, we found that when these micropeptides were mutated, the gene regulatory networks that establish cerebellar cells and oligodendrocytes were most significantly affected. Intriguingly, these cell types were recently proposed to have appeared and evolved in jawed vertebrates 9. Is it possible that the emergence of these micropeptides co-evolved with the gene regulatory networks that established cerebellar and oligodendrocyte cell types in vertebrates 10–12? This is yet an open question.
Recently, there has been a renewed urgency to understand the existence and vast possible functions of micropeptides, particularly in the brain 13–15. Although there is evidence for thousands of putative micropeptides, the validation and characterization of these proteins will require high-throughput efforts across species, conditions, and cell types 16. Key implications from this field include identifying therapeutic or cell targets for neurodevelopmental diseases or disorders; engineering strategies for therapies directed towards de novo protein or drug design; and identifying molecular strategies for co-evolution of chromatin regions that harbor cryptic ORFs in physiologic, stressed, or disease neural states.
As I was working on this problem, I reflected on what drew me to my fascination with small proteins to begin with. I realized that the scientific question appealed to me because I saw myself and so many of my colleagues in this story. Consider the arbitrary limits placed on the definition of a protein. Evidence for, and acceptance of, changed definitions across fields has enabled a whole world of genes to now be deemed worthy of investigation. As such, this work comes at a time not only of scientific innovation, but also of social transformation. What are we missing when we limit our definitions to only the most dominant, visible, acceptable, status quo? What creativity has been ignored or stifled because it didn’t fit the mold? What are the outsized roles of the forces that shape creative strategies of survival – even thriving – and evolution?
This work also got me thinking about the evolutionary history of these micropeptides 17, and how gene networks and cell types may have co-evolved. Thinking about some of the ways that these micropeptide genes emerge, adapt, evolve, or disappear in different contexts provided me a lens through which to understand and confront some of the societal challenges that the life sciences – and academia at large – are, and have been, facing worldwide 18–21. Around the time I was wrapping up this work on micropeptides in zebrafish neurodevelopment 6, the NASEM report on “Advancing Antiracism, Diversity, Equity, and Inclusion in STEMM Organizations: Beyond Broadening Participation” was published 21. In particular, one section drew my attention:
“…the noteworthy ways in which [minoritized] individuals respond to bias in STEMM environments…can be categorized into three general groups: exiting the field, implementing strategies to fit in, and collectively mobilizing to transform the STEMM environment.” 21
How individuals respond to persistent, systemic biases in their environments – exit, adapt, or mobilize – is reflected in what often occurs in biological systems 22,23. Our environments and lived experiences inevitably shape the scientific questions that we ask, how we ask them, and who gets to ask them. The confluence of this report and my own scientific journey highlighted to me how impossible it is to remove ourselves – the experiences and environments of the people doing the science – from the science itself.
So, what are the “micropeptides” in your own work, in your story? I iteratively reflect on these questions both as a basic (neuro)developmental biologist and as an emerging bioethicist 24. As scientists, we can learn from the many unexpected discoveries regarding micropeptides – and any number of yet undervalued fields – to reimagine the tiny changes that can influence entire systems. When they are taken together, they’re not so small after all.
Acknowledgements
In reverse alphabetical order by first name (perhaps you can guess why from my own name), I am grateful to V. Greco, L. Grmai, L. Miao, L. Weiss, E. Strayer, C. Bartman, and A. Giraldez for feedback and/or workshopping through some of these ideas. I am supported by an award from the U.S. Eunice Kennedy Shriver National Institute of Child Health and Human Development (5K99HD105001).
Author Information
Valerie Tornini is currently an associate research scientist at Yale School of Medicine, and an incoming assistant professor in the Department of Integrative Biology and Physiology and the Institute for Society and Genetics at the University of California, Los Angeles (UCLA), USA.
References
1. Benezra, R., Davis, R. L., Lockshon, D., Turner, D. L. & Weintraub, H. The protein Id: a negative regulator of helix-loop-helix DNA binding proteins. Cell61, 49–59 (1990).
2. Ingolia, N. T., Ghaemmaghami, S., Newman, J. R. S. & Weissman, J. S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science324, 218–223 (2009).
3. Slavoff, S. A. et al. Peptidomic discovery of short open reading frame–encoded peptides in human cells. Nat. Chem. Biol.9, 59–64 (2013).
4. Ulitsky, I., Shkumatava, A., Jan, C. H., Sive, H. & Bartel, D. P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell147, 1537–1550 (2011).
5. Bazzini, A. A. et al. Identification of small ORFs in vertebrates using ribosome footprinting and evolutionary conservation. EMBO J.33, 981–993 (2014).
6. Tornini, V. A. et al. linc-mipep and linc-wrb encode micropeptides that regulate chromatin accessibility in vertebrate-specific neural cells. eLife12, e82249 (2023).
7. Mowery, C. T. et al. Trisomy of a Down Syndrome Critical Region Globally Amplifies Transcription via HMGN1 Overexpression. Cell Rep.25, 1898-1911.e5 (2018).
8. Abuhatzira, L., Shamir, A., Schones, D. E., Schäffer, A. A. & Bustin, M. The Chromatin-binding Protein HMGN1 Regulates the Expression of Methyl CpG-binding Protein 2 (MECP2) and Affects the Behavior of Mice. J. Biol. Chem.286, 42051–42062 (2011).
9. Lamanna, F., Hervas-Sotomayor, F. et al. A lamprey neural cell type atlas illuminates the origins of the vertebrate brain. Nat. Ecol. Evol.7, 1714–1728 (2023).
10. Zalc, B. The acquisition of myelin: An evolutionary perspective. Brain Res.1641, 4–10 (2016).
11. González-Romero, R., Eirín-López, J. M. & Ausió, J. Evolution of High Mobility Group Nucleosome-Binding Proteins and Its Implications for Vertebrate Chromatin Specialization. Mol. Biol. Evol.32, 121–131 (2015).
12. Deng, T. et al. Interplay between H1 and HMGN epigenetically regulates OLIG1&2 expression and oligodendrocyte differentiation. Nucleic Acids Res.45, 3031–3045 (2017).
13. Mudge, J. M. et al. Standardized annotation of translated open reading frames. Nat. Biotechnol.40, 994–999 (2022).
14. Sandmann, C.-L. et al. Evolutionary origins and interactomes of human, young microproteins and small peptides translated from short open reading frames. Mol. Cell83, 994-1011.e18 (2023).
15. Duffy, E. E. et al. Developmental dynamics of RNA translation in the human brain. Nat. Neurosci.25, 1353–1365 (2022).
