We want to understand how development is controlled and evolves to generate new phenotypes. To answer this, we study segmented worms (annelids) with spiral cleavage, a unique and ancient mode of development found in many marine invertebrates. We are interested in discovering how different strategies to specify the early progenitors of all cell types and organs of an animal evolved in spiral cleavage and how changes in the timing and maternal contribution defining those cells might influence phenotypic evolution during subsequent development, such as in the adult morphology and the life cycles. We approach these questions by combining multiple methodologies, from genomics, epigenomics and transcriptomics to more classical embryology, and using non-model organisms, some of which we are establishing as tractable research species for the first time.
Can you give us a lab roll call?
Currently, there are two postdoctoral (Yan Liang and Allan Carrillo-Baltodano) and four postgraduate researchers (Kero Guynes, Francisco Manuel Martín-Zamora, Rory Donnellan and Billie Davis) in my lab.
Yan studies the dynamics of gene expression and regulation during spiral cleavage and their correlation with the evolution of different life cycles and cell fate specification strategies.
Allan works on the evolution of maternal determinants and early cleavage in the specification of the body axes.
Kero investigates the role of DNA methylation during spiral cleavage and in annelids.
Fran has spearheaded the study of gene regulation and histone post-translational modifications during annelid embryogenesis and spiral cleavage.
Rory is investigating the regeneration of anterior structures in our favourite annelid, Owenia fusiformis.
Billie is exploring the evolution and role of 3D genome architecture in annelids and spiral cleavage.
Lab photo taken in 2022, including former PhD and MSc students.
Favourite technique, and why?
Chema: My favourite technique is in situ hybridisation because I love to see where genes are expressed. But I’m sure each member of the lab has their own!
Apart from your own research, what are you most excited about in developmental and stem cell biology?
Chema: I love evolutionary and comparative developmental biology generally. But I am particularly interested in the embryology of weird and understudied creatures, what they can tell us about animal development, and the evolution of the fascinating diversity of animal forms.
How do you approach managing your group and all the different tasks required in your job?
Chema: I like to support and train the members of my lab to become independent and have a sense of ownership of their projects. This is essential to stay motivated and willing to discover new things, even during the challenging moments in every research project. Efficient time management and ensuring time for hobbies, friends, and family is the key!
What is the best thing about where you work?
Chema: Queen Mary and the Department of Biology is a diverse and collegial environment. People work on many different organisms and questions, from plants and microbes to bats and fossils. That is very stimulating! It helps you learn something new daily and think about your interests and problems from new perspectives.
Allan: The community in the biology department at QMUL is very strong. Most postdocs initially come without a sense of what to do in a big, diverse city like London. However, the PhD students, in particular, make everyone welcome, and soon enough, you cannot skip a pub night on Fridays. The informal settings allow for a natural discussion of things that can help in our research and careers.
What’s there to do outside of the lab?
Chema: We are in East London! It is one of London’s most vibrant, diverse, young, and exciting parts.
Allan: London has everything you can imagine, but the music scene is excellent. From small bohemian venues to listen to folk music to mega stadiums to see your favourite rock band. There is something for everyone.
Browse through other ‘Lab meeting’ posts featuring developmental and stem cell biology labs around the world.
In a recent review published in Acta Neuropathologica, Matuleviciute, Akinluyi & co-authors focus on the evidence obtained from humans to examine how microglia, the brain’s macrophages, participate in brain development. The review focuses on neurodevelopmental disorders, a set of complex disorders with significant heterogeneity in aetiology and presentation, such as autism spectrum conditions (ASCs) and schizophrenia (SZ). The authors make the case for this focus by specifying that many of the observed symptoms in ASCs and SZ are cognitive and characteristically human such as language difficulties, executive function breakdown and positive auditory symptoms. Furthermore, they also highlight key differences between human and non-human microglia. The review specifies that we are far from identifying mechanisms because human studies are descriptive and though, microglia are part of the pathological signature of these disorders, it remains challenging to concur whether they are reactive to an initial insult or causative of altered neurodevelopment. The authors highlight cutting-edge methodologies that are based on human tissues to help elucidate mechanisms of the interactions between microglia and the topography. This includes high-resolution transcriptomic studies and human tissue-based models. Finally, the co-authors critically examine the success of treatments in neurodevelopmental disorders that have targeted microglia[1]. Check it out!
1. Matuleviciute, R., et al., Microglial contribution to the pathology of neurodevelopmental disorders in humans. Acta Neuropathologica, 2023.
First, I’d be remiss not to acknowledge that most of my posts have been on vertebrate systems when many invertebrates are also studied for their regenerative capacity. One such example is Hydra (Vogg et al., 2019) but Hydra papers don’t seem to appear in JEEM/Development until the 1960s. While exploring the early issues of JEEM/Development, I found fish manuscripts were similarly rare, which is surprising because – today – zebrafish are a prominent model for development and regeneration (Marques et al., 2019). Indeed, there was also a notable absence of any studies of nervous system regeneration, maybe because such articles were submitted to more specialist neurobiology and neuroscience journals. In this final post of the series, we fill this missing link by discussing the highlights from the following articles:
Caltech researcher Roger Wolcott Sperry is most famous for his work on the ‘split brain hypothesis’ and the corpus callosum, for which he won the Nobel Prize for Medicine/Physiology in 1981 (Hubel, 1994; De León Reyes et al., 2020). Alongside his work in cats, Sperry published a few articles on cichlid fish. One such article was published in JEEM/Development in 1957 (Arora and Sperry, 1967). His colleague, R. W. Arora, seems to have left little trace but I think it’s likely he brought the fish to the whole operation. Their study continued previous work on the basis that humans failed to regenerate motor neurons because of aberrant guidance and muscle innervation following wounding or amputation, which doesn’t occur in other vertebrates. To learn more about peripheral nervous system (Murtazina and Adameyko, 2023) regeneration, Arora and Sperry focused on the mandible of Astronotus ocellatus, severing the left mandibular nerve of ten fish and causing partial paralysis had completely recovered by 16 days post-operation (Fig. 1, left). Next, they cut the individual branches of the mandibular nerve and crossed them so that the nerves lay next to muscles they wouldn’t normally innervate (Fig. 1, right). Overall, as with the complete nerve cuts, mandible function was recovered a couple of weeks later with the nerves innervating the adjacent muscle rather than their original target (I have simplified the results here, do see the manuscript for full details). Arora and Sperry even used an electric induction coil to prove innovation and show that stimulating the regenerated nerves would cause the contraction of the new muscle target. This work showed the plasticity of the peripheral nervous system to adapt to the rewiring of the motor neurons with new muscles. Furthermore, this study showed that the guidance cues that direct motor neuron migration and innovation during development might not be present during regeneration, signalling the importance of the microenvironment for pathfinding and differentiating the requirements for neuronal targeting vs. neuronal innervation.
.
