The Karam Teixeira laboratory (https://www.gen.cam.ac.uk/research-groups/karam-teixeira) at the University of Cambridge (Department of Genetics) is looking to recruit an outstanding Postdoctoral scientist to investigate the molecular mechanisms sheltering totipotency and controlling germline stem cell behavior in vivo. Using the Drosophila germline as a model for studying stem cells, we employ an integrated approach, combining high-throughput molecular analysis (next-generation sequencing) and computational investigation with developmental, microscopy, and genetic analyses (including CRISPR-Cas9 gene editing, tissue-specific RNAi knockdown, etc). We were previously able to assemble the complete genetic framework controlling germline stem cell self-renewal and differentiation in vivo, revealing conserved new aspects of stem cell biology (Teixeira et al, Nature Cell Biology, 2015; Sanchez et al, Cell Stem Cell, 2016). Moving forward, our goal is to build a refined molecular understanding of how protein synthesis control – a new frontier in gene regulation – governs stem cell fate transitions in vivo. Our lab is generously funded by the Wellcome Trust.
Candidates must have experience in a wide range of molecular biology techniques, and prior expertise in next generation sequencing would be an advantage. Experience working with fly genetics is a plus but not required. The successful candidate will be highly motivated, willing to join a young and dynamic research group, have good communication skills, and possess strong problem solving capacities.
How do phenotypic differences between cells of the same clonal origin emerge? How exactly does the transition between the initial and final phenotypes occur? What happens in the cell during the transition? When there are two or more options, how is the choice made between them? How long does it take to acquire a new phenotype? What is the minimal difference required to consider two cells as phenotypically different? To define a cell type, should we consider only morphological, molecular differences or both simultaneously?
Despite the plethora of studies, these simple questions remain unanswered. Most of the studies focus on identifying the essential genes or environmental factors usually at the level of cell populations. The huge amount of molecular data accumulated over the past decades gave us the illusion of knowledge. Knowing the players is obviously essential, but this is only the starting point for the understanding. Unfortunately, our understanding of the process of differentiation remains desperately scarce, as the lack of clear responses to the above listed simple questions shows.
Recently, two important challenges came to modify the perception of differentiation. The first comes from the spectacular development of single-cell technologies. The second is that we have to come to realize how much phenotypic plasticity is a genuine characteristic of cells. The main lesson from the rapid development of single-cell detection techniques is the unambiguous demonstration of how different individual cells are and how poorly population-level averages represent them. The bulk of our knowledge on differentiation comes from studies of cell populations. Perhaps unconsciously, we took for granted that individual cells all follow with small variation the same sequence of events as what we could see at the level of cell populations. The unexpectedly high variation of individual cell phenotypes in populations that were believed to be homogenous (because of the morphological similarity of the cells, their clonal origin, their expression of some markers etc.) draw the attention to the phenotypic plasticity of the cells and to the fact that fate decisions are “taken” by individual cells.
Clearly, a coherent, systemic level explicative frame is needed that can account for the coherent population-level behaviour emerging from highly variable individual cell phenotypes and behaviour.
Our recently published work [1] was motivated by the wish to contribute to this effort. Although we used the extensively studied hematopoietic stem cell model, we were surprised how much these general questions fit to the model. The definition of the hematopoietic stem cell is widely debated, no precise description of the earliest events of differentiation and only very scarce information on the morphological changes during the same period were available. The study of hematopoietic stem cells is made difficult by the lack of exact criteria to identify them. Nonetheless, there is a consensus that CD34+ cell fraction in the umbilical cord blood contains high number of these cells. We decided therefore to privilege an integrated view and work on the whole population of CD34+ cells. Individual cells were randomly sorted from the population at different fixed time-points and their gene expression profile was analysed by single-cell RT-PCR. The structure of the population and its components were identified on the basis of the collected single-cell gene expression data. Parallelly, we set up a time-lapse system that allowed the continuous monitoring of the cells and their progeny during the first 96hrs after stimulation. Adding the continuous observations of the morphological changes and cell division timing of individual cells to the single time-point single-cell molecular analysis sampling of the same population provided a glimpse of the true dynamic nature of cellular fate decision.
The CD34+ cell fraction is traditionally considered as heterogeneous. Indeed, before cytokine stimulation every cell displayed a unique gene expression profile. However, no groups could be identified on the basis of their statistical similarity, this population is not a mixture of a limited number of “cell types”. When cytokines were added to the culture, every cell responded in a unique way. Again, every cell displayed a unique gene expression pattern that was different of the previous seen at t=0 hours. It was characterized by the simultaneous expression of different lineage-specific genes. This state is known as a multi-lineage primed pattern [2, 3]. A second round of change occurred during the second 24 hours. Two days after the stimulation of the cells two distinct transcription patterns emerged. One pattern was typical for myelo-erythroid progenitors, while the second was reminiscent of multipotent cells. Until now, the results overall confirmed the previous studies; the main novelty was the apparent rapidity of transition from the initial to a multilineage primed gene expression pattern and, just 24 hours later, to two distinct profiles. However, these snapshots did not allow us to deduce on the dynamics of the changes.
Time-lapse video of a cell clone with cells conserving polarized morphologies. GIF made from from Movie S1 in the paper
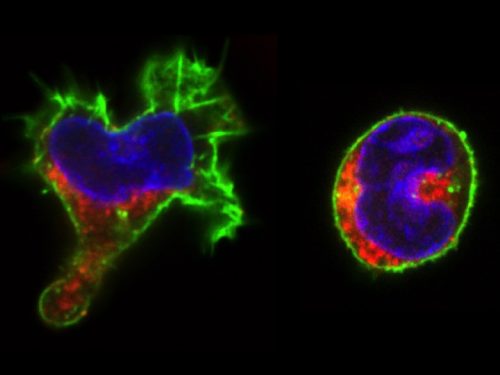
The real surprise came from the analysis of the time-lapse records. Individual CD34+ cells were placed in microwells and imaged for a week at 1 image/min. The resulting time-lapse records allowed us to record cell cycle lengths and morphological changes of each individual cell within individual clones. After stimulation, the cells usually displayed a polarized shape with a strong protrusion on one side called uropod. The first unexpected observation was to see that the unusual length of the first cell cycle. The cells made more than 50 hours on average to divide. This means, that the first major transcriptional change occurred during the first cell cycle and the second around the end of the first or the very beginning of the second cell cycle. After the first division, we could see two different morphologies; one was strongly polarized with a uropod, the second is spherical. The daughter cells usually inherited the morphology of their mothers. Polarized cells gave two polarized and round cells two round daughters. However, we were surprised to observe that a significant proportion of the cells did not conserve a stable morphology; they switched from one morphology to another and back many times during the cell cycle. The majority of their daughter cells also conserved the fluctuating phenotype
Red color: CD133; Green: actin; Blue: DNA
We called these cells “hesitant”. The overall picture suggest that stimulated CD34+ cells, after a brief passage through a multilineage primed state reach (without cell division!) one of the two alternative states characterized by a typical transcription pattern and cellular morphology. However, a significant proportion of cells fluctuate between the two morphologies. Does this morphological instability reflect transcriptome fluctuations? To answer this question, we have isolated individual cells with the three different – stable round, polarized or “hesitant” – behaviours and analysed their gene expression pattern. It appeared that the cells with stable morphology displayed one of the two expression patterns first observed at the 48 hors time point. The “hesitant” cells were characterized by an intermediate profile. The molecular analysis correlated to the time-lapse data suggests therefore that these cells are in an unstable state; their transcriptome undergoes fluctuations that are reflected in their fluctuating morphology also.
