The research group of Dr. Elvira Mass is looking to recruit a highly motivated PhD student to join her group Developmental Biology of the Innate Immune System.
Our research focuses on tissue-resident macrophages and their homeostatic functions. Specifically, we are interested how macrophages impact organ development and function during embryogenesis and postnatal stages on the cellular and molecular level. Based on new developmental and mechanistic knowledge, we seek to improve our understanding of disease pathophysiology and treatment.
We are part of the Life & Medical Science (LIMES) Institute, which is supported by DFG and Cluster of Excellence funding and which offers a great scientific environment for young researchers.
Requirements:
Previous experience in cell biology/immunology/hematopoiesis/animal models
The willingness to work with mice is a prerequisite
basic immunological and histological techniques such as flow cytometry and immunofluorescent stainings are beneficial
Programming skills (R/Python) for transcriptome analysis are advantageous
Your application should include:
CV
brief summary of your previous work experience (max. 1/2 page)
contact information of referees and dates when you wish to start the position
optional: 1 page research proposal for your PhD project, which builds on the study Mass et al. Science 2016
We are looking for someone working on Models of Animal Regeneration to join the CRTD Dresden as soon as possible!
Your permanent position will come with a generous package, full facility support, and the lively environment of a growing, interdisciplinary campus with tight links to the medical clinics.
Our latest monthly trawl for developmental biology (and other cool) preprints. Let us know if we missed anything.
October was a monster month for preprinting (a burst of post-summer productivity?), notable for the number of preprints covering plant development, disease modelling, modelling modelling, neurodevelopment, and organisms ranging from polychaete worms to hemipterans, Arctic charr to roses (plus all the usual suspects).
The preprints were hosted on bioRxiv, PeerJ, andarXiv. Use these links to get to the section you want:
Transcriptional regulation by NR5A2 couples cell differentiation and inflammation in the pancreas. Isidoro Cobo, Paola Martinelli, Marta Flandez, Latifa Bakiri, Mingfeng Zhang, Enrique Carrillo-de-Santa-Pau, Jinping Jia, Liv Thommesen, Torunn Bruland, Natalia del Pozo, Sara Olson, Jill Smith, William R. Bamlet, Gloria M. Petersen, Nuria Malats, Laufey Amundadottir, Erwin Wagner, Francisco X. Real
The Transcriptional Logic of Mammalian Neuronal Diversity. Ken Sugino, Erin Clark, Anton Schulmann, Yasuyuki Shima, Lihua Wang, David L. Hunt, Bryan M. Hooks, Dimitri Trankner, Jayaram Chandrashekar, Serge Picard, Andrew Lemire, Nelson Spruston, Adam Hantman, Sacha B. Nelson
RNA velocity in single cells. Gioele La Manno, Ruslan Soldatov, Hannah Hochgerner, Amit Zeisel, Viktor Petukhov, Maria Kastriti, Peter Lonnerberg, Alessandro Furlan, Jean Fan, Zehua Liu, David van Bruggen, Jimin Guo, Erik Sundstrom, Goncalo Castelo-Branco, Igor Adameyko, Sten Linnarsson, Peter Kharchenko
Autosomal recessive coding variants explain only a small proportion of undiagnosed developmental disorders in the British Isles.Hilary C. Martin, Wendy D. Jones, James Stephenson, Juliet Handsaker, Giuseppe Gallone, Jeremy F. McRae, Elena Prigmore, Patrick Short, Mari Niemi, Joanna Kaplanis, Elizabeth Radford, Nadia Akawi, Meena Balasubramanian, John Dean, Rachel Horton, Alice Hulbert, Diana S. Johnson, Katie Johnson, Dhavendra Kumar, Sally Ann Lynch, Sarju G. Mehta, Jenny Morton, Michael J. Parker, Miranda Splitt, Peter D. Turnpenny, Pradeep C. Vasudevan, Michael Wright, Caroline F. Wright, David R. FitzPatrick, Helen V. Firth, Matthew E. Hurles, Jeffrey C. Barrett
Identification, Isolation, and Characterization of Human LGR5-positive Colon Adenoma Cells. Michael K Dame, Durga Attili, Shannon D McClintock, Priya H Dedhia, Peter Ouilette, Olaf Hardt, Alana M Chin, Xiang Xue, Julie Laliberte, Erica L Katz, Gina M Newsome, David Hill, Alyssa Miller, Yu-Hwai Tsai, David Agorku, Christopher H Altheim, Andreas Bosio, Becky Simon, Linda C Samuelson, Jay A Stoerker, Henry D Appelman, James Varani, Max S Wicha, Dean E Brenner, Yatrik M Shah, Jason R Spence, Justin A Colacino
The Polycomb-dependent epigenome controls β-cell dysfunction, dedifferentiation and diabetes. Tess Tsai-Hsiu Lu, Steffen Heyne, Erez Dror, Eduard Casas, Laura Leonhardt, Thorina Boenke, Chih-Hsiang Yang, Sagar, Laura Arrigoni, Kevin Dalgaard, Raffaele Teperino, Lennart Enders, Madhan Selvaraj, Marius Ruf, Sunil Jayaramaiah Raja, Huafeng Xie, Ulrike Boenisch, Stuart H. Orkin, Francis Lynn, Brad G. Hoffman, Dominic Grün, Tanya Vavouri, Adelheid Lempradl, Andrew Pospisilik