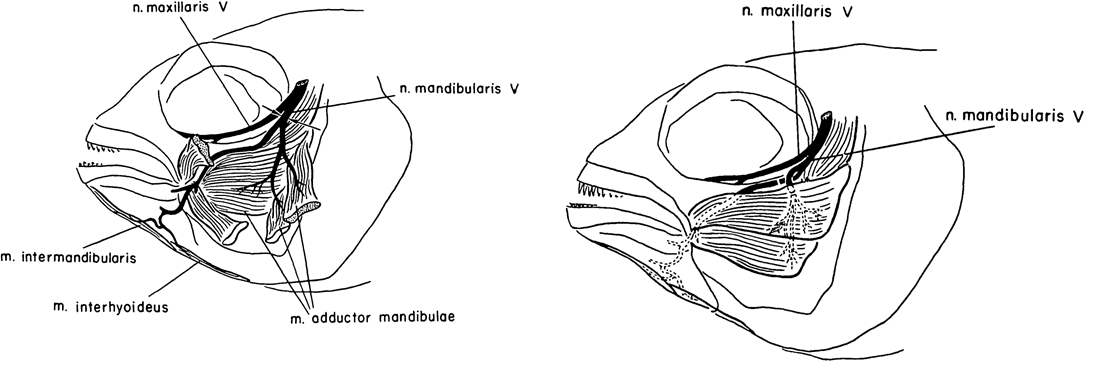
Fig. 1. Left: “Sketch of jaw musculature and its innervation in Astronotus ocellatus. The two superficial adductor muscles are cut and deflected to expose the deeper adductor muscle. Arrows indicate point of section of mandibularis V nerve in Series I.” Right “Sketch showing cross-union of levator and depressor branches of mandibularis V nerve.” Taken from Arora and Sperry, 1957.
Teleost fish, including cichlids, are prominent in today’s research, although I think it’s fair to say that zebrafish are a dominant system. The explosion of zebrafish studies can largely be attributed to the Boston and Tübingen screens, culminating in the publication of a special issue of Development a little over 25 years ago (Nüsslein-Volhard 2012; Mullins et al., 2019). Since, the generation of hundreds (more?) of genetic lines, including fluorescent genetic reporters that capitalise on the zebrafish’s amenability to microscopy and imaging. As with cardiac regeneration, which we heard about earlier this week, the zebrafish (unlike most mammals, including humans) can regenerate its spinal cord (Becker and Becker, 2022).
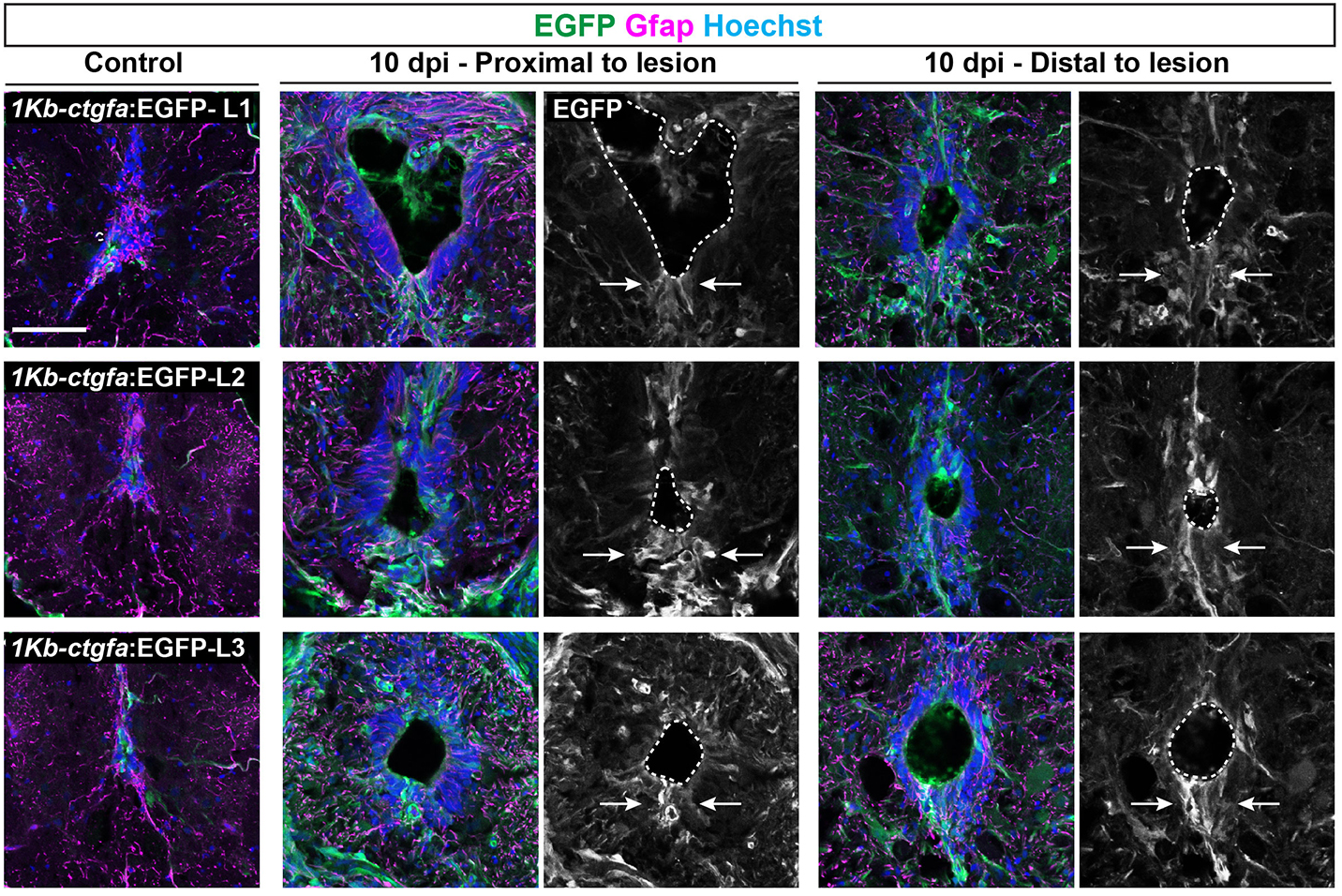
In addition to the peripheral nervous system, zebrafish have a remarkable capacity for regenerating the central nervous system, including the spinal cord, following injury – a phenomenon that doesn’t occur in mammals due to glial cell-dependent scarring. Thus, understanding how to prevent scaring and/or induce regeneration has huge therapeutic potential for spinal cord injuries. In their recent Development paper, Zhou and colleagues use a suite of sophisticated genetic tools to ask how glial cells respond to injury in zebrafish (Zhou et al., 2023). The authors first generate ctgfa-Tracer zebrafish for lineage tracing ctgfa-expressing cells, such as bridging glia, which emerge following spinal cord injury. Using this line, the authors show that bridging glia, ventral ependymal progenitors and regenerating glial cells are derived from ctfga-expressing cells but they minimally contribute to neurons and oligodendrocytes. The authors next turn to ctfga regulation, revealing the gene-regulatory sequences that drive ctfga expression during regeneration. Finally, the authors specifically ablate ctgfa-expressing cells, and show that axon outgrowth and swimming behaviour are affected following spinal injury. Together, Zhou and colleagues determine that ctfga-expressing cells have a pro-regenerative role in spinal cord regeneration.
Fig. 2. “A distal enhancer element directs glial ctgfa expression after SCI.” Taken from Fig. 4B (Zhou et al., 2023). Please see the original publication for the full legend.
Both these papers address the ability of teleost fish to regenerate the nervous system and highlight the importance of the local microenvironment, as well as using animal behaviour (e.g. eating and swimming) as a read-out of phenotype. Arora and Sperry showed the regenerative plasticity of the peripheral nervous system, whereas Zhou and colleagues highlighted a particular cell population that supports and contributes to central nervous system regeneration.