16. Tornini, V. A. Small protein plays with big networks. Trends Genet. TIG S0168-9525(23)00236–6 (2023)
17. Weisman, C. M. The Origins and Functions of De Novo Genes: Against All Odds? J. Mol. Evol.90, 244–257 (2022).
18. Thorp, H. H. It matters who does science. Science380, 873 (2023).
19. Maina, M. B. African neuroscience: Desperately seeking diversity. UNESCO Cour.2022, 15–16 (2022).
20. Silva, A. et al. Addressing the opportunity gap in the Latin American neuroscience community. Nat. Neurosci.25, 1115–1118 (2022).
21. National Academies of Sciences, Engineering, and Medicine. Advancing Antiracism, Diversity, Equity, and Inclusion in STEMM Organizations: Beyond Broadening Participation. (The National Academies Press, 2023).
22. Montgomery, B. L. Lessons from Plants. (Harvard University Press, 2021).
23. Montgomery, B. L. Lessons from Microbes: What Can We Learn about Equity from Unculturable Bacteria? mSphere5, e01046-20 (2020).
24. Tornini, V. A., Peregalli Politi, S., Bruce, L. & Latham, S. R. Maximizing biomedical research impacts through bioethical considerations. Dis. Model. Mech.16, dmm050046 (2023).
The webinar on 5 December 2023 was on the topic of germ cell development and was chaired by Development Editor, Swathi Arur (MD Anderson Cancer Center). Below are the recordings of the talks.
Gabriele Zaffagnini (Centre for Genomic Regulation)
Talk and Q&A by Gabriele Zaffagnini
Diego Sainz de la Maza (University College London)
Talk and Q&A by Diego Sainz de la Maza (1 votes) Loading...
In the middle of the Aix-Marseille University Campus, at the heart of the Calanques National Park, at the border of city. The most beautiful academic spot in France ;)
We are mainly investigating temporal patterning, and how it links development with regeneration and pediatric cancers. Along the years, we have identified key factors that are sequentially expressed in the embryo and that are important to ensure that developmental programs unfold with the correct timing. We have found that failure to do so can lock tissues into permanent development leading to aggressive tumors. This mechanism likely underlies the emergence of pediatric cancers. We are therefore working on the various mechanisms that regulate temporal transitions during development or that coopt temporal patterning to promote regeneration 💪🏻 or pediatric cancers 😫. Our model organisms are Drosophila and the chick embryo.
Lab roll call
Cédric: As a PI,my everyday work consists in making sure that everybody is happy to come in the lab every morning to make exciting science.
Dylan: I joined Cedric Maurange’s team as postdoc to understand how miRNAs can regulate the cellular composition, hierarchy, and growth of pediatric tumors in an established Drosophila cerebral tumor.
Paul: I am the team bioinformatician, working as a research engineer on deciphering how the cell-of-origin affects the differentiation trajectory of rhabdomyosarcoma.
Lauranne: As a non-permanent engineer, I investigate how MYCN or c-Myc overexpression perturbs cell lineage progression and cerebellum’s organization by using chick embryo as study model.
Shobana: I am a PhD student in the team investigating the dynamic role of microRNAs in neuronal maturation in Drosophila
Emma: I am a PhD student working on the self-organizing principles governing tumor growth and I am using a model of Drosophila brain tumor as well as a numerical model of tumor growth.
Favourite technique, and why?
Cédric: I am very interested in single-cell techniques. It was such an amazing moment when my student came with our first single-cell RNA-seq data, back in 2017, which so clearly showed the cellular heterogeneity of neuroblast tumors and how temporal programs are recapitulated in them. It became clear that this technique would be key to unravelling how cellular heterogeneity and hierarchy are regulated in cancer.
We are now trying to use single-cell multiomics combined with computational simulations to decipher how perturbed developmental/temporal trajectories can be corrected in tumors. I am very grateful to the mathematicians and computer scientists who enable us to make sense of these complex datasets.
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Cédric: In general, the ability to measure/visualise a biological process at the single cell level, but in the context of a tissue, is really exciting. At the moment, I am particularly excited about the possibility of precisely measuring the transcriptional activity of a gene at a given developmental time in a single cell and correlating it with its chromatin environment. Self-organisation at all molecular and cellular scales is also fascinating, particularly challenging and attractive because understanding it will require the collaboration of biologists, mathematicians, physicists and computer scientists.
How do you approach managing your group and all the different tasks required in your job?
Cédric: Science should be about passion and fun. People in the lab should be self-motivated. There are so many mysteries to be solved in biology, it is easy to pick up one that you are particularly interested in and contribute to solve it. It is very fascinating also to see where the quest leads us.
I like to give time to newcomers in the lab so that they get familiar with the model and take ownership of the project. Hopefully, after some time they will also come with new ideas of how to tackle the problem. We have weekly lab meetings and my door is always open (except when I am on Zoom !). I expect people to naturally come to me when they want to discuss. We frequently do one-to-one meetings to more precisely assess the current situation and which points to concentrate on. We also have a growing number of projects that relies on tight interdisciplinary collaborations. The combined expertise is usually a talisman for the unexpected.
What is the best thing about where you work?
Cédric: The good atmosphere, the multi-disciplinarity of the institute and the multiple possibilities of collaborations with the other institutes on the campus and throughout the city – everything in a fantastic natural environment and sun all year long.
Dylan: Working in the calanques national park, the Luminy campus site is exceptional.
Paul: The good atmosphere and the possibility to work with scientists with really various research fields and interests. Moreover, the lab and the scientific community of Marseille also offer many social events.
Lauranne: We work in a lovely campus surrounded by nature where the positive atmosphere among the 21 IBDM’s teams not only during work hours but also during after- work gatherings adds to the overall pleasure and satisfaction we find in our work environment.
Shobana: In addition to the institute being multi-disciplinary, the friendly and engaging atmosphere makes it easy to approach people for help or discussion, further facilitated by regular seminars and after-work sessions.
Emma: The IBDM, ideally located in the calanques national park, benefits from the very nice weather of Marseille all year long. Moreover, the institute gathers many people from different backgrounds which makes it very interdisciplinary in addition of the nice and friendly atmosphere.
What’s there to do outside of the lab?
Cédric: Walk in the Calanques and discover the various landscape of Provence, enjoy all the activities of the vibrant city that is Marseille.
Paul: The lab is ideally placed in the “Parc National des Calanques”, a sort of nature spot in the periphery of the big city of Marseille, offering the possibility to do some hiking. In the city center, there are some museums, malls etc. and during winter, a beautiful Christmas market on the Vieux-Port.
Shobana: There’s tons to do right outside the lab like hiking, climbing or just walking. Even looking out of the lab window, one can see how beautiful and serene it is being surrounded by the Calanques. A well-equipped sports complex is quite nearby as well, offering training in different sports.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
“We knew back in the late 1960s that half the human genome was repetitive DNA. And so, where did this myth arise that those scientists were incredibly stupid? I mean, these guys, Jacob and Monod, they got a Nobel Prize!”
