It allows replicating the figures presented in our article using your favorite gene(s), without the need to implement bioinformatic analyses. If you are interested in pallium development and want to visualise the expression of specific genes in progenitors and neurons, explore their spatial and temporal variations or compare ventral and dorsal differentiation trajectories, this App is for you. We hope the community of developmental neurobiologists will find it useful and convenient.
Integrative analysis of the 3D genome and epigenome in mouse embryonic tissues Miao Yu, Nathan R. Zemke, Ziyin Chen, Ivan Juric, Rong Hu, Ramya Raviram, Armen Abnousi, Rongxin Fang, Yanxiao Zhang, David U. Gorkin, Yang Li, Yuan Zhao, Lindsay Lee, Anthony D. Schmitt, Yunjiang Qiu, Diane E. Dickel, Axel Visel, Len A. Pennacchio, Ming Hu, Bing Ren
HOPX governs a molecular and physiological switch between cardiomyocyte progenitor and maturation gene programs Clayton E. Friedman, Seth W. Cheetham, Richard J. Mills, Masahito Ogawa, Meredith A. Redd, Han Sheng Chiu, Sophie Shen, Yuliangzi Sun, Dalia Mizikovsky, Romaric Bouveret, Xiaoli Chen, Holly Voges, Scott Paterson, Jessica E. De Angelis, Stacey B. Andersen, Sohye Yoon, Geoffrey J. Faulkner, Kelly A. Smith, Richard P. Harvey, Benjamin M. Hogan, Quan Nguyen, Kazu Kikuchi, James E. Hudson, Nathan J. Palpant
Comprehensive multiomic profiling of somatic mutations in malformations of cortical development Changuk Chung, Xiaoxu Yang, Taejeong Bae, Keng Ioi Vong, Swapnil Mittal, Catharina Donkels, H. Westley Phillips, Ashley P. L. Marsh, Martin W. Breuss, Laurel L. Ball, Camila Araújo Bernardino Garcia, Renee D. George, Jing Gu, Mingchu Xu, Chelsea Barrows, Kiely N. James, Valentina Stanley, Anna Nidhiry, Sami Khoury, Gabrielle Howe, Emily Riley, Xin Xu, Brett Copeland, Yifan Wang, Se Hoon Kim, Hoon-Chul Kang, Andreas Schulze-Bonhage, Carola A. Haas, Horst Urbach, Marco Prinz, Corrine Gardner, Christina A. Gurnett, Shifteh Sattar, Mark Nespeca, David D. Gonda, Katsumi Imai, Yukitoshi Takahashi, Robert Chen, Jin-Wu Tsai, Valerio Conti, Renzo Guerrini, Orrin Devinsky, Wilson A. Silva Jr, Helio R. Machado, Gary W. Mathern, Alexej Abyzov, Sara Baldassari, Stéphanie Baulac, Focal Cortical Dysplasia Neurogenetics Consortium, Brain Somatic Mosaicism Network, Joseph G. Gleeson
Lessons on fruiting body morphogenesis from genomes and transcriptomes of Agaricomycetes László G. Nagy, Peter Jan Vonk, Markus Künzler, Csenge Földi, Máté Virágh, Robin A. Ohm, Florian Hennicke, Balázs Bálint, Árpád Csernetics, Botond Hegedüs, Zhihao Hou, Xiao-Bin Liu, Shen Nan, Manish Pareek, Neha Sahu, Benedek Szathmári, Torda Varga, Hongli Wu, Xiao Yang, Zsolt Merényi
p73 controls cell junction dynamics during sprouting angiogenesis and acts via Angiomotin Laura Maeso-Alonso, Hugo Alonso-Olivares, Nicole Martínez-García, Javier Villoch-Fernández, Laura Puente-Santamaría, Natalia Colas-Algora, Alfonso Fernández-Corona, María Elena Lorenzo-Marcos, Benilde Jiménez, Lars Holmgren, Margareta Wilhelm, Jaime Millan, Luis del Peso, Lena Claesson-Welsh, Margarita M. Marques, Maria C. Marin
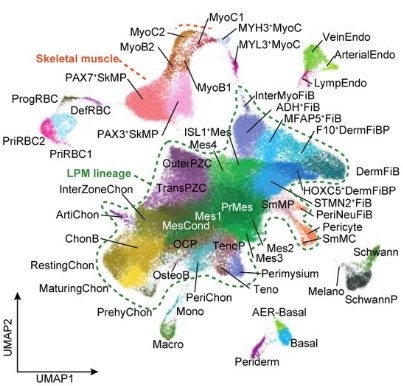
UMAP visualisation of human embryonic limb cells from Zhang, et al.
