There is a vacancy for a 4-year PhD student position at the Sars International Centre for Marine Molecular Biology in the research group of Dr. Pawel Burkhardt (www.sars.no). The position is funded by Sars Centre core funding. The Sars Centre belongs to the University of Bergen and is partner of the European Molecular Biology Laboratory (EMBL) (www.embl.de). The place of work will be at the Sars Centre. The starting date is flexible, but should be no later than 01 October 2019.
About the project/work tasks:
The goal of the Burkhardt group is to reconstruct the evolutionary origin of synapses and neurons. The group is particularly interested in studying synaptic protein homologs in choanoflagellates, sponges and ctenophores. We are looking for a highly self-motivated and enthusiastic PhD student with interests in evolutionary biology, neurobiology or cell biology. The successful candidate will undertake research with the possibility to use a variety of techniques, ranging from various biochemical methods to X-ray crystallography, super resolution immunofluorescence and electron microscopy, to study synaptic protein homologs in choanoflagellates and sponges (for examples see: Burkhardt et al, 2011 PNAS; Burkhardt et al, 2014 Mol Bio Evo; Bhattacharyya et al, 2016 eLife; Laundon et al, 2019 PLoS Biology). The PhD project will focus on when protein signalling complexes required for synaptic activity first evolved and how they functioned at a molecular level.
Qualifications and personal qualities:
The applicant must hold a master’s degree or the equivalent or must have submitted his/her master’s thesis for assessment prior to the application deadline. It is a condition of employment that the master’s degree has been awarded.
Routine experience in standard molecular and cellular biology techniques is required
Practical experience in a variety of biochemical techniques (e.g. protein expression & purification, Co-IPs, mass spec) and with different fluorescence imaging techniques is highly desirable
Specific experience with choanoflagellates is beneficial, but not essential.
The ability to work both independently and to cooperate with others and a high motivation/enthusiasm is essential.
Proficiency in both written and oral English
About the PhD position:
The duration of the PhD position is 4 years, of which 25 per cent of the time each year comprises required duties associated with research, teaching and dissemination of results. The employment period may be reduced if you have previously been employed in a recruitment position.
About the research training:
As a PhD Candidate, you must participate in an approved educational programme for a PhD degree within a period of 4 years. A final plan for the implementation of the research training must be approved by the faculty within three months after you have commenced in the position. It is a condition that you satisfy the enrolment requirements for the PhD programme at the University of Bergen.
We can offer:
A professional, challenging and international working environment.
Well-equipped, modern laboratories and facilities
Salary at pay grade 51 (Code 1017/Pay range 20, alternative 9) in the state salary scale. Currently equal to NOK 449.400. Further promotions are made according to qualifications and length of service in the position
Enrolment in the Norwegian Public Service Pension Fund.
A position in an inclusive workplace (IA enterprise).
A brief account of the applicant’s research interests and motivation for applying for the position
The names and contact information for two reference persons. One of these must be the main advisor for the master’s thesis or equivalent thesis
CV
Transcripts and diplomas showing completion of the bachelor’s and master’s degrees, or official confirmation that the master’s thesis has been submitted
Relevant certificates/references
List of published papers and pre-prints or other relevant scientific work.
Please note that the entire application and relevant appendices with certified translations into English or a Scandinavian language must be uploaded at Jobbnorge. Applications submitted without required information or applications sent as e-mails will not be considered. Only submitted documents will be subjected to an expert assessment.
Detailed information about the position can be obtained by contacting: Group Leader Pawel Burkhardt, phone: +47 906 48 539, email: Pawel.Burkhardt@uib.no
A postdoctoral position is available in the Laboratory of Gareth Fraser (www.fraser-lab.net) at the Department of Biology, University of Florida. Our lab is focused on a range of research projects based around the central theme of vertebrate evolutionary developmental biology – they include tooth development and regeneration, craniofacial diversity, EvoDevOmics, and skin appendage evolution and development.
We are looking for a motivated and creative scientist to continue to develop the shark and ray as comparative evodevo models for a range of research projects. The primary focus of this position will be stem cell mediated tooth regeneration in sharks and rays, however we do encourage development of the candidates own creative research ideas and interests, so there is potential here to develop a project that suits the candidate in line with the focal themes of the lab. The salary is competitive and commensurate with qualifications and experience, and includes a full benefits package. The applicant selected for this position would begin the beginning of the Fall 2019 term. The anticipated date would be August 16, 2019.