In the latest episode of the Genetics Unzipped podcast, we discover how 500,000 whole genomes will help medical research, plumb the depths of the ‘dark genome’, and ponder how much of our DNA is just junk.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
In the first webinar of 2024, Development’s Deputy Editor, Steve Wilson (UCL), hosts three early career researchers studying signalling and early embryogenesis.
Wednesday 31 January – 15:00 GMT
Jonathan Wells (Cornell University) ‘How does the early embryo make heterochromatin?’
Stephanie Telerman (University of Cambridge) ‘Post-translational regulation of the Numb/Notch pathway in neurogenesis and cancer by Dlk2’
Yuki Shindo (Dartmouth College) ‘Nucleocytoplasmic control of early embryogenic cell cycles’
At the discretion of the speakers, the webinar will be recorded for viewing on demand. To see the other webinars scheduled in our series, and to catch up on previous talks, please visit: thenode.biologists.com/devpres
I’m excited to announce that we have now opened applications for the second round of Development’s Pathway to Indpendence Programme. This programme is aimed at supporting postdocs who will be going on the job market next year – providing mentorship, training and networking opportunities. Having piloted this programme with our first talented cohort of PI fellows through the course of 2023, I’m looking forward to working with a new group of postdocs over the course of next year.
You can find out more about the aims this programme, and the application and selection process, in this editorial and on the journal webpage. We encourage applications from all corners of the globe and all areas of developmental and stem cell biology. Those interested in applying can do so via the Company of Biologists’ grants portal, and you are welcome to reach out to me for more information. The deadline for application is 31 January 2024.
One of the main elements of the programme is an in-person meeting of all the PI fellows. In late October, the first cohort of Development’s Pathway to Independence (PI) fellows got together here at The Company of Biologists’ office on the outskirts of Cambridge for a two-day gathering. Having been in contact with these eight outstanding postdocs over the course of this year, it was great finally to meet them all in person and find out a bit more about them, their research and their career plans.
From L-R: Leah Greenspan, Loic Fort, Priti Agarwal (front), Thomas Juan (back), Clotilde Cadart (with one of the next generation of scientists?!), James Gahan, Yuchuan Miao and Polina Kameneva – outside The Company of Biologists’ office in Histon, Cambridge
Navigating the job market and setting up a lab are huge challenges for which many postdocs feel underprepared, and the PI programme aims to help in some small way by providing training, mentorship and peer support – and by helping to raise the profile of our group of PI fellows. At the meeting in Cambridge, our fellows took part in an intense and interactive leadership training course organised by hfp consulting – many thanks to Franck and Dana for facilitating this event. As one of the fellows commented, the course was “extremely informative and gave me real actionable ideas on how to organise my group”; another noted that they “learned a lot about myself as a trainee and a future PI”.
As well as the training course, each of the PI fellows gave a short talk about their future research plans, and received feedback on their presentation from some of the journal’s editors and directors of the Company of Biologists. Personally, I was hugely impressed by the research directions presented by all eight fellows, and I left the meeting feeling energised and excited about the diverse and innovative programmes they all hope to initiate in their independent labs.
As a fledgling programme, it was important to us to find out how well the various elements of the scheme have worked for our PI fellows, so that we can ensure we provide the best support we can going forwards. We’ve received some really valuable feedback from this year’s group that will help us tweak the programme for next year. But overall, it was really great to hear how useful our PI fellows have found the programme. Here’s what some of them had to say:
This programmed has been a game changer for me as I search for an independent position, giving me unprecedented visibility in the community as well as amazing training that will help me in my transition to being a PI .
Through their multiple networking and training opportunities, the PI program empowered me with tools to help launch my independent group and connect with peers at the same career stage, creating a supportive environment.
This programme made me feel part of a community of talented future group leaders and provided me with great opportunities to promote my applications. Sharing is caring!
I wish all our current cohort of PI fellows luck in finding their first independent position – and congratulate those who have already done so! – and I’m very much looking forward to reviewing the next set of applications, and to working with what I’m sure will be an equally talented group of postdocs next year! Please do get in touch if you have any questions about the programme.
I am glad to announce a new Research Topic / Special Issue for the journal Frontiers in Molecular Biosciences (IF 5.0), edited by Prof Rio Sugimura (University of Hong Kong), Prof Emanuele Azzoni (University of Milan-Bicocca), Dr Antonella Fidanza (University of Edinburgh) and Dr Giovanni Canu (University College London).
Michaela Raab, Ersi Christodoulou, Roopesh Krishnankutty, Nicholas T Younger, Konstantinos Gournopanos, Alexander von Kriegsheim, Scott H Waddell, Luke Boulter
Laura Currey, Benjamin Mitchell, Majd Al-Kahlily, Sarah-Jayne McElnea, Danyon Harkins, Alexandra Pelenyi, Nyoman D. Kurniawan, Thomas H. Burne, Lachlan Harris, Stefan Thor, Michael Piper
Samuel Abassah-Oppong, Brandon J. Mannion, Matteo Zoia, Raquel Rouco, Virginie Tissieres, Cailyn H. Spurrell, Virginia Roland, Fabrice Darbellay, Anja Ljubojevic, Julie Gamart, Tabitha A. Festa-Daroux, Carly S. Sullivan, Eddie Rodríguez-Carballo, Yoko Fukuda-Yuzawa, Riana Hunter, Catherine S. Novak, Ingrid Plajzer-Frick, Stella Tran, Jennifer A. Akiyama, Diane E. Dickel, Javier Lopez-Rios, Iros Barozzi, Guillaume Andrey, Axel Visel, Len A. Pennacchio, John Cobb, Marco Osterwalder
