A postdoctoral position is available immediately in Yang Chai‘s laboratory at the Center for Craniofacial Molecular Biology, University of Southern California in Los Angeles, California. We are interested in the regulation of developmental patterning, organogenesis, and mesenchymal stem cells. Our studies will seek to define molecular mechanisms governing both normal and abnormal craniofacial development, providing scientific rationales for future therapeutic strategies to prevent and treat craniofacial birth defects, as well as stem cell based craniofacial tissue regeneration. The candidate must have a PhD and be experienced with molecular and developmental biology. Supported by the NIDCR, NIH. For details, please visit http://chailab.usc.edu/
Send application, resume, and three letters of recommendation to
Wen, Q., Jing, J., Han, X., Feng, J., Yuan, Y., Ma, Y., Chen, S., Ho, T.H., and Chai, Y. (2020) Runx2 regulates mouse tooth root development via activation of Wnt inhibitor Notum. JBMR,PMID 32569388.
Jing, J., Feng, J., Li, J., Han, X., He, J., Ho, T., Du, J., Zhou, X., Urata, M., and Chai, Y. (2019) Antagnistic interaction between Ezh2 and Arid1a coordinate dental root patterning via Cdkn2a. eLife, Jul 1;8. e46426.
Guo, Y., Yuan, Y., Wu, L., Ho, T., Jing, J., Sugii, H., Li, J., Han, X., Guo, C., and Chai, Y. (2018) BMP-IHH-mediated interplay between mesenchymal stem cells and osteoclasts supports calvarial bone homeostasis and repair. Bone Research. 6, 355-367. PMCID: PMC6193039
Li, J., Parada, C., and Chai, Y. (2017) Cellular and Molecular Regulatory Mechanism of Tooth Root Development. Development, 144, 374-384. PMCID: PMC5341797
Brinkley, J.F., Fisher, S., Harris, M., Holmes, G., Hooper, J.E., Wang Jabs, E., Jones, K.L., Kesselman, C., Klein, O.D., Maas, R.L., Marazita, M.L., Selleri, L., Spritz, R.A., van Bakel, H., Visel, A., Williams, T.J., Wysocka, J., the FaceBase Consortium, and Chai, Y. (2016) The FaceBase Consortium: A comprehensive resource for craniofacial researchers. Development, 143, 2677-2688.PMCID: PMC4958338.
Zhao, H., Feng, J., Ho, T. V., Grimes, W. C., Urata, M., and Chai, Y. (2015) The suture provides a niche for mesenchymal stem cells of craniofacial bones. Nature Cell Biology, 17, 386-396. PMCID: PMC4380556
Zhao, H., Feng, J., Seidel, K., Shi, S., Klein, O., Sharpe, P., and Chai, Y. (2014) Secretion of Shh by a neurovascular bundle niche supports mesenchymal stem cell homeostasis in the adult mouse incisor. Cell Stem Cell 14, 160-173. PMCID:PMC3951379
In this episode we’re taking a look at the story and the characters behind one of the most transformative – and ubiquitous – techniques in modern molecular biology: the polymerase chain reaction.
Anyone who has worked with DNA in the laboratory is undoubtedly familiar with the polymerase chain reaction – PCR, as it’s usually known. Invented in 1985, PCR is an indispensable molecular biology tool that can replicate any stretch of DNA, copying it billions of times in a matter of hours, providing enough DNA to use in sequencing or further research, or for applications like forensics, genetic testing, ancient DNA analysis or medical diagnostics.
It’s hard to overstate the transformation that PCR brought to the world of molecular biology and biomedical research. Suddenly, researchers could amplify and study DNA in a way that had been simply impossible before, kickstarting the genetic revolution that’s still going strong today. But where did this revolutionary technology come from? Officially, PCR was invented in 1985 by a colourful character called Kary Mullis, who won a Nobel Prize for the discovery, but, as we’ll see, all the components of PCR were in place by the early 1980s – it just took a creative leap to assemble them into one blockbusting technique.
Image: Illustration depicting semi-conservative DNA replication. Three generations of DNA are shown. After separation of the DNA double helix, two new complementary DNA strands are synthesised (indicated by a new colour). Complementary base pairing and hydrogen bonding results in formation of a new double helix. Credit: Susan Lockhart. Attribution-NonCommercial 4.0 International (CC BY-NC 4.0)
If you enjoy the show, please do rate and review on Apple podcasts and help to spread the word on social media. And you can always send feedback and suggestions for future episodes and guests to podcast@geneticsunzipped.com Follow us on Twitter – @geneticsunzip
A postdoctoral research position is available to study the cellular, genetic, and epigenetic mechanisms of maternal age effects on offspring health and lifespan. The project will focus on the role of mitochondrial dynamics and function in maternal age effects, using molecular, bioinformatic, biochemical, and imaging techniques.
This is an NIH-funded project in the laboratory of Dr. Kristin Gribble at the Marine Biological Laboratory, Woods Hole, MA. The lab researches the mechanisms and evolution of aging and maternal and transgenerational effects on offspring health. We use rotifers as a model system for our work. For more information about the lab’s research and publications, see mbl.edu/jbpc/gribble.
Applicants should possess a Ph.D. molecular biology, cell biology, biochemistry, genetics, bioinformatics, or a related field. The ideal candidate will have a record of scientific rigor, productivity, and creativity. Excellent oral and written communication skills are required. Knowledge of rotifer biology is not required; highly motivated individuals with experience in other model systems and with a background in bioinformatics, cell biology, biochemistry, epigenetics, and/or imaging are encouraged to apply. Salary commensurate with experience and qualifications.
Applicants must apply for this position via the Marine Biological Laboratory careers website. Please submit (1) a cover letter with a brief description of your research experience and how you will contribute to research on the mechanisms of maternal effects on offspring, (2) a CV, and (3) contact information for at least three references.
Development invites you to submit your latest research to our upcoming special issue: Imaging development, stem cells and regeneration.