Qualifications
Candidates should have an interest in evolutionary developmental biology, and vertebrate diversity. Knowledge of developmental biology techniques, RNAseq analyses or stem/regenerative biology is desirable.
Application Instructions
For full consideration, applications must be submitted online at http://apply.interfolio.com/62577 and must include: (1) a letter of application summarizing the applicant’s qualifications and interests, (2) a complete curriculum vitae, (3) a statement on research goals, and (4) ) a list of at least three references. After initial review, applicants who are chosen to receive further consideration will be asked to request confidential letters of recommendation from the references. If you would like further information about the position please contact Gareth Fraser. Email: g.fraser@ufl.edu.
Application deadline is May 29, 2019.
All candidates for employment are subject to a pre-employment screening which includes a review of criminal records, reference checks, and verification of education.
The selected candidate will be required to provide an official transcript to the hiring department upon hire. A transcript will not be considered “official” if a designation of “Issued to Student” is visible. Degrees earned from an educational institution outside of the United States require evaluation by a professional credentialing service provider approved by the National Association of Credential Evaluation Services (NACES), which can be found at http://www.naces.org/.
The University of Florida is an equal opportunity institution dedicated to building a broadly diverse and inclusive faculty and staff. Searches are conducted in accordance with Florida’s Sunshine Law. If an accommodation due to disability is needed in order to apply for this position, please call (352) 392-2477 or the Florida Relay System at (800) 955-8771 (TDD).
The Department particularly welcomes applicants who can contribute to a diverse and inclusive environment through their scholarship, teaching, mentoring, and professional service. The university and greater Gainesville communities enjoy a diversity of cultural events, restaurants, year-round outdoor recreational activities, and social opportunities.
Applications are invited from highly motivated individuals who are interested in fundamental mechanisms of neurodevelopment and neurogenetic disorders. The focus of the project is to understand neural developmental and neurological phenotypes in mouse models ofTuberous Sclerosis Complex.The fellowship is in the laboratories of Sara Wilson at the Department of Integrative Medical Biology (IMB), and Leif Carlsson at Umeå Center for Molecular Medicine (UCMM) both laboratories at Umeå University, Sweden. The facilities provide an interactive modern environmentwith easy access to good core facilities. The fellowship is administratively placedat IMB, which is an interdisciplinary department focusing on questions in basic and medical sciences. The fellowshipis funded for two years and is available immediately. The working ‘day to day’ language in the laboratories is English.
Requirements:Individuals with a background in mouse genetics and handling, developmental biology, neuroscience, molecular and/or cell biology with a keen interest in developmental neuroscienceare encouraged to apply. Experience with rodent motor circuits or rodent motor or seizure behavior analysis is a big advantage.Technical experience with imaging, molecular biology, immunohistochemistry,in situhybridization, vertebrate embryonic model systemsare required. Technical experience with mouse embryo electroporation and /or neurite outgrowth/migration assayswill also be positively evaluated. The successful candidate will have or about to receive a Ph.D.in a relevant discipline, have good communication skills and be proficient in written and spoken English. The most successful candidatewill have ahigh level of motivation, be organized, rigorous and have the ability to work both independently and within a team.
Application:Please submit your application (reference 2019SW7) by 31stMay 2019 to sara.wilson@umu.seby sending the following documents as a single pdf file:
A short cover letter (not more than 1 page) to include a description of your research experience and suitability for the position.
Curriculum Vitaeincluding: publication list, technical expertise, names and contact information for three referees.
Informal enquiries may be directed to Dr. Sara Wilson (sara.wilson@umu.se) or Prof. Leif Carlsson (leif.carlsson@umu.se).
How can a genome’s cis-regulatory potential drive robust gene expression throughout embryonic development? Do topologically associating domains (TADs) truly contribute to gene expression by spatially constraining the regulatory actions of cis-sequences? And might phase separation by multi-valent proteins also be involved in the regulation of genes and genomic regions? The recent Company of Biologists Workshop on “Chromatin-based regulation of development”, organized by Benoit Bruneau (Gladstone Institutes, UCSF) and Joanna Wysocka (Stanford University), was the perfect playground to discuss important questions like these, and to stimulate interactions between early career and established scientists. Why was this Workshop so great to me (currently a postdoc in the lab of François Spitz, Institut Pasteur, France)? And why should you – a young investigator – apply to attend a future one? (more…)
Have you got a beautiful image of a developing organism you’d like to share with the world? Or a striking computer generated visualisation, or even something you’ve drawn or painted or stitched or crafted? We’re planning a Node calendar for the academic year 2019-20, and want to fill it with your images.