WDR11-mediated Hedgehog signalling defects underlie a new ciliopathy related to Kallmann syndrome. Yeon-Joo Kim, Daniel Osborn, Ji-Young Lee, Masatake Araki, Kimi Araki, Timothy Mohun, Johanna E M Kansakoski, Nina Brandstack, Hyun-Taek Kim, Francesc Miralles, Cheol-Hee Kim, Nigel A Brown, Hyung-Goo Kim, Juan Pedro Martinez-Barbera, Paris Ataliotis, Taneli Raivio, Lawrence C Layman, Soo-Hyun Kim
Molecular evolutionary trends and feeding ecology diversification in the Hemiptera, anchored by the milkweed bug genome. Kristen A. Panfilio, Iris M. Vargas Jentzsch, Joshua B. Benoit, Deniz Erezyilmaz, Yuichiro Suzuki, Stefano Colella, Hugh M. Robertson, Monica F. Poelchau, Robert M. Waterhouse, Panagiotis Ioannidis, Matthew T. Weirauch, Daniel S.T. Hughes, Shwetha C. Murali, John H. Werren, Chris G.C. Jacobs, Elizabeth J. Duncan, David Armisén, Barbara M.I. Vreede, Patrice Baa-Puyoulet, Chloé S. Berger, Chun-che Chang, Hsu Chao, Mei-Ju M. Chen, Yen-Ta Chen, Christopher P. Childers, Ariel D. Chipman, Andrew G. Cridge, Antonin J.J. Crumière, Peter K. Dearden, Elise M. Didion, Huyen Dinh, harshavardhan doddapaneni, Amanda Dolan, Shannon Dugan-Perez, Cassandra G. Extavour, Gérard Febvay, Markus Friedrich, Neta Ginzburg, Yi Han, Peter Heger, Thorsten Horn, Yi-min Hsiao, Emily C. Jennings, J. Spencer Johnston, Tamsin E. Jones, Jeffery W. Jones, Abderrahman Khila, Stefan Koelzer, Viera Kovacova, Megan Leask, Sandra L. Lee, Chien-Yueh Lee, Mackenzie R. Lovegrove, Hsiao-Ling Lu, Yong Lu, Patricia J. Moore, Monica C. Munoz-Torres, Donna M. Muzny, Subba R. Palli, Nicolas Parisot, Leslie Pick, Megan Porter, Jiaxin Qu, Peter N. Refki, Rose Richter, Rolando Rivera-Pomar, Andrew J. Rosendale, Siegfried Roth, Lena Sachs, M. Emília Santos, Jan Seibert, Essia Sghaier, Jayendra N. Shukla, Richard J. Stancliffe, Olivia Tidswell, Lucila Traverso, Maurijn van der Zee, Séverine Viala, Kim C. Worley, Evgeny M. Zdobnov, Richard A. Gibbs, Stephen Richards
Mechanochemical feedback and control of endocytosis and membrane tension. Joseph Jose Thottacherry, Anita Joanna Kosmalska, Alberto Elosegui-Artola, Susav Pradhan, Sumit Sharma, Parvinder P. Singh, Marta C. Guadamillas, Natasha Chaudhary, Ram Vishwakarma, Xavier Trepat, Miguel A. del Pozo, Robert G. Parton, Pramod Pullarkat, Pere Roca-Cusachs, Satyajit Mayor
Summarizing Performance for Genome Scale Measurement of miRNA: Reference Samples and Metrics. P. Scott Pine, Steven P. Lund, Jerod R. Parsons, Lindsay K. Vang, Ashish A. Mahabal, Luca Cinquini, Sean C. Kelly, Heather Kincaid, Daniel J. Crichton, Avrum Spira, Gang Liu, Adam C. Gower, Harvey I. Pass, Chandra Goparaju, Steven M. Dubinett, Kostyantyn Krysan, Sanford A. Stass, Debra Kukuruga, Kendall Van Keuren-Jensen, Amanda Courtright-Lim, Karol L. Thompson, Barry A. Rosenzweig, Lynn Sorbara, Sudhir Srivastava, Marc L. Salit
ACEseq – allele specific copy number estimation from whole genome sequencing. Kortine Kleinheinz, Isabell Bludau, Daniel Huebschmann, Michael Heinold, Philip Kensche, Zuguang Gu, Cristina Lopez, Michael Hummel, Wolfram Klapper, Peter Moeller, Inga Vater, Rabea Wagener, ICGC MMML-Seq project, Benedikt Brors, Reiner Siebert, Roland Eils, Matthias Schlesner
HiGlass: Web-based Visual Exploration and Analysis of Genome Interaction Maps. Peter Kerpedjiev, Nezar Abdennur, Fritz Lekschas, Chuck McCallum, Kasper Dinkla, Hendrik Strobelt, Jacob M. Luber, Scott B. Ouellette, Alaleh Azhir, Nikhil Kumar, Jeewon Hwang, Soohyun Lee, Burak H. Alver, Hanspeter Pfister, Leonid A. Mirny, Peter J. Park, Nils Gehlenborg
Barriers to Integration of Bioinformatics into Undergraduate Life Sciences Education. Jason Williams, Jennifer Drew, Sebastian Galindo-Gonzalez, Srebrenka Robic, Elizabeth Dinsdale, William Morgan, Eric Triplett, James Burnette, Sam Donovan, Sarah Elgin, Edison Fowlks, Anya Goodman, Neal Grandgenett, Carlos Goller, Charles Hauser, John R. Jungck, Jeffrey Newman, William Pearson, Elizabeth Ryder, Melissa Wilson Sayres, Michael Sierk, Todd Smith, Rafael Tosado-Acevedo, William Tapprich, Tammy Tobin, Arlin Toro-Martínez, Lonnie Welch, Robin Wright, David Ebenbach, Mindy McWilliams, Anne Rosenwald, Mark Pauley
A Data Citation Roadmap for Scholarly Data Repositories. Martin Fenner, Mercè Crosas, Jeffrey Grethe, David Kennedy, Henning Hermjakob, Philippe Rocca-Serra, Gustavo Durand, Robin Berjon, Sebastian Karcher, Maryann Martone, Timothy Clark
The scent of the fly. Paul G. Becher, Sebastien Lebreton, Erika A. Wallin, Erik Hedenstrom, Felipe Borrero-Echeverry, Marie Bengtsson, Volker Jorger, Peter Witzgall
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our fifth report from the 2017 group of student awardees comes from Jake Cornwall Scoones(student at University of Cambridge), who undertook his studentship with Anna Philpottat the Dept. of Oncology in Cambridge.