Imaging-based approaches have long played a role in the field of developmental biology. However, recent technical advances now provide us with the ability to visualise cell and developmental processes at extraordinary resolution and in real-time. From progress in light sheet and super-resolution microscopy, to the development of tissue-clearing techniques and sophisticated image analysis platforms, we are now able to capture and quantitatively analyse the beauty and dynamics of development across different scales – from individual molecules and cells, to complete tissues and embryos. This Special Issue aims to showcase articles that, at their core, have applied such advanced techniques in innovative ways to further our understanding of developmental and regenerative processes. We also encourage the submission of articles that report the development or application of a novel imaging-based technique.
Prospective authors are welcome to send pre-submission enquiries to dev.specialissue@biologists.com. We also invite proposals for Review articles: if you are interested in contributing a Review, please send a summary of your proposed article to us by 15 December 2020.
The Special Issue will be published in mid-2021 (although note that, in our new continuous publication model, we will aim to publish your article as soon as it is accepted*). The issue will be widely promoted online and at key global conferences, guaranteeing maximum exposure for your work.
For information about article types and manuscript preparation, please refer to our author guidelines. To submit your article, visit our online submission system; please highlight in your cover letter that the submission is to be considered for this Special Issue.
The deadline for submitting articles is 30 March 2021.
Why choose Development?
Submissions handled by expert Academic Editors
Competitive decision speeds and rapid publication
Format-free submission
Strong commitment at first decision – over 95% of invited revisions accepted
Free to publish – no page or colour charges, no hidden fees
Easy one-click transfer option to Biology Open
Not-for-profit publisher
* Please note that not all articles accepted for publication will be included in the Special Issue; they may instead be published in earlier or later issues of the journal based on timing and editorial discretion.
Oleic acid triggers hippocampal neurogenesis by binding to TLX/NR2E1
Prasanna Kandel, Fatih Semerci, Aleksandar Bajic, Dodge Baluya, LiHua Ma, Kevin Chen, Austin Cao, Tipwarin Phongmekhin, Nick Matinyan, William Choi, Alba Jiménez-Panizo, Srinivas Chamakuri, Idris O. Raji, Lyra Chang, Pablo Fuentes-Prior, Kevin R. MacKenzie, Caroline L. Benn, Eva Estébanez-Perpiñá, Koen Venken, David D. Moore, Damian W. Young, Mirjana Maletic-Savatic
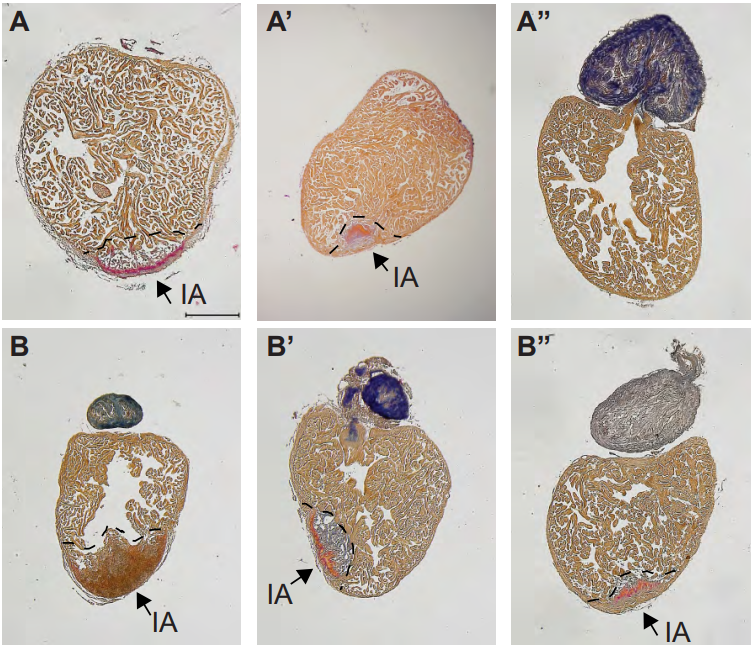
Cardiac Sex Differences are Established Prior to Gonad Formation
Wei Shi, Xinlei Sheng, Kerry M. Dorr, Josiah E. Hutton, Haley A. Davies, Tia D. Andrade, Todd M. Greco, Yutaka Hashimoto, Joel D. Federspiel, Zachary L. Robbe, Xuqi Chen, Arthur P. Arnold, Ileana M. Cristea, Frank L. Conlon
Olig3 acts as a master regulator of cerebellar development
Elijah D. Lowenstein, Aleksandra Rusanova, Jonas Stelzer, Marc Hernaiz-Llorens, Adrian E. Schroer, Ekaterina Epifanova, Francesca Bladt, Eser Göksu Isik, Shiqi Jia, Victor Tarabykin, Luis R. Hernandez-Miranda
Human Naïve Epiblast Cells Possess Unrestricted Lineage Potential
Ge Guo, Giuliano Giuseppe Stirparo, Stanley Strawbridge, Daniel Spindlow, Jian Yang, James Clarke, Anish Dattani, Ayaka Yanagida, Meng Amy Li, Sam Myers, Buse Nurten Özel, Jennifer Nichols, Austin Smith
A Library of Induced Pluripotent Stem Cells from Clinically Well-Characterized, Diverse Healthy Human Individuals
Christoph Schaniel, Priyanka Dhanan, Bin Hu, Yuguang Xiong, Teeya Raghunandan, David M. Gonzalez, Sunita L. D’Souza, Arjun Yadaw, Jens Hansen, Gomathi Jayaraman, Bino Mathew, Moara Machado, Seth Berger, Joseph Tripodi, Vesna Najfeld, Jalaj Garg, Marc Miller, Colleen Lynch, Katherine Michelis, Neelima Tangirala, Himali Weerahandi, David C. Thomas, Robert Sebra, Milind Mahajan, Eric Schadt, Dusica Vidovic, Stephan C Schürer, Joseph Goldfarb, Evren U. Azeloglu, Marc R. Birtwistle, Eric A. Sobie, Jason C. Kovacic, Nicole C. Dubois, Ravi Iyengar
Cathepsin K maintains the number of lymphocytes in vivo