Entering is easy: simply send your image, along with a description of what’s being shown, tothenode@biologists.com, with the subject line ‘Calendar competition’.
Content-wise, the only limitation is that it should have a developmental biology angle to it. We’d love to have as much diversity in organisms and systems as possible. The image dimensions on the calendar are 24 cm L x 19 cm H, and some cropping may occur. One entry per person but individual labs can enter multiple images.
The deadline for submissions is 13.00 (GMT+1), Friday 10 May
We’ll collate all the entries and then run a public vote the following week. Good luck!
The idea for the calendar didn’t come from nowhere: Development used to print calendars each year and distribute them to researchers until it became economically unfeasible. But people would often ask us about them at conferences (or rather, complain about not having them any more!), and some of us in the Development team remember how nice the calendars looked hanging up in the lab before we left those labs for publishing. We hope our Node calendar will be equally beautiful and useful!
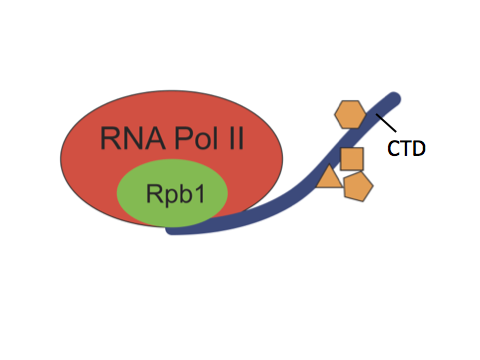
RNA polymerase II (Pol II) is the enzyme responsible for transcribing most genes in eukaryotes. The C-terminal domain (CTD) is a highly repetitive, unstructured domain on the largest Pol II subunit, Rpb1. It consists of numerous repeats of seven amino acids and serves as a binding platform for numerous factors involved in gene regulation, including chromatin modifying enzymes, transcription factors and RNA processing enzymes. Both length and sequence complexity of the CTD greatly vary among species of different lineages. For example, the CTD of S. cerevisiae has 26 repeats, the majority of which are consensus repeats of YSPTSPS. In contrast, mammalian CTDs have 52 repeats, the distal repeats are mostly divergent repeats whose sequence differ from the consensus at one or more positions.
The CTD (blue ribbon) is a repetitive, disordered tail on the Rpb1 subunit of RNA Pol II that interacts with a myriad of essential factors (orange).
The length and sequence complexity of the CTD were thought to regulate gene expression and be essential for the development of complex organisms. In the past decade, this proposition has been reinforced by several studies in mammalian cells. These studies showed that post-translational modifications occur on certain residues of the divergent repeats, some of which are recognized by factors only present in higher eukaryotes, and that mutations which prevent such modifications lead to misexpression of genes. Yet the significance of the divergent residues has never been systematically examined in the context of development.
Considering the above findings in mammalian cells, one would anticipate that a mouse would die if all of its CTD repeats were replaced with the consensus repeats that predominate the yeast CTD. However, generating transgenic mouse lines is very time and cost consuming. Instead, we decided to perform systematic mutagenesis of the CTD in Drosophila, an organism that is also highly genetically malleable but faster and cheaper to mutate compared to mice. In addition, 40 of the 42 CTD repeats in the fly diverge from the consensus, so it is an ideal model to interrogate the significance of divergent repeats.
The project was started at a time when CRISPR/Cas9 was not yet widely adopted by the fly research community, so initially we relied on RNAi knockdown to deplete the endogenous Rpb1 subunit. We also tested if the co-expression of an RNAi-resistant Rpb1 harboring mutations in the CTD would rescue the RNAi phenotype, making approximately 20 transgenic fly lines expressing various mutant forms of Rpb1 in an attempt to identify essential regions of the fly CTD. To our surprise, despite the high conservation in the amino acid sequence of the CTD within the fruit fly genus, most of our mutant CTD flies were viable, even when internal deletions removed up to 30% of the entire CTD. The only essential region we identified was an 8-repeat region encompassing the only two consensus repeats in the fly CTD (Gibbs et al., 2017). This suggests that the majority of the divergent repeats in the fly CTD are redundant.