Lara M. Siebert-Kuss, Verena Dietrich, Sara Di Persio, Jahnavi Bhaskaran, Martin Stehling, Jann-Frederik Cremers, Sarah Sandmann, Julian Varghese, Sabine Kliesch, Stefan Schlatt, Juan M. Vaquerizas, Nina Neuhaus, Sandra Laurentino
Ian C Tobias, Sakthi D Moorthy, Virlana M Shchuka, Lida Langroudi, Zoe E Gillespie, Andrew G Duncan, Ruxiao Tian, Mariia Cherednychenko, Natalia A Gajewska, Raphaël B Di Roberto, Jennifer A Mitchell
Csenge Földi, Zsolt Merényi, Bálint Balázs, Árpád Csernetics, Nikolett Miklovics, Hongli Wu, Botond Hegedüs, Máté Virágh, Zhihao Hou, Xiao-Bin Liu, László Galgóczy, László G. Nagy
George C. Gabriel, Hisato Yagi, Tuantuan Tan, Abha Bais, Benjamin J. Glennon, Margaret C. Stapleton, Lihua Huang, William T. Reynolds, Marla G. Shaffer, Xinxiu Xu, Madhavi Ganapathiraju, Dennis Simon, Ashok Panigrahy, Yijen L. Wu, Cecilia W. Lo
Maria Victoria Hinckelmann, Aline Dubos, Victorine Artot, Gabrielle Rudolf, Thu Lan Nguyen, Peggy Tilly, Valérie Nalesso, Maria del Mar Muniz Moreno, Marie-Christine Birling, Juliette D. Godin, Véronique Brault, Yann Herault
J. Guillermo Sanchez, Scott Rankin, Emily Paul, Heather A. McCauley, Daniel O. Kechele, Jacob R. Enriquez, Nana-Hawa Jones, Siri AW Greeley, Lisa Letourneau-Friedberg, Aaron M. Zorn, Mansa Krishnamurthy, James M. Wells
Lana Vukadin, Bohye Park, Mostafa Mohamed, Huashi Li, Amr Elkholy, Alex Torrelli-Diljohn, Jung-Hyun Kim, Kyuho Jeong, James M Murphy, Caitlin A. Harvey, Sophia Dunlap, Leah Gehrs, Hanna Lee, Hyung-Gyoon Kim, Seth N. Lee, Denise Stanford, Robert A. Barrington, Jeremy B. Foote, Anna G. Sorace, Robert S. Welner, Blake E. Hildreth III, Ssang-Taek Steve Lim, Eun-Young Erin Ahn
Connie S. Chamberlain, Archana Prabahar, John A Kink, Erika Mueller, Yiyao Li, Stephanie Yopp, Christian M. Capitini, Peiman Hematti, William L. Murphy, Ray Vanderby, Peng Jiang
Sara Cascione, Elena Fontana, Rebecca Scarfò, Rosita Rigoni, Valentina Capo, Elena Draghici, Kerry Dobbs, Luigi D. Notarangelo, Anna Villa, Andrea Ditadi
Chenchen Li, Hanna Shin, Dheeraj Bhavanasi, Mai Liu, Xiang Yu, Scott A. Peslak, Xiaolei Liu, Juan R. Alvarez-Dominguez, Gerd A. Blobel, Brian D. Gregory, Jian Huang, Peter S. Klein
Jessica B Cruz, Carolina M Nogueira, Juliano R Sangalli, Ricardo P Nociti, Dewison R Ambrizi, Alessandra Bridi, Jorge Pinzon, Maira BR Alves, Vera FMH de Lima, Yeda F Watanabe, Fabiana F Bressan, Flavio V Meirelles, Rafael V Sampaio
Christian Wiese, Miriam Abele, Benjamin Al, Melina Altmann, Alexander Steiner, Nils Kalbfuss, Alexander Strohmayr, Raksha Ravikumar, Chan Ho Park, Barbara Brunschweiger, Chen Meng, Eva Facher, David W. Ehrhardt, Pascal Falter-Braun, Zhi-Yong Wang, Christina Ludwig, Farhah F. Assaad
Yuan Zhang, Deepak Sharma, Yan Liang, Nick Downs, Fleur Dolman, Kristen Thorne, Jose Henrique Pereira, Paul Adams, Henrik V. Scheller, Malcolm O’Neill, Breeanna Urbanowicz, Jenny C. Mortimer
Ambre Guillory, Mauricio Lopez-Obando, Khalissa Bouchenine, Louis Lambret, Philippe Le Bris, Alain Lécureuil, Jean-Paul Pillot, Vincent Steinmetz, François-Didier Boyer, Catherine Rameau, Alexandre de Saint Germain, Sandrine Bonhomme
Elise Parey, Olga Ortega-Martinez, Jérôme Delroisse, Laura Piovani, Anna Czarkwiani, David Dylus, Srishti Arya, Samuel Dupont, Michael Thorndyke, Tomas Larsson, Kerstin Johannesson, Katherine M Buckley, Pedro Martinez, Paola Oliveri, Ferdinand Marlétaz
Rion Brattig Correia, Joana M. Almeida, Margot J. Wyrwoll, Irene Julca, Daniel Sobral, Chandra Shekhar Misra, Sara Di Persio, Leonardo G. Guilgur, Hans-Christian Schuppe, Neide Silva, Pedro Prudêncio, Ana Nóvoa, Ana S. Leocádio, Joana Bom, Sandra Laurentino, Moisés Mallo, Sabine Kliesch, Marek Mutwil, Luis M. Rocha, Frank Tüttelmann, Jörg D. Becker, Paulo Navarro-Costa
Anastasia I. Kurtova, Alexander D. Finoshin, Margarita S. Aparina, Guzel R. Gazizova, Olga S. Kozlova, Svetlana N. Voronova, Elena I. Shagimardanova, Evgeny G. Ivashkin, Elena E. Voronezhskaya
Melissa J., Cristina Capitanio, Ian R. Smith, Julia C. Paoli, Anna Bieber, Yizhi Jiang, Joao A. Paulo, Miguel A. Gonzalez-Lozano, Wolfgang Baumeister, Florian Wilfling, Brenda A. Schulman, J. Wade Harper
Alexandra V. Bruter, Ekaterina A. Varlamova, Nina I. Stavskaya, Zoia G. Antysheva, Vasily N. Manskikh, Anna V. Tvorogova, D. S. Korshunova, Alvina I. Khamidullina, Marina V. Utkina, Viktor P. Bogdanov, Alyona I. Nikiforova, Eugene A. Albert, Denis O. Maksimov, Jing Li, Mengqian Chen, Alexander A. Shtil, Igor B. Roninson, Vladislav A. Mogila, Yulia Y. Silaeva, Victor V. Tatarskiy
Brendan J Houston, Joseph Nguyen, D. Jo Merriner, Anne E O’Connor, Alexandra M Lopes, Liina Nagirnaja, Corinna Friedrich, Sabine Kliesch, Frank Tuettelmann, Kenneth I Aston, Don Conrad, Robin M Hobbs, Jessica EM Dunleavy, Moira K O’Bryan
Ankita Chavan, Randi Isenhart, Son C. Nguyen, Noor Kotb, Jailynn Harke, Anna Sintsova, Gulay Ulukaya, Federico Uliana, Caroline Ashiono, Ulrike Kutay, Gianluca Pegoraro, Prashanth Rangan, Eric F. Joyce, Madhav Jagannathan
Laura J. A. Hardwick, Benjamin P. Davies, Sara Pensa, Maedee Burge-Rogers, Claire Davies, André Figueiredo Baptista, Robert Knott, Ian McCrone, Eleonora Po, Benjamin W. Strugnell, Katie Waine, Paul Wood, Walid T. Khaled, Huw D. Summers, Paul Rees, John W. Wills, Katherine Hughes
Yosuke Mai, Yasuaki Kobayashi, Hiroyuki Kitahata, Takashi Seo, Takuma Nohara, Sota Itamoto, Shoko Mai, Junichi Kumamoto, Masaharu Nagayama, Wataru Nishie, Hideyuki Ujiie, Ken Natsuga
Quan Xu, Lennard Halle, Soroor Hediyeh-zadeh, Merel Kuijs, Umut Kilik, Qianhui Yu, Tristan Frum, Lukas Adam, Shrey Parikh, Manuel Gander, Raphael Kfuri-Rubens, Dominik Klein, Zhisong He, Jonas Simon Fleck, Koen Oost, Maurice Kahnwald, Silvia Barbiero, Olga Mitrofanova, Grzegorz Maciag, Kim B. Jensen, Matthias Lutolf, Prisca Liberali, Joep Beumer, Jason R. Spence, Barbara Treutlein, Fabian J. Theis, J. Gray Camp
Recently, one of the preLights Ambassadors, Martin Estermann, launched a new webinar series that discusses the journey of a preprint in becoming a journal publication. As part of this initiative, selected preprint authors explain their biological research, focussing on the original story presented in the preprint and how this changed during revisions. They will also be able to reflect on the potential benefits of having their research available to the scientific community before formal peer review.