I hope this little series has been entertaining – I’ve certainly enjoyed learning more about these early papers and the researchers who wrote them. Would you be interested in reading more ‘now and then’ or ‘past and present’ posts on other topics? Share your thoughts in the comments.
References
H. L. Arora, R. W. Sperry; Myotypic Respecification of Regenerated Nerve-fibres in Cichlid Fishes. Development 1 September 1957; 5 (3): 256–263. doi: https://doi.org/10.1242/dev.5.3.256
Thomas Becker, Catherina G. Becker; Regenerative neurogenesis: the integration of developmental, physiological and immune signals. Development 15 April 2022; 149 (8): dev199907. doi: https://doi.org/10.1242/dev.199907
Noelia S. De León Reyes, Lorena Bragg-Gonzalo, Marta Nieto; Development and plasticity of the corpus callosum. Development 15 September 2020; 147 (18): dev189738. doi: https://doi.org/10.1242/dev.189738
Ines J. Marques, Eleonora Lupi, Nadia Mercader; Model systems for regeneration: zebrafish. Development 15 September 2019; 146 (18): dev167692. doi: https://doi.org/10.1242/dev.167692
Mary C. Mullins, Joaquín Navajas Acedo, Rashmi Priya, Lilianna Solnica-Krezel, Stephen W. Wilson; The zebrafish issue: 25 years on. Development 15 December 2021; 148 (24): dev200343. doi: https://doi.org/10.1242/dev.200343
Aliia Murtazina, Igor Adameyko; The peripheral nervous system. Development 1 May 2023; 150 (9): dev201164. doi: https://doi.org/10.1242/dev.201164
Christiane Nüsslein-Volhard; The zebrafish issue of Development. Development 15 November 2012; 139 (22): 4099–4103. doi: https://doi.org/10.1242/dev.085217
Stefano Sandrone. Roger W. Sperry (1913-1994). J Neurol. 2022 Sep;269(9):5194-5195. doi: 10.1007/s00415-022-11232-6. Epub 2022 Jul 22. PMID: 35867150; PMCID: PMC9363358.
Matthias C. Vogg, Brigitte Galliot, Charisios D. Tsiairis; Model systems for regeneration: Hydra. Development 1 November 2019; 146 (21): dev177212. doi: https://doi.org/10.1242/dev.177212
Lili Zhou, Anthony R. McAdow, Hunter Yamada, Brooke Burris, Dana Klatt Shaw, Kelsey Oonk, Kenneth D. Poss, Mayssa H. Mokalled; Progenitor-derived glia are required for spinal cord regeneration in zebrafish. Development 15 May 2023; 150 (10): dev201162. doi: https://doi.org/10.1242/dev.201162
Our first webinar in October will be chaired by Development Editor Debby Silver (Duke University) and features three early-career researchers studying neurodevelopment and regeneration. The webinar will be held using Zoom with a Q&A session after each talk.
Wednesday 4 October 2023 – 15:00 BST
Baptiste Libé-Philippot (VIB-KU Leuven Center for Brain & Disease Research) ‘Study of the role of hominid-specific genes in the development and function of human cortical neurons’
Idoia Quintana Urzainqui (EMBL, Heidelberg) ‘Studying the shark embryo to understand the origin and evolution of telencephalic cell types’
Leo Otsuki (Institute of Molecular Pathology) ‘Editing positional memory alters signalling centre output during axolotl limb regeneration’
Wallace is endlessly fascinating. The magnitude of his scientific contributions, where he came from, what he did to make those achievements… That in itself makes him quite remarkable, right?
In the latest episode of the Genetics Unzipped podcast, we’re exploring the epic life of Alfred Russel Wallace; adventurer, naturalist and co-discoverer of natural selection. Biologist and author, Jim Costa, recounts the life of this often forgotten founding father of evolution.
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
Yesterday, we went through a couple of examples of amphibian tail regeneration. Limb regeneration is another example of epimorphic regeneration, which is expertly implemented by some amphibians. Unlike the tail, the limb is patterned in three dimensions: anterior-posterior, ventral-distal and media-proximal to produce the proper number and position of the digits, making the patterning process more complex (McQueen and Towers, 2020). But the potential to – one day – regenerate lost human limbs makes limb regeneration a tantalising subject for researchers and science-fiction writers alike (Cox et al., 2019).
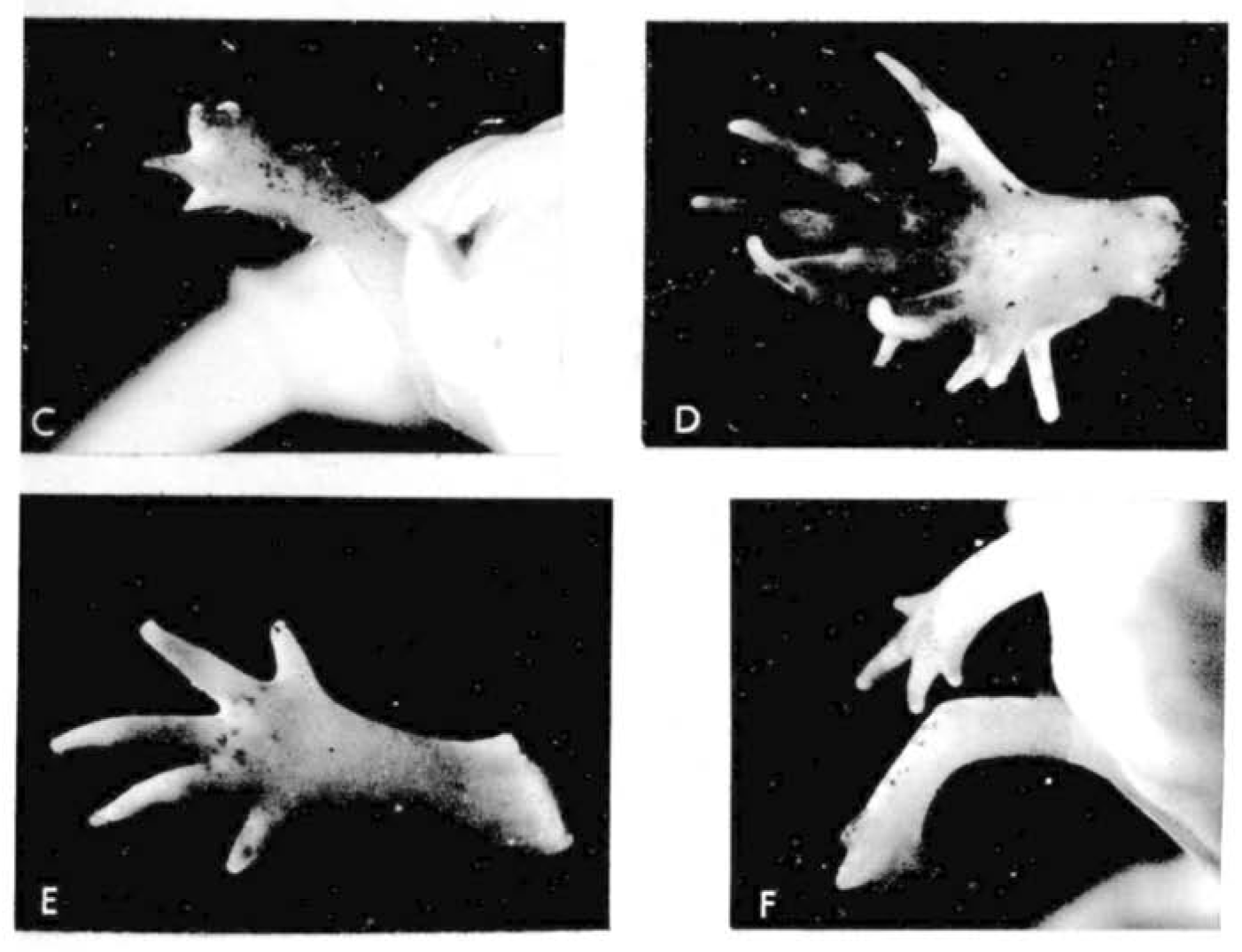
Let us return to the work of our old friend Dr D. R. Newth. In the previous post, we learnt how regeneration could produce a tail more perfect than the original; there’s a similar theme in today’s flashback. A few months after the 1958 publication on tails, Newth posted the second half of his work in JEEM/Development on Triturus helveticus newt limbs (Newth, 1958). There seemed to be an abundance at the time of biologists researching the regenerative abilities of ectopic (supernumerary) limbs by grafting bits of stump tissue, as well as other bits. Such transplants usually produced abnormal limbs with too few or too many digits, which were occasionally resolved during regeneration but there was a lot of conflicting evidence bouncing between journals at the time. One peculiar observation from the past literature is that grafting otic vesicles into the flank induced ectopic limb growth. Newth took a similar approach, engrafting nasal placodes into the body wall to produce an extra foreleg – if anyone has any idea about the mechanism behind placode-dependent limb outgrowth, please do let me know in the comments! This induction approach was quite inefficient (only 19/163 newts grew ectopic limbs) and, like previous reports, these limbs were abnormal with defects ranging from digit-less stumps to polydactyly (Fig. 1). Amputating these defective limbs, however, caused the regeneration of more ‘normal’ looking limbs, with a second amputation of the regenerated limb further improving the phenotype. For example, an ectopic limb that developed with 12 digits, regenerated eight digits following the first amputation and four digits (the normal number) following the amputation of the regenerated limb (Fig. 1). Newth owed these successful regeneration experiments to the amount of limb material, arguing that grafting an inducing centre to form a whole limb was a better approach than grafting stump tissue and provided more material for precise amputation. Again, the position of amputation was deemed significant, but the fascinating mechanism behind this phenomenon remained a mystery.