Renate Hausinger, Marianne Hackl, Ana Jardon-Alvarez, Miriam Kehr, Sandra Romero Marquez, Franziska Hettler, Christian Kehr, Sandra Grziwok, Christina Schreck, Christian Peschel, Rouzanna Istvanffy, Robert A.J. Oostendorp
Early Stem Cell Aging in the Mature Brain
Albina Ibrayeva, Maxwell Bay, Elbert Pu, David Jörg, Lei Peng, Heechul Jun, Naibo Zhang, Daniel Aaron, Congrui Lin, Galen Resler, Axel Hidalgo, Mi-Hyeon Jang, Benjamin D. Simons, Michael A. Bonaguidi
Cell size is a determinant of stem cell potential during aging
Jette Lengefeld, Chia-Wei Cheng, Pema Maretich, Melanie R. McReynolds, Marguerite Blair, Hannah Hagen, Emily J Sullivan, Kyra Majors, Christina Roberts, Joon Ho Kang, Joachim Steiner, Teemu P Miettinen, Scott Manalis, Adam Antebi, Jacqueline Lees, Laurie Boyer, Omer H. Yilmaz, Angelika Amon
Endogenous Galectin-3 is Required for Skeletal Muscle Repair
Daniel Giuliano Cerri, Lilian Cataldi Rodrigues, Vani Maria Alves, Juliano Machado, Víctor Alexandre Félix Bastos, Isis do Carmo Kettelhut, Luciane Carla Alberici, Sean R. Stowell, Maria Cristina R. Costa, Richard D. Cummings, Marcelo Dias-Baruffi
A CRISPR-Cas9–engineered mouse model for GPI-anchor deficiency mirrors human phenotypes and exhibits hippocampal synaptic dysfunctions
Miguel Rodríguez de los Santos, Marion Rivalan, Friederike S. David, Alexander Stumpf, Julika Pitsch, Despina Tsortouktzidis, Laura Moreno Velasquez, Anne Voigt, Karl Schilling, Daniele Mattei, Melissa Long, Guido Vogt, Alexej Knaus, Björn Fischer-Zirnsak, Lars Wittler, Bernd Timmermann, Peter N. Robinson, Denise Horn, Stefan Mundlos, Uwe Kornak, Albert J. Becker, Dietmar Schmitz, York Winter, Peter M. Krawitz
Patient iPSC-astrocytes show transcriptional and functional dysregulation in schizophrenia
Marja Koskuvi, Šárka Lehtonen, Kalevi Trontti, Meike Keuters, Ying Chieh Wu, Hennariikka Koivisto, Anastasia Ludwig, Lidiia Plotnikova, Pekka L. J. Virtanen, Noora Räsänen, Satu Kaipainen, Ida Hyötyläinen, Hiramani Dhungana, Raisa Giniatullina, Ilkka Ojansuu, Olli Vaurio, Tyrone D. Cannon, Jouko Lönnqvist, Sebastian Therman, Jaana Suvisaari, Jaakko Kaprio, Markku Lähteenvuo, Jussi Tohka, Rashid Giniatullin, Claudio Rivera, Iiris Hovatta, Heikki Tanila, Jari Tiihonen, Jari Koistinaho
A comprehensive overview of computational tools for RNA-seq analysis
Dhrithi Deshpande, Karishma Chhugani, Yutong Chang, Aaron Karlsberg, Caitlin Loeffler, Jinyang Zhang, Agata Muszynska, Jeremy Rotman, Laura Tao, Lana S. Martin, Brunilda Balliu, Elizabeth Tseng, Eleazar Eskin, Fangqing Zhao, Pejman Mohammadi, Pawel P Labaj, Serghei Mangul
Comparing quality of reporting between preprints and peer-reviewed articles in the biomedical literature
Clarissa F. D. Carneiro, Victor G. S. Queiroz, Thiago C. Moulin, Carlos A. M. Carvalho, Clarissa B. Haas, Danielle Rayêe, David E. Henshall, Evandro A. De-Souza, Felippe E. Amorim, Flávia Z. Boos, Gerson D. Guercio, Igor R. Costa, Karina L. Hajdu, Lieve van Egmond, Martin Modrák, Pedro B. Tan, Richard J. Abdill, Steven J. Burgess, Sylvia F. S. Guerra, Vanessa T. Bortoluzzi, Olavo B. Amaral
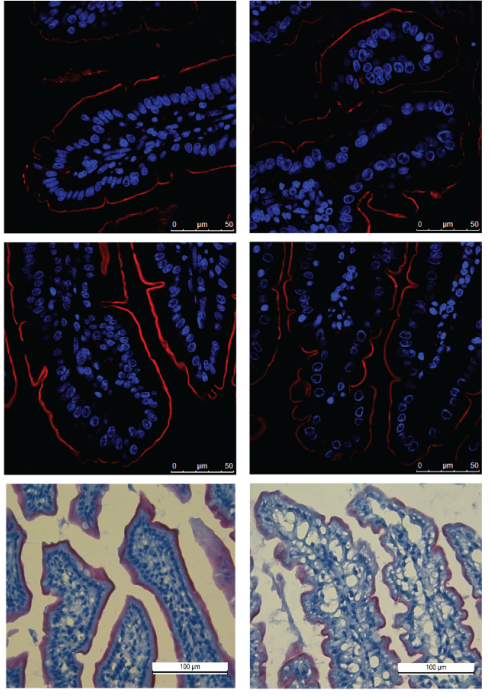
The study of regenerative biology aims to elucidate the innate ability of organisms to repair tissues or organs after they have been removed or damaged. During vertebrate regeneration, tissue damage causes the immediate release of signals that initiate wound closure and initiate regeneration. This project uses larval zebrafish to study how cells respond to when the epithelia is damaged. Zebrafish repair wounds quickly and efficiently, and their small size and transparency allow us to follow cell behaviour easily. Our previous research found that there are very surprising movements of cells over the first few hours after damage, and this current project aims to understand the forces involved in these movements and the signals that orchestrate the wound response. We plan to image fluorescently labelled cells over time to give positional data across the fish using a custom built lightsheet microscope. Data sets will be analysed using physical and computational modelling to calculate passive and active forces such as compression, stretching, shear and friction. Once a physical model of whole animal cell movement is established we will interrogate our predictions by manipulating known early wound signals.