Transdifferentiation of pancreatic organoids
This summer, I had the amazing opportunity of undertaking a Gurdon Summer Studentship, working in Prof. Anna Philpott’s lab, under the guidance of Dr. Roberta Azzarelli at the Cambridge Oncology Department. Having completed first-year Natural Sciences, I was intrigued to learn more about the molecular mechanisms that underpin development. Focused on a potential transdifferentiation pathway in pancreatic cells, the project has been both fascinating and informative, teaching me many invaluable skills, from tissue culture, to growing organoids, to having the patience required for many of the complex procedures involved.
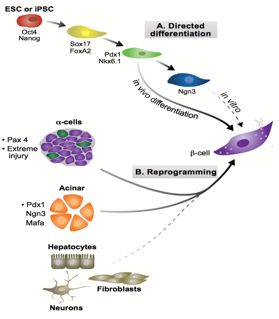
In recent years, many have championed regenerative medicine as a solution to diseases associated with cell loss like diabetes mellitus. This technique, where cells are grown and differentiated to replace dead or diseased cells, has the potential to overcome many of the pitfalls of transplantation, most notably immune rejection. To minimise rejection, inserted cells must display high genomic identity with the recipient. These cells can be derived by one of two methods:1, 2 through a differentiation programme, transitioning embryonic(-like)3 stem cells into the fate desired; or via transdifferentiation, through introducing factors that tweak the cell’s genome and epigenome.
Figure 1: Two possible pathways for generating β-cells. Adapted from Pagliuca and Melton, 2013
During development, cell-lineages sequentially accrue epigenetic modifications that consign them to their fate, transforming from a totipotent zygote to terminally differentiated cells. Waddington’s visual metaphor, envisaging this sequential differentiation as a ball rolling down a terrain of bifurcating valleys, helps to frame questions in developmental biology.4 Upon transplanting the nucleus of somatic Xenopus cells into enucleated eggs, Gurdon found that the resultant cell was pluripotent, capable of differentiating into all cell types.5 In so doing, Gurdon demonstrated that factors within an egg have the capacity to modify the epigenetic state of a nucleus, implying that the ball can roll back up the valley. In the case of pancreatic transdifferentiation, with sufficient knowledge of this terrain and with the correct molecular intervention, we should able to push the ball into our chosen valley.
Pancreatic development proceeds through two stages: the primary transition (embryonic day 9 (E9) to E12.5); and the secondary transition (E12 to birth).6 The primary transition sees endodermal thickening, followed by the pancreatic progenitor proliferation leading to pancreatic bud formation. Through a process of epithelial stratification and micro-lumen formation, the initial tubular morphology that will later characterise the pancreas begins to emerge. This initial expansion phase is followed by rounds of specification and patterning, forming a bipotent trunk and a multipotent tip. Adult pancreata are comprised of three different cell types, namely acinar, duct and endocrine cells, the former derived from tip progenitors and the latter two from trunk cells.
Endocrine development, part of the secondary transition, is triggered by the transient expression Neurogenin 3 (Ngn3) above a threshold level, and proceeds through delamination from the epithelium and aggregation into Islets of Langerhans.Ngn3+ cells are specified to one of five fates in a temporally regulated manner:7 α-cells produced upon earliest Ngn3-activation synthesising glucagon; followed by β-cells producing insulin; then δ-cells producing somatostatin; then PP-cells producing pancreatic polypeptide; and finally ε-cells producing ghrelin. Previous work from the lab by Azzarelli et al.8 demonstrated Ngn3 is phosphoregulated by proline-directed kinases (e.g. CDKs), regulating the balance between proliferation, (Ngn3 hyper-phosphorylation), and differentiation, (Ngn3 under-phosphorylation). β-cell maturation is promoted by the transcriptional regulator Pdx1, among others, whose expression remains confined to β-cells in the adult pancreas. Working in tandem with Pdx1, MafA transcriptionally regulates genes involved in insulin secretion and biosynthesis and it is thus considered a marker of mature β-cell identity.
Ngn3, crucial in assigning endocrine fate initially, and MafA and Pdx1, important in β-cell maturation, have been previously over-expressed in combination to induce transdifferentiation of acinar cells to β-fate in an in vivo model. These factors were used in our experiment to see if such transdifferentiation is possible in an in vitro model, specifically pancreatic organoids, a form of 3D cell culture grown in matrigel. Upon over-expression of these three factors, it is possible to test for changes in cell fate, by looking at symptomatic traits of β-cells, namely the production of insulin.
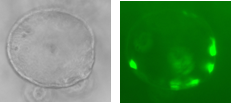
Over-expression can be achieved through the infection of organoids with lentiviral vectors carrying genetic loci that upon infection will be inserted into the cells’ nuclear genomes. The activation of these imported genes should be controllable so a transactivator is used which, when bound to the antibiotic doxycycline (Dox), becomes active, binding the promoter of our genes of interest, inducing their transcription. The three factors are necessarily expressed together through their transcription within a single ORF, containing MafA, Pdx, Ngn3 and GFP (a fluorescent marker) sequences, separated by loci encoding the cleavage peptide 2A, meaning upon translation, cleavage ensues resulting in four functional proteins. Hence a GFP signal is sufficient to determine the expression of all three factors, leaving other wavelengths free to test for pancreatic hormones through immunohistochemistry.
Figure 2: Pancreatic orgnaoids infected with a virus encoding GFP
Two lentiviral vectors were used to infect organoids, one carrying the transactivator gene Tet3G, and the other carrying the locus of interest, Pdx-MafA-Ngn3-GFP (PMN), or GFP as our control. Viruses were produced by the transfection of HEK cells with several plasmids, each containing genes encoding different viral proteins plus the locus of interest, followed by incubation, viral purification and titration. Organoids were incubated with viruses for a week with Dox and then fixed with PFA. Immunohistochemistry was performed, staining organoids for insulin. Preliminary results are very encouraging and I look forward to seeing how my contribution fits into the larger project as a whole.