Fig. 1. Plate 1 from Newth et al., 1958. D shows the original ectopic limb with 10 digits. E shows the regenerated five-fingered limb. F shows the second regenerated limb with, supposedly, four digits.
The limb development and regeneration field has really flourished in the past 65 years and is impossible to adequately summarise here. Evidence, largely from chick and mice, has revealed the many genetic, epigenetic and molecular regulators that control the position of limb budding, outgrowth and digit patterning (McQueen and Towers, 2019). The field has moved in all directions, investigating the roles of the microenvironment, immune system and progenitor populations, as well as their differentiation and lineages, using genome editing, high-resolution imaging and next-generation sequencing technologies. Grafting remains an extremely useful experimental approach (e.g. Vierira et al., 2023) with salamanders, and the axolotl in particular, the main model systems for limb regeneration experiments (Joven et al., 2019). Like in the 1950s, cross-species comparisons are crucial for understanding how tissue regeneration evolved, the factors that control it, and for identifying approaches to induce regeneration in non-regenerative species.
Takeuchi and colleagues published their paper investigating the role of Hox genes during new limb development regeneration last year (Takeuchi et al., 2022; Fig. 2). Their work follows studies in mammals, which have shown that 5′ Hox genes (e.g. Hox13) are crucial for limb development, but whether this requirement is conserved in other tetrapods has not been clear. Takeuchi and team show that Hox13 genes Hoxa13, Hoxc13 and Hoxd13, are expressed both in developing limbs and in blastemas of the newt Pleurodeles waltl. Using CRISPR/Cas9 to knockout Hoxa13, they reveal that Hoxa13 is required for digit formation with the mutants having fewer digits than wild type. Hoxc13 and Hoxd13, however, seem to be dispensable but overexpression of Hoxd13 in Hoxa13 knockout newts partially rescues limb development; Hoxd13 expression in limb buds is strongly dependent on Hoxa13a. Lastly, the authors show that, following the amputation of the forelimbs, Hox13 genes, and Hoxa13 in particular, are also required for digit formation during limb regeneration.
Fig. 2. Movie 1 taken from Takeuchi et al, 2022 showing regeneration of a wild type limb in Pleurodeles waltl.
Both of the articles discussed today concern digit formation in regenerating newt limbs. D.R. Newth showed that regeneration can rescue proper digit development in abnormal limbs and, given that Hox genes have specific positional expression patterns, the work from Takeuchi and colleagues suggests that Hox13 genes might be one of the factors involved in this process. Furthermore, Takeuchi and colleagues demonstrate another parallel, but not identical, link between development and regeneration. Tomorrow, in the final post of this mini-series, we look towards the nervous system.
References
Ben D. Cox, Maximina H. Yun, Kenneth D. Poss; Can laboratory model systems instruct human limb regeneration? Development 15 October 2019; 146 (20): dev181016. doi: https://doi.org/10.1242/dev.181016
Alberto Joven, Ahmed Elewa, András Simon; Model systems for regeneration: salamanders. Development 15 July 2019; 146 (14): dev167700. doi: https://doi.org/10.1242/dev.167700
Caitlin McQueen, Matthew Towers; Establishing the pattern of the vertebrate limb. Development 1 September 2020; 147 (17): dev177956. doi: https://doi.org/10.1242/dev.177956
D. R. Newth; On Regeneration after the Amputation of Abnormal Structures: II. Supernumerary Induced Limbs. Development 1 September 1958; 6 (3): 384–392. doi: https://doi.org/10.1242/dev.6.3.384
Takashi Takeuchi, Haruka Matsubara, Fumina Minamitani, Yukio Satoh, Sayo Tozawa, Tomoki Moriyama, Kohei Maruyama, Ken-ichi T. Suzuki, Shuji Shigenobu, Takeshi Inoue, Koji Tamura, Kiyokazu Agata, Toshinori Hayashi; Newt Hoxa13 has an essential and predominant role in digit formation during development and regeneration. Development 1 March 2022; 149 (5): dev200282. doi: https://doi.org/10.1242/dev.200282
W.A. Vieira, M. Raymond, K. Kelley, M. A. Cherubino, H. Sahin, C. D. McCusker; Integration failure of regenerated limb tissue is associated with incongruencies in positional information in the Mexican axolotl. Front. Cell Dev. Biol. 2023; 11:1152510. doi: https://doi.org/10.3389/fcell.2023.1152510
Hello, I am Tzer Han and I am very happy to introduce myself as a contributor to the ‘New PI Diaries’. I am starting my lab at the Physics Department of UC San Diego, with a broad interest in understanding how symmetry and order emerge in living matter. We are currently focusing on two main directions: (i) using organoids to study tissue mechanics and chart the morphological landscape of multicellular systems; and (ii) using marine invertebrates to study developmental patterning, cilia dynamics and collective behavior. If any of this sounds interesting to you, feel free to check out my website here.
