The Company of Biologists’ Workshopsprovide leading experts and early career scientists from a diverse range of scientific backgrounds with a stimulating environment for the cross fertilization of interdisciplinary ideas.
The April 2019 Workshop looks a treat for fans of developmental gene regulation. Co-organised by Benoit Bruneau and Joanna Wysocka, ‘Chromatin-based regulation of development‘ will bring together scientists who can or would like to bridge chromatin-level gene regulation, quantitative approaches and developmental biology. It will be held at the beautiful Wiston House, a 16th century Grade I listed building at the foot of the South Downs in West Sussex, England.
The Company of Biologists are offering around 10 funded places for early-career researchers to attend our Workshops along with the 20 speakers. It’s an incredible opportunity for young researchers – find out more here:
Group Leader in Biological Imaging (based at Monash University, Melbourne , Australia)
Expressions of interest are invited from scientists pushing technological boundaries in biological imaging, who seek to make major discoveries running their own independent research program. This EMBL Australia Group Leader position is hosted within the Biomedicine Discovery Institute at Monash University, Australia.
The successful applicant will be housed within the Department of Anatomy and Developmental Biology, one of the five Monash BDI departments. Areas of research focus include renal, lung and reproductive biology, inflammation and cell death, epigenetics, embryology, cancer, stem cells, and cellular reprogramming.
Candidates with a background in developing and/or applying cutting edge imaging technology to address major biological questions are invited to apply. The University particularly welcomes applicants with significant experience and interest in lightsheet microscopy, super-resolution microscopy, and correlative light and electron microscopy.
Welcome to our monthly summary of developmental biology (and related) preLights.
It’s been almost eight months since the launch of preLights, and we are very excited about the hugely positive response from the community so far and preLights’ rapid growth – we recently reached two hundred preprint highlights! To further grow this service, we will be announcing a call for recruiting new team members next month, so watch this space if you would like to join us as a preLighter.
The preLights posts from September turned out to be very diverse: from worms and insects to plants and fish, and also plenty of exciting novel techniques. We hope you enjoy this selection!
From Development to Disease
The early C. elegans embryo is a powerful model to tackle questions like how polarity initially emerges in a developing organism. Giuliana Clemente covered two studies that revealed a new role for Aurora A in symmetry breaking and polarity establishment in the worm.
Sarah Bowling discussed why cells die when they are not in the “right place” and highlighted a preprint showing that disrupted EGFR signaling, rather than a fitness recognition event through cell competition, is responsible. The preprint authors gave further insight into the study (check out the author’s response section) and pointed out that the simplest explanation to their question proved to be the correct one.
Erik Clark’s preLight showcased a conserved role for a posttranslational module (involving the micropeptide millepattes) in segmentation across many insect species, but a lack of segmentation phenotypes in Drosophila mutants. While they may not be good models for insect segmentation, Drosophila has provided numerous mechanistic insights into cell and tissue dynamics through the study of its developing egg chamber. Sundar Naganathan highlighted a preprint showing that in this model, 3D tissue elongation occurs through a mechanochemical signaling pathway that translates the external stiffness gradient into altered cadherin trafficking. Amanda Haage’s preLight also featured some fine mechanobiology in cancer cells, showing that actomyosin contractility regulates the activity of PKA, which is important for mechanically guided cell migration (durotaxis).
Figure taken from preprint by Ray et al.; highlighted by Erik Clark
From the plant development field, Martin Balcerowicz wrote about a study showing that two new proteins (Sol1 and Sol2) regulate fate transitions and cell divisions during Arabidopsis stomatal development.
Moving to zebrafish, Alberto Rosello-Diez featured a novel mechanism that initiates regeneration upon injury: in response to ROS signaling, Hegdehog-expressing cells are repositioned to create a new signaling center. Andreas van Impel reviewed zebrafish vasculature development and how the 50:50 balance of intersegmental arteries and veins is regulated by a flow-dependent mechanism. Excessive blood vessel formation, on the other hand, is a hallmark of cancer; Coert Margadant highlighted a promising strategy to inhibit blood vessel formation by blocking recycling of phosphoinositide lipids.