This studentship has been an invaluable experience, allowing me to gain an understanding of real-world lab science, a practice venturing into the unknown, far from the rushed experiments for which answers were already known performed as a part of my course. I have developed lab-skills, most notably having the rare opportunity to work with organoids, a relatively novel technology. I urge other students to consider applying for this fantastic opportunity in future years.
MSc/PhD and postdoc positions available in the Zaidel-Bar Cellular and Tissue Morphogenesis Lab.
We study the regulation of the cytoskeleton from single proteins to the entire organism and system levels, using multiple approaches (including bioinformatics, genetics, biochemistry and live imaging) to understand how cells and tissues change shape, move, sense, and generate forces (for more info: celladhesionlab.com).
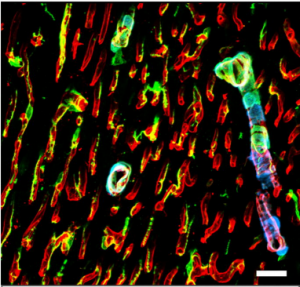
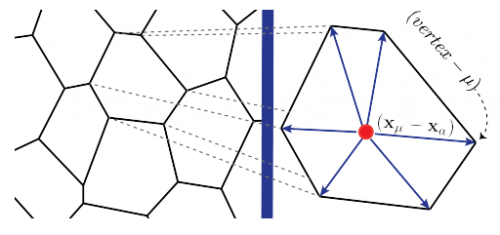
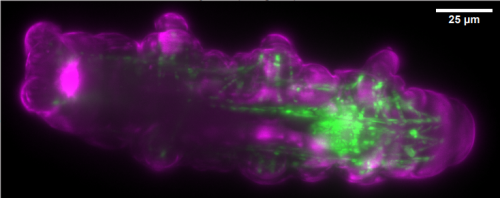
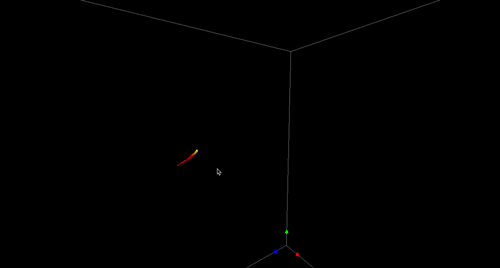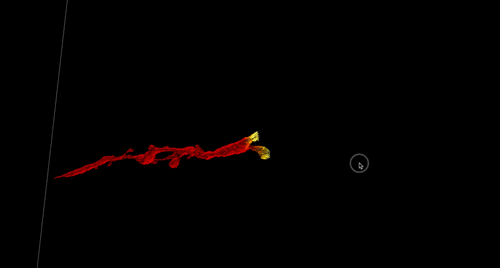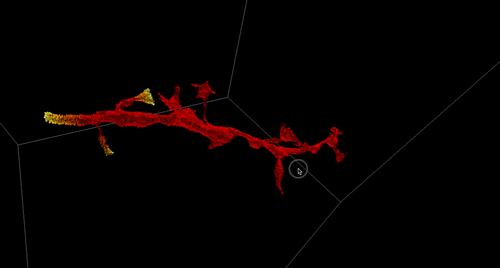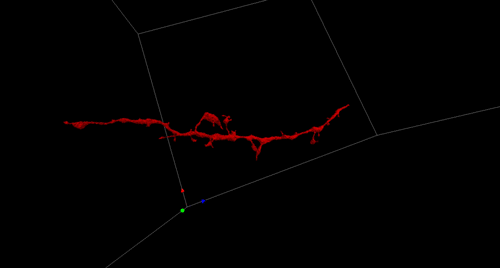
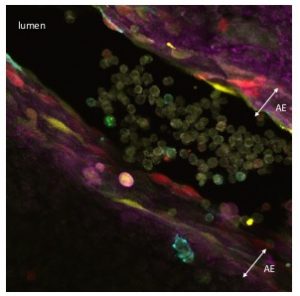
Figure 1. Aortic endothelium (AE) lit up by the Confetti cassette in an embryonic day (E) 11.5 VE-Cadherin (+/Cre) Confetti (+/T) mouse embryo.
My PhD focused in deciphering molecular mechanisms implicated in cell cycle regulation in embryonic and adult tissues. As many others in this field I became fascinated by the experiments by Yamanaka and colleagues and this prompted me to transition into the stem cell world. For this reason I joined Shannon McKinney-Freeman’s laboratory in 2012. Here, I became interested in understanding how Hematopoietic Stem Cells (HSCs) arise during embryonic development.
HSCs are mesodermal in origin and emerge from a subset of endothelial cells (known as hemogenic endothelium) mainly in the major arteries of the embryo. The hemogenic endothelium undergoes and endothelial-to-hematopoietic transition during midgestation. This process can be visualized as clusters of cells budding from the arterial endothelium into the lumen of these embryonic arteries.
In order to better understand how these clusters are formed I thought it would be interesting to analyze the expression profiles of cell cycle regulators within these cells in murine embryos. I was discussing some of these results with a brilliant former postdoc in the lab, Per Holmfeldt. He mentioned the existence of a genetic tool (the Confetti allele) that had been used to study the behaviour of other types of stem cells for example in the intestine (Snippert et al., 2010). I thought it was a great idea that could bring some light into the formation of these clusters.
The Confetti allele is a conditional multicolored reporter composed of four fluorescent proteins (GFP, CFP, YFP and RFP) flanked by different lox P sites in particular orientations. In the unrecombined orientation the cassette does not lead to the expression of any fluorescent protein. Upon Cre expression the Confetti allele recombines and the cells get randomly and stably label with one of the four colors. Importantly all the progeny of a particular cell will maintain the expression of the same color.