These are the key observations and the immediate conclusions of this work. However, these observations may contribute to the current tendency to reframe the issue of cell differentiation and stem cells in general. Cell differentiation can be approached using the concepts of stability and change – two complementary concepts widely used in biology. Stem cells may represent a highly unstable cell fraction contrary to the cells with stable differentiated phenotype. Unstable stem cells are actively exploring the space of available phenotypes before getting trapped by one of them. This is a kind of trial-and-error process. Under normal conditions, the unstable period is relatively short lasting; this is why the unstable cells we consider as stem cells are so rare in a normal tissue. However, a substantial change in the environment can destabilize many cells at the same time. This is what we see when CD34+ cells are stressed by the sudden addition of a cytokine cocktail. Due to the progressive adaptation to the new environment, the proportion of the “hesitant” stem cells decreases gradually as they attracted to the more adapted phenotypes. Importantly, this process seems to depend only indirectly on cell divisions. This interpretation is in remarkable agreement with earlier theoretical predictions and experimental work [4-8] and supported by recent experimental observations also [9-12].
Our paper was initially submitted to a well-known journal in stem cell biology. Beyond the disappointment of the rejections (every scientist is used to that), we were surprised by the poor quality of the reviews. The referees raised some technical concerns about the single-cell RT-PCR versus single-cell RNA sequencing, but not a single word about the time-lapse experiments that represented the major part of the paper, nor about the coupling the molecular and cellular scales, which is the true originality of the work. The reviews were very different when the manuscript was submitted to PloS Biology. The comments concerned all aspects of the work and the suggestions truly helped to improve the final version.
It would be naïve to think that a single paper can answer the fundamental questions raised at the beginning of this text. Clearly, we need a fresh view on cell differentiation that goes beyond the simple gathering and classification of molecular data and takes into account the true dynamics.
Mojtahedi M, Skupin A, Zhou J, Castano IG, Leong-Quong RY, Chang H, et al. Cell Fate Decision as High-Dimensional Critical State Transition. PLoS Biol. 2016;14(12):e2000640. doi: 10.1371/journal.pbio.2000640. PubMed PMID: 28027308; PubMed Central PMCID: PMC5189937.
Doing a PhD is tough, the data from surveys supports that. However it is not insurmountable, and here we have a collection of some guidance from the Twitter community. Let us know in the comments if you have any thoughts to add.
The Department of Biology within the College of Natural and Health Sciences at The University of Tampa invites applications for a tenure track position in developmental biology at the rank of Assistant Professor starting in August 2018.
The University of Tampa is a medium-sized, comprehensive, residentially-based private institution of 8,913 undergraduate and graduate students. The University is ideally situated on a beautiful 110-acre campus next to the Hillsborough River, adjacent to Tampa’s dynamic central business district, which is a growing, vibrant, diverse metropolitan area. UT reflects this vibrancy; with 20 consecutive years of enrollment growth UT boasts 229 student organizations, a multicultural student body from 50 states and 140 countries, and “Top Tier” ranking in U.S. News and World Report.
Primary responsibilities will include an undergraduate teaching load of 12 contact hours per semester. The candidate is expected to teach introductory biology for majors, an upper division course in developmental biology, and other courses as needed.
Secondarily, the candidate is expected to engage in scholarly and research activity that involves undergraduates, advise students, and provide service to the department, college, university and broader community. Research activities must yield peer-reviewed publications.
PhD preferred (advanced ABD candidates considered), prior teaching and research experience with undergraduates is desirable.
Salary for this position is competitive and commensurate with experience.
Review of applications will begin January 2, 2018, and continue until the position is filled. Limited start-up packages and modest research space are available for tenure-track positions.
In 2016, the BSDB introduced the Cheryll Tickle Medal, which is being awarded annually to a mid-career, female scientist for her outstanding achievements in the field of Developmental Biology. The BSDB is proud to announce the 2018 awardee Christiana Ruhrberg. The medal will be presented at next year’s Spring Meeting where Christiana will give the Cheryll Tickle Award Lecture.
Christiana Ruhrberg studied Biology at the Justus Liebig University (Giessen, Germany), and obtained her first class Diploma/MSc degree in 1992. After taking on an MSc/research assistant position at the University of Sussex for two years to elucidate genetic changes in ovarian cancer, she moved to Imperial College London to work for another two years to study the genomic organisation of the gene-rich human ‘surfeit’ locus. She then carried out her PhD project in the laboratory of Fiona Watt at the Imperial Cancer Research Fund (1994-97) where she identified and described the function of the envoplakin and periplakin genes. Her postdoctoral research in the laboratories of Robb Krumlauf at the National Institute for Medical Research in London (1997-1999) was funded by a MRC postdoctoral training fellowship and dedicated to studying the role of Hoxa1, Hoxb1 and Hoxb2 during hindbrain development. During her second postdoc with David Shima at the Imperial Cancer Research Fund in London (2000-2002), she was funded by a ICRF fellowship and worked on VEGF-A-mediated blood vessel branching. Having received an MRC Career Development Award in 2003, she became an independent investigator at University College London’s Institute of Ophthalmology studying links between vascular and neuronal development, with particular focus on the roles of VEGF and SEMA3A signalling during facial nerve and blood vessel formation. Staying at that same institute, she was appointed Lecturer in 2007, promoted to Reader in 2008 and then full Professor in 2011. Christiana has an excellent record in obtaining research funding, as illustrated by a Wellcome Trust Junior Investigator Award in 2011 and a Wellcome Trust Investigator Award in 2016 for her research on vascular biology and neurovascular interactions.
Figure: Using the mouse embryo hindbrain to elucidate neuronal and vascular development. (A) The hindbrain from an embryonic day (E) 10.5 mouse was dissected to perform visualise the expression of Hoxb1 in rhombomere 4 (A), the origin of the Isl1-positive facial branchiomotor (FBM) neurons, which can be observed during their caudal migration by Isl1 at E12.5 (B). (B-F) Main cell types (B) and cell interactions (C-F) in the developing mouse hindbrain. In (C-F), dissected hindbrains were stained with the vascular endothelial marker IB4 (red) and markers for mitotic neural progenitors, microglia or neural progenitor processes, shown in green in C-E, respectively). Note that neural progenitors attract sprouting blood vessels (C), the physical contact between microglial and endothelial processes (D) and that neural progenitor processes contact vasculature (E).
Christiana has been a mentor to 11 PhD students and 6 postdocs, 3 of whom have now established themselves as independent investigators. She has published many influential papers in the fields of vascular, neuronal and neural crest cell development, including primary research articles and reviews, methods papers and book chapters. Twelve of her research papers have been listed as recommended reads by the Faculty of 1000, five have been featured with cover images and six have been featured in editorials in influential journals such as JCB, PNAS, Nature, Science, Neuron and JCI.