Our initial systematic mutagenesis of the fly CTD emboldened us to test the idea that all divergent repeats could be replaced with consensus repeats. However, most previous findings in mammalian cells argue that the divergent CTD repeats are essential, so we still expected that flies would die with an all-consensus CTD. The most straightforward way to test this would be to simply replace the fly CTD with 42 consensus repeats. However, since many of the CTD repeats appear redundant, consensus repeats might be able to do what divergent repeats do, but with more or fewer repeats to achieve the same function. Therefore, we tested a series of consensus CTDs ranging from 10 to 52 repeats.
Surprisingly, we obtained normal looking flies with solely consensus repeats. This was achieved with only 20 to 29 repeats, which approximates the length of the yeast CTD. In contrast, flies with 42 consensus repeats, which matches the length of the wild-type fly CTD, barely survived to adulthood. In hindsight, we were fortunate to have tested varying numbers of consensus repeats because had we only tested the 42 consensus repeat CTD, we would have concluded that the divergent motifs are indispensable, which would have driven the project in a different direction. Additionally, flies with 52 consensus repeats died, whereas flies with the human CTD, which is also composed of 52 repeats but contains a mixture of consensus and divergent repeats, were able to survive. Therefore, it seems as though consensus repeats alone are sufficient, yet having too many of them is bad for fly development.
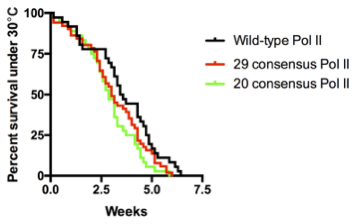
We were shocked by the results of the RNAi rescue experiment but at the same time concerned that residual endogenous Rpb1 might contribute to survival. Therefore, we decided to use CRISPR/Cas9 to mutate the endogenous Rpb1 gene. Remarkably, the CRISPR results agreed with our RNAi experiment. In particular, our flies with 20 to 29 consensus repeats did just as well as flies with the wild-type CTD even under temperature stress.
Death plots of fly lines with wild-type versus consensus Pol II when raised under heat stress (30°C). Pol II with 20 or 29 consensus repeats show comparable to wild-type survival rates.
As flies can survive with solely consensus repeats, consensus and divergent repeats should both be able to interact with the same group of factors. Our data also suggests that interactions provided by the consensus repeats are likely stronger, since in the case of an all-consensus CTD, fewer than half the number of wild-type repeats are needed for such interactions to occur properly. This fits nicely with the phase separation mechanism proposed by Weber and Brangwynne where protein interactions could be mediated by weak, multivalent interactions (Weber and Brangwynne, 2012). In reviews published when our project was being developed, Hnisz et al and Harlen et al proposed that such forces could also drive CTD:factor interactions (Harlen and Churchman, 2017; Hnisz et al., 2017). Also consistent with the phase separation theory was that the deleterious effects of having too many consensus repeats could be reverted either by shortening the CTD (as was the case with our truncated consensus CTDs), or by replacing some of the stronger repeats with weaker ones (as was the case with 52 consensus CTD versus the human CTD), both of which could reduce the overall valency of the CTD.
Coincidentally, our co-author Bede Portz had a chat with Stirling Churchman at the Cold Spring Harbor Mechanisms of Eukaryotic Transcription Meeting about our CTD mutagenesis in flies. Stirling was interested in our findings and was curious to know if the CTD by itself could target transcription sites. Bede thought that the fruit fly salivary glands would be an ideal system to test this. The salivary gland cells undergo rounds of replication without cell division, giving rise to polytene chromosomes where numerous copies of sister chromatids are fused together, which allows for visualization of each transcription site as a distinct band (or ‘puff’). Before us, John Lis’ group had shown that many transcription components such as Pol II are compartmentalized at heat shock puffs in salivary glands (Yao et al., 2006; Yao et al., 2007; Zobeck et al., 2010). Transcription compartments have also been documented recently using live super-resolution microscopy or in other systems where numerous copies of the same DNA sequences were introduced into a genomic location (Cho et al., 2018; Chong et al., 2018). However, the polytene chromosomes provide the power to visually determine a factor’s spatial location with respect to euchromatin, heterochromatin and nucleoplasm without having to rely on super-resolution microscopy or amplifying the number of copies of a candidate gene.