A human embryonic limb cell atlas resolved in space and time Bao Zhang, Peng He, John E Lawrence, Shuaiyu Wang, Elizabeth Tuck, Brian Williams, Kenny Roberts, Vitalii Kleshchevnikov, Lira Mamanova, Liam Bolt, Krzysztof Polanski, Rasa Elmentaite, Eirini S Fasouli, Martin Prete, Xiaoling He, Nadav Yayon, Yixi Fu, Hao Yang, Chen Liang, Hui Zhang, David R. FitzPatrick, Helen Firth, Andrew Dean, Roger A Barker, Mekayla A Storer, Barbara J Wold, Hongbo Zhang, Sarah A Teichmann
Unlocking the microblogging potential for science and medicine Aditya Sarkar, Augustin Giros, Louis Mockly, Jaden Moore, Andrew Moore, Anish Nagareddy, Yesha M. Patel, Karishma Chhugani, Varuni Sarwal, Nicholas Darci-Maher, Yutong Chang, Lana X. Garmire, Riyue Bao, Rayan Chikhi, Serghei Mangul
An open-source tool to assess the carbon footprint of research Jérôme Mariette, Odile Blanchard, Olivier Berné, Olivier Aumont, Julian Carrey, Anne Laure Ligozat, Emmanuel Lellouch, Philippe-e Roche, Gäel Guennebaud, Joel Thanwerdas, Philippe Bardou, Gérald Salin, Elise Maigne, Sophie Servan, Tamara Ben-Ari
In their recent Development paper, published in our Immune Special Issue, Manisha Goyal, Tina Mukherjee, and colleagues examine the pathways controlling ROS homeostasis during hematopoietic growth control. They identify an important role for odor-sensing and the GABA pathway in myeloid ROS regulation, which has downstream effects on hematopoietic growth. Now, Manisha gives us some insights into the story behind the paper.
Dr. Mukherjee’s past work shows the importance of olfaction and GABA signaling in blood-progenitor maintenance (Shim et al., 2013). This work always intrigued me and also motivated me to join the lab. I was fortunate to work on another interesting and fascinating work where we show that olfaction-derived GABA catabolism in blood-progenitor cells regulates their immune response to parasitic wasp infections (Madhwal et al., 2020). Knowing the importance of GABA signaling in blood-progenitor homeostasis and the role of GABA catabolism in immune response, we asked about the involvement of this pathway in homeostatic conditions as well.
How did you get started on this project?
The project started with the observation that perturbation of the GABA catabolic pathway in blood-progenitor cells leads to lymph gland growth retardation. Conversely, we found that perturbation of TCA cycle enzymes in blood-progenitor cells doesn’t affect lymph gland growth in homeostasis. This made us curious to begin exploring the role of GABA catabolism and the TCA cycle in blood-progenitor homeostasis.
What was the key experiment?
There has always been an excitement to do the experiments and analyze the results. The findings that GABA catabolism regulates TCA cycle activity and this regulation is specifically acting on phosphorylation of pyruvate dehydrogenase kinase and not on the total enzyme levels were really fascinating to me. This finding was one of the key experiments connecting GABA catabolism, reactive oxygen species (ROS), and TCA cycle regulation. Moreover, working with fruit flies has always been fun and the tiny flies have made exploring the mechanism of complex pathways simple.
When doing the research, did you have any particular result or eureka moment that has stuck with you?
We had some interesting discussions about our results showing that treatment with an antioxidant (N-acetyl cysteine) was sufficient to rescue the lymph gland growth defect and high ROS levels in larvae with a perturbation in the GABA catabolic pathway in blood-progenitor cells (domeMeso>GatRNAi and domeMeso>SsadhRNAi). This experiment was a turning point for the story where we were able to find an independent role/effect of ROS regulation on lymph gland growth and could take the story further.