Funding Notes
White Rose BBSRC Doctoral Training Partnership in Mechanistic Biology
4 year fully-funded programme of integrated research and skills training, starting October 2021:
• Research Council Stipend (estimated £15,600 per year)
• Tuition Fees at the UK fee rate (£4,473 per year)
• Research support and training grant (RTSG)
Please note: international tuition fees for 2021 entry are £23,750
To Apply:
Informal inquiries: h.roehl@sheffield.ac.uk
The deadline for applications will be the 10th January 2021 with selection of final candidates for interview shortly after.
Updated 30 October . Let us know if we missed anything
Various organisations and looser assemblies of locked down researchers have begun to put together online seminar and talk series, many of which are open to anyone (usually with registration), and many of which also have previous talks recorded.
Here’s a list of what we’ve found recently, developmental biology and adjacent – please let us know if we missed anything so we can keep it up to date. For upcoming virtual developmental biology conferences/symposia, see our recently updated Events calendar page.
Big news from us is Development presents…, the webinar series hosted each month by a different Development Editor which will be a platform for early career researchers to share their work. As well as the talks, you also have the chance to meet the speakers and other participants at interactive video tables – giving the developmental biology community the chance to network virtually.
Talking about promoting early career researchers, the Society for Developmental Biology in the USA is running a monthly postdoc seminar series. Next one is November 13, speakers TBD (you can see recordings of the October session with Michael Piacentino and Natasha O’Brown on the homepage).
The role of the next generation sequencing (NGS) specialist is to ensure daily operations at the Genomics Platform and to support researchers at two centres working on fundamental biological mechanism, the Novo Nordisk Centres for Protein Research (CPR) and Stem Cell Biology (DanStem), with expertise in functional genomics and single-cell sequencing applications. The advertised job is an exciting opportunity for strong candidates with a background in genomics technology to establish themselves in the field of genomics services. The genomics platform represents a joint venture that bridges the vibrant scientific environment that spans DanStem and the CPR and is central to scientific activities in both centers.
Background
DanStem addresses fundamental research questions in stem cell and developmental biology and has activities focused on the translation of promising basic research results into new therapeutic strategies for cancer and chronic diseases. DanStem is a vibrant, internationally diverse and ambitious research center with state-of-the-art facilities. The setting is ideally suited for seamless collaboration and exchange with other centers and departments of the Faculty and Copenhagen science community.
CPR promotes basic and applied research on human proteins of medical relevance. CPR spans broad areas of protein research, ranging from structural and mechanistic understanding of proteins and their cellular functions in chromatin biology and genome maintenance, to development of clinical applications of proteomics and systems biology. The vision of CPR is to combine the power of integrative protein technologies and their application to accelerate understanding of the biological processes underlying health and disease.
The genomics platform was established in 2017 by DanStem and CPR and is run by a sequencing specialist and a bioinformatics specialist. We are seeking a highly motivated and ambitious candidate to join the platform as sequencing specialist with a focus on user support, method development and training. The genomics platform has access to its own NextSeq500, an Agilent Bravo, Fragment analysers and other necessary genomics equipment. Groups at CPR and DanStem are seeing a steady increase in genomics based experiments and assays that massively benefit from input and advice by the combined expertise of the genomics platform. The advertised job is an exciting opportunity for strong candidates to establish themselves in the field of genomics services in an exciting scientific environment provided by our well-established centers.
Job description
The role of the next generation sequencing (NGS) specialist is to ensure daily operations of the wet lab part of Genomics Platform. The tasks include: Supporting experimental design and advising potential users, training and assistance on library preparations (DNA and RNA based), running and overseeing the NextSeq500 sequencer, training the users on platform instruments (e.g. Fragment Analyser, liquid handling robot), providing advice on genomics tools and troubleshooting. It also includes ordering, stocking and supplying consumables specific to the genomics platform. Importantly, the applicant will be expected to introduce and support the establishment of new genomics applications as a service in collaboration with users.
Your profile
A degree in biotechnology, molecular biology, or appropriate biological or biomedical science with either PhD or MSc.
Theoretical and practical knowledge of NGS sample preparation workflows and applications.
Previous experience with NGS bioinformatics is a plus.
Team oriented communication style and ability to manage experiment associated cost transparently and efficiently
Good English communication skills, both oral and written, are required.
Ability to connect and actively participate in exchange with other genomics platforms nationally and internationally
We offer
Stimulating, challenging and multifaceted research environment
A combination of active scientific service platforms to interact and to develop ideas together and expand available methodology at CPR and DanStem
Possibility for continued education and training
Attractive employment conditions
Central located workplace.
Employment conditions
The employment is a permanent position and is scheduled to start 1 March 2021 or upon agreement with the chosen candidate. The place of work is at DanStem, University of Copenhagen, Blegdamsvej 3B, Copenhagen. Salary, pension and terms of employment are in accordance with the provisions of the collective agreement between the Danish Government and AC (the Danish Confederation of Professional Associations).
In addition to the basic salary, a monthly contribution to a pension fund is added (17.1% of the salary) and depending on qualifications, a supplement may be negotiated.
The employment will be as Research consultant (Specialkonsulent) depending on the selected candidate’s experiences and qualifications and the working time are 37 hours per week on average.
Application
Your application must be submitted in English by clicking “Apply now” below and must include:
Motivation letter
Curriculum vitae incl. education, experience, previous employments, language skills and other relevant skills
Copy of diplomas/degree certificate(s).
Only applications received in time and consisting of the above listed documents will be considered. Applications and/or any material received after the deadline will not be taken into consideration.
Deadline for applications: 9 December 2020.
Questions
For further information about the position, please contact Professor Joshua Brickman by e-mail: joshua.brickman@sund.ku.dk.