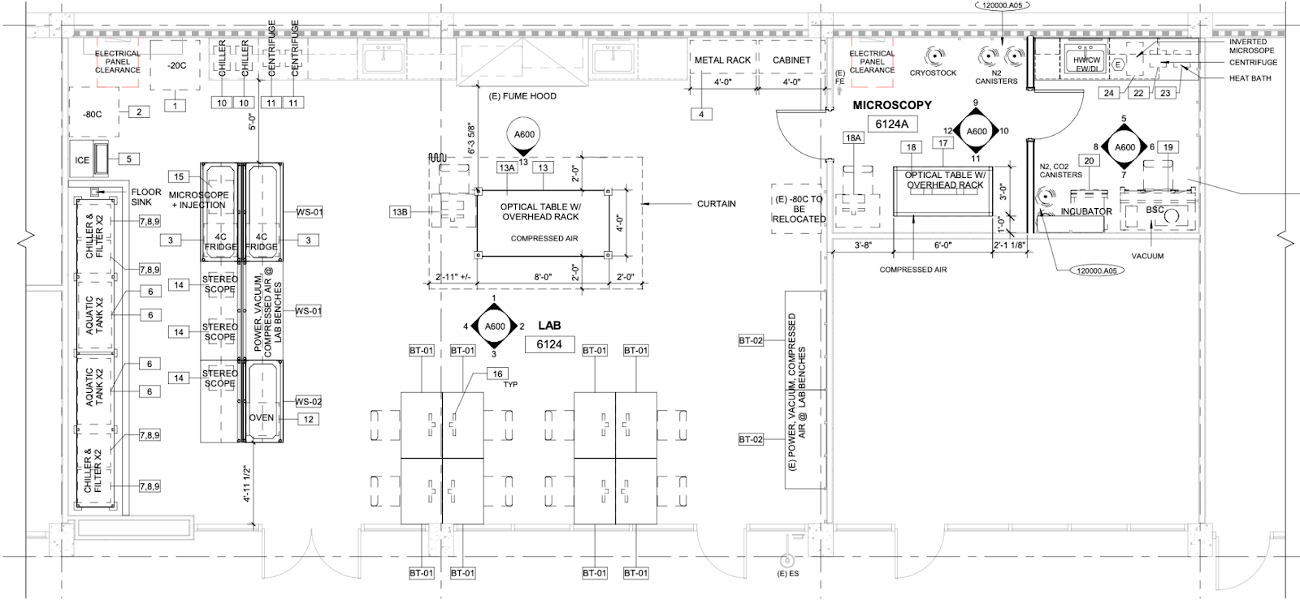
No doubt, becoming a new PI is rewarding but also challenging. Many tasks to be dealt with are unexpected: no one warned you about them and you have exactly one chance to get everything right. One of these tasks, if you are an experimentalist like me, is designing the lab space that suits your experimental needs. Beyond the aspects with which most of us are familiar with, like allocating benches and equipment spaces, there are infrastructures hidden from plain view that matter as much: Where to put the HEPA filter to ensure clean airflow? Should the optical table be on active or passive damping? What architectural precautions to take in case the 75 gallons aquarium tank starts leaking over a long weekend? So, after many rounds of back and forth with the design architects, electrical and mechanical engineers, safety inspectors, I am excited to see the lab design finally taking shape. It will be a few more months before the bidding, contract award, fire marshall approval and actual construction happen. But I will consider this a win for the moment.
What is it like starting your own lab? What exactly needs to be done? In this ‘New PI diaries’ blog series, we will follow the journey of a few new PIs over the course of their first year of starting their research group, giving the Node readers an inside view of the challenges and highlights of being a new PI.
In the coming months, we will be hearing from three upcoming new PIs: Tzer Han Tan, Margot Smit and Elena Camacho. Watch this space!
Tzer Han has just started his lab at University of California, San Diego, looking at how order, symmetry and dynamics emerge in living matter. You can read his first diary entry here.
Margot’s lab at the University of Tübingen, Germany, will be studying plant cell identity and development, in particular, how plants control the timing of fate transitions.
Elena will be starting her group at Centro Andaluz de Biología del Desarrollo in Spain, investigating human embryonic development by leveraging stem cells and mathematics.
Will you be starting your own lab soon? The Node is especially looking for new PIs who will be setting up their lab somewhere outside of Europe and the US. Get in touch if you are interested!
Epimorphic regeneration, the process of replacing lost appendages, is probably the most impressive example of tetrapod vertebrate regeneration. Vertebrate appendages, such as limbs and tails, are composed of many different cell types from different germ layers (including, for example, neurons, skin, muscle, bone and endothelium), which require the complex integration of positional information and patterning to regenerate functional structures (Cox et al., 2019). Amphibians are probably the best-known executors of epimorphic regeneration, with the pink, smiley-faced axolotl intriguing scientists since the 16th century (Joven et al., 2019). Even five centuries later, researchers are continuing to study amphibians to understand epimorphic regeneration. While studies have revealed the importance of blastema and a handful of signalling factors, there’s still a great deal about epimorphic regeneration that remains to be solved. Today, I summarise insights from the following two articles:
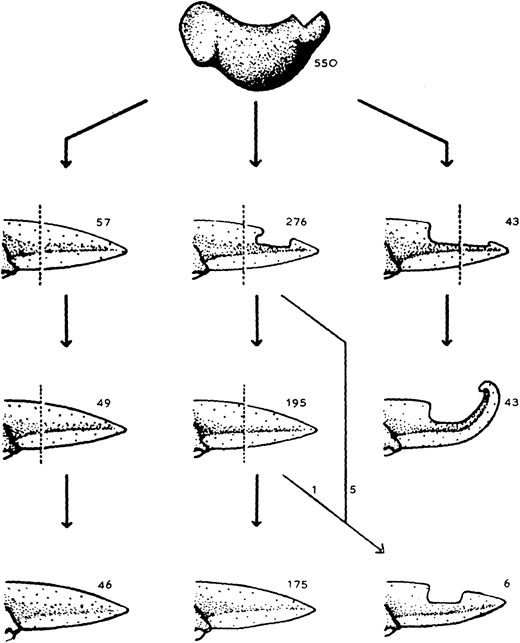
The almost-aptly-amphibian named David Richmond Newth was a Birmingham-born zoologist. After serving in India and Burma during the Second World War, he returned to the UK as a lecturer at Univesity College London. In his 1958 JEEM/Development publication (Newth, 1958), David sought to clarify some previous studies asking whether a regenerated amphibian tail would resemble the original: can animals restore defective appendages and regenerate structures that they didn’t have in the first place? Newth used both axolotl and the frog Rana temporaria as experimental systems, and operated on the embryos to induce the animals to develop an incomplete tail fin. In the frogs, he removed the dorsal half of the tail bud, whereas in axolotl he manipulated posterior neural folds to create ventral fin defects. Next, Newth amputated the tail at different positions along the anterior-posterior axis, cutting anterior to the defect or within the missing tissue. In the case of the Rana tadpoles, David repeated the amputation again (Fig. 1). As long as the amputation occurred anterior to the missing tail tissue, the amphibians could regenerate whole tails but dissection through the missing tissue did not generate complete tail outgrowth from the wound (Fig. 1). By showing that regeneration could form structures that were missing in the original appendage, this work indicates that embryonic development and regeneration might use different mechanisms of growth and patterning. However, the author is careful to point out that, in this case, the mechanism of regeneration in normal animals might not be the same as ‘abnormal’ ones; regeneration of ‘normal’ tissue could still follow developmental mechanisms. Second, this study demonstrates the importance of the position of amputation and indicates that the cells that form the blastema dictate which structures can be formed during regrowth.