And what about the flipside: any moments of frustration or despair?
COVID-19 limited the time we could spend in the lab and there were also other constraints on us, labs, and support workers, which heightened the pressure to work. But the excitement to do the experiments and finding the answers helped us to overcome our frustrations.
Where will this story take the lab?
Currently, we are exploring this research further to unravel another aspect of ROS regulation in blood-progenitor cells. This work bridges GABA catabolism and TCA cycle in blood-progenitor cells and given that ROS homeostasis and TCA cycle are regulated by odor-sensing, we are curious to investigate the mechanism and key metabolic changes in homeostasis, as well as in immune response conditions. Hopefully, we will see the outcomes of the work soon.
What is next for you after this paper?
I continue to be a part of this work and I am exploring other mechanisms of ROS regulation by GABA catabolism.
References:
Shim, J., Mukherjee, T., Mondal, B. C., Liu, T., Young, G. C., Wijewarnasuriya, D. P., & Banerjee, U. (2013). Olfactory control of blood progenitor maintenance. Cell, 155(5), 1141–1153. https://doi.org/10.1016/j.cell.2013.10.032
Madhwal, S., Shin, M., Kapoor, A., Goyal, M., Joshi, M. K., Ur Rehman, P. M., Gor, K., Shim, J., & Mukherjee, T. (2020). Metabolic control of cellular immune-competency by odors in Drosophila. eLife, 9, e60376. https://doi.org/10.7554/eLife.60376
“They ruled out nature. They ruled out nurture. There must be something else, and that something else is what I like to call ‘the wobble'”
Dr Kat Arney
In the latest episode of the Genetics Unzipped podcast, presenter Dr Kat Arney explores the importance of randomness in genetics. How can we explain differences between individuals with identical nature and nurture? We look at how Professor Ben Lehner’s worm-breaking research has changed our understanding of epigenetics.
This post highlights the approach and finding of a new research article published by Disease Models and Mechanisms (DMM). This feature is written by Katelyn E. Senkus, MS as a part of a seminar at The University of Alabama (taught by DMM Editorial Board member, Professor Guy Caldwell) on current topics related to use of animal and cellular model systems in studies of human disease.
Neurodegenerative diseases encompass a range of conditions, each with a unique pathophysiology and manifestation of symptoms. Although seemingly disparate, conditions like amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD) share a key underlying feature, TDP-43 abnormalities, and are classified as TDP-43 proteinopathies (de Boer et al., 2020). TDP-43, a nuclear protein encoded by the TARDBP gene, is critical to cellular function through its role in transcriptional repression and exon skipping activation (Ratti and Buratti, 2016). Hallmark features of TDP-43 proteinopathies include cytoplasmic TDP-43 protein aggregations and TDP-43 nuclear depletion. TDP-43 can form cytoplasmic aggregates that deleteriously affect neuronal health, including cellular toxicity, signaling cascade interference, and sequestration of additional TDP-43, thus rendering it inactive. Such aggregates are present in 97% and 50% of ALS and FTLD patients, respectively (de Boer et al., 2020; Jo et al., 2020). Generally, TDP-43 nuclear depletion is considered secondary to TDP-43 aggregation in the onset of ALS. However, could nuclear depletion be a driving factor of disease? A meta-analysis reported that 24% of healthy older adults exhibited TDP-43 aggregations (Nascimiento et al., 2018). Acknowledging that genetic mutations account for only 1-5% of cases, what is different between healthy individuals and their ALS counterparts (de Boer et al., 2020)? Perhaps the answer lies in epigenetic modifications controlling TDP-43 expression.
A recent report in Disease Models & Mechanisms by Pacetti et al., (2022) explored the hypothesis that levels of endogenously synthesized TDP-43 may influence an individual’s ALS risk. Accordingly, the purpose of this study was to characterize TDP-43 levels across tissues throughout the lifespan and to elucidate mechanisms underpinning potential variations in TDP-43 production.
TDP-43 mRNA and protein levels were quantified in various tissues of 10- and 90-day-old mice. Significant reductions in TDP-43 mRNA and protein were observed in brain tissue from 90-day-old mice, but not liver tissue. When extended to 360-day-old mice, no further reductions occurred.