Tonight, at 17:00 GMT, Martin will talk to Laura Kerosuo (NIH/NIDCR, USA), whose lab featured in the Node’s ‘Lab meeting’ series (link to post). They will discuss a preprint the lab posted at the beginning of this year, titled: “Maintenance of pluripotency in the entire ectoderm enables neural crest formation”. This preprint was highlighted by Andrew Montequin, a PhD student at Northwestern University, as part of preLights (see preLights post) and was recently published in Nature Communications.
We’ll have another session on the topic of NLR immune receptor–nanobody fusions and plant disease resistance next week. This discussion will include preprint authors Jiorgos Kourelis, Clemence Marchal and Jose Salguero-Linares. For more information about this webinar, please follow this link.
Doing great science depends on teamwork, whether this is within the lab or in collaboration with other labs. However, sometimes the resources that support our work can be overlooked. Our ‘Featured resource’ series aims to shine a light on these unsung heroes of the science world. In this post, the team behind Xenbase introduces the key features on the database and suggests how the community can contribute to the mission of Xenbase.
When was the Xenbase established and what are its aims?
Xenbase was the brainchild of Peter Vize. Originally conceived of as an online catalog of gene expression images in the late 1990’s, the golden age of gene expression screens, where essentially every lab was producing 100s of images of gene expression throughout embryonic development, yet those pictures were sitting on a lab computer not being shared or annotated in any meaningful way. Peter saw that a resource to share this information would have a huge impact to cut down on wasted time, effort and resources (i.e. save funding dollars).
The first iteration of Xenbase was launched in 2000, and by 2002, Xenopus tropicalis was earmarked for whole genome sequencing by the Joint Genome Institute (JGI), so the vision for Xenbase quickly morphed into a bigger project: integrated genomics and gene expression on a fully searchable database.
The initial challenge faced when building Xenbase was to combine the research from two Xenopus species used in complementary but (almost) non-overlapping fields of cell biology and embryology. Xenopuslaevis had a long history as a lab frog, with decades of literature covering organogenesis, cell fate maps and cell biology, gene function and gene expression, but as a polyploid of hybrid origin, the genome hadn’t been sequenced (and was a long way off). In comparison, the diploid Xenopus tropicalis genome was being sequenced and this smaller frog was being adopted for disease modeling, and had lots of EST data, but there wasn’t a lot of other biological data for ‘trops’. It was clear both Xenopus species were in need of database support, and the Xenbase founders met that challenge head on, in large part by learning from the already established MODs like MGI (mouse) and Zfin (zebrafish). Xenbase was the first MOD to support two species and essentially three genomes (i.e., X. tropicalis and the 2 subgenomes of X. laevis).
The overarching aim of Xenbase is to simultaneously support labs using Xenopus as a research model, share the genomes and bioinformatic information about genes/proteins, codify the results of the research via deep and expert curation and thus support basic and applied science to accelerate discovery. Having all the data about Xenopus in one place has huge advantages. Within a few years enough people found that ‘what works for X. laevis, works for X.tropicalis’, e.g., gene expression in X. laevis and X. tropicalis are nearly always the same (or very similar), so the same reagents (such as MOs, gRNAs, and antibodies) can be used in both species.
Who are the people behind the resource/ who runs the resource?
Xenbase has two teams and two performance sites. The curation team is based at the Division of Developmental Biology, at Cincinnati Children’s in Ohio, USA, led by Prof. Aaron Zorn. We currently have four curators and bioinformatician/genome analyst in the Cincinnati-based curation team: VG Ponferrada, Malcolm Fisher, Andrew Bell, Christina James-Zorn, and Ngoc Ly. We also have a student assistant, Nguyen Thuy Vy Ngo, who helps triage the new literature. The development team, headed by Prof. Peter Vize, is based in the Department of Biological Sciences at the University of Calgary, in Alberta, Canada. The Calgary development team is led by Kamran Karimi, with support of the database architect Troy Pells, and the Bioinformatian/genome specialist, Vaneet Lotay, who are further supported by software developers, Joe Wang and Stan Chu. We also share code and development with our sister-website, Echinobase, so the Calgary based team is ably supported by Brad Arshinoff and Sergei Agalakov.
What tools/resources are available for researchers?
Xenbase is a gene-centric database, with a single ‘umbrella’ gene page showing the X. tropicalis gene and the two X. laevis paralogs, typical gene expression images at different embryonic stages (when available) and other data such as reagents, orthologs and OMIM/DO associated diseases (and many more links to associated data). Each gene page then has a series of tabs, like a folder, starting with the Expression (all images in database with expression), Phenotypes (all experiments that either manipulate or assay the gene(s)) and Literature (all literature that cite the gene(s)). The next set of tabs cover GO terms, Nucleotides, Proteins and Interactants, which collate annotations and/or accession about the genes/protein products with data pulled in from, and linked to, numerous trusted databases (NCBI, ENSEMBL, UniProt, InterPro/TrEMBL, GO and IntAct).