As I started working with this genetic tool (Figure 1, video 1), I realized that it could be used to answer more sophisticated questions. In particular, I thought it could be employed to understand long-standing issues like the clonal origin of the adult blood system at different embryonic stages.
In mammals, all previous studies have estimated that blood emerges from just a handful of blood progenitors during embryonic development. All these studies were based on the transplantation of embryonic tissues, that had to be previously dissected, dissociated, sometimes cultured ex vivo and finally transplanted. I thought all these stresses could have an impact on the final readout. Here a non-invasive approach should better reveal the actual clonal embryonic origin of the adult blood.
The challenge part here was that the Confetti allele only allows to label the blood with up to four colors. So it did not seem enough to study larger complexities. But here I had an idea.
Let’s say that we have a bag full of marbles of four different colors. These marbles can be green, yellow, blue or red. If we ask two persons to grab one marble. It is very likely that the marble that each of them have grabbed is different. In contrast, if each of these persons pick 100 marbles and then they compare the distribution of the colors they got it is more likely that it will be more similar. The reason is that the more marbles they get the distribution of the colors will be a better representation of what it is in the bag.
In this line, since the Confetti cassette labels the cells randomly with one of four colors (yellow, green, blue or red), this same mathematical concept could be applied for our purposes. The more blood precursors (marbles picked) then the distribution of the confetti colors among the blood of adult mice (persons grabbing marbles) will be more similar. If the number of blood precursors was small, then the distribution of the Confetti colors among different mice would be very different.
Here, you can probably imagine that the idea was now to use the mouse-to-mouse variance in the distribution of the Confetti colors in the adult blood as a predictor to estimate the number of blood precursors. Hence the required experiment was to plate known numbers of Confetti cells and then for each cell dose calculate the well-to-well variance in the distribution of the Confetti colors.
I have loved Mathematics all my life and this has been critical for me to develop this idea.
Still, I did not have the right expertise to derive a formula that could correlate these two variables. In order to crystallize this concept, we brought into the project a fantastic statistician, David Finkelstein, how was able to create a formula where we can use the mouse-to-mouse variance in the distribution of Confetti colors in the adult blood to infer the number of blood precursors.
After a number of experiments to validate this formula we were ready to apply this equation to our major question. We generated cohorts of mice where blood precursors were “Confetti-labeled” at particular developmental stages. Finally, we were able to estimate that lifelong hematopoiesis is established by hundreds of embryonic precursors at different stages during mouse development (Ganuza et al., 2017).
For me, this project has been extremely instructive further than from just the exciting data we found. I think it is a good example that illustrates how important is to interact with your colleagues in the lab to get new ideas that could lead to a completely new project. It definitively shows how important is to merge different scientific disciplines to address previous or new questions employing a fresh approach. I deeply believed that Mathematics need to be more heavily used in Biology.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our fourth report from the 2017 group of student awardees comes from Eleanor Sheekey(student at University of Cambridge), who undertook her studentship with Peter Rugg-Gunnat the Babraham Institute in Cambridge.
Much ado about Nanog
Named after the mythical “Land of the young”, Tír na nÓg, Nanog is a homeobox transcription factor expressed in embryonic stem cells (ESCs) aiding continual cellular proliferation alongside the other main pluripotency factors, Oct4 and Sox2. However, despite its contribution to pluripotency, much is still unknown of the mechanistic roles that Nanog plays within stem cells, since cells kept in culture deficient in Nanog retain the ability to self-renew (1). Interestingly, it was recently found that Nanog provides a link between pluripotency and chromatin organisation suggesting important functions beyond transcriptional regulation (3). With the BSDB Gurdon Studentship, I was privileged to spend my 8 weeks at the Babraham Institute alongside Dr Clara Novo to continue to explore this link.
Centromeres and chromatin
A key hallmark of stem cells is their ability to divide. However, this division must be executed precisely to avoid aneuploidy, an abnormal chromosomal number in a cell, and prevent unregulated division and cancer formation. Centromeres play a crucial role in chromosome separation, composing centric heterochromatin (CH) for kinetochore formation and pericentromeric heterochromatin (PCH) to hold chromatids together (2). Although centromeres are thought to be defined epigenetically, the centro- and pericentromere contain minor and major satellite DNA repeats, respectively. Nanog binds the major satellite DNA repeats within the pericentromere, maintaining the PCH in a ‘more’ open state with increased transcription of major satellites and lower levels of H3K9me3(3), bridging the gap between pluripotency and chromatin organisation. It also promotes the formation of chromocenters, which are clusters of major satellite repeats from several chromosomes. Deletion of Nanog leads to chromatin compaction including the PCH. Since PCH is essential for genetic stability and Nanog levels are known to fluctuate in stem cell cultures, the consequences of an altered PCH organisation will need to be understood before successfully developing more stably reprogrammed stem cells for future medical treatments (4).
Meet E14 and BT12
Throughout my project, I was working with mouse ESCs (mESCs). Much like in a vending machine, each of the different genetically modified mESCs are assigned a systematic name. E14 is like salted crisps, a male, wild type mESC cell line, originally isolated from the inner cell mass of a developing embryo. BT12, on the other hand is spiced up, no longer containing Nanog, but instead expressing a GFP transgene (1). Maintaining these two cell lines in culture was essential for conducting my experiments to study the effect of Nanog-/-.