TDP-43 levels are mediated by autoregulatory mechanisms, but this does not fully explain the differential findings by tissue and stability over time. Thus, the effect of epigenetic modifications on TDP-43 production was investigated. In the TARDBP promoter, 113 CpG sites, clustered into four CpG-rich islands, were identified. A significant inverse association between CpG sites 109-113 in island 4 methylation and TDP-43 expression was observed in brain tissue of 90-day-old mice. As anticipated, liver DNA methylation remained unchanged. Findings were substantiated in an in vitro model using mouse motor neuron-derived NSC-34 cells. Following induction of hypomethylation, significant increases in TDP-43 mRNA and protein levels were observed; whereas, hypermethylation significantly reduced promoter activity.
Epigenetic investigation continued with histone modification assessment in an in vivo model. Generally, reduced gene expression is observed with increased H3K27me3 and decreased acH2.A.Z. ChIP-qPCR analysis revealed a 4-fold increase in H3K27me3 and 1.8-fold decrease in acH2.A.Z in brain tissue of 90-day old mice.
To support research translation, experiments were repeated in human SH-SY5Y cells and 125 CpG sites, clustered into three CpG-rich islands, were identified in the TARDBP promoter. Sites in the most upstream island were methylated which is in contrast to the animal findings where methylation occurred at the downstream islands. Nonetheless, hypomethylation and hypermethylation experiments generated similar results.
Collectively, findings support the hypothesis that epigenetic modifications in the TARDBP promoter underpin the differential expression of TDP-43 in mouse tissues at two different ages. Authors postulate that variations in ALS onset may be attributed, in part, to different levels of epigenetic modification of this gene. For example, TDP-43 aggregates in combination with low levels of endogenous TDP-43 production in the brain may predispose an individual to ALS. Whereas, the same concentration of TDP-43 aggregates paired with moderate-to-normal levels of TDP-43 production may prevent onset, as endogenously produced TDP-43 compensates for sequestration by aggregates. Variations in epigenetic modifications were not explored in this study, as inbred animal models generally exhibit homogenous features. However, it is well-established that humans’ environments influence epigenetic processes; thus, the aforementioned presumption is plausible (Alegría-Torres et al., 2013).
To date, potential ALS therapies have targeted TDP-43 aggregates in the brain, including small molecular inhibitors and heat shock proteins (de Boer et al., 2020). Pacetti et al., (2022) proposes a new mechanism of equal importance. Interventions that reduce DNA methylation and/or influence histone modification of the TARDBP promoter may restore TDP-43 functionality and overcome the consequences of TDP-43 aggregates. It is critical to explore if compensation is indefinite or only slightly extends disease-free time. Nevertheless, acknowledging that many neurodegenerative diseases present with TDP-43 abnormalities, the current study results have the potential to significantly impact public health through development of epigenetic-targeted therapies.
REFERENCES
Alegría-Torres, J.A., Baccarelli, A., Bollati, V. (2013). Epigenetics and lifestyle. Epigenomics. 3, 267-277.
de Boer, E.M., Orie, V.K., Williams, T., Baker, M.R., De Oliveira, H.M., Polvikoski, T., Silsy, M., Menon, P., van den Bos, M., Halliday, G.M., et al. (2020). TDP-43 proteinopathies: a new wave of neurodegenerative diseases. J. Neurol. Neurosurg. Psychiatry. 92, 86-95.
Jo, M., Lee, S., Jeon, Y-M., Kim, S., Kwon, Y., Kim, H-J. (2020). The role of TDP-43 propagation in neurodegenerative diseases: integrating insights from clinical and experimental studies. Exp. Molec. Med. 52, 1652-1662.
Nascimiento, C., Di Lorenzo Alho, A., Amaral, C.B.C., Leite, R.E.P., Nitrini, R., Jacob-Filho, W., Pasqualucci, C.A., Hokkanen, S.R.K., Hunter, S., Keage, H., et al. (2018). Prevalence of TDP-43 proteinopathy in cognitively normal older adults: systematic review and meta-analysis. Neuropathol. Appl. Neurobiol. 44, 286-297.