The Faculty of Health and Medical Sciences comprises approximately 7,850 students, 1,700 PhD students and 4,800 employees. The Faculty advances the field of health sciences through its core activities: research, teaching, knowledge sharing and communication. With basic research fields ranging from molecular studies to studies of society, the Faculty contributes to a healthy future through its graduates, research findings and inventions benefitting patients and the community. The University of Copenhagen wishes to reflect the surrounding community and invites all regardless of personal background to apply for the position.
The new Center for Stem Cell & Organoid Medicine (CuSTOM) at Cincinnati Children’s Hospital Medical Center (CCHMC) is launching a major new initiative to recruit outstanding tenure-track or tenured faculty at the Assistant to Associate Professor level.
CuSTOM (www.cincinnatichildrens.org/custom ) is a multi-disciplinary center of excellence integrating developmental and stem cell biologists, clinicians, bioengineers and entrepreneurs with the common goal of accelerating discovery and facilitating bench-to-bedside translation of organoid technology and regenerative medicine. Faculty in CuSTOM benefit from the unique environment and resources to studies of human development, disease and regenerative medicine using pluripotent stem cell and organoid platforms.
CCHMC is a leader in organoid biology and one of the top ranked pediatric research centers in the world, providing a unique environment for basic and translational research. Among pediatric institutions CCHMC is the third-highest ranking recipient of research grants from the National Institutes of Health. CCHMC continues to make major investments in research supporting discovery with 1.4 million square feet of research space and subsidized state-of-the-art core facilities including a human pluripotent stem cell facility, CRISPR genome editing, high-throughput DNA analysis, biomedical informatics, a Nikon Center of Excellence imaging core and much more.
We invite applications from innovative and collaborative investigators focused on basic or translational research in human development and/or disease using stem cells or organoid models. Successful candidates must hold the PhD, MD, or MD/PhD degrees, and will have a vibrant research program with an outstanding publication record.
Applicants should submit their curriculum vitae, two to three page research statement focused on future plans, and contact information for three people who will provide letters of recommendation to CuSTOM@cchmc.org. Applications must be submitted by December 1st, 2020
The Cincinnati Children’s Hospital Medical Center, and the University of Cincinnati are Affirmative Action/Equal Opportunity Employers, fostering diversity and inclusion. Qualified women and minority candidates are especially encouraged to apply.
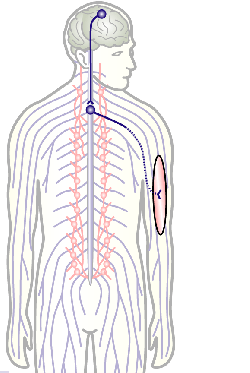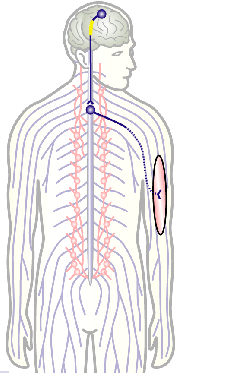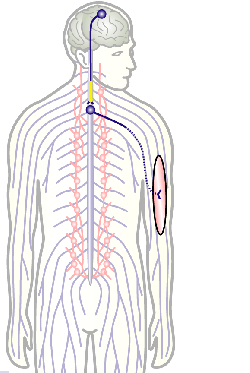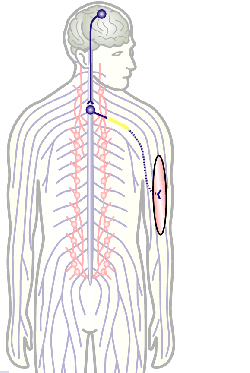
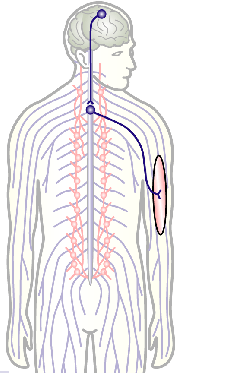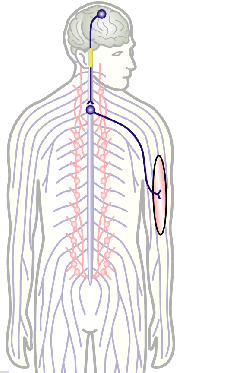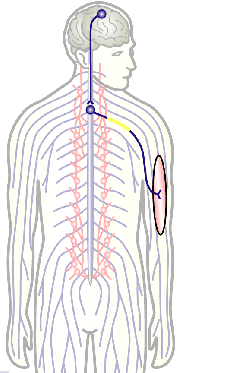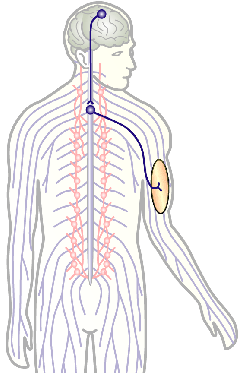
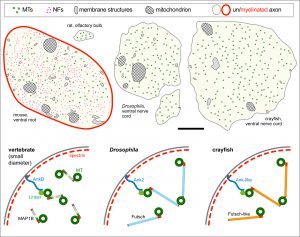
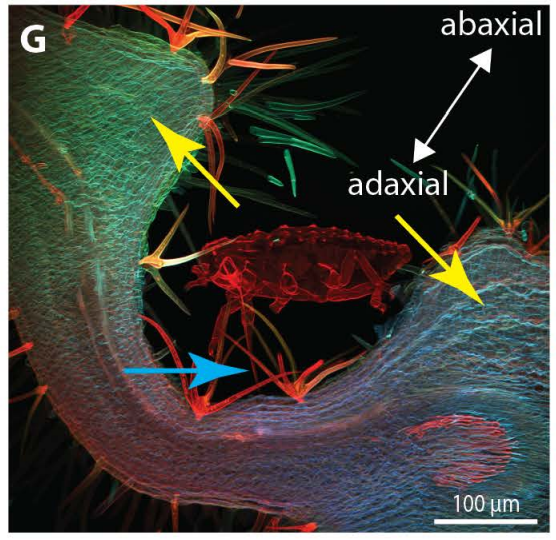
To develop remedial strategies for neurodegeneration in age and disease, we need to improve our understanding of the cell biology of neurons – in particular their axons. Axons are the cable-like, up-to-meter long processes of neurons that wire our nervous system (Fig.1); we lose 40% of axons towards high age and they are key target sites for degenerative processes (Fig.2).