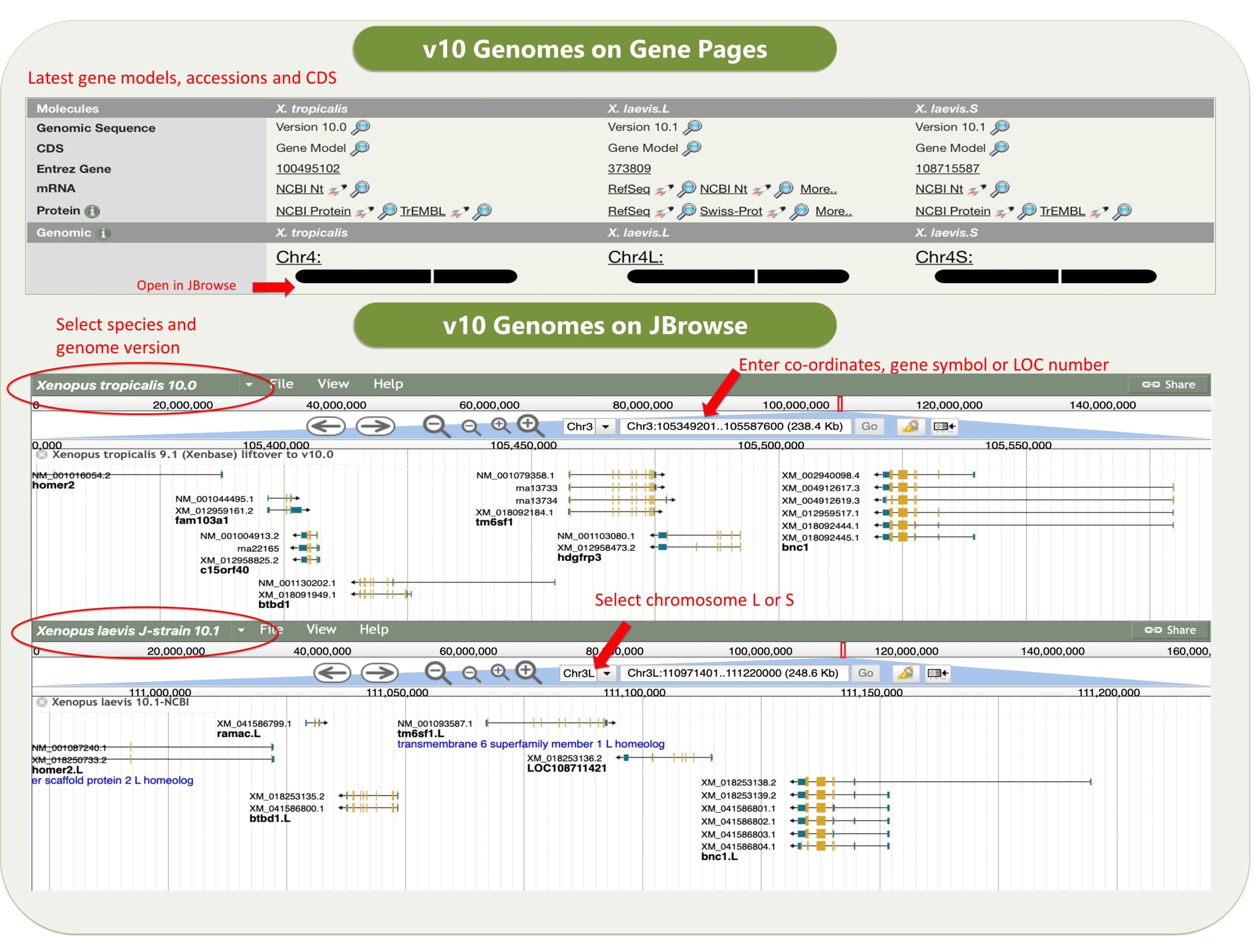
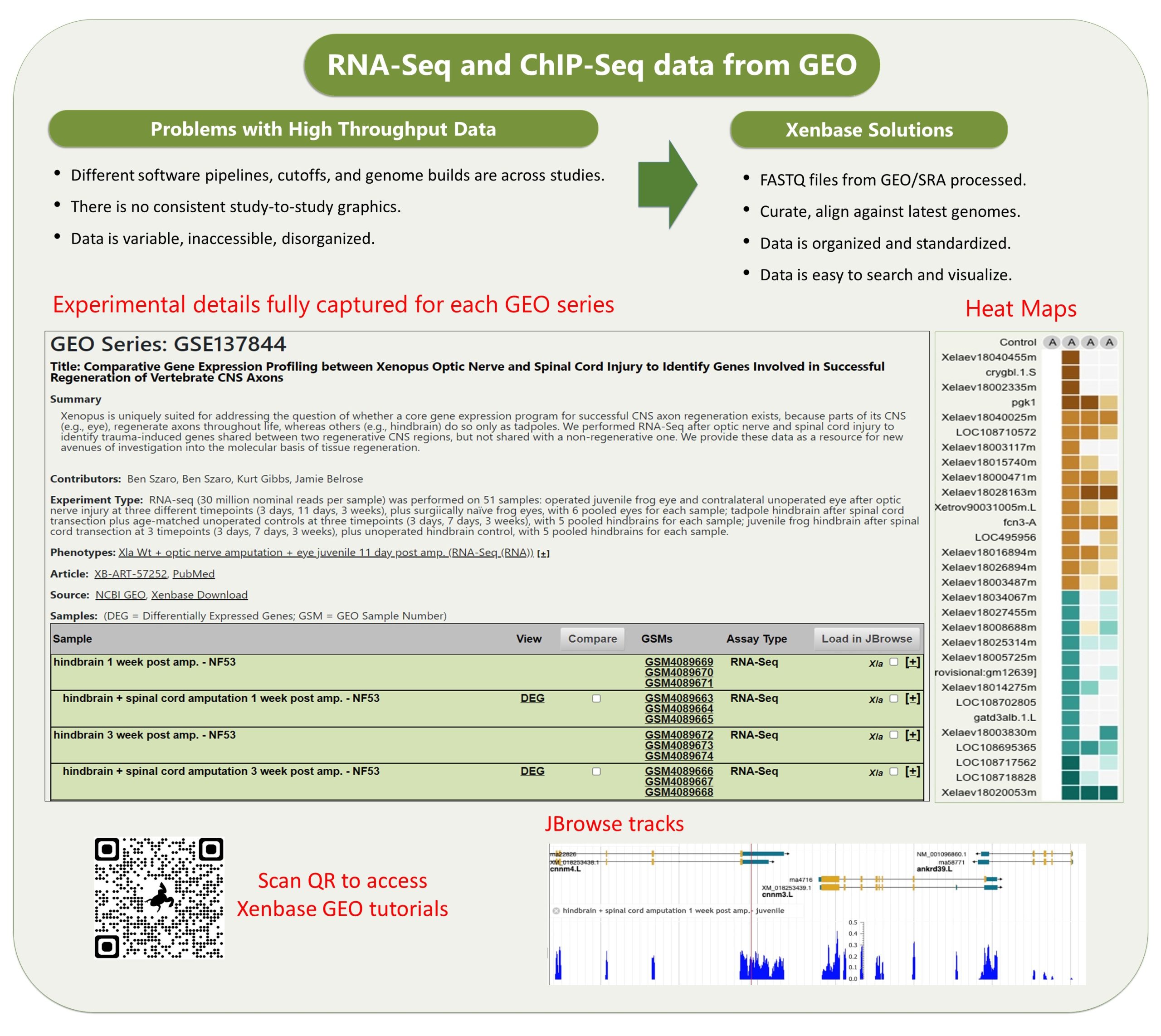
Xenbase has various tools that are widely used. The genome viewer JBrowse has the latest v10 Xenopus genomes, and a huge variety of other useful tracks such as CRISPR-Scan predicted guide RNAs to help design your mutant lines, a morpholino track, and an enormous number of RNA-seq and ChIP-seq tracks, tracks for histone marks and transcriptions factor binding sites, and many more.