Pacetti, M., De Conti, L., Marasco, L.E., Romano, M., Rashid, M.M., Nubie, M., Baralle, F.E., Baralle, M. (2022). Physiological tissue-specific and age-related reduction of mouse TDP-43 levels is regulated by epigenetic modifications. Dis. Model. Mech.
Ratti, A., Buratti, E. (2016). Physiological functions and pathobiology of TDP-43 and FUS/TLS proteins. J. Neurochem.138, 95-111.
This post highlights the approach and finding of a new research article published by Disease Models and Mechanisms (DMM). This feature is written by Melanie Mew as part of a seminar at The University of Alabama (taught by DMM Editorial Board member, Professor Guy Caldwell) on current topics related to use of animal and cellular model systems in the studies of human disease.
Prions are misfolded proteins that can induce other proteins to fold abnormally and are transmissible from one individual to another. Prion diseases (PrDs) range from sporadic forms of bovine spongiform encephalopathy (“mad cow” disease) to genetically inherited Creutzfeldt-Jacob disease (CJD). They are uniformly fatal and untreatable. The key to preventing and treating PrDs may lie in understanding the structure of prions and how they mediate neurodegeneration. A major challenge to studying PrDs is that they occur only in mammals, whose long generation times and high maintenance costs limit the speed and statistical power of research. Smaller models such as Drosophila are a useful alternative because evolutionary conservation of basic cellular processes allows them to recapitulate PrD phenotypes. A remaining mystery in PrD research is the mechanisms through which prions cause disease. In a recent study published in Disease Models and Mechanisms, Myers et al., (2022) use a transgenic Drosophila model expressing human or rodent prion protein (PrP) to elucidate intrinsic and extrinsic mediators of prion toxicity. The researchers identified a new “glassy eyes” phenotype, associated with reduced size and organization of the eyes, in flies expressing toxic forms of PrP.
An initial investigation of human PrP compared to the less toxic rodent PrP found that, while these proteins share many similarities, amino acid changes in the C-terminal 3D (CT3D) may mediate the greater instability and toxicity of human PrP. Expressing hamster PrP in fruit flies permits normal eye development (Fernandez-Funez et al., 2017), whereas random insertion of human PrP into the Drosophila genome is deleterious to eye development (Myers et al., 2022). Specifically, comparison of two naturally occurring human PrP polymorphisms: Met129 and Val129—the latter associated with associated with a higher probability of developing CJD—induced a glassy-eyed phenotype. In separate experiments, a conditional activator system was used to induce pan-neuronal hamster or human PrP expression in adult flies. These individuals were subjected to a climbing assay, in which the human PrP flies developed locomotor deficits significantly worse than those of hamster PrP-expressing flies. Human PrP expression was also associated with reduced size and structure of Drosophila mushroom bodies, which are non-essential features responsible for learning and memory in flies and other insects.
To directly compare the toxicity of rodent and human PrP, Myers et al. inserted codon-optimized versions of each gene at a previously validated attP2 locus (Bischof et al., 2007, Moore et al., 2018). They created flies expressing either mouse PrP, human PrP-M129, or human PrP-V129. Flies expressing mouse PrP had eyes comparable to those of controls, whereas flies homozygous for either form of human PrP displayed glassy eyes. Having established a clear pattern of toxicity associated with human PrP, the researchers next investigated genetic and molecular mediators of human PrP toxicity. Specific extrinsic mediators of PrP toxicity may include members of the unfolded protein response (UPR) pathway, associated with ER stress. The UPR is activated by various sensors, including Ire1α and PERK. Ire1α and a protein activated downstream of it, X-box binding protein 1 (XBP1), can protect against human PrP toxicity. Silencing either of these proteins alone has no effect on eye development, but produces a small-eye phenotype when combined with human PrP expression. Conversely, silencing PERK or its downstream affiliate, ATF4, independently or in combination with human PrP expression allows for normal eye development. Therefore, activating the PERK branch of the UPR or inhibiting the Ire1α branch could be valuable treatment strategies for PrDs. Other therapeutic options for PrDs could include modulating the structure of PrP itself. In this vein, Myers and colleagues investigated the protective effects of amino acid substitutions found in dogs, horses, rabbits, and pigs. Asp167Ser, the canine residue mutation, was protective against eye disorganization and mushroom body degeneration in flies.