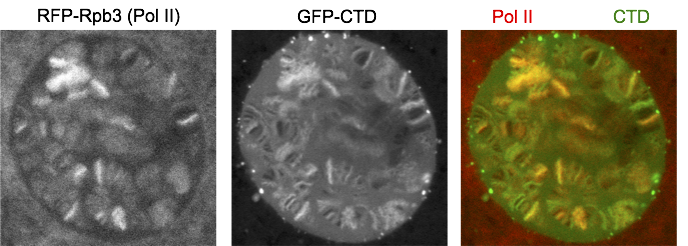
We fused just the CTD to GFP and expressed it in salivary glands. Interestingly, GFP-CTD colocalizes with Pol II on fixed polytene chromosome spreads. One possibility is that the CTD alone dynamically associates with the transcription sites. The alternative is that the GFP-CTD is stably bound by CTD binding partners that are abundant at transcription sites. To distinguish the two possibilities, we turned to live imaging and showed that GFP-CTD on puffs can rapidly recover upon photobleaching, suggesting that the CTD is dynamically recruited to transcription compartments. In addition, phase separation is likely the driving force for the behavior of the CTD, as 1,6-hexanediol, which has been shown to disrupt weak hydrophobic interactions, can disrupt the association of GFP-CTD to chromosomes.
Single Z confocal stack of a live nucleus from prepupal salivary glands with Pol II Rpb3 subunit labeled in red and the CTD alone labeled in green.
The partitioning characteristic of a molecule is influenced by its valency. This led us to investigate how the number of consensus repeats impacts the behavior of the CTD. To address this, we fused our series of consensus CTDs to GFP. Phase separation models predict that changes in valency could lead to pathological aggregates (Weber and Brangwynne, 2012). Indeed, our 42 and 52 repeats of consensus CTDs, which do not support normal function in the fly, showed less GFP on puffs and correspondingly more extrachromosomal foci when fused to GFP. Much of the extrachromosomal foci were neither recovered upon photobleaching nor dispersed by 1,6-hexanediol, suggesting that they were stable aggregates. Interestingly, none of our consensus CTDs partition into transcription compartments as readily as the wild-type CTD. These results suggest that the divergent repeats play a role in either preventing aggregation, or facilitating partitioning into transcription compartments, both of which could be forces that constrain the sequence conservation of the CTD in evolution.
At the start of this project, we had anticipated that the divergent repeats would be essential for fly viability since the sequences of the CTD in 12 species of Drosophila are highly conserved. Moreover, the ratio of synonymous to nonsynonymous mutations among the 12 species is high, thus indicating that the fly CTD is under significant purifying selection. Our discovery that the entire fly CTD can be replaced by consensus repeats poses a conundrum since it calls into question the basis for the purifying selection. One possibility is that intra and intermolecular interactions intrinsic to the CTD constrain the sequence. Our SAXs analysis showed that the CTD adopts a compact random coil structure and the compaction implies transient interactions within the CTD (Gibbs et al., 2017). Also, recent results show that CTD molecules self-associate to form droplets (Boehning et al., 2018). Both types of interactions might need to be finely tuned to prevent aggregation and allow for factor binding to the CTD. Despite the sequence differences, the 29 consensus and wild-type Drosophila CTDs appear to meet these constraints.
The theory of constructive neutral evolution provides another answer to the conundrum. This theory was formulated to explain the gap between the simplicity and complexity of molecular machines that serve the same function in different organisms (Gray et al., 2010). In our case, the 29 consensus CTD represents the simple end of the spectrum while the Drosophila CTD represents the complex end. For both to be functionally equivalent, we posit that the essential functions of the CTD are mediated by a limited number of proteins that interact with the consensus repeats. It is notable that while the Drosophila CTD only contains two repeats that exactly match the consensus, these are embedded in a region encompassing seven other near-consensus motifs. Deletion of this region completely eliminated the capacity of Rpb1 to support fly viability. Constructive neutral evolution posits that chance mutations in the consensus sequences are tolerated by the fortuitous binding of other proteins which compensate for the loss of the consensus sequences. This fortuitous interaction might offset the tendency of the mutation to cause the CTD to aggregate or become mistargeted. Alternatively, the loss of consensus repeat could diminish the affinity of an essential protein for the CTD but this could be offset by the fortuitous binding of another protein that simultaneously associates with the mutant repeat and the protein that normally binds directly to the consensus repeat. Occurrence of the fortuitous binding caused by one mutation sets the stage for mutating additional consensus repeats. The evolutionary trajectory is then dictated by chance and could explain why the consensus CTD, the human CTD, and the Drosophila CTD all support Rpb1 function in the fly. If the consensus CTD truly represents the simplest CTD, then there should exist a class of mutations in genes outside of the Rpb1 gene that are deleterious to wild-type flies but not to those carrying the 29 consensus CTD.