Fig.3 Click to see original
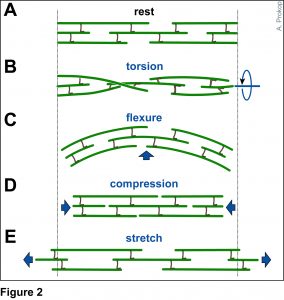
The overarching aim of this project is therefore to gain a better understanding of the architecture of axons and how it evolved from invertebrates to mammals. For this, we focus on the bundles of microtubules (MTs; Fig.3) that extend through the entire axon; they provide the structural backbones of axons and highways for life-sustaining cargo transport and organelle dynamics. Therefore, our studies of the mechanisms that uphold these MT bundles [Refs.1,2] aim to advance our understanding of axon architecture and how they are maintained long-term (Fig.4).
Fig.4 Click to see original
Here we focus on the role of so-called MT cross-linker proteins, expected to hold MTs in parallel bundled arrangements (Figs.3,5). Cross-linkage was one of the first mechanisms proposed to explain axonal MT bundle conformation, and proposed cross-linkers such as Tau or MAP1B have close links to neurodegeneration including Alzheimer’s disease. However, the experimental evidence and molecular understanding of MT cross-linkage is surprisingly sparse [reviewed in Refs. 2,4], leaving an important gap in our understanding of axon architecture and maintenance.
Fig.5 Click for further information
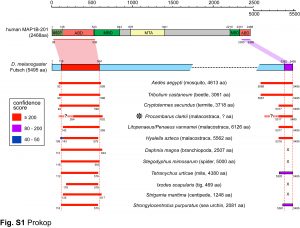
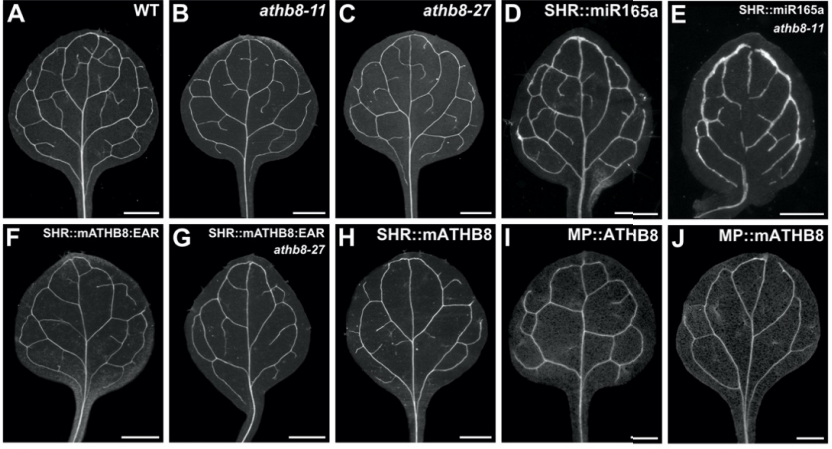
We are particularly interested in MAP1B which plays conserved roles in MT bundle architecture from invertebrates to humans, whilst showing a very particular evolutionary profile (Fig.6): the N- and C-terminal domains have proposed actin-binding properties and are well-conserved throughout the animal kingdom, whereas the middle region is highly variable: it is extremely long (>4,500 aa) in many arthropods correlating with wider spacing of their MTs, whereas other animals including mammals have short forms (1,687 aa in humans) correlating with narrower spacing and the presence of neurofilaments (absent in arthropods). Furthermore, the middle region undergoes rapid change, with sequences not conserved even between closely related species (e.g. Drosophila melanogaster and D. grimshawi flies from the same family). Our detailed studies of MAP1B and its homologues will be doubly beneficial: they will bring important new understanding of mechanisms underpinning evolution, as well as of axon architecture providing information that is also relevant for work on axon degeneration.
During this project, we will capitalise on Matthew Ronshaugen‘s expertise in evolutionary biology to perform phylogenetic analyses of MAP1B and its homologues, aiming to extract concepts and rules that explain MAP1B’s evolutionary behaviour, and develop experimentally testable working hypotheses. Experiments will build on Andreas Prokop‘s expertise on axon structure and MT regulation using Drosophila neurons as a genetically amenable system for fast and efficient experimentation; thus, we will modify the fly MAP1B gene to test our phylogenetics-derived hypotheses. Many experiments will involve electron microscopy for which Karl Kadler is a long-standing expert; EM will reveal structural aberrations, subcellular positions of MAP1B and changes in MT spacing.
Detailed project description
Obj. 1: To understand the evolution of MAP1B homologues: We will perform in-depth evolutionary analysis of the MAP1B protein family by (a) collecting full length MAP1B protein sequences from a broad set of metazoan animal species, (b) building an accurate protein alignment and (c) establishing a stable phylogeny for MAP1B. We will examine how selection has acted on MAP1B domains, for example whether rapid middle region variations are due to neutral drift or possibly positive selection acting to diversify MAP1B crosslinking function and neuronal morphology. Finally, we will use a bioinformatic approach to identify conserved motifs, predict functions based on structure and homology, and examine their gain and loss throughout the MAP1B evolutionary history. Our results will inform the genetic structure function analysis described in Obj. 3 to examine how sequence conservation and divergence within MAP1B proteins correlate with changes in neuronal morphology.
Fig.6 Click image to see original
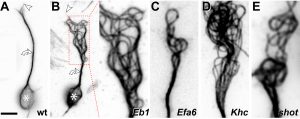
Obj. 2: To establish the mechanisms of Drosophila MAP1B in MT cross-linkage: We will test whether the fly MAP1B is positioned in between MTs and whether it imposes a constant MT-MT spacing in this position. For this, we will perform EM studies using MT contrast enhancement (lanthanum or tannic acid) in combination with MAP1B detectable via a central enzymatic APEX2 tag (generated via CRISPR/Cas9). We know that the conserved N- and C-terminal domains of fly MAP1B are functionally relevant but have no knowledge of the underlying mechanisms [Ref. 3]. We will establish whether they directly bind MTs using biochemical pull-down strategies.