Christiana received an impressive amount of honours, including the title ‘Young Cell Biologist of the Year’ (BSCB, 1996), the ‘Werner-Risau-Prize’ (German Society for Cell Biology, 2003), the MRC’s ‘Science Heirloom’ (Suffrage Science, 2011). She was also named an ‘Academic Role Model’ (UCL, 2013) and is listed in EMBO’s ‘Expert Women in Life Sciences’ (2013) and ‘AcademiaNet’ (Robert Bosch Stiftung, Germany, 2014). Besides all these achievements, Christiana takes on regular institutional responsibilities, is a member of various grant committees and a contributing member of the Faculty 1000, editor for PLoS One, and serves on programme committees of a number of scientific meetings.
The BSDB makes it a tradition to ask the Cheryll Tickle Medal awardees a number of questions concerning our field and its future. Please, read Christiana’s answers below.
What were the questions that inspired you to work in the field of Developmental Biology?
I was initially attracted to working in the field of developmental biology when the first mouse knockouts became available. Many of them had lethal phenotypes, making embryological studies imperative to determine the physiological functions of the ablated genes. I initially examined mouse knockout models to identify molecules that regulate the migration of facial branchiomotor neurons and subsequently to determine how the growth factor VEGF orchestrates blood vessel morphogenesis. By answering two different biological questions with the mouse embryo hindbrain as a model system, I serendipitously identified VEGF as the elusive migratory cue for facial branchiomotor neurons. This finding inspired me to continue investigating VEGF functions in neuronal and vascular development, with a more recent strive to apply knowledge gained through developmental studies also to further our understanding of disease processes in the adult.
Why should young researchers continue to engage in Developmental Biology?
Personally, I have always been fascinated by how the vertebrate body develops to enable postnatal life. Developmental studies also yield striking images of a multitude of diverse cellular processes that coordinate organ morphogenesis, making this type of research not only exciting, but also aesthetically pleasing. The PhD students, postdocs and technicians who train in my laboratory share these sentiments and have gained much deserved appreciation for their developmental biology research work through journal cover images and when winning presentation prizes at conferences. Developmental biology research also impacts on public health, because understanding how embryonic processes yield functional organs informs regenerative medicine. In particular, knowing how functional tissues are built normally might one day soon provide a gold standard for designing therapeutic strategies to recreate or repair dysfunctional tissues. That said, the developmental biology community is increasingly faced with the challenge of having to convince funding agencies that developmental biology research can underpin research into tissue repair and regeneration!
Which were the key events or experiences in your life that influenced your career decisions and paved your path to success?
My career path has not been straightforward, but encompassed a series of obstacles and opportunities. For example, the failure to appoint a successor for our retiring genetics professor at my home university in Germany could have persuaded me to switch subjects from molecular biology and genetics to a different one to avoid a significant delay to graduation, but I took this challenge as an opportunity to study for a year in the UK, being the first student on my course to embark on such an overseas placement. I ended up staying at the University of Sussex for almost 2 years to complete a research project all the way to publication, returning to Germany only to submit my thesis. Incredibly, after all the long hard work, I almost did not graduate, because those in charge at my German University deemed an English-written thesis unacceptable! This experience encouraged me to return to the UK to continue academic research in an English-speaking environment. Unfortunately, I initially chose a PhD supervisor at the Imperial Cancer Research Fund who turned out to be rather unsupportive of my endeavours; yet, I was able to make a ‘lateral’ move within the same organisation to re-start my PhD with Fiona Watt. She was a most inspirational PhD supervisor and mentor, and later matched me with an ideal postdoc supervisor in Robb Krumlauf at the National Institute of Medical Research. In Robb’s lab, I discovered both my love for developmental biology and the inspirational community of scientists working in this area. In a nutshell, it was not careful planning that allowed me to get to my current career stage, but my unwavering enthusiasm for research combined with resilience when faced with adversity and the will to take advice and encouragement from my fabulous mentors Fiona and Robb.
What advice do you give young researchers towards a successful career?
Whether you choose to continue on an academic career or embark on an alternative career, I recommend everyone to take advantage of transferable skills training to complement the technical training gained in the research environment. Strengthening your verbal and written communication skills, learning about project and team management and developing effective networking skills will provide a strong foundation to equip you for success in a diverse range of career options.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our first report from the 2017 group of student awardees comes from Josie Elliott(student at University of Oxford), who undertook her studentship withAlison Woollard at the Dept. of Biochemistry in Oxford.
What is the role of wrt-2 and wrt-4 in left-right asymmetry in C.elegans?
Once upon a time a genetic screen identified a signalling pathway that caused Drosophila melanogaster embryos to develop a ‘lawn’ of denticles rather than forming them only at parasegment boundaries. Thus the so-called Hedgehog signaling pathway was born (Nusslein-Volhard & Wieschaus, 1980). This pathway has revealed itself to be one of the core signal transduction pathways in regulating animal development. Through gene duplication and diversification events, different types of hedgehog proteins have been found across vertebrates. The brilliantly named Sonic Hedgehog ligand is the best studied ligand present in mammals. Loss of expression of Sonic Hedgehog in mice causes defects in left right asymmetry (Levin et al., 1995).
The story was expanded into C.elegans by bioinformatics (Burglin, 1996). A search of the C.elegans genome for homologues of the carboxyl terminal ‘hog’ domain revealed a family of proteins which did not contain the amino terminal ‘Hedge’ domain but instead a novel domain dubbed ‘Wart’. Thus the warthog (wrt) genes were named. Further genome analysis revealed genes which contained the amino wart domain but lacked the carboxy hog domain. Together this made a family of ten warthog genes.
Previous research in my lab (the Woollard lab in the Biochemistry department at the University of Oxford) had linked the wrt-2 branch of the warthog family (figure 2) to defects in left right asymmetry, with wrt-2 and wrt-4 giving the highest penetrance phenotypes. However it was found that the GFP marker used in the previous strains to help quantify another phenotype (related to vulval development) caused defects of its own. This is where I come in. My project was to cross and create strains of worms without this GFP background and then requantify the left right asymmetry defects. I studied the single mutants wrt-2, wrt-4, wrt-8, as well as the double mutants wrt-4;wrt-2, wrt-8;wrt-2, wrt-8;wrt-4 and the triple mutant wrt-8;wrt-4;wrt-2. The double and triple mutant strains I created myself. The combined mutations allow investigation into redundancy between the genes, giving a glimpse into their evolutionary history. The wrt-7 gene has been shown to have no expression pattern and is likely a pseudogene, so due to my limited time in the lab I didn’t quantify the wrt-7 mutations.
Fig. 2 The tree presents the origin of the ‘Wart’ domain in C. elegans’ most distant relative in the Rhabditiada order, Brugia malayi, as well as its two closet relatives in the Caenorhabditis genus, C. briggsae and C. remanei. A multiple sequence alignment of Wart domains was used to generate this unrooted Neighbour Joining tree (default setting Clustal_X). Results of 1000 bootstrap trials are shown. Figure from Emily Baker’s dissertation.