Several preprints harnessed the power of single-cell transcriptomics to gain insight into neural development and neurodegenerative disease. Zheng-Shan Chong covered a study exploring microglial development through scRNA-seq, which discovered a subset of postnatal neural cells resembling degenerative disease-associated microglia (DAM). Yen-Chung Chen preLighted two preprints looking at why not all neuron types are equally vulnerable in neurodegenerative diseases. Applying new transcriptomic techniques, LCM-seq and Axon-seq, potential protective mechanisms utilized by resistant somatic neurons were found. Finally, Carmen Adriaens’ preLight also dealt with high-throughput sequencing (promoter capture Hi-C), and explained the links between SNP-s associated with type 2 diabetes and chromatin architecture of human pancreatic islet cells.
Tools & Technologies
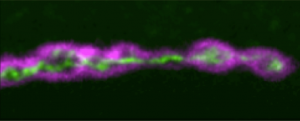
It’s difficult to imagine where the developmental biology field would be today without in situ hybridization techniques that detect mRNA localization. This month, Yen-Chung Chen highlighted work that developed a new fluorescent in situ hybridization method termed SABER, which is more sensitive, cheaper and quicker compared to previous FISH methods. To look at fluctuations in mRNA levels and whether these persist over multiple cell divisions, the preprint covered by Leighton Daigh combined Luria and Delbrück’s fluctuation analysis with population based mRNA sequencing (this new method was termed MemorySeq).
Figures taken from preprint by Kishi et al.; preLighted by Yen-Chung Chen
The number of preprints dealing with CRISPR technology is rapidly growing, but there is still a lot to be understood regarding the targeting and editing accuracy and efficacy in this system. Here, Rob Hynds preLighted a study that uncovered some general principles of CRISPR gene editing by analyzing a large set of indel profiles.
Jacky Goetz wrote about a new method that is able to quantify mechanical stress in vivo by injecting polyacrylamide beads into living embryos and analyzing their deformations. Finally, one of the favorites from this month was Samantha Seah’s preLight that discussed a new way of visualizing data using raincloud plots.
A Postdoctoral position is available in the Spagnoli lab. in the Centre for Stem Cell and Regenerative Medicine at King’s College, London. Our team uses interdisciplinary approaches to study pancreas development and stem cells. The candidate will be part of an exciting EU-funded FET-Open consortium aiming at bioengineering pancreatic tissue.
Developing therapies for pancreatic diseases, such as diabetes and pancreatic cancer, is hampered by a limited access to pancreatic tissue in vivo. Engineering three-dimensional (3D) tissue models, which accurately mimic the native organ, have great potential in biomedical applications, providing powerful platforms for studying tissue development and homeostasis and for modeling diseases. We have established a multi-disciplinary FET European consortium with the ambitious goal of developing an innovative bio-printing approach for generating pancreatic tissue. Tissues and organs comprise multiple cell types with specific biological functions that must be recapitulated in the printed tissue. We will biomimic developmental processes to fabricate 3D bio-printed pancreatic tissue units that allow sustained cell viability, expansion and functional differentiation ex vivo.
The Spagnoli lab. is coordinating the Consortium and will be primarily responsible for establishing a 3D-imaging and transcriptome atlas of pancreatic cells and defining cellular sources for engineering pancreatic tissue units. The candidate will be part of a young dynamic team and help in developing this ambitious research program. Moreover, the lab. is based at the Centre for Stem Cells & Regenerative Medicine (CSCRM) at King’s College London, which acts as the nucleus for a vibrant and collaborative stem cell research community, being an ideal environment for a successful postdoc experience.
Qualifications:
Applicants should have a recent Ph.D. degree or M.D./Ph.D. degree. Candidates with experience in stem cell and developmental biology, confocal microscopy, image acquisition and analysis will be preferred. This position seeks a highly motivated individual with a strong interest in developmental biology.
Starting date:
The position is open starting from October 2018 but later entry dates are possible. The initial contract is for one year and can be renewed until a maximum of three years.
Requirements:
A CV and a statement of research interests along with names of 2 referees should be sent via e-mail to the address below.