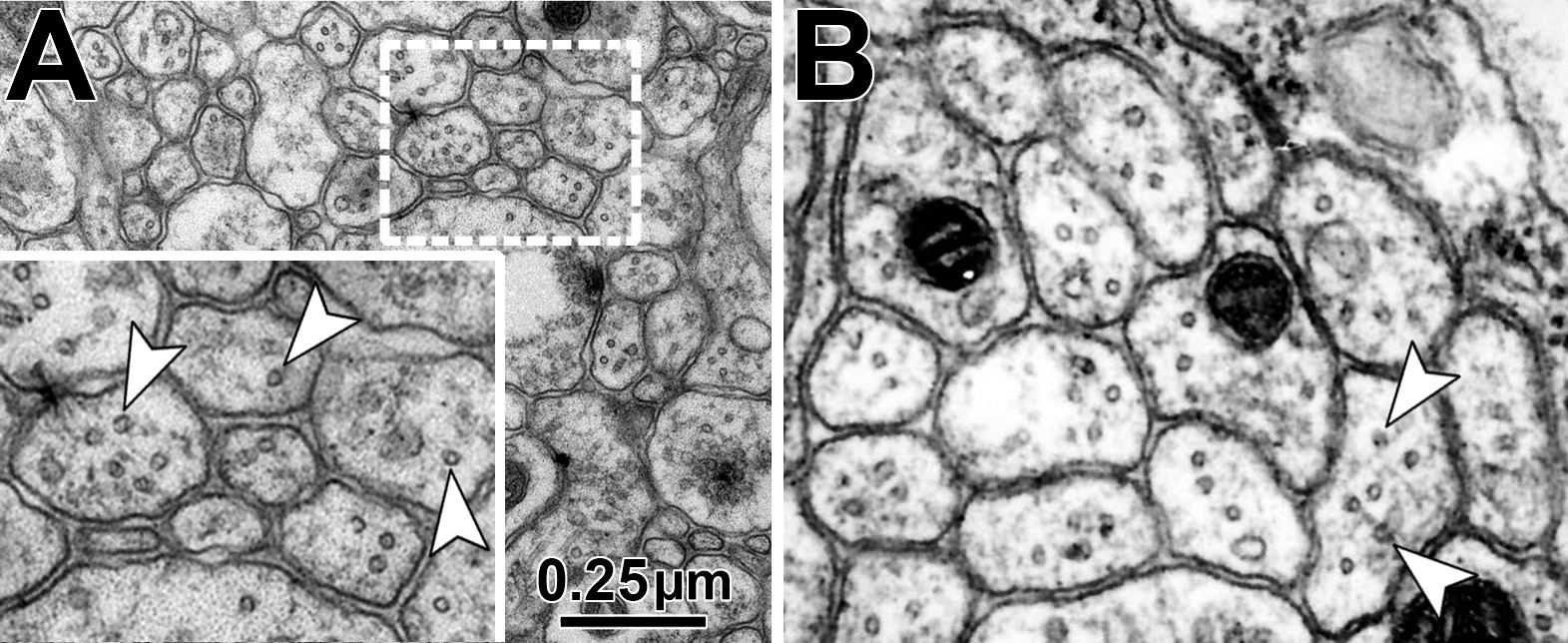
Fig.7 Examples of MT profiles (arrow heads) in axonal cross-sections in the Drosophila CNS (A) and peripheral nerve (B).
Obj. 3: To functionally assess the meaning of evolutionary changes: Using readouts established during Obj. 2, we will introduce changes to fly MAP1B and assess their impacts on axon structure. We already hold constructs of rat MAP1B and mini constructs of fly MAP1B (containing only N- and C-terminal domains) which will be expressed and tested in MAP1B-deficient background to assess changes, for example in MT spacing. We will build on our phylogenetic analyses and use gene blocks strategies to generate and then analyse interspecies hybrid versions of MAP1B; for example we will assess whether the exchange of the middle region has a structural impact.
Taken together, we will establish how MAP1B proteins contribute to the bundled conformation of axonal MTs across the animal kingdom, and why part of this molecule has undergone such a severe change. This will provide new insights into evolutionary mechanisms and relevant understanding of neurodegenerative processes. This project is therefore highly interdisciplinary and provides training opportunities including phylogenetic in silico analyses, molecular biology, classical genetics, biochemistry and electron microscopy.
References
Hahn, I., Voelzmann, A., Parkin, J., Fuelle, J. B., Slater, P. G., Lowery, L. A., Sanchez-Soriano, N., Prokop, A. (2020). Tau, XMAP215/Msps and Eb1 jointly regulate microtubule polymerisation and bundle formation in axons. bioRxiv, 2020.08.19.257808 — [LINK]
Hahn, I., Voelzmann, A., Liew, Y.-T., Costa-Gomes, B., Prokop, A. (2019). The model of local axon homeostasis – explaining the role and regulation of microtubule bundles in axon maintenance and pathology Neural Dev 14, 10.1186/s13064-019-0134-0 — [LINK]
Hummel, T., Krukkert, K., Roos, J., Davis, G., Klämbt, C. (2000). Drosophila Futsch/22C10 is a MAP1B-like protein required for dendritic and axonal development. Neuron 26, 357-370 — [LINK]
Prokop, A. (2020). Cytoskeletal organization of axons in vertebrates and invertebrates. J Cell Biol 219, e201912081 — [LINK]
Qu, Y.*, Hahn, I.*, Lees, M., Parkin, J., Voelzmann, A., Dorey, K., Rathbone, A., Friel, C., Allan, V., Okenve Ramos, P., Sánchez-Soriano, N., Prokop, A. (2019). Efa6 regulates axon growth, branching and maintenance by eliminating off-track microtubules at the cortex. eLife 8, e50319 — [LINK]
 (No Ratings Yet)
(No Ratings Yet) In this episode we’re taking a look at the story and the characters behind one of the most transformative – and ubiquitous – techniques in modern molecular biology: the polymerase chain reaction.