FISH, IF and ChIPs
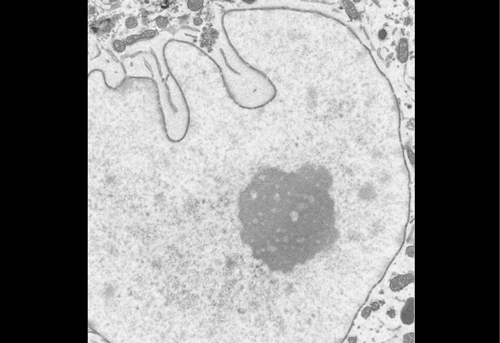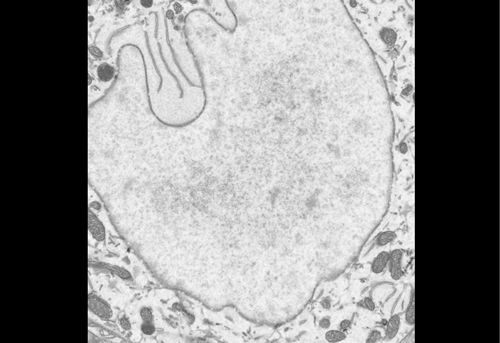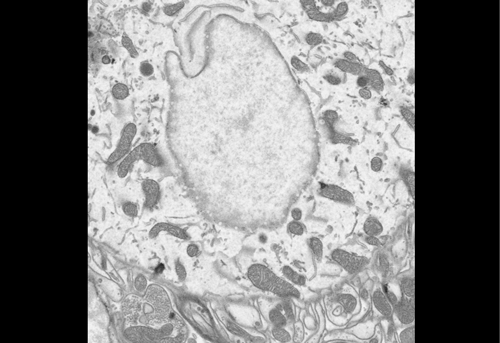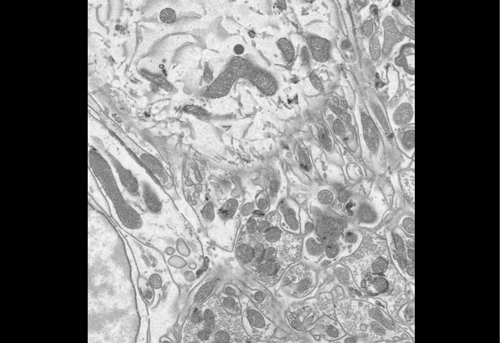
As Nanog was shown to affect heterochromatin at pericentromeres, we wondered if this effect extended to the centromere. To tease apart any differences in protein levels and localisation at the centromeres between E14 and BT12, we used a combination of techniques. Immunofluorescence (IF) provides a clear single cell insight and is beautiful to visualise under the microscope. Fluorophores had to be carefully chosen to avoid the wavelength clashing with that of GFP (already being expressed in BT12) and to differ from each other. Fluorescence in situ hybridisation (FISH) visualises specific locations of DNA using single-stranded probes and thus in combination with IF provides a good indication of whether proteins localise to a particular DNA region.
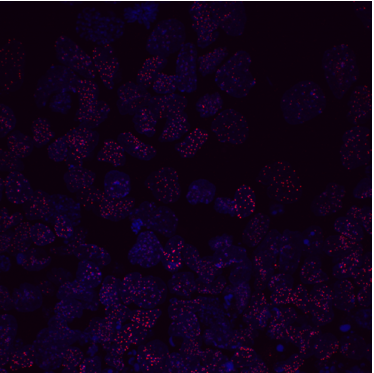
Figure 1 IF images of BT12
Figure 2 Using the 3D Imaris interphase to quantify my images
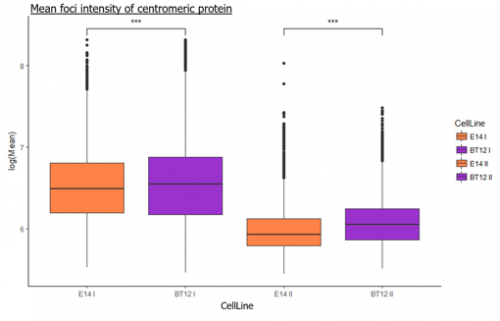
We were able to take some stunning images (Figure1) which could be used to quantify the levels of centromeric proteins assayed in wild-type and Nanog knockout cells. We then used the Imaris software (5) to quantify signals obtained from multiple 3D stacks, allowing for easy identification of fluorescent foci (Figure 2). Once the quantification of protein intensity was gathered I analysed it on R-studio to test for significant differences between both cell types (Figure 3).
Figure 3 R-studio analysis
To further this analysis, we used Chromatin immunoprecipitation (ChIP) that can answer many biological questions associated with DNA-protein interactions by fixing proteins to chromatin. In brief, by using antibodies specific to the protein of interest and magnetic beads that bind to these antibodies, a ‘pull-down’ of captured DNA can be isolated. The eluted DNA can then be analysed using ChIP-quantitative polymerase chain reaction (ChIP-qPCR) to quantify protein intensity at specific DNA regions. In our case, by using primers complementary to the major and minor satellites, we could assess if the binding of our proteins of interest differed between the cell lines.
Epilogue
Much like predicting the murderer in an Agatha Christie novel, one of my favourite aspects of the project was piecing together the experimental evidence to suggest hypotheses to explain our results. It is quite feasible for the absence of Nanog to have a diverse range of effects on the mESCs due to its interactions with DNA and other pluripotency factors, which could result in mislocalisation and/or altered post-translational modifications of its partners. Nevertheless, my preliminary results will require further validation and follow up before any conclusions can be made. However, in the manner of Shakespeare’s comedy, it has definitely been a fun experience, one from which I have gained much more lab confidence and exposure from, of which I am grateful both to the Rugg-Gunn lab at the Babraham Institute and the BSDB studentship that helped fund it.