On a side note: the C.elegans species is great. They’re heamaphrodites so you can just leave them on an agar plate to reproduce by themselves. Males do exist – enabling you to perform genetic crosses. They’re transparent, so no need to dissect anything. You can even freeze them and they’ll be alright in liquid nitrogen until someone needs them!
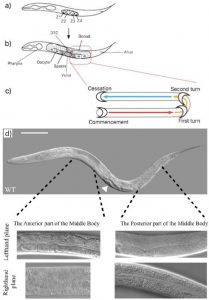
At first glance C.elegans may not seem very asymmetric. However one source of left right asymmetry is the relative positioning of the gonads and intestine either side of the vulva in the worm. This is best shown through pictures. In the wildtype worm (figure 3d) the picture in the lefthand plane shows that in the anterior part of the worm one can see intestinal cells and the intestinal lumen going down the centre. In the posterior part of the worm one can see the U shaped gonad that migrates away from the vulva (just visible in the bottom left hand corner of that picture) along the body, turns twice then travels back along the body (schematic figure 3c). The top picture of a whole worm (figure 3d) shows the opposite situation (the worm is laying on its other side) where the gonad can be seen at the anterior part of the body and the intestine at the posterior.
Fig. 3a) L1 stage animal. b) L4 stage animal. The gonad turns twice due to the migration of the distal tip cell (DTC). c) Commencement sees the gonad arms depart from the ventral midpoint and travel along the ventral sides of the worm (red arrow); firstly turning to the dorsal side of the animal and secondly along the anteroposterior axis (yellow arrow) until they meet again, this time at the dorsal midpoint (blue arrow). The gonad migration at the anterior end of the worm is the mirror-image. Furthermore, the anterior gonad is located on the right-hand lateral side of the worm (shown as the bottom plane), whereas the posterior gonad lies along the left-hand lateral side (top plane); however, this is not shown in the image. d) Arrowhead points to vulva. The right-hand lateral side is shown and therefore the anterior, but not the posterior, gonad arm is visible. The alternate scenario in which the posterior gonad is visible in the top plane of view is hypothetically depicted; in which instance, the anterior gonad would be hidden from view by the intestine. Scale bar = 25μm. Figure taken from Emily Baker’s dissertation.
Thus the anterior/posterior distribution of gonad and intestine gives us a tool to study and quantify left right asymmetry in worms. After discussions with Emily (a recent biology graduate who had been working on the project before me) we decided to only quantify worms in which the posterior gonad was visible down the microscope. Thus, worms which I counted looked like those in the two lefthand plane pictures in figure 3d. This decision, although arbitrary in itself, allows our results when published to be more understandable and easier to replicate.
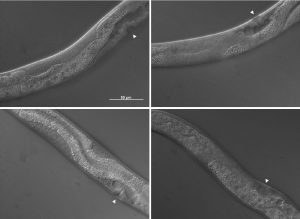
The pictures shown in figure 4 demonstrate some of the mutant phenotypes scored in this project. Basically I was looking for gonad tissue poking through anterior intestine tissue and intestine tissue poking through posterior gonad tissue. Worms with this phenotype were scored as mutant and were also further divided into anterior or posterior defects.
Fig. 4Top left shows the anterior gonad of a wrt-2;wrt-4 worm in which gonad tissue can be seen as well as intestine. Bottom left shows the anterior gonad of a wrt-8 worm in which gonad can be seen above and below the intestine cells. Top right shows the posterior gonad of a wrt-2;wrt-4 worm in which the gonad has migrated unsuually thus intestine tissue can be seen. Bottom right shows the posterior gonad of a wrt-2;wrt-4 worm in which half the gonad has disappeared to the other side of the worm. Photos taken by me on a Zeiss microscope.
One of the most enjoyable parts of this project has been pipetting alongside Emily, who worked on this project before me and is continuing to develop it. In my 8 weeks I have seen first hand a small portion of the amount of research that has gone into studying the warthog genes. However chatting with Emily I have seen how my short project may in future count towards producing a scientific paper. At the moment Emily is taking the story of warthog genes in C.elegans back to its bioinformatics heart, producing a phylogenetic tree of all the warthog genes across the nematode family – trying to link evolution with phenotype. She is also expanding from the wrt-2 branch of the gene family into as many other members as she can get her hands on.
The highlight of my time in the project has been having so much independence in the lab, deciding day to day, week to week how I wanted to spend my time. This has been a fantastic insight into the life of a researcher and totally different to undergraduate labs. The lowlight would have to be the Saturday when I broke a pipette and in the process spilled a box of pipette tips all over my desk…
Thank you so much to everyone in the Woollard lab for being more friendly and welcoming than I could have hoped for.
Scientists at DanStem discovered that cells’ sense of direction (cell polarity) determines their destiny. By discovering the signals that control cell polarity in the developing organism, scientists can now mimic it in a test tube and control the fate of human stem cells. The results accelerate the efforts to generate specialized cells, e.g. insulin-producing beta cells, from stem cells to treat and maybe even cure chronic diseases
The paper, published in Nature Cell Biology, by the Semb Group at DanStem, shows that cell polarity (the cell’s ability to sense what is up and down) in progenitor cells not only contribute to the architecture and shape of organs, e.g. tubular systems, but also governs the fate progenitor cells.
Video: 3D volume rendering of two endocrine progenitor cells (green membrane and red nuclei). The cell on the left is immature and has a larger apical domain facing the lumen of the duct (labeled in red) while the other cell has downregulated the apical domain. Embryonic mouse pancreas grown and imaged live in vitro
Major scientific findings of the paper
The scientists answer a fundamental question as to how the limited number of genes in our genome can control the large complexity of our organism. The results illustrate how genes are re-utilized in a context-dependent manner to expand their ability to control, not only one, but a many complex cellular events during organ formation.
Epidermal growth factor (EGF) signaling is identified as an essential regulator of cell polarity throughout pancreas formation to regulate tube formation (early) as well as the birth of insulin-producing beta cells (late).
The paper answers a long-sought question: how progenitors in the developing pancreas are instructed to become insulin-producing beta cells.
This discovery can be utilized to increase the efficiency and robustness of differentiating human pluripotent stem cells into insulin-producing beta cells for future cell therapy in type 1 diabetes, and deliver new general concepts for how cues from the environment instruct organ-specific multipotent progenitors into their different fates.
The O’Brien Lab (http://www.stemdynamics.org) is part of the Department of Molecular and Cellular Physiology at Stanford University School of Medicine. Our goal is to uncover the ongoing dialog that adult stem cells have with the tissues they support, and understand how this dialog happens on the molecular and cellular levels. We use a simple invertebrate organ, the Drosophila midgut, to pioneer new imaging and computational approaches that address basic questions in stem cell biology.
The O’Brien Lab is seeking a highly motivated, detail-oriented Life Science Research Professional 1 (LSRP1) to join our dynamic laboratory environment. The LSRP1 will perform functions and activities involved in defined research projects, which principally involve live and fixed tissue microscopy of Drosophila organs, computerized image analysis (ImageJ/Imaris), basic MATLAB coding, and construction and maintenance of sophisticated Drosophila genetic stocks. The goals of the projects are to understand cellular interactions and signaling in stem cell based Drosophila organs.
*- At the time of the interview, applicants will be asked to provide evidence of their imaging skills in the form of publications, posters, report, or other documentation.