Contact:
Dr. Francesca M. Spagnoli, Centre for Stem Cell and Regenerative Medicine, KCL, London, UK
On 2nd October 2018, the Mechanobiology Institute in Singapore hosted a meeting of Singapore-based researchers using fish in their work. We wanted to bring together the local community to build support and promote future research collaborations within Singapore. This meeting follows successful international conferences last year in Singapore, where the importance of fish in both fundamental and translational research was apparent (https://thenode.biologists.com/developmental-biologists-meet-lion-city/events/, https://www.zdmsociety.org/past-events).
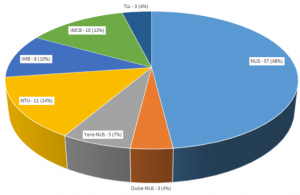
The fish community in Singapore, despite running up against the usual funding challenges and regulations, remains a formidable group. Over 80 people attended during the afternoon, with the room literally full to bursting, Figure 1. Researchers came from National University of Singapore, Nanyang Technological University (both universities recently built state-of-the-art fish facilities), A*Star and Duke-NUS (Figure 2).
Figure 1. Top: Timothy Saunders addressing an overwhelming crowd of scientists in the Singapore Fish Meeting 2018. Bottom: Anticipation of the first talk.
The half-day meeting saw presentations by twelve young scientists (students, post-docs and research associates). There was also a poster session going on late into the evening, including one on a wide-screen LED TV (Figure 3). Notably, women presenters made up more than half (9/12 talks and 6/11 posters) of the presenters, deriving from a roughly equal proportion of male and female participants. The talks comprised a wide-ranging topics from fundamental biology, through to medical applications and engineering. A clear strength was in the use of fish in disease modelling, but there were also interesting presentations related to ageing, behaviour, and fluid mechanics. It was a powerful demonstration of the versatility of fish models for studying a broad range of topics. It was particularly stimulating to get fresh research ideas and tools, and also to immerse ourselves in the work of our colleagues covering unfamiliar territory.
Figure 2. Affiliation breakdown of participants in the Singapore Fish Meeting 2018. NUS – National University of Singapore (including the Cancer Science Institute and Mechanobiology Institute); IMB – Institute of Medical Biology; IMCB – Institute of Molecular and Cell Biology; NTU – Nanyang Technological University; TLL – Temasek Life Sciences Laboratory.
The day finished with pizza and wine, which provided a platform for cross-fertilization of expertise to spawn new ideas and projects. The fish meeting was a wonderful success – great talks, lively discussions, and good food. It was also an important opportunity to bring together the local fish community. We believe that the fish community in Singapore will continue to do great science and be a driving force of scientific discoveries through inter-disciplinary cooperation.
Figure 3. Abraha (hidden) presenting his Next-Gen poster to a group of curious scientists.
The community would also like to take this opportunity to remember the late Professor Hong Yunhan from National University of Singapore, who pioneered the generation of haploid medaka stem cells.
About the Authors:
Dr. Jason Lai has just graduated from the Stainier Lab in Bad Nauheim, Germany to pursue his post-doctoral training with Timothy Saunders. He is interested in growth and form, particularly related to the Hippo/YAP pathway (https://www.researchgate.net/profile/Jason_Lai6)
Emergence of functional polarity in a tubular epithelium
Epithelial tubes often have a functional polarity written along their proximo-distal (P-D) axis, with different segments of specialised cell-types carrying out distinct physiological activities. With a handful of notable exceptions, we know very little about how P-D axes and segment-specific differentiation are regulated during organogenesis.
The major objective of this project is to understand the molecular and cellular mechanisms that pattern and maintain functional polarity along the P-D axis in a structurally simple, but functionally sophisticated epithelial tube: the insect renal tubule.
The project will be carried out in the laboratory of Barry Denholm (Biomedical Sciences, University of Edinburgh)
Location: School of Biological Sciences, University of East Anglia, Norwich, UK
Project Description:
There is much interest in the possibility of growing human organs from patient stem cells in vitro (a.k.a. ‘organoids’). In addition to regenerative therapies, this could furnish new disease models and facilitate drug development. By learning how tissues and organs self-organise in the embryo, we can rationally design strategies to improve and expand organoid technologies. Our research uses the chick embryo to investigate the development of our primary sense organ, the eye.