Fig. 1. “The regeneration of tails in Rana tadpoles which had earlier lost the dorsal half of their tail-buds. The numbers are of animals surviving to the stages indicated.” Taken from Fig. 3 in Newth (1958).
It seems that the Axolotl is a species that has truly stood the test of time and is just as popular a model today as 500 years ago (Joven et al., 2019). However, technical scientific advances have advanced the toolbox available for asking complex research questions, including the generation of sophisticated axolotl genetic lines and reporters (e.g. Duerr et al., 2022). Although a wide range of frog species have been used in classical studies, present research largely concentrates on Xenopus spp. (Phipps et al., 2020). Whether regeneration employs the same development mechanisms to produce tissues during development remains unclear. Similar developmental signalling pathways (e.g. FGF, Shh) are involved in regenerating appendages; however, single-cell technologies are revealing that the progenitors deriving from the regeneration-specific organising centre – the blastema – have distinct molecular identities from those in the embryo and follow convergent, but different, trajectories (Čapek and Müller, 2019; Tsai et al., 2019). As we heard for wound healing yesterday, many recent investigations have focused on the roles of immune cells in amphibian tail regeneration, showing that myeloid cells and the factors they secrete are required for a regeneration-permissive environment (e.g. Tsai et al., 2019; Aztekin et al., 2020).
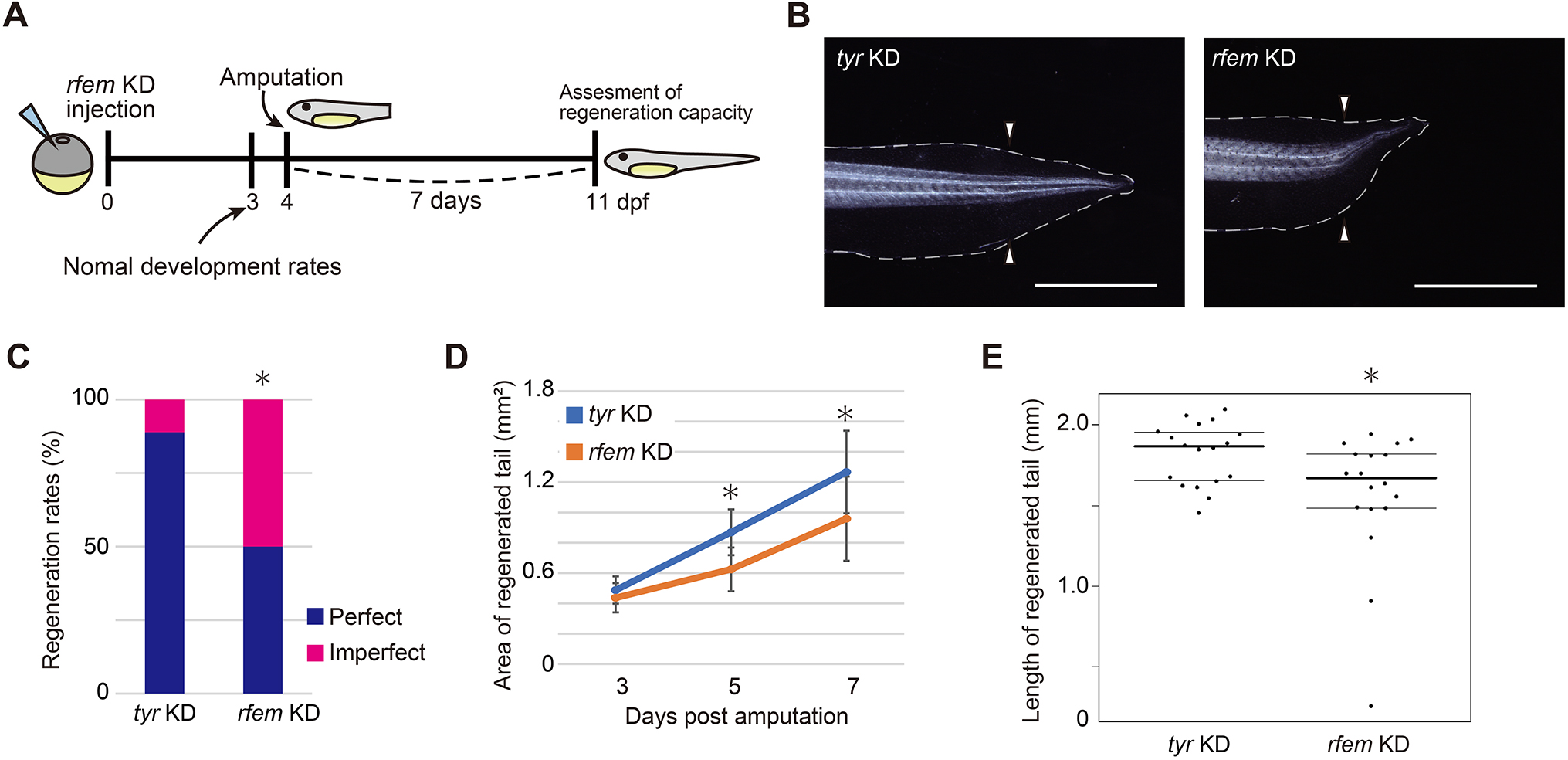
In their article published last month in Development, Deguchi and colleagues continue interrogating the intervention of the immune system in Xenopus laevis tail regeneration (Deguchi et al., 2023). Through a screen to identify cells that support progenitor proliferation, they find that regeneration factors expressed on myeloid (of which there are two paralogues in Xenopus, hereinafter collectively referred to as rfem) increases expression following tail amputation. It’s also required for regeneration because knockdown of rfem generates significantly shorter regenerated tails (Fig. 2). Using RNA sequencing of tail stumps and intact tails, the authors show that rfem-expressing cells cluster with leukocyte markers expressed by macrophages and dendritic cells. Deguchi and colleagues reduce the macrophage population, and thus rfem-expressing cells, by knocking down key monocyte-lineage gene csf1, which also caused tail regeneration abnormalities. Conversely, the addition of rfem-expressing macrophage-like cells in rfem knockdown animals rescues tail regeneration. Although the mechanism of Rfem activity is yet to be elucidated, this study builds upon previous research by identifying a factor through which immune cells might regulate epimorphic regeneration (Aztekin et al., 2020).
Fig. 2. “rfem.L and/or rfem.S are required for normal tail regeneration.” Taken from Fig. 2 in Deguchi et al., 2023 (see publication for full legend).
These two examples from the past and present both ask about the regenerative potential of the amphibian tail. Newth shows that regenerative programs can restore tissues that are effectively formed during development and aren’t possessed by the original structure and indirectly hints about various progenitor populations that might be involved. Deguchi and colleagues take this further to demonstrate that supporting cells, most likely macrophages, have an important role in regulating tail regeneration through the action of rfem-encoded proteins. However, this tale is far from over! Tomorrow, we indulge in a second helping of epimorphic regeneration, focusing on a dexterous but incredibly handy experimental technique: tissue grafts.