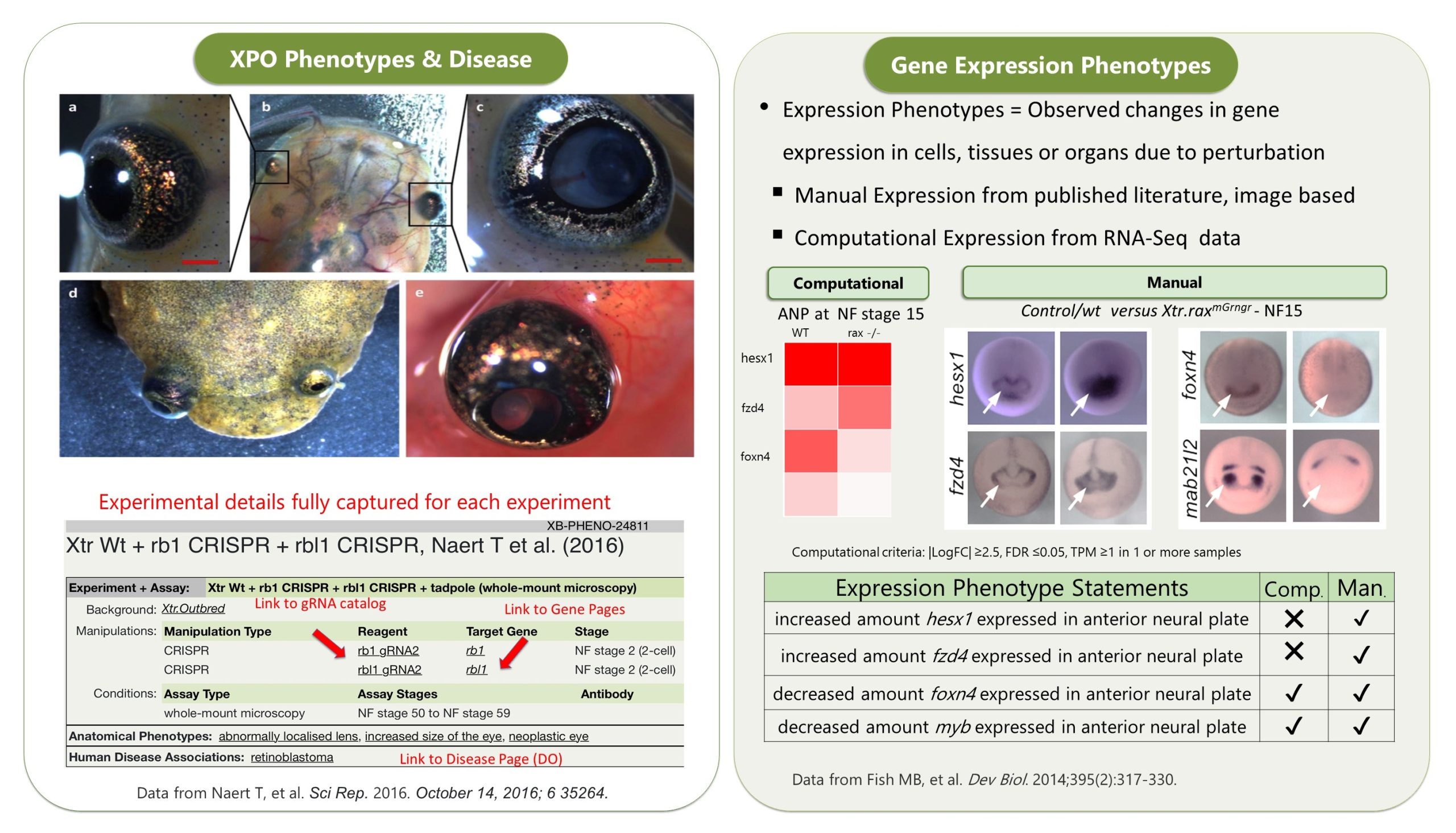
The other data module which is really cool is our GEO data ( accessed via Expression Menu/GEO data @ Xenbase). We took the publicly submitted high throughput sequence data from the NCBI’s GEO database, manually curated the supporting metadata and then we processed it through a pipeline that ‘harmonizes’ the different studies (see Fortriede et al 2020 for all the details). Even though the data are from different studies, researchers can view all of this data in a standardized format, aligned to the latest v10 genomes and via heat maps that visualize DEGs (differentially expressed genes). In addition, we have 1000’s of ‘computational’ gene expression phenotype statements from these experiments, where we generate statements in the readable format, e.g., “manipulation X increases/decreases the expression of gene Y in tissue Z at NF stage #”, all linked to genes, literature and the original GEO data. These ‘gene expression phenotypes’ are most easily returned via our Phenotypes search (e.g., search for six1 . We hope this mass of curated RANSeq and ChIPSeq data will help inform GRNs, and let researchers see all the results from other peoples experiments that pertain to their gene of interest.
Any hidden gems, features that are new, or that researchers might be less aware of?