It is known that stem cell cultures can spontaneously self-organise to form retinas in vitro, but it is not know how they do this. This PhD project aims to investigate a Pax6 gene network that may drive self-organisation of the early retina. Pax6 was named a ‘master control gene’ for eye development, and we recently identified a network of Pax6-interacting genes with the potential to spontaneously self-organise the early retina, either in the embryo or in vitro. The project will directly test this possibility by measuring the biophysical properties of key gene products and comparing them with values predicted by computer simulations of early retinal development.
Through this project you will develop transferable ‘wet lab’ skills in molecular, cell, and developmental biology, and advanced imaging. You will receive rigorous training in quantitative analytical approaches, with an added opportunity to gain new skills in computational biology (computer programming, mathematical modelling).
For informal enquiries and further information, please contact: Dr Tim Grocott (t.grocott@uea.ac.uk)
The Mammalian Genetics and Development Workshop is an annual meeting focusing on the development and genetics of mammals. The Meeting is based on the submitted abstracts, and include diverse topics ranging from mammalian development (not exclusively human or mouse) and identification of disease genes and developmental mechanisms, to human genetics and epigenetics. Other model systems (including Drosophila, zebrafish and chick) are also welcomed where these relate to general developmental questions and/or disease models.
The meeting will be similar format to the 28th Meeting in 2017, which had a fantastic set of short talks covering various aspects of developmental biology, mouse models of human disorders and genetics of human disease (abstracts are published at https://doi.org/10.1017/S0016672317000076)
The workshop is traditionally a venue for post-docs and PhD students to talk rather than laboratory heads and is an excellent training ground and a friendly, informal forum. In keeping with this objective, we offer TWO PRIZES of £150 to individual post-graduate/post-doctoral presenters. In addition, thanks to the generosity of Mammalian Genome (https://link.springer.com/journal/335) publishers, we will offer an additional PRIZE of £150.
Registration: A £10 registration fee is payable by all attendees on arrival at the meeting. This fee covers the abstract booklet, tea and coffee refreshments and the wine reception. Speakers and chairpersons will be provided with lunch, free of charge, on the day of their presentation.
Abstract Submission: All Workshop presentations will be in lecture format (15 or 20 minutes). Please send your abstract by e-mail (Word or rtf file) to ich.mgdwshop@ucl.ac.uk by 5pm on 8th November. In addition please indicate the length of talk you prefer.
Abstract format is as follows: Title (bold), AUTHORS (ALL CAPS), Address (italics) and text (200 word limit). Abstracts will be published (with the authors’ permission) in Genetics Research (https://www.cambridge.org/core/journals/genetics-research).
We have a vacancy for a postdoctoral research associate, funded by a BBSRC grant, to work on the roles of spastic paraplegia gene products in formation of axonal ER, and identification of new genes and cellular processes involved in this. For some background on the project, see: https://elifesciences.org/articles/23882.
Applicants should have a PhD in a biological science, interests in neuronal cell biology, and experience in as many as possible of the following: model organism genetics, genetic screens, neuronal cell biology, light or electron microscopy, live imaging, molecular biology, and bioinformatics.
The Academy of Medical Sciences is convening a scientific meeting on the 19 and 20 March 2019, in Oxford. Abstract submission deadline: 5 November 2018.
This meeting will explore key areas of neurodevelopmental research in a unique forum that emphasises discussion and collaboration between disciplines, career stages and sectors.
One of the key aims will be to highlight the latest advances in neurodevelopmental research and identify key research questions that could bring a real impact to the field. The meeting’s talks and discussions will span a broad range of neurodevelopmental and disorder research, from in utero to adolescence, and from multiple perspectives, including molecular, cellular, circuit/systems and behavioural levels.
We welcome researchers from all backgrounds to attend this meeting and strongly encourage early career researchers, from PhD level onwards, to apply. Registration is free.

 (No Ratings Yet)
(No Ratings Yet)
 (1 votes)
(1 votes)