A postdoctoral position is available in the laboratory of Dr. David Matus at Stony Brook University to investigate symbiotic and developmental processes with Selective Plane Illumination Microscopy. We have recently received funding for three years of postdoctoral support to develop protocols for extensive in vitro and in vivo imaging of cell invasion processes with a focus on the tissue and cellular entry of endosymbiotic algae as they enter their spotted salamander embryo hosts (Ambystoma maculatum). This work is funded by the Gordon and Betty Moore Foundation in collaboration with researchers from Columbia University, The American Museum of Natural History, and Gettysburg College (see Shelf Life Episode 11). The project will combine molecular biology, embryology, cell biology, and extensive light sheet imaging. Preferred candidates will have backgrounds in light sheet and/or confocal microscopy as well as an interest in advanced imaging methods and the intersections of cell and developmental biology with ecology and evolution. Opportunities will exist to develop projects and assist in advanced imaging using C. elegans and zebrafish as well. My laboratory is a part of a modern and well-equipped Department of Biochemistry and Cell Biology at Stony Brook University on Long Island, NY. For further information on our work, please see the following publications on techniques,cell invasion, and the symbiosis.
To apply, please send a letter of interest detailing your expertise, CV and names and contact information of three references to david.matus AT stonybrook.edu. You can also apply directly through the Stony Brook University Human Resources portal here or by searching for position #1702902 at the Stony Brook HR site.
It’s all about the wires. But what about the glue?
Networks make us who we are. I am not talking about social networks but about neural networks that define how we perceive the world and how we act. For a century, neuroscientists have sought to understand functions of neural networks in condition and how such networks are established in the first place still remains an active area of research. Neuronal networks consist of a large set of wires that need to be connected and organized in defined, functional ways, similarly to telecommunication wiring pervading our cities or like the motherboards of our modern computers. To work, signal carriers of such networks must be of appropriate bandwidth, with proper paths, and connected in a specific way. Likewise, in neuronal networks, cell fate must be specified, neurons must migrate to proper locations, grow and navigate their axon processes through specific paths and generate well-defined synaptic connections.
In the 1990’s, the “Decade of the Brain,” as declared by the United States Congress, developmental neuroscientists made extraordinary progress in understanding cell fate determination, axon guidance and synaptogenesis. Seminal studies identified the principal families of guidance molecules and established major paradigms, such as the role of floor plate Netrin, a secreted guidance cue, in commissural axon guidance 1,2. At the same time while some principles were established others were revisited. The non-neuronal cells of the nervous system, called glia (from the Greek γλία/ γλοία for glue), which as their name implies were long though to provide a passive substrate for neuronal growth, were realized to play active roles in neuronal physiology and function. Glia were first acknowledged to be neuron’s best friend, housekeeper, insulator, nurse 3,4and the list of glial roles expanded to include roles of glia as neural progenitors, tracts for migratory neurons and synaptic plasticity facilitators. Despite this progress, today’s classical neuroscience textbooks count only a handful of mentions of glia and their extensive roles. When I was an undergraduate, one would be lucky to attend a seminar on glial biology by one of the few scientists of the field.
I discovered the extensive glia literature when planning my post-doctoral research path. Focusing on the unknowns of neural development, I realized the enormous potential of the view that glia can actively regulate neuronal development and physiology. I was a descendant of the C. elegans research community, which I joined because of my first excitement about the C. elegans short life cycle and its recovery after freezing (allowing easy storage of strains for decades), but mainly for the ease of genetics, molecular biology and functional studies, possible at single-cell resolution. Yet, glial studies in this animal were few and far between. C. elegans axons are not myelinated by glia and glia were not shown to give rise to neurons nor to appose the neuromuscular junctions I studied during my PhD. In fact the presence of glia in C. elegans was not even clearly accepted. Then, midway during my PhD, Shai Shaham, my post-doc supervisor to-be, demonstrated that the so-called “support cells” of my favorite nematode are, in fact, glia 5, and soon after his lab demonstrated glial roles in neuronal ensheathment, dendrite shape and axon extension 6–8. Exciting! So many glial functions to be uncovered in C. elegans glia!
Yet, why would one study glial functions in C. elegans, where glial cells are limited to the 1/6 of the total neuron number, axons are non-myelinated and and microglia-like immune-specialized cells do not exist? For one, C. elegans has proven so useful for understanding the basic biology common to so many living things, that there was no reason to believe glia would be an exception! From a technical standpoint, most C. elegans neurons are born and grow independently of glia division and trophic support, thus perturbing glia does not cause neuronal death. In other systems, glial roles in neuron survival and support hamper attempts to address the active roles of glia in vivo in the nervous system. Finally, unlike most other settings, C. elegans glia can be studied in vivo, at single-cell resolution, using facile genetics, an invariant cell lineage and embryonic morphogenesis progresses in a transparent egg laid free in the environment. Moreover, live imaging of the same exact cell over and over again is possible in C. elegans. Who wouldn’t want to study glia in this setting!
The most complex circuit of the most well studied worm, and the forgotten cells
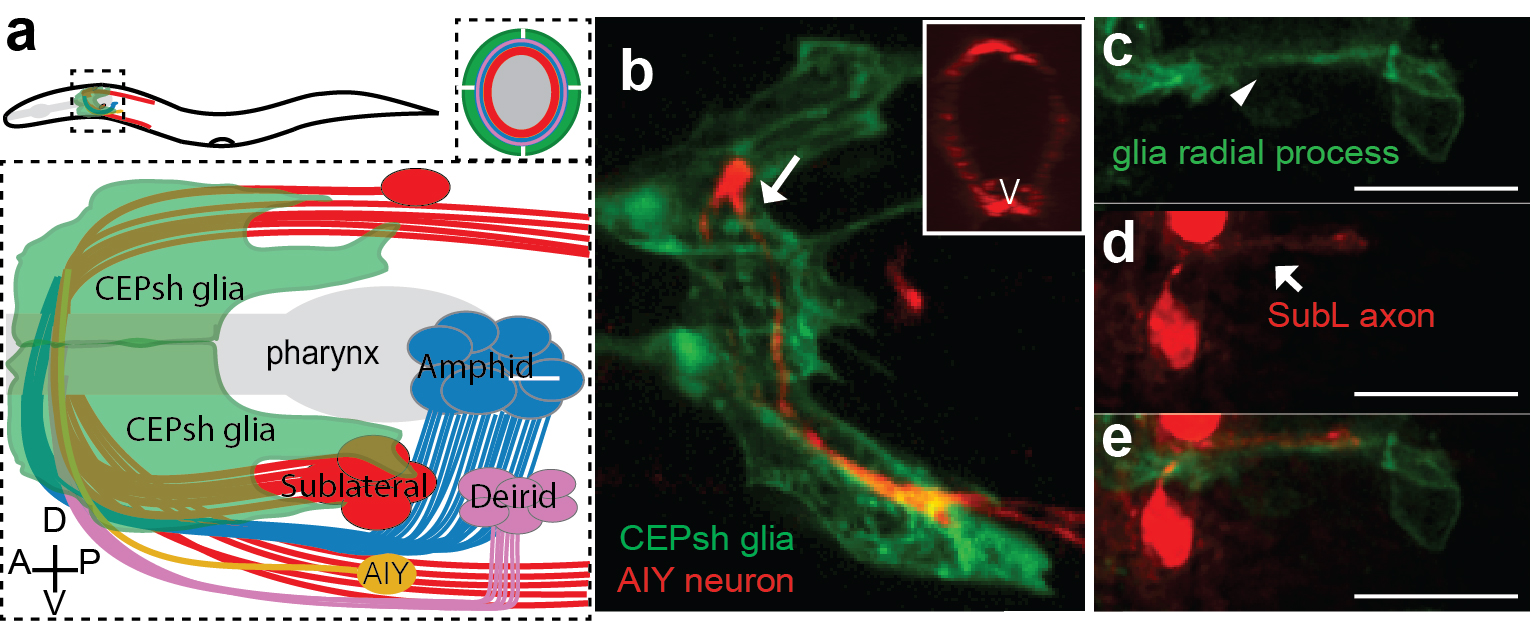
Armed with these thoughts, I joined Shai’s lab, decided to study developmental functions of glia, focusing on glial roles in the formation of the C. elegans brain-like neuropil, the nerve ring. The nerve ring (NR) consists of ~180 axons, presents the majority of synapses and interneuron connections in the animal, and is enveloped in its final form by four CEPsh glia, with astrocyte-like terminal morphology and molecular content (Figure 1; Katz & Shaham, personal communication,9). CEPsh glia express Netrin in early embryogenesis 10. Moreover, ablation of their precursors results in more profound NR defects than those of Netrin null mutants, suggesting unknown glia roles in NR assembly 7,11. Thus, I was interested to focus on glial roles in NR assembly, using biased approaches.