Considering the number of CTD mutants that we had to generate to come to our final conclusion, we feel extremely lucky to have started this project in flies: the fruit fly is indeed an amazing model system to study the CTD. In addition to this work, the divergent nature of the fly CTD has allowed our collaborators Dr. Scott Showalter at PSU, Drs. Yan “Jessie” Zhang and Jennifer Brodbelt to map structural changes and post-translational modifications to each individual repeat without having to introduce additional mutations to the CTD (Gibbs et al., 2017; Mayfield et al., 2016). Furthermore, the fly salivary gland present a unique system to characterize the spatial distribution of transcription factors with just a standard confocal microscope. We envision that the currently available gene-editing and optogenetic tools will allow more exciting discoveries of transcription to be made in the salivary glands.
One of the things I learned during my stay in another lab is that you can’t succeed completely by yourself. You can try, sure, but a relevant unanswered question in biology is a hard nut to crack. It usually takes a multidisciplinary approach and great ideas coming from more than one brain. Collaborations are key to overcome this issue.
Most of the times, what keeps you from collaboratingwith peer scientists is money. Money to travel and money to pay for your accommodation. Renting a room in the US can cost as much as your entire monthly wage in Spain. Time is another big factor. Traveling takes time. As a Ph.D. student though, my time is fully dedicated to research anyway, so that factor is negligible for me.
Collaborate or perish
When Kate McDole and colleagues published their paper (McDole et al., 2018) describing their innovative live imaging microscope, I thought, ‘That is a method I could use for my project’. However, in my research center, I don’t have the facilities nor the expertise to carry it out.
In Miguel Torres Lab, at the Spanish Center for Cardiovascular Research, I am using live imaging to study how the heart establishes its heterogeneous cell types during gastrulation and how mouse embryos manage to form a primitive tube that later on starts beating. However, our methodology does not support imaging live specimens for longer than 20h. Keller’s Lab, at Janelia Reseach Institute, managed to build a fast light-sheet microscope that adapts to the mouse embryo growth and optical properties, allowing its imaging throughout 48h at an unprecedented resolution.
A travel grant to make it happen
The Company of Biologists provided with the funds to cover my expenses, and so I was able to join Keller’s lab for 5 weeks. At Janelia, I found a welcoming and collaborative environment surrounded by cutting-edge facilities. Besides getting some precious datasets from their microscope, I learned how to analyze the vast amount of data generated by time-lapse acquisitions (they run for 48h and take 300 stacks every 4 minutes, do the math…). Learning to program in Python wasn’t in my plans but it ended up being the most valuable thing I learned during the stay. Programming was completely unknown to me before I did the stay, and now I don’t know what I would do without it. To be able to quantify the cell dynamics I am observing in the videos and extract meaningful information becomes way more efficient when one uses programming.
It’s true that a Skype call can help you work together with other scientists, but sometimes you need to be there to learn other techniques, bring up together a project or receive inputs from clear-eyed researchers. That, I think, makes a project more solid and likely to succeed in answering the proposed question. I strongly believe collaborations blossom science, and the role of these traveling fellowships is key to make them happen. At the end of the day, to get to your goal you need a team, and some members of that dream team might be in another part of the world.
If you want to know a bit more about my Ph.D. project, here’s a 4min video explaining it:
The Craft lab at Boston Children’s Hospital and Harvard Medical School is looking for an outstanding, highly motivated postdoctoral fellow to join our developmental biology and pluripotent stem cell-based team.
We established directed differentiation protocols to generate distinct articular and growth plate-like cartilage lineages from human and mouse pluripotent stem cells (Craft et al., Development 2013; Craft et al., Nature Biotech 2015). NIH and foundation funded projects in our lab include investigating mechanisms of articular chondrocyte lineage commitment and stability of this fate through deep sequencing and functional assays, specification and characterization of joint progenitor cells using newly generated fluorescent reporter lines, translational/preclinical experiments of cartilage repair in large animals, and developing iPSC models of congenital cartilage disease. Collaborative projects with Harvard faculty include studies of how modifications in regulatory elements of GDF5 impact gene expression and differentiation of joint lineages (with Dr. Terence Capellini, Harvard University), and evaluating the cartilage and tendon-inducing functions of small molecules and their associated signaling pathways (identified by Dr. Jenna Galloway, Massachusetts General Hospital, through zebrafish screens) in embryonic stem cell (ESC) differentiation cultures.