The research performed by Myers and colleagues suggests that human PrDs may be treatable with drugs that stabilize the CT3D of PrP or modulate the UPR response to PrP. Future research will likely further elucidate the interacting partners of human PrP that mediate neurodegeneration, as well as the roles of specific residues in contributing to intrinsic protein stability. Understanding the evolution of human PrP could also help identify benefits of stability-sacrificing human PrP mutations. Microbial prions, for example, have been shown to sometimes confer evolutionary adaptations such as stress tolerance and memory (Levkovich et al., 2021). For now, this work informs our general understanding of neurodegeneration induced by protein aggregation—a hallmark of other disorders such as Parkinson’s disease and Alzheimer’s disease (Soto and Pritzkow, 2018)—and is a step toward discovering effective treatments for neurological diseases.
REFERENCES
Bischof J., Maeda R.K., Hediger M., Karch F. and Basler K. (2007) An optimized transgenesis system for Drosophila using germ-line-specific phiC31 integrases. Proc Natl Acad Sci U S A. 104, 3312-7.
Fernandez-Funez P., Sanchez-Garcia J. and Rincon-Limas D.E. (2017) Drosophila models of prionopathies: insight into prion protein function, transmission, and neurotoxicity. Curr. Opin. Genet. Dev.44, 141-148.
Levkovich S.A., Rencus-Lazar S., Gazit E., Laor Bar-Yosef D. (2021) Microbial Prions: Dawn of a New Era. Trends Biochem. Sci. 46, 391-405.
Moore, B. D., Martin, J., de Mena, L., Sanchez, J., Cruz, P. E., Ceballos-Diaz, C., Ladd, T. B., Ran, Y., Levites, Y., Kukar, T. L., Kurian, J. J., McKenna, R., Koo, E. H., Borchelt, D. R., Janus, C., Rincon-Limas, D., Fernandez-Funez, P. and Golde, T. E. (2018). Short Aβ peptides attenuate Aβ42 toxicity in vivo. J. Exp. Med. 215, 283-301.
Myers, R., Sanchez-Garcia, J., Leving, D., Melvin, R. and Fernandez-Funez, P. (2022) New Drosophila models to uncover the intrinsic and extrinsic factors mediating the toxicity of the human prion protein. Dis. Model. Mech.
Soto, C. and Pritzkow, S. (2018) Protein misfolding, aggregation, and conformational strains in neurodegenerative diseases. Nat. Neurosci.21, 1332–1340.
The annual BSDB meeting is always on the ‘must-attend’ list for the team at Development. This year was, of course, extra-special because it was our first in-person meeting for two years. For me, it was particularly exciting as it was meant to be my first in-person meeting in my new role as Community Manager of the Node. I was looking forward to catching up with old friends and introducing new researchers to the Node community. Unfortunately, the in-person part was not meant to be, as COVID caught up with me the day before the meeting started. However, in light of the ongoing discussion surrounding conference accessibility and sustainability, it was an excellent opportunity to check out the virtual experience of the joint meeting with the BSCB.
Of course, the fact I got to ‘attend’ the meeting at all was my first big thumbs up for the hybrid concept. I found that it was possible to feel the excitement of the attendees even without being there (and I was only slightly jealous!). Almost every speaker started their talks with ‘I’m really happy to be here presenting at my first in-person conference for two years,’ and the delight in their voices prevented any feeling of the phrase becoming repetitive. The meeting kicked off with a plenary lecture from John Wallingford, who wisely got the whole audience onside by telling us that we are all developmental biologists before going on to explain that he would be talking entirely about cell biology featuring the ignoreome, which is composed of completely uncharacterised genes.
The next talk was the BSDB Cheryll Tickle medal lecture from Emma Rawlins. Unfortunately, Emma was also stuck at home because of COVID, but we went smoothly over to her live-streamed presentation. Emma gave us a whistle-stop tour of her career before focusing on the latest work from her lab on human lung development. The first evening concluded online with the BSCB Raff medal lecture from Florence Young, who presented her PhD (and ongoing) work on microtubule-based cargo transport in neurons. Although I was disappointed to not be able to follow up these excellent talks with more discussion with colleagues, the main thing that I missed was the laser pointer, which was an ongoing problem throughout the meeting. A few of the speakers did use the computer mouse, but for the talks I attended, only Dolf Weijers had a laser pointer set up to show for the online and in-person audience. Fortunately, however, this issue should be an easy fix for future meetings.