References
Can Aztekin, Tom W. Hiscock, Richard Butler, Francisco De Jesús Andino, Jacques Robert, John B. Gurdon, Jerome Jullien; The myeloid lineage is required for the emergence of a regeneration-permissive environment following Xenopus tail amputation. Development 1 February 2020; 147 (3): dev185496. doi: https://doi.org/10.1242/dev.185496
Daniel Čapek, Patrick Müller; Positional information and tissue scaling during development and regeneration. Development 15 December 2019; 146 (24): dev177709. doi: https://doi.org/10.1242/dev.177709
Ben D. Cox, Maximina H. Yun, Kenneth D. Poss; Can laboratory model systems instruct human limb regeneration? Development 15 October 2019; 146 (20): dev181016. doi: https://doi.org/10.1242/dev.181016
Momoko Deguchi, Taro Fukazawa, Takeo Kubo; regeneration factors expressed on myeloid expression in macrophage-like cells is required for tail regeneration in Xenopus laevis tadpoles. Development 1 August 2023; 150 (15): dev200467. doi: https://doi.org/10.1242/dev.200467
Timothy J. Duerr, Eun Kyung Jeon, Kaylee M. Wells, Antonio Villanueva, Ashley W. Seifert, Catherine D. McCusker, James R. Monaghan; A constitutively expressed fluorescent ubiquitination-based cell-cycle indicator (FUCCI) in axolotls for studying tissue regeneration. Development 15 March 2022; 149 (6): dev199637. doi: https://doi.org/10.1242/dev.199637
Alberto Joven, Ahmed Elewa, András Simon; Model systems for regeneration: salamanders. Development 15 July 2019; 146 (14): dev167700. doi: https://doi.org/10.1242/dev.167700
D. R. Newth; On Regeneration after the Amputation of Abnormal Structures: I. Defective Amphibian Tails. Development 1 June 1958; 6 (2): 297–307. doi: https://doi.org/10.1242/dev.6.2.297
Lauren S. Phipps, Lindsey Marshall, Karel Dorey, Enrique Amaya; Model systems for regeneration: Xenopus. Development 15 March 2020; 147 (6): dev180844. doi: https://doi.org/10.1242/dev.180844
Stephanie L. Tsai, Clara Baselga-Garriga, Douglas A. Melton; Blastemal progenitors modulate immune signaling during early limb regeneration. Development 1 January 2019; 146 (1): dev169128. doi: https://doi.org/10.1242/dev.169128
Carlo Donato Caiaffa, Yogeshwari S. Ambekar, Manmohan Singh, Ying Linda Lin, Bogdan Wlodarczyk, Salavat R. Aglyamov, Giuliano Scarcelli, Kirill V. Larin, Richard Finnell
Andrew Liu, Jessica O’Connell, Farley Wall, Richard W. Carthew
Dynamics of Ds and Fat protein distributions across the Drosophila wing pouch from Liu et al. This image is made available under a CC-BY-NC 4.0 International license.
Eduardo D. Gigante, Katarzyna M. Piekarz, Alexandra Gurgis, Leslie Cohen, Florian Razy-Krajka, Sydney Popsuj, Hussan S. Ali, Shruthi Mohana Sundaram, Alberto Stolfi
Cell division and differentiation in C. robusta embryos from Gigante et al. This image is made available under a CC-BY-NC 4.0 International license.
Marcia Mesquita Peixoto, Francisca Soares-da-Silva, Valentin Bonnet, Gustave Ronteix, Rita Faria Santos, Marie-Pierre Mailhe, Xing Feng, João Pedro Pereira, Emanuele Azzoni, Giorgio Anselmi, Marella de Bruijn, Charles N. Baroud, Perpétua Pinto-do-Ó, Ana Cumano
Juan Manuel Gomez, Hendrik Nolte, Elisabeth Vogelsang, Bipasha Dey, Michiko Takeda, Girolamo Giudice, Miriam Faxel, Alina Cepraga, Robert Patrick Zinzen, Marcus Krüger, Evangelia Petsalaki, Yu-Chiun Wang, Maria Leptin
Yogeshwari S. Ambekar, Carlo Donato Caiaffa, Bogdan Wlodarczyk, Manmohan Singh, Alexander W. Schill, John Steele, Salavat R. Aglyamov, Giuliano Scarcelli, Richard H. Finnell, Kirill V. Larin
Akanksha Jain, Gilles Gut, Fátima Sanchís Calleja, Ryoko Okamoto, Simon Streib, Zhisong He, Fides Zenk, Malgorzata Santel, Makiko Seimiya, René Holtackers, Sophie Martina Johanna Jansen, J. Gray Camp, Barbara Treutlein
Cora Demler, John Coates Lawlor, Ronit Yelin, Dhana Llivichuzcha-Loja, Lihi Shaulov, David Kim, Megan Stewart, Frank Lee, Thomas Schultheiss, Natasza Kurpios
Simon Schneider, Andjela Kovacevic, Michelle Mayer, Ann-Kristin Dicke, Lena Arévalo, Sophie A. Koser, Jan N. Hansen, Samuel Young, Christoph Brenker, Sabine Kliesch, Dagmar Wachten, Gregor Kirfel, Timo Strünker, Frank Tüttelmann, Hubert Schorle
Paula Cubillos, Nora Ditzer, Annika Kolodziejczyk, Gustav Schwenk, Janine Hoffmann, Theresa M. Schütze, Razvan P. Derihaci, Cahit Birdir, Johannes E. M. Köllner, Andreas Petzold, Mihail Sarov, Ulrich Martin, Katherine R. Long, Pauline Wimberger, Mareike Albert
A. Lardenois, A. Suglia, CL. Moore, B. Evrard, L. Noël, P. Rivaud, A. Besson, M. Toupin, S. Léonard, L. Lesné, I. Coiffec, S. Nef, V. Lavoué, O. Collin, A. Chédotal, S. Mazaud-Guittot, F. Chalmel, AD. Rolland
Joab Camarena, Sofia E. Luna, Jessica P. Hampton, Kiran R. Majeti, Carsten T. Charlesworth, Eric Soupene, Vivien A. Sheehan, M. Kyle Cromer, Matthew H. Porteus
Michał Brouze, Agnieszka Czarnocka-Cieciura, Olga Gewartowska, Monika Kusio-Kobiałka, Kamil Jachacy, Marcin Szpila, Bartosz Tarkowski, Jakub Gruchota, Paweł Krawczyk, Seweryn Mroczek, Ewa Borsuk, Andrzej Dziembowski
Prakhar Bansal, Erin C Banda, Heather R Glatt-Deeley, Christopher Stoddard, Jeremy W Linsley, Neha Arora, Darcy T Ahern, Yuvabharath Kondaveeti, Michael Nicouleau, Miguel Sabariego-Navarro, Mara Dierssen, Steve Finkbeiner, Stefan F Pinter
Jos Smits, Dulce Lima Cunha, Jieqiong Qu, Nicholas Owen, Lorenz Latta, Nora Szentmary, Berthold Seitz, Lauriane N. Roux, Mariya Moosajee, Daniel Aberdam, Simon J. van Heeringen, Huiqing Zhou
Lucas G. A. Ferreira, Marina M. L. Kizys, Gabriel A. C. Gama, Svenja Pachernegg, Gorjana Robevska, Andrew H. Sinclair, Katie L. Ayers, Magnus R. Dias da Silva
Temperature control and simultaneous monitoring of transcriptional activity from Martini and Garcia. This image is made available under a CC-BY-ND 4.0 International license.