A postdoctoral position is available in the laboratory of Dr. Sophie Astrof at Thomas Jefferson University to study roles of cell-extracellular matrix (ECM) interactions in cardiovascular development and congenital heart disease. We have recently discovered that progenitors within the second heart field (SHF) give rise to endothelial cells composing pharyngeal arch arteries. Projects in the lab focus on the role of ECM in regulating cell fate and migration of SHF-derived cells, and in the regulation of signaling between endoderm-, mesoderm-, and ectoderm-derived tissues during the morphogenesis of the pharyngeal vasculature. The project will combine genetic manipulation, embryology, cell biology, and confocal imaging. My laboratory is a part of a modern and well-equipped Center for Translational Medicine at Jefferson Medical College (http://www.jefferson.edu/university/research/researcher/researcher-faculty/astrof-laboratory.html) located in the heart of Philadelphia. For further information on our work, please see the following publications: Wang et al., Development, 143:88-100, 2016 and Wang et al, Developmental Biology 421:108-217, 2017. To apply, please send a letter of interest detailing your expertise, CV and names and contact information of three references to sophie.astrof@gmail.com
An NIH funded T32 postdoctoral fellow position is available in the Pediatric Heart Lung Center (PHLC), at the University of Colorado School of Medicine, in Aurora CO. The focus of the interdisciplinary programs within the PHLC is research with a special emphasis on new understanding and novel strategies for the treatment of childhood cardiovascular and pulmonary diseases. This position will report to Dr. Douglas Shepherd and Dr. Steven Abman. Animal models, spatial transcriptomics, optical tissue clearing, and a novel cleared-tissue specific light-sheet fluorescence microscope will all be utilized to develop new insights into the molecular mechanisms governing endothelial cell differentiation during fetal development (see: Ryan et al 2017; Singh et al., 2017; Jun Yun et al., 2016).
The ideal candidate has experience with animal models and advanced fluorescence imaging. Opportunities will exist for both instrument development and new biological models of vascular development in the lung. Therefore, candidates with a microscopy background and/or a biological background are invited to apply.
Additionally, Dr. Shepherd has existing projects as part of the University of Colorado Center for Neuroscience on imaging within intact murine brains. Through these projects, opportunities will also be available to apply spatial transcriptomics to the developing brain.
Because this an NIH T32 funded position, candidates must hold US citizenship or a US permanent residence.
Established by the British Society for Developmental Biology in 2014, The Gurdon/The Company of Biologists Summer Studentship scheme provides financial support to allow highly motivated undergraduate students an opportunity to engage in practical research during their summer vacation. Each year, ten successful applicants spend eight weeks in the research laboratories of their choices, and the feedback we receive is outstanding.
Our third report from the 2017 group of student awardees comes from Rachael Adams (student at University of Cambridge), who undertook her studentship withPeter Lawrenceat the Dept. of Zoology in Cambridge.
This summer, thanks to the Gurdon/The Company of Biologists Summer Studentship, I was fortunate to spend 8 weeks working in Dr Peter Lawrence’s lab in the Department of Zoology.
The group studies PCP, a pathway which coordinates cell polarity and helps to align epidermal patterns in the Drosophila abdomen. Drosophila larvae are covered with a cuticle decorated by denticles which help the larvae to grip substrate in order to move. These denticles form a specific pattern and changes in the pattern can be used to investigate the properties of the PCP pathway. PCP genes are highly conserved and have been identified as being involved in processes such as vertebrate gastrulation, demonstrating their fundamental importance in animal development and patterning.
My project aimed to investigate the importance of Rab GTPases to PCP. Much research has been carried out on the Rab family of proteins, as they act as master regulators of intracellular membrane trafficking. The accurate delivery of cargo between organelles is crucial for normal eukaryotic cell function. Rabs facilitate this by coordinating vesicle formation, transport and fusion.
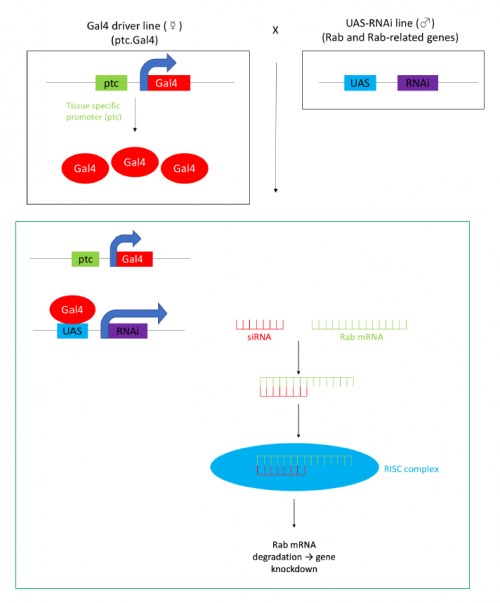
Figure 1- The w;ptc.Gal4 line of flies have the yeast Gal4 gene under the control of the patched(ptc) gene promoter, Gal4 binds to UAS sites to promote transcription. This means that when the driver line is crossed to the UAS-RNAi line the resultant progeny (in green box) express RNAi in the same location as the patched gene. This RNAi is complementary to a specific Rab mRNA, the resultant double stranded RNA is targeted for degradation by the RISC silencing complex, this results in gene knockdown.
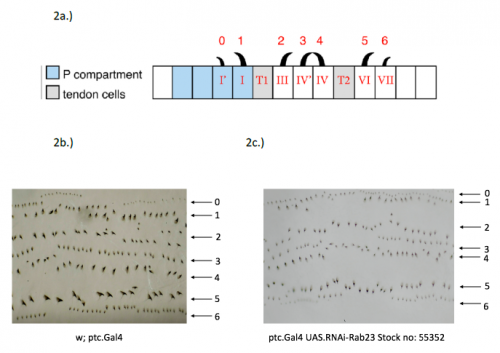
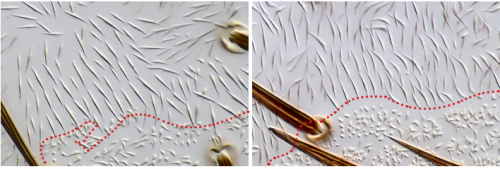
In order to test the role of as many Drosophila Rabs as possible I carried out a UAS RNAi knockdown screen of 34 different Rab and Rab-related genes, see figure 1. This screen took 7 weeks to carry out as I had to cross a collection of 103 different UAS-RNAi lines with ptc.Gal4 virgin females. This involved maintaining a population of ptc.Gal4 flies in several bottles. I crossed the virgins with males obtained from each UAS-RNAi stock and incubated the flies at 29℃, the optimal temperature to see the effect of the RNAi. Once each cross was made it was approximately 5 days before third instar larvae could be collected. To identify any change in the phenotype, I mounted several third instar larval progeny of each Gal4-UAS- RNAi cross and examined the denticle pattern produced under the microscope. Most larvae exhibited a wild type pattern, this may be because most of the Rabs I examined don’t have a function in PCP or redundancy of Rab proteins may mask certain phenotypes. The low efficacy of some of the RNAi constructs may have also affected the proportion of successful knockdowns. However, I found that several of the Rab23 crosses produced an unusual phenotype where the tendon cell gaps appeared larger than normal, see figure 2. In order to test whether this result is repeatable, I have crossed the UAS-RNAi Rab23 stocks to different driver lines (sr.Gal4 and en.Gal4). As Gal4 expression is under the control of a different promoter in each driver line, this results in a different expression pattern of the RNAi, see figure 3. Whilst different drivers will not produce the same phenotype, if the denticle pattern is disrupted it may help in the investigation of the role of Rab23 in PCP.