Day two of the joint meeting highlighted another couple of advantages of virtual attendance. Firstly, it was possible to jump between the two parallel sessions without disturbing anyone. Another advantage was that I didn’t need to queue for my comfort break or caffeine and cookie hit. However, Mike Fainzilber pointed out on Twitter that great collaborations can be set up in ‘caffeine-queues’:
No comment about comfort breaks, but we have published papers that started from comments in caffeine queues at meeting. Virtual conferences just cannot replace that!
This brings me nicely on to social media: I also followed the meeting on Twitter using the meeting hashtag #BSCBDB22, which I did manage to get wrong a couple of times while tweeting about the meeting – I’m blaming ‘COVID-brain’. I would recommend following conferences on Twitter, especially if you are attending virtually. I think that it helped me feel more connected with the in-person attendees. People were tweeting about the talks, posters and the social side of the meeting. On the other hand, the ‘biggie’ that I was sad to be missing on the second day was the poster session. It was great that the posters were available online and it was possible to type in a question for the author, but it is just not the same. At in-person meetings, posters are where discussion happens, ideas for experiments are formed and new connections are made. Sadly, I don’t think that we have found a way to replicate this online.
Day two concluded with the announcement of the Wolpert and Waddington medal winners awarded by the BSDB. This year’s Wolpert medal for extraordinary contributions to the teaching and communication of developmental biology was awarded to Andreas Prokop. Instead of giving a medal lecture, the prize comes with funding for a small number of lectures around the country. Andreas has been a big supporter of the Node, contributing numerous articles, as well as allowing us to host the resource page that he curated for the BSBD. You can read more about Andreas’ teaching and science communication work here. The Waddington medal for major contributions to any aspect of developmental biology in the UK, was awarded to Val Wilson. In her medal lecture, Val took us on a tour of her favourite embryos, including ‘the embryo that Rosa liked’, describing some of her most important contributions to the field.
Day three, aka the day of the disco, saw me taking advantage of my virtual attendance to jump between the sessions again. The link ups to the speakers unable to attend the meeting in person continued to work seamlessly. The flash talks were outstanding, and their inclusion meant I could hear a little more about the research that I was missing out on by not being able to physically attend the poster sessions. The science part of day three concluded with the BSCB Hooke medal lecture from Jeremy Carlton. Jeremy presented selected highlights from his research journey, focusing on his work on the many membranes abscission events that are dependent on the ESCRT proteins. Day three concluded with the conference dinner and disco. Always a highlight of these meetings, it was a shame to miss out, but the tweets and videos showed everyone having a good time and the dancing was as good (enthusiastic!) as ever.
The organisers had saved the ‘big guns’ for the final morning (possibly to ensure that everyone had vacated their rooms by 9 am as promised to the conference venue), with plenary lectures from Anne Straube and Jody Rosenblatt, and medal lectures from Laura Greaves (BSCB Women in Science medal), Adam Shellard (BSCB Postdoc medal) and Guillermo Serrano Najera (BSDB Beddington medal for an outstanding PhD thesis). I was meant to be conducting my very first in-person interview with Guillermo, which unfortunately couldn’t happen, but we managed to catch up over Teams and I will be posting the interview on the Node soon. Guillermo has so many different interests and this was a really fun interview. Hopefully this comes across in the final article!
Overall, I really enjoyed attending the BSCB/BSDB joint meeting. Do I think the virtual experience is the same as attending in-person? No. Would I have preferred to attend in person? Yes, but I think hybrid meetings should be an ongoing feature of major conferences. There could be any number of reasons a delegate can’t attend in-person, such as sustainability considerations, financial reasons, family commitments, health reasons, etc. and the virtual experience is a great alternative. Being able to follow the meeting via the conference platform and on Twitter still made it a worthwhile and enjoyable experience. Thanks to the organisers from the BSCB and BSDB!
Read on for our news roundup of the past two weeks, with an emphasis on what has caught our eyes on twitter. We have also include a list of meetings with upcoming deadline and a selection of preLights.
It would be remiss not to mention the biggest twitter news this week; Elon Musk has struck a deal to buy the platform. Researchers are speculating what this means for the future of #SciTwitter
Where is science twitter going to end up if we all jump ship one day? Wrong answers only.
Academic writing is one of the top topics on our twitter feed. We enjoyed the thread below as their were so many contributors offering their own best practices. We have collated some of the Node writing advice in a blog post that you can find here.
What are some good #academic writing practices? Please share what has worked for you.
If you are passionate about preprints and would like to gain some additional experience in science communication, then become part of the preLights community. Applications are open until this Friday and you can find out more here.
If you would like to write for the Node, check out our recent list of writing ideas. If you would like to contribute to our ‘Developing news’ blog, please get in touch at thenode@biologists.com
Owen Sansom, the director of the recently launched MRC National Mouse Genetics Network, describes the objectives of the project and his vision for the future of the network.
A new model for translation of mouse research
Over the last 20 years, the UK has become a leading force in the generation and use of complex mouse models in the precise investigation of human disease. Nevertheless, there remains a great challenge to improve how research in animals is translated to clinical benefits.
Key to this will be developing and expanding connections between basic scientists and clinicians to ensure that animal models accurately compare to human disease. This is the focus of the new MRC National Mouse Genetics Network and, I believe, will make a huge impact on our ability to take advantage of recent advances in mouse genetics.
The National Mouse Genetics Network is a major new £22 million investment initially comprised of 7 challenge-led research clusters with members distributed across the UK. At its centre, the Mary Lyon Centre at MRC Harwell will act as a repository for, and provider of, genetically altered mice, as well as generating and sharing access to data, training, and specialist facilities and resources. Importantly, each cluster will integrate expertise in fundamental biology with clinical findings in order to better address pertinent and important research questions.
Having been involved in previous, smaller-scale, network initiatives, I know that this model can produce results, but I believe that it’s a structure that will work better when done on a larger scale, as the scope for collaboration and the capacity of the system grow.
Cross-cluster synergy
Importantly, the Network won’t just connect up a linear series of technicians, scientists, and clinicians with a shared disease interest. The clusters will be focused on distinctive, but complementary, research themes, with plans for cross-cluster and cross-disease interactions, while working on shared goals.
These shared objectives include areas that can broadly benefit animal research, such as the development or improvement of techniques used to thoroughly characterise mouse models of disease, methods used to generate new kinds of mouse models, opportunities for alternatives to animal usage, as well as specific points of common ground between diseases.
Scientists can often become focused on their particular cell-type or tissue of interest, but many diseases can be very wide-ranging in their effects, creating cross-overs between otherwise very different disorders. Through creation of a wider network of experts across many disease areas, we can ensure that our understanding is as deep and broad as possible.
Capacity creation
A large network not only creates more points of contact to allow these kinds of interactions, but also creates greater capacity for a central organising hub. Indeed, one of the core strengths of the National Mouse Genetics Network will be its central hub, the Mary Lyon Centre.
We are in an age of big data, in which single-cell sequencing and deep molecular phenotyping will support the disease positioning of our mouse models so that they are as relevant as possible to human disorders. As well as acting as the repository for the mouse models themselves, the Mary Lyon Centre, along with a data platform initiative within the Network, will build the infrastructure required for open sharing of these data so that scientists within and outside of the network can better work together to improve our understanding of disease.
Our impact will also be accelerated through the Mary Lyon Centre’s training centre, Advance, as it helps to disseminate our new methods both within and outside of the Network.
A platform for access to mouse genetics
My vision is that the Network will create a broad platform to improve access to mouse genetics for those working in translational medicine. This is aimed beyond our starting membership, as I believe that the Network will create a structure that is better able to engage with industry on the development of new disease therapeutics and to initiate new collaborations across academia.
We’re excited to announce the first set of clusters joining the National Mouse Genetics Network and look forward to seeing how they will work together on a number of fantastic projects to develop the next generation of disease modelling in mice and to improve our ability to detect disease at the earliest stages of life. But I’m equally excited to see how others might join or interact with the network, either by working with our existing clusters or by suggesting new research challenges around which to build future clusters.
 (5 votes)
(5 votes)

 (No Ratings Yet)
(No Ratings Yet)