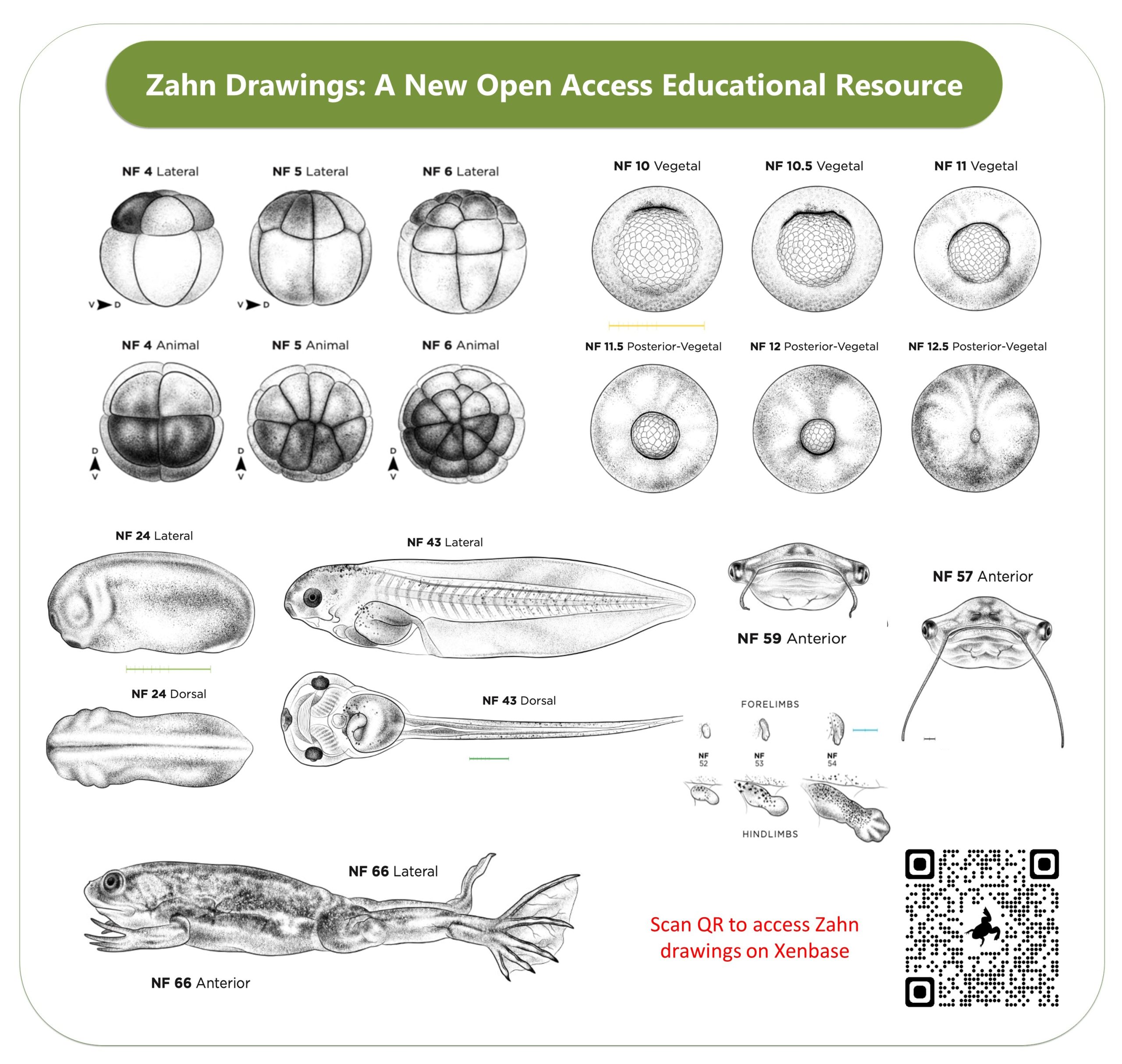
Two new features we are super proud of are the open access drawings of Xenopus embryonic stages (the Normal table) by Natalya Zahn and the accompanying Landmarks Table. The Zahn drawings, which are in the same style as the classic 1950’s Nieuwkoop and Faber drawings, are open access and cover more views- especially anterior and ventral views. Also, the accompanying Staging Landmarks Table that we built to help researchers in the wet lab stage embryos also includes internal developmental milestones and gene markers. We hope both of these resources become indispensable for university courses, in wet labs and embryology courses.
I think the Xenopus Community pages, which include personal profiles and lab pages, are both great ways to promote one’s research and attract students and collaborators. I recommend PIs appoint a trainee in the lab to make sure their Xenbase profiles & Lab page is up-to-date, and that all members of the Lab have a profile to record their research interests and a list of their publications. The Xenbase Jobs Board is also available to post any open positions, from graduate students to postdocs and group leaders and department chairs.
We are constantly working to keep the information on Xenbase up to date and synchronized with the other major databases and repositories, and we recently joined the Alliance of Genome Resources. By collating data from diverse model organisms (worm, yeast, fly, mice, frogs, rats and fish) the Alliance aims to improve the understanding of the genetic and genomic basis of human biology, health and disease. Frogs have played, and continue to play, an important part of that discovery process.
How can the community contribute?
The single most effective way for researchers to contribute to the mission of Xenbase, is to choose open access (OA) options to publish their research, whenever possible. When data is locked up behind a paywall, we literally can’t see it, so we can’t curate it. If we don’t curate the data, the results become essentially invisible over time, and its immediate and long term impact is greatly reduced. Accessible data becomes curated data, which is discoverable and will be cited more too. Uncurated data is easily overlooked.
Another way people can contribute is to send us their images of novel gene expression, especially images of stages beyond what is included in their papers. Instead of being saved in an inaccessible folder on a lab computer, send them to Xenbase and share them with the world!. All community submitted images are fully attributed to the people and that lab that makes them!
We also run a help desk: xenbase@cchmc.org. Contact us anytime with community submissions, questions, feedback, concerns, troubleshooting help and your ideas to make Xenbase an even better, more useful resource.
Where does funding come from?
Xenbase is currently supported by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD). Previous financial support also came from the National Science Foundation (NSF), the Wellcome Trust (UK), the BBSRC (UK), the Alberta Network for Proteomics Innovation (ANPI).
If money was no object, what would you like to add to Xenbase?
We’re looking into some new great tools including updating to JBrowse2 (to allow viewing multiple genomes at once), and new data graphics to show/assess synteny across Xenopus genomes, and supporting single-cell data. We’d also like to develop more educational resources to support students and teaching labs, including a high tech histology or a 3D atlas of Xenopus from embryos to adults, more anatomy atlas modules, perhaps including virtual dissection of adult Xenopus frogs. We would like to fund more illustrations of Xenopus development, to really fill in the Normal Table we published in Zahn et al 2022 in Development! We would also love to get movies (which are more and more common in publications) embedded on the articles pages. All of these ideas have been floated, so we’ll see what we can get done.
References
Fortriede JD, Pells TJ, Chu S, Chaturvedi P, Wang D, Fisher ME, James-Zorn C, Wang Y, Nenni MJ, Burns KA, Lotay VS, Ponferrada VG, Karimi K, Zorn AM, Vize PD, Xenbase: deep integration of GEO & SRA RNA-seq and ChIP-seq data in a model organism database, Nucleic Acids Res., Volume 48, Issue D1, Pages D776-D782, doi:10.1093/nar/gkz933
Zahn N, James-Zorn C, Ponferrada VG, Adams DS, Grzymkowski J, Buchholz DR, Nascone-Yoder NM, Horb M, Moody SA, Vize PD, Zorn AM, Normal Table of Xenopus development: a new graphical resource, Development, 2022 Jul 15;149(14):dev200356, doi:10.1242/dev.200356
Fisher M, James-Zorn C, Ponferrada V, Bell AJ, Sundararaj N, Segerdell E, Chaturvedi P, Bayyari N, Chu S, Pells T, Lotay V, Agalakov S, Wang DZ, Arshinoff BI, Foley S, Karimi K, Vize PD, Zorn AM. Xenbase: key features and resources of the Xenopus model organism knowledgebase. Genetics. 2023 May 4;224(1):iyad018. doi: 10.1093/genetics/iyad018. PMID: 36755307; PMCID: PMC10158840.

 (6 votes)
(6 votes)
 (No Ratings Yet)
(No Ratings Yet)