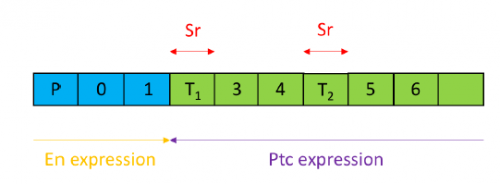
Figure 2- 2a.) A schematic of the wild type denticle pattern (Saavedra, Brittle, Palacios, Strutt, Casal and Lawrence. (2016)) 2b.) The patched driver line denticle pattern. As in the wild type, rows 0,1 and 4 face anteriorly whilst rows 2,3,5 and 6 face posteriorly. The tendon cell gaps are relatively small. 2c.) The Rab 23 knockdown denticle pattern, the tendon cell gaps are enlarged.
Figure 3- An illustration of where patched (ptc), stripe (sr) and engrailed (en) are expressed relative to the position of denticles and tendon cells. En is expressed in the posterior compartment (illustrated in blue). Ptc is expressed in the anterior compartment (illustrated in green) outside of the domain of en where it acts as a receptor for Hedgehog (Hh). Sr expression is limited to the tendon cells.
The results from the ptc.Gal4- UAS-RNAi Rab 23 crosses raised the question of why the tendon cell gap appeared larger. Possibilities include: that the tendon cells were enlarged, divided more frequently or the surrounding cells were smaller. To investigate this phenotype, I set up a series of crosses and aim to ultimately generate larvae carrying the Rab23 knockdown and expressing spaghetti squash protein (sqh) labelled with the fluorescent marker mCherry. Sqh encodes the regulatory light chain of non-muscle myosin so is expressed throughout cells. The sqh.mCherry construct therefore allows individual cells to be visualised with a confocal microscope and could help identify why the tendon cell gap is expanded.
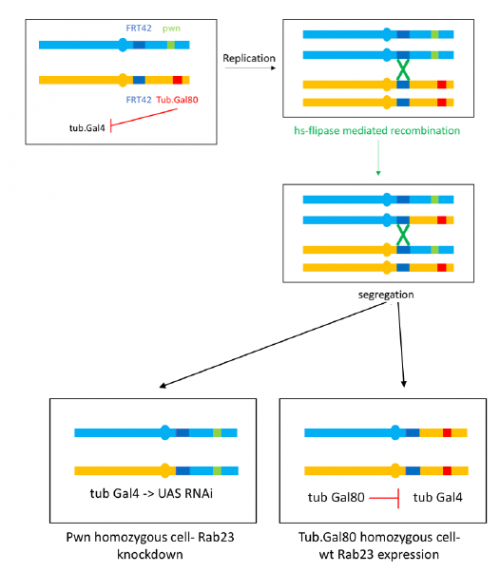
To further investigate the phenotype I set up crosses to produce marked clones in adult flies where Rab23 is knocked down, the resultant mosaic animal allows comparison with wild type cells. This is achieved by using the Flp-FRT system to bring about site-specific recombination, see figure 4.
Figure 4- I aim to create flies with the above genotype by a series of crosses. When flipase (Flp) expression is activated by heat shock, recombination between FRT42 sites on the second chromosome occurs. This produces a mixture of clones with the knockdown and pawn (pwn) truncated bristle marker and wild type clones where Gal80 inhibits Gal4 activity and Rab 23 is expressed. Figure inspired by Prudêncio and Guligur, 2015, FLP/FRT induction of mitotic recombination in Drosophila germline.
I have really enjoyed my time in the lab and have learned a great deal. I am looking forward to continuing to investigate the Rab23 phenotype and seeing the results of my crosses as part of my Part II Zoology project this term. My special thanks to José Casal for supervising and to everyone in the lab for providing a great deal of help and advice.
 (No Ratings Yet)
(No Ratings Yet)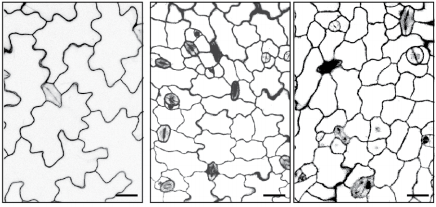









 (1 votes)
(1 votes)
 Named after the mythical “Land of the young”, Tír na nÓg, Nanog is a homeobox transcription factor expressed in embryonic stem cells (ESCs) aiding continual cellular proliferation alongside the other main pluripotency factors, Oct4 and Sox2. However, despite its contribution to pluripotency, much is still unknown of the mechanistic roles that Nanog plays within stem cells, since cells kept in culture deficient in Nanog retain the ability to self-renew (
Named after the mythical “Land of the young”, Tír na nÓg, Nanog is a homeobox transcription factor expressed in embryonic stem cells (ESCs) aiding continual cellular proliferation alongside the other main pluripotency factors, Oct4 and Sox2. However, despite its contribution to pluripotency, much is still unknown of the mechanistic roles that Nanog plays within stem cells, since cells kept in culture deficient in Nanog retain the ability to self-renew (