A few weeks into the project, writing for fellowship applications, I was struck by an important realization. While C. elegans was an important model for axon guidance cue discovery 10,12,13 and for studies of axon growth and fasciculation 12–16, how its most complex neuropil is formed during embryogenesis was subject to extensive speculation but was not yet described in detail. Only a handful of studies focused on NR structure, axon guidance or positioning 7,11,13,17, and many important questions remained unanswered. When and where does NR assembly start in the embryo? How and when do NR components enter the structure? What are the first axons to pioneer the NR and do they have functional roles in guiding later “follower axons”? Do the CEPsh glia extend early processes to define the structure or do they only enclose the structure in later embryogenesis, after axons have fasciculated, to protect them from the mechanical forces during embryonic elongation? Is NR integrity regulated only by Netrin-expressing glia (ventral CEPsh glia) or also by the non-expressing ones (dorsal CEPsh glia)? Which molecules, besides Netrin, may function from glia or from early neurons to address NR formation? Are there molecules that control specifically axon guidance, independently from neuronal migration and axon-growth initiation? I was surprised that so many questions remained open in C. elegans, a model system well appreciated for embryonic studies, tractable neuronal identities and single-cell resolution. Were the answers to these questions trivial, and not worth exploring? Or, as I have heard some neuroscientists say, is neural development mostly explained, and only the dynamic plasticity of neuronal circuits remains to be understood? Was I doomed to engage in a problem solved in other organisms, to provide the C. elegans version “merely” for the record?
I had to really understand the state of affairs in the field, and in reading the literature realized more and more how little was known. Not only in C. elegans, but at all. Indeed, my attempts to prepare introductory slides for my early scientific presentations did not prove easy. Transcriptional and morphogen signaling pathways for neuronal specification were defined in detail 18–20 and growth cone morphology distinguished pioneer and follower axons 21–23. However, how neuronal fates dictated circuit assembly initiation through a series of comprehensible axon-guidance events was, with few exceptions, not at all clear. The molecular identities of pioneer axons and the molecular mechanisms driving their functional roles remained understudied. Glia were known to form transient structures associated with axonal bundles and to express guidance cues 24. However, their functional roles in axon guidance independently of neuronal survival, growth, migration and their molecular interactions with pioneer axons appeared unresolved 25–27. I was reminded of a century-old Cajal’s quote: “What functional significance can be attributed to the neuroglia? Unfortunately, the present state of science does not allow to answer this important question but through more or less rational conjectures.” (Ramón y Cajal, 1899).
First things first: CEPsh glia and SubL pioneers initiate the nerve ring neuropil
I decided to start from scratch. Instead of biasing my studies to focus on glia, I decided to observe and image NR development, and let nature be my guide. The post-embryonic NR contains two major commissures, the amphid and deirid commissures composed primarily of sensory neuron axons 17. The posterior part of the amphid commissure is also referred to as the sublateral (SubL) commissure, since its axons extend out of the NR to populate the ventral sublateral cords. Since the amphid commissure is the largest characterized bundle populating the NR we hypothesized that this may be the one growing out first. I started imaging amphid-neuron axons in the beginning of morphogenesis, initially by using recently published reporters 28. Imaging bundled membranes using the by Pdyf-7::membrane-GFP reporter, or single axons, by photoconversion of specific neurons in animals expressing Pdyf-7::kaede, revealed axon growth only after 400 min of embryogenesis (known as the comma stage). Yet, electron microscopy (EM) images acquired by Yun Lu in the lab, showed that the pharyngeal primordium was surrounded by a thick layer of axons, corresponding to the early NR, already at 440min of embryogenesis (1.5-fold embryonic stage). So many axons were present already at 440min, making us wonder whether amphid neurons were truly early NR components.
I found myself, again, going one step back, realizing that I needed a more comprehensive effort to visualize as many embryonic neurons as I could. Since C. elegans has been a valuable model system for studies of neuronal specification and diversity 29, I was not too worried about finding neuron-subtype specific markers. I started to evaluate known neuronal markers. This screen left me with a handful of useful markers, and a couple of important take-home lessons. First, markers with post-embryonic expression restricted to specific neurons are usually more broadly expressed during embryonic morphogenesis. The C. elegans research community would, therefore, benefit greatly from characterizing expression patterns also during embryonic morphogenesis. Indeed, several labs today focus on expression profiling of C. elegans embryos 30,31, a blessed endeavor. This realization, that sparse, cell-specific embryonic reporters were few and far between, also drove the development in Shai’s lab of a method for labeling individual C. elegans embryonic cells without cell-specific drivers32. Second, the willingness of the C. elegans research community to share reagents, published dozens of years ago or others unpublished, to help me with my project, proved the supportive nature of this community, something that became rare in biological research.
I was now armed with reporters for many axons populating distinct NR commissures. By performing in vivo live embryonic timelapses of process growth into the NR, I saw that axons of the sublateral bundle grow early to define the NR neuropil while other commissures, amphid and deirid, were established in an orderly fashion later, with sequential axon growth occurring even within commissures (Figure 2a-b, 9). After months of sampling and imaging live embryos, I could not sustain my excitement when we also identified two bundles of similar position and composition in EM images of comma-stage embryos, taken by Yun Lu in the lab. I kept going back to analyze those EM images; yes, there were these two bundles, and no other multiprocess-bundles were obvious at that time, navigating to the NR presumptive position!