Requirements: The successful candidate will have received a PhD or MD/PhD within the past 2 years, a minimum of 3 years laboratory experience including tissue culture, at least one first author publication, and excellent communication skills in English. Individuals with experience in one or more of the following are encouraged to apply: limb/joint/cartilage or early embryonic development, signal transduction pathways, ESC/iPSCs, single cell RNAseq/programming.
Apply: Interested applicants should email their CV, a cover letter describing their research background/interests, and contact information for three references to april.craft at childrens.harvard.edu
Press release from Development. You can also read the Research Highlight for this article.
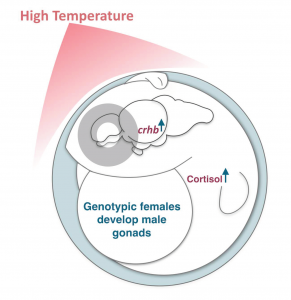
Researchers have identified the hormone that causes sex reversal of medaka fish growing in high temperatures. This study from the Instituto Tecnologico de Chascomus (INTECH) in Argentina is the first to report that the brain is involved in the masculinization of females, which has implications for fish populations as temperatures rise. The research has just been published in the scientific journal, Development.
Dr Juan Fernandino led the team investigating sex reversal, a phenomenon seen in some species whereby environmental conditions, such as high temperatures, cause genetically female fish to develop testes rather than ovaries. Now, researchers show for the first time that the brain can influence this process. “In the past, sex determination studies were specially focused on the gonad, probably because in genetic sex-determining genes are initially active in the gonad, driving the development of the testis,” explains Fernandino, “I was surprised at how long it took us to change our focus that sexual determination begins exclusively in the gonad.”
A schematic representation of the proposed mechanism for sex reversal in medaka fish. Credit. Diana Castañeda
After identifying the hormone produced in fish growing in high temperatures, the researchers used CRISPR/Cas9 genome-editing technology to block the ability of cells to detect it. Importantly, this stopped genetic females developing into males, “I was happily surprised that using gene editing we obtained a complete suppression of masculinization induced by temperature” says Fernandino, “our results highlighted for the first time the participation of the brain as a transducer of environmental stressors, directing development of the testis in genotypic females.”
Identifying how sex reversal occurs has important implications for how some species of fish reproduce. Increasing temperatures and warmer waters might result in more male fish and fewer females. Not only would this affect the ability of fish to reproduce, but also the repercussions on fish populations might have knock-on effects for the ecosystem and fishing industry. “Understanding the molecular mechanisms behind heat-induced masculinization is of great importance for controlling sex ratios in aquaculture and to predict the potential effects of climate change in an important group of animals,” adds Fernandino.
The Maehr Laboratory is seeking a highly motivated postdoctoral research fellow to investigate the molecular basis of immune syndromes and immune cell development.
The applicant will be taking on experimental and/or computational projects in a collaborative cross-disciplinary group. Envisioned projects integrate pluripotent stem cell differentiation approaches with applied developmental immunology approaches and functional genomics, to decipher the molecular underpinnings of human immune syndromes such as autoimmunity and immunodeficiency. In addition, the collaborative project will apply disease models, single-cell omics, and computational analysis strategies based on data-integration and machine learning.
The dynamic and collaborative Maehr lab is embedded in the cutting edge research environment of the Program in Molecular Medicine and Diabetes Center Of Excellence at the University of Massachusetts Medical School. Please visit our webpage for full list of affiliations and more information about us: http://maehrlab.net/ (web) and @MaehrLab (twitter)
Candidates should possess a Ph.D. and have a strong background in immunology, developmental and/or computational biology. Experience with stem cell differentiation, bioengineering and/or computational approaches is desirable. Excellent communication, writing, and collaboration skills are essential.
Interested candidates should email a cover letter and CV to Dr. René Maehr (rene.maehr@umassmed.edu).
 (No Ratings Yet)
(No Ratings Yet) (15 votes)
(15 votes)
 (2 votes)
(2 votes)