Prakhar Bansal, Erin C Banda, Heather R Glatt-Deeley, Christopher Stoddard, Jeremy W Linsley, Neha Arora, Darcy T Ahern, Yuvabharath Kondaveeti, Michael Nicouleau, Miguel Sabariego-Navarro, Mara Dierssen, Steve Finkbeiner, Stefan F Pinter
Alexa Giovannini, Sabrina Piechota, Maria Marchante, Kathryn S Potts, Graham Rockwell, Bruna Paulsen, Alexander D Noblett, Samantha L Estevez, Alexandra B Figueroa, Caroline Aschenberger, Dawn A Kelk, Marcy Forti, Shelby Marcinyshyn, Ferran Barrachina, Klaus Wiemer, Marta Sanchez, Pedro Belchin, Merrick Pierson Smela, Patrick R.J. Fortuna, Pranam Chatterjee, David H McCulloh, Alan Copperman, Daniel Ordonez-Perez, Joshua U Klein, Christian C Kramme
Wai Kit Chan, Danilo Negro, Victoria M Munro, Helen Marshall, Zrinko Kozić, Megan Brown, Mariana Beltran, Neil C Henderson, David J Price, John O Mason
Mercedes Barrero, Anna V. López-Rubio, Aleksey Lazarenkov, Enrique Blanco, Moritz Bauer, Luis G. Palma, Anna Bigas, Luciano Di Croce, José Luis Sardina, Bernhard Payer
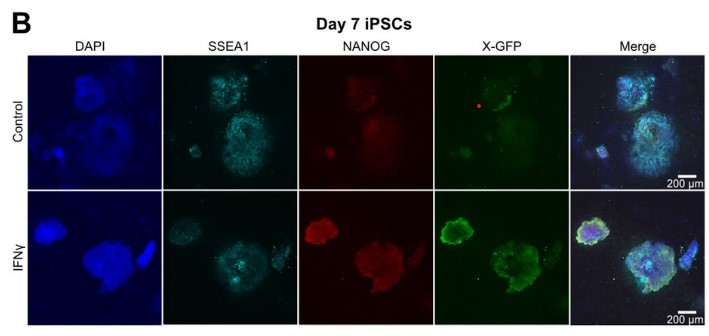
iPSC colonies upon early interferon γ treatment from Barrero et al. This image is made available under a CC-BY-NC 4.0 International license.
Tammy T Nguyen, Zinger Yang Loureiro, Anand Desai, Tiffany DeSouza, Shannon Joyce, Lyne Khair, Amruta Samant, Haley Cirka, Javier Solivan-Rivera, Rachel Ziegler, Michael Brehm, Louis M Messina, Silvia Corvera
Xiaoyu Yang, Ian R. Jones, Poshen B. Chen, Han Yang, Xingjie Ren, Lina Zheng, Bin Li, Yang E. Li, Quan Sun, Jia Wen, Cooper Beaman, Xiekui Cui, Yun Li, Wei Wang, Ming Hu, Bing Ren, Yin Shen
Diana R. Rodríguez-García, Yossmayer del Carmen Rondón Guerrero, Lucía Ferrero, Andrés Hugo Rossi, Esteban A. Miglietta, Ariel A. Aptekmann, Eliana Marzol, Javier Martínez Pacheco, Mariana Carignani, Victoria Berdion Gabarain, Leonel E. Lopez, Gabriela Díaz Dominguez, Cecilia Borassi, José Juan Sánchez-Serrano, Lin Xu, Alejandro D. Nadra, Enrique Rojo, Federico Ariel, José M. Estevez
Craig L. Cowling, Arielle L. Homayouni, Jodi B. Callwood, Maxwell R. McReynolds, Jasper Khor, Haiyan Ke, Melissa A. Draves, Katayoon Dehesh, Justin W. Walley, Lucia C. Strader, Dior R. Kelley
Matēj Drs, Samuel Haluška, Eliška Škrabálková, Pavel Krupař, Andrea Potocká, Lucie Brejšková, Karel Muller, Natalia Serrano, Aline Voxeur, Samantha Vernhettes, Jitka Ortmannová, George Caldarescu, Matyas Fendrych, Martin Potocký, Viktor Žárský, Tamara Pečenková
Sebastjen Schoenaers, Hyun Kyung Lee, Martine Gonneau, Elvina Faucher, Thomas Levasseur, Elodie Akary, Naomi Claeijs, Steven Moussu, Caroline Broyart, Daria Balcerowicz, Hamada AbdElgawad, Andrea Bassi, Daniel Santa Cruz Damineli, Alex Costa, Jose A Feijo, Celine Moreau, Estelle Bonnin, Bernard Cathala, Julia Santiago, Herman Hofte, Kris Vissenberg
Simon Amiard, Léa Feit, Lauriane Simon, Samuel Le Goff, Loriane Loizeau, Léa Wolff, Falk Butter, Clara Bourbousse, Fredy Barneche, Christophe Tatout, Aline. V. Probst
Linkan Dash, Maxwell R. McReynolds, Melissa A. Draves, Rajdeep S. Khangura, Rebekah L. Muench, Jasper Khor, Jodi B. Callwood, Craig L. Cowling, Ludvin Mejia, Michelle G. Lang, Brian P. Dilkes, Justin W. Walley, Dior R. Kelley
Avilash S. Yadav, Lilan Hong, Patrick M. Klees, Annamaria Kiss, Xi He, Iselle M. Barrios, Michelle Heeney, Anabella Maria D. Galang, Richard S. Smith, Arezki Boudaoud, Adrienne H.K. Roeder
David Bolumar, Javier Moncayo-Arlandi, Javier Gonzalez-Fernandez, Ana Ochando, Inmaculada Moreno, Ana Monteagudo-Sanchez, Carlos Marin, Antonio Diez, Paula Fabra, Miguel Ángel Checa, Juan José Espinos, David K. Gardner, Carlos Simon, Felipe Vilella
Isabel Tundidor, Marta Seijo-Vila, Sandra Blasco-Benito, María Rubert-Hernández, Gema Moreno-Bueno, Laura Bindila, Rubén Fernández de la Rosa, Manuel Guzmán, Cristina Sánchez, Eduardo Pérez-Gómez
Khursheed Iqbal, Brandon Nixon, Benjamin Crnkovich, Esteban M. Dominguez, Ayelen Moreno-Irusta, Regan L. Scott, Ha T.H. Vu, Geetu Tuteja, Jay L. Vivian, Michael J. Soares
Jesús A. Andrés-San Román, Carmen Gordillo-Vázquez, Daniel Franco-Barranco, Laura Morato, Cecilia H. Fernández-Espartero, Gabriel Baonza, Antonio Tagua, Pablo Vicente-Munuera, Ana M. Palacios, Maria P. Gavilan, Fernando Martín-Belmonte, Valentina Annese, Pedro Gómez-Gálvez, Ignacio Arganda-Carreras, Luis M. Escudero
Nitesh Kumar Sharma, Ram Ayyala, Dhrithi Deshpande, Yesha M Patel, Viorel Munteanu, Dumitru Ciorba, Andrada Fiscutean, Mohammad Vahed, Aditya Sarkar, Ruiwei Guo, Andrew Moore, Nicholas Darci-Maher, Nicole A Nogoy, Malak S. Abedalthagafi, Serghei Mangul

 (1 votes)
(1 votes) (No Ratings Yet)
(No Ratings Yet)