But what about the CEPsh glia? I had begun attempts to image CEPsh glia membrane growth the minute I joined Shai’s lab, but at the time no useful information could be gleaned, since no CEPsh-specific embryonic promoters were known. I decided to dissect regulatory elements of a couple of known CEPsh-expressing promoters, but this approach was not fruitful, as deletion analysis of cis-regulatory elements resulted in derepressed expression in non-glial cells. I finally decided to use a transgenic pan-glial marker, expressed from the onset of embryonic morphogenesis, and to follow stochastic segregation of the marker in my cells with mosaic analysis. Despite the method’s low throughput nature, I was able to visualize CEPsh membranes growing in the NR early and coalescing with growing SubL axons. Those early CEPsh membranes did not present the elaborate astrocyte-like endfeet seen post-embryonically but grow thin, non-branched processes reminiscent of vertebrate radial glia.
SubL pioneers and CEPsh populated the NR early! But who actually took the lead? Were SubL and CEPsh equally important in NR initiation or does one of the cell types guide the other? The answer came from ablation experiments I performed several months later. When specific CEPsh glia are ablated by expressing an apoptotic gene and following cell killing by mosaic analysis, pioneer axons grow ectopically, while follower axons can be short or mis-guided. Cell ablation of the SubL bundle perturbed follower axon pathfinding but CEPsh membrane morphology appeared normal. Thus, the most exciting scenario I could have imagined emerged: glia are not just bystander support cells maintaining the neuropil; they are necessary for initiating NR assembly. Moreover, our later studied revealed that CEPsh also cooperate with SubL pioneers to guide later NR components (Figure 2). This was an important realization. Although glia in other systems have been reported to appose axon bundles, functional analysis of glial roles in pioneer axon guidance was lacking, or was tested in settings where defects in neuronal viability or migration complicated the analysis. 26,27,33–35.
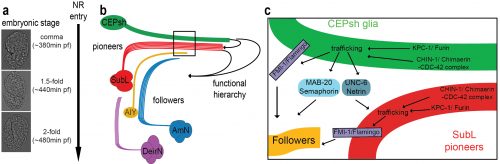
The NR is populated by neuronal commissures and CEPsh glia. Schematic (a) and imaging (b) of NR in postembryonic larval animals in lateral view. Inset: cross-sectional view. Pttx-3::mCherry, AIY and Phlh-17::membrane-GFP label AIY interneuron and CEPsh glia respectively. (c-e) In ~ 420min embryos, SubL pioneer axon bundle (SMDD neuron in red, Pttx-3::mCherry) and CEPsh glia (in green, Pmir-228::membrane-GFP) coalesce to define the NR. Figure adapted from Rapti et al, 2017 (9).
An “unsuccessful screen” & the first viable mutant with severe NR assembly defects
Defining a working model of cellular interactions for NR initiation was amazing, but not enough! Long devoted to unbiased genetics, like many C. elegans researchers, I decided to search for genes regulating NR formation through forward genetic screens. I screened tens of thousands of mutagenized worms under a compound microscope, recovering from the slides, back to bacteria-seeded plates, worms with abnormal axon trajectories. I started the screen well before my embryonic imaging uncovered the identities of early NR components. Back then I was thinking about neuropil formation, and selective fasciculation of axons. I hypothesized that fasciculation molecules, specific to neuron-subtypes, would ensure stereotypical interactions between axonal partners. I, therefore, designed screens for mutants in which two normally fasciculated axons, are properly guided but fail to adhere. My screens gave no viable mutants harboring the defects I was expecting. The reason for this remains unknown, but may suggest a different scenario of axon partnership based on the relative cell positioning of specific neurons at the time of their axon growth.
Regardless, in these screens I did find mutants with other NR axon defects. By now, I knew of the requirement of glia for NR assembly and decided to focus on mutants I found whose NR axon defects resembled those of glia-ablated animals. After cloning several previously-identified guidance genes with mild NR guidance defects, I decided to focus on a mutant with very severe NR defects, in which no mutations in the classical guidance genes were found by whole genome sequencing (WGS). And here started the painstaking tale of identifying the relevant gene. During my PhD, I mapped the first mutant of an uncharacterized gene without resorting to whole genome sequencing (WGS). It was difficult for me to understand why cloning my current mutant was so challenging, even with WGS data. The answer came when I proved in multiple ways that not one, but two mutations in two different unlinked genes were causal for the defects in the mutant. These mutations were synergistic, such that either single mutation caused only mild defects (5-15%) compared to the severe NR defect of the double mutant (>75%). To add to the complexity, one of the mutations was semi-dominant and subject to partial maternal rescue. After months of effort, I could finally announce that mutations in the Chimaerin gene chin-1, and the Furin gene kpc-1 were the cause of the mutant defects in NR axon guidance. When, in a joint group meeting, Shai, Leslie Vosshall, and Cori Bargmann commented on the low probability of recovering and identifying such a mutant, I felt that my hard work had really paid off. I was also filled with optimism that today’s ease of WGS, facilitating such complicated endeavors, will hopefully help identification of mutants that in previous decades would have been frozen and forgotten. This should allow researchers to choose their mutants of interest mainly based on interesting phenotypes and not how easy their identification is. For one, our mutant was definitely of interest; overall it showed ~80% navigation defects in any NR axon we imaged by optical means and a severe defective overall NR structure by electron microscopy of newly-hatched animals.
A wrong educated guess and the non-canonical functions of “old players”
The identities of the genes we found and previous studies in C. elegans or vertebrate nervous systems prompted me to favor cell-autonomous functions of these genes 36–38. I devoted several months to unsuccessfully prove such cell-autonomous roles of those in axon guidance. I was finally forced to consider the possibility of non-cell-autonomous roles. Rescue studies using combinations of cell-specific embryonic promoters finally revealed that the C. elegans Chimaerin and Furin act from glia to guide pioneer SubL axons and from both glia and pioneer SubL to guide follower axons. And they do so at the onset of NR assembly, by specifically affecting NR axon guidance, and not neuronal or glia survival, fate specification, migration or differentiation. To our knowledge, this is the first mutant affecting NR formation in such a specific way.
This mutant gave us an inroad in studying what turned out to be highly redundant signaling pathways for neuropil assembly initiation, a problem that has plagued genetic analysis of the process not only in C. elegans, but in vertebrates as well. Using the synergistic nature of Chimaerin and Furin mutants, we screened for mutations enhancing either single mutant and identified a network of conserved axon guidance cues redundantly regulating NR assembly. Importantly using our system of hierarchical NR assembly we could precisely define the cell-specific contributions of those factors. It turned out that knowing the site of action of these cues was crucial for modeling their actions. We demonstrated that C. elegans glia use distinct Netrin and Semaphorin signaling pathways to guide pioneer and follower axons respectively. We further showed glia and pioneer neurons together use the C. elegans Celsr/Flamingo homolog, FMI-1, to ensure proper navigation of follower axons into the neuropil. Moreover, our results support roles for Chimaerin and Furin in trafficking guidance cues, a novel combined function for this pair of proteins (Figure 2).