In this episode we’re taking a look at the story and the characters behind one of the most transformative – and ubiquitous – techniques in modern molecular biology: the polymerase chain reaction.
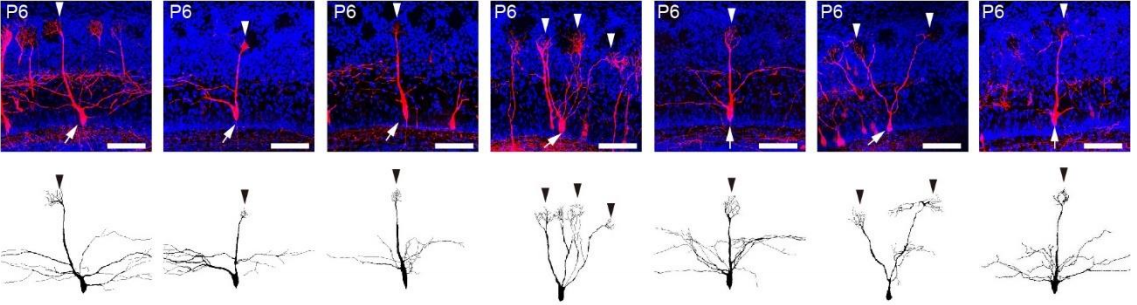
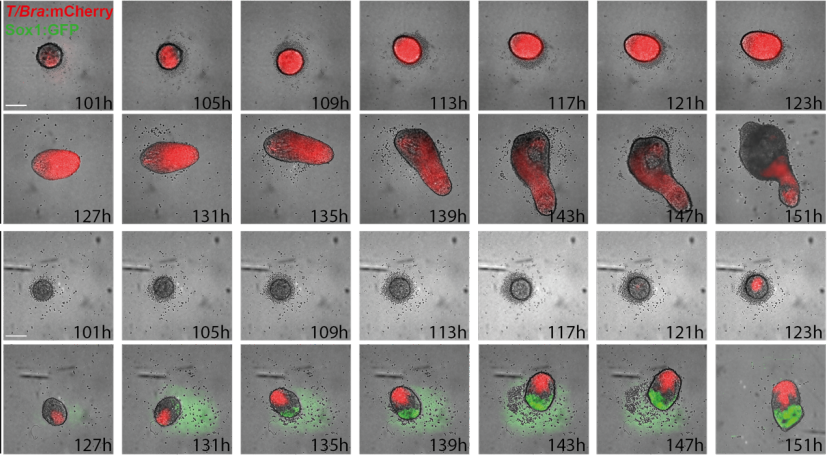
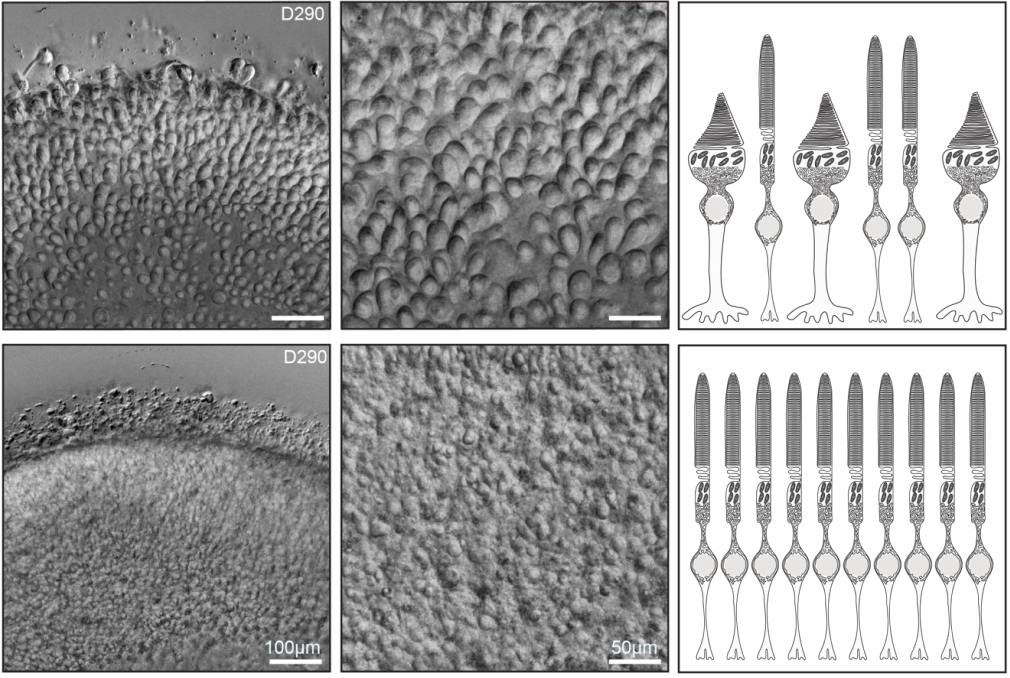
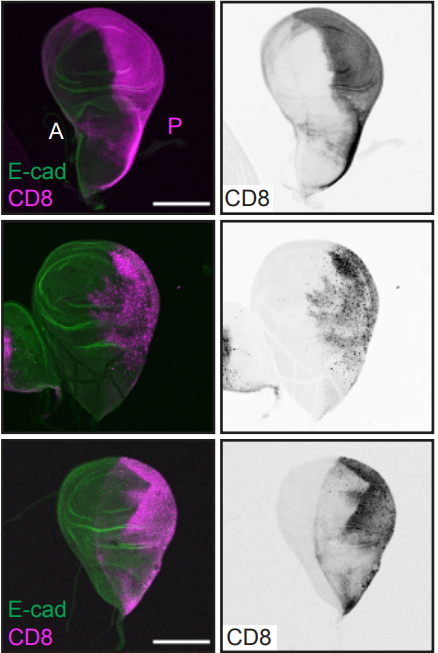










 (12 votes)
(12 votes)