Model of cellular and molecular events of embryonic NR assembly (a)The NR is assembled, orderly, from the comma till after the 2-fold embryonic stage. (b) CEPsh glia and pioneer SubL axons enter the path first, later followed by “follower” axons of commissures or non-commissural paths. This follows a functional hierarchy of CEPsh glia guiding SubL pioneers and cooperating with them to guide follower axons. (c) Molecular pathways function redundantly from CEPsh glia and SubL pioneers to drive NR assembly. Figure adapted from Rapti et al, 2017 (9).
In summary, our studies uncovered the initiating events of NR formation in C. elegans, identified key pioneer roles for glia, defined the identities of the first pioneer neurons that enter this brain-like neuropil, and uncovered a molecular framework governing axon guidance events early on. Our studies uncovered a genetic strategy for dissecting the highly redundant cellular and molecular interactions driving axon guidance, and defined a new role for two old players in guidance factor trafficking. This sounds like a lot of work. And it was. But really, it is just the beginning. There are so many questions left unanswered, and the logic of assembly is still not understood for most of the neurons entering the NR. How relevant these studies are to other systems also remains to be explored, but some exciting similarities have recently emerged (see below).
New ways of thinking about old molecules and established concepts
In retrospect, our findings did not provide a short, easy-to-digest, easy-to-sell story. Instead, by tackling a multifaceted problem without shying away from the complexity we took to heart Einstein’s quip that phenomena must be explained in the simplest possible terms, but no simpler. This is an important lesson- while there is great temptation to understand biology in well-packaged sound bites, life turns out usually to be a lot more complex. Oversimplifying can have the unwanted effect of preventing fundamental understanding of a process, and can lead to proclamations of entire scientific fields being “solved”, even when this is not the case. In our case, the benefit of revisiting old problems with unbiased approaches was the discovery of new concepts, like the new roles of previously known molecules and importantly the glia roles in initiating assembly.
Remarkably, as our manuscript was nearing publication, two papers were published that questioned the precise roles of Netrin in commissural axon guidance in vertebrates 39,40. While questioning the long-standing role of the floor plate in commissural axon guidance, they revealed important roles for Netrin derived from ventricular-zone neural progenitors that correspond to radial glia. These studies help put our discoveries in a much broader context. In our paper, we suggest that embryonic CEPsh glia resemble radial glia, both morphologically and molecularly, suggesting that vertebrate radial glia may guide pioneer axons to initiate circuit assembly, in addition to their recognized roles in neuronal migration. The recent Netrin papers greatly strengthen our hypothesis, and predict that the proteins and gene interactions we described in our paper are likely to exist in vertebrates as well.
This project taught me a valuable lesson: keep an open mind; you may be surprised.
 (2 votes)
(2 votes)
 (1 votes)
(1 votes)
 Christiana Ruhrberg studied Biology at the Justus Liebig University (Giessen, Germany), and obtained her first class Diploma/MSc degree in 1992. After taking on an MSc/research assistant position at the University of Sussex for two years to elucidate genetic changes in ovarian cancer, she moved to Imperial College London to work for another two years to study the genomic organisation of the gene-rich human ‘surfeit’ locus. She then carried out her PhD project in the laboratory of Fiona Watt at the Imperial Cancer Research Fund (1994-97) where she identified and described the function of the envoplakin and periplakin genes. Her postdoctoral research in the laboratories of Robb Krumlauf at the National Institute for Medical Research in London (1997-1999) was funded by a MRC postdoctoral training fellowship and dedicated to studying the role of Hoxa1, Hoxb1 and Hoxb2 during hindbrain development. During her second postdoc with David Shima at the Imperial Cancer Research Fund in London (2000-2002), she was funded by a ICRF fellowship and worked on VEGF-A-mediated blood vessel branching. Having received an MRC Career Development Award in 2003, she became an independent investigator at University College London’s Institute of Ophthalmology studying links between vascular and neuronal development, with particular focus on the roles of VEGF and SEMA3A signalling during facial nerve and blood vessel formation. Staying at that same institute, she was appointed Lecturer in 2007, promoted to Reader in 2008 and then full Professor in 2011. Christiana has an excellent record in obtaining research funding, as illustrated by a Wellcome Trust Junior Investigator Award in 2011 and a Wellcome Trust Investigator Award in 2016 for her research on vascular biology and neurovascular interactions.
Christiana Ruhrberg studied Biology at the Justus Liebig University (Giessen, Germany), and obtained her first class Diploma/MSc degree in 1992. After taking on an MSc/research assistant position at the University of Sussex for two years to elucidate genetic changes in ovarian cancer, she moved to Imperial College London to work for another two years to study the genomic organisation of the gene-rich human ‘surfeit’ locus. She then carried out her PhD project in the laboratory of Fiona Watt at the Imperial Cancer Research Fund (1994-97) where she identified and described the function of the envoplakin and periplakin genes. Her postdoctoral research in the laboratories of Robb Krumlauf at the National Institute for Medical Research in London (1997-1999) was funded by a MRC postdoctoral training fellowship and dedicated to studying the role of Hoxa1, Hoxb1 and Hoxb2 during hindbrain development. During her second postdoc with David Shima at the Imperial Cancer Research Fund in London (2000-2002), she was funded by a ICRF fellowship and worked on VEGF-A-mediated blood vessel branching. Having received an MRC Career Development Award in 2003, she became an independent investigator at University College London’s Institute of Ophthalmology studying links between vascular and neuronal development, with particular focus on the roles of VEGF and SEMA3A signalling during facial nerve and blood vessel formation. Staying at that same institute, she was appointed Lecturer in 2007, promoted to Reader in 2008 and then full Professor in 2011. Christiana has an excellent record in obtaining research funding, as illustrated by a Wellcome Trust Junior Investigator Award in 2011 and a Wellcome Trust Investigator Award in 2016 for her research on vascular biology and neurovascular interactions.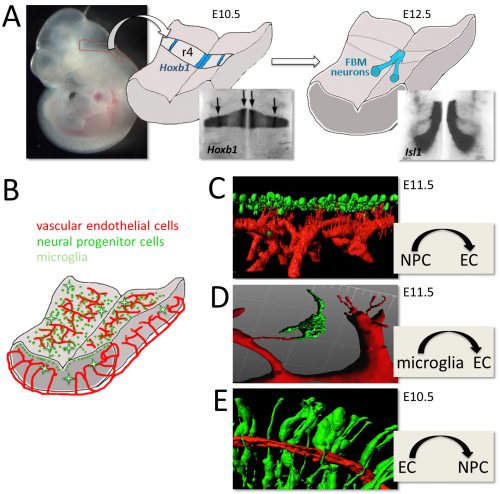
 Once upon a time a genetic screen identified a signalling pathway that caused Drosophila melanogaster embryos to develop a ‘lawn’ of denticles rather than forming them only at parasegment boundaries. Thus the so-called Hedgehog signaling pathway was born (
Once upon a time a genetic screen identified a signalling pathway that caused Drosophila melanogaster embryos to develop a ‘lawn’ of denticles rather than forming them only at parasegment boundaries. Thus the so-called Hedgehog signaling pathway was born (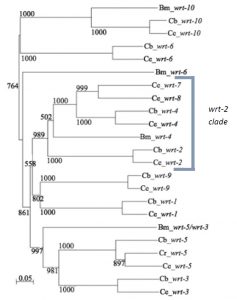

 (5 votes)